
Abstract
XPNPEP2 is a proline hydrolytic enzyme that hydrolyzes several biologically active peptides and causes a loss of substrate activity. However, its function in cancer is still unknown. Our study showed that XPNPEP2 expression was significantly upregulated in cervical cancer tissues compared with normal cervical tissues and cervical intraepithelial neoplasm tissues. Statistical analysis showed that XPNPEP2 expression was associated with the International Federation of Gynecology and Obstetrics stage and lymph node metastasis. Overexpression of XPNPEP2 in SiHa and HeLa cells promoted cell invasion and migration without affecting cell proliferation and apoptosis. Mechanistically, we found that XPNPEP2 facilitated cervical cancer cell invasion and migration by inducing epithelial–mesenchymal transition. Furthermore, we demonstrated that XPNPEP2 had significant effects on the metastasis of xenografted tumors in vivo. Collectively, our findings identify the novel function of XPNPEP2 in the metastasis of cervical cancer and suggest that XPNPEP2 could be a novel potential therapeutic target for the treatment of cervical cancer.
Introduction
XPNPEP2 (also known as aminopeptidase P or APP2) is a membrane-bound metalloprotease that catalyzes the removal of a penultimate prolyl residue from the N-termini of peptides, for example, Arg-Pro-Pro. 1 It is encoded by the human XPNPEP2 gene located on chromosome Xq25. 2 Biologically active peptides such as hormones, growth factors, neurotransmitters, coagulants, toxins, and cytokines are potential substrates for XPNPEP2. 3 XPNPEP2 may play a role in the metabolism of the vasodilator bradykinin, and most studies on XPNPEP2 are focused on cardiovascular disease. Some have reported that XPNPEP2 can affect the incidence of angiotensin-converting-enzyme (ACE) inhibitor–induced side effects of angioedema. 4 However, the relationship between XPNPEP2 and cancer has not been reported. The role of XPNPEP2 in cancer is still unknown.
Cervical cancer (CC) is the third most common female malignancy worldwide. 5 With the progress in surgery, radiotherapy, and chemotherapy, the 5-year survival rate of women with CC who undergo standardized treatment can be greater than 80%. However, once patients develop a metastasis (especially lymph node metastasis), the 5-year survival rate drops below 50%. 6 The metastasis of CC is the main cause of CC-related mortality. 7 Therefore, it is critical to elucidate the molecular mechanism of CC metastasis and identify suitable targeted therapeutics. In our study, we found that XPNPEP2 was highly expressed in CC tissues and was correlated with metastasis and the International Federation of Gynecology and Obstetrics (FIGO) stage of CC. Therefore, we hypothesized that XPNPEP2 may be associated with CC metastasis.
In this study, we first investigated the relationship between elevated XPNPEP2 expression and CC metastasis and progression. Correlations between XPNPEP2 expression and the clinicopathological characteristics of cervical tissues were further analyzed. Enhancement and suppression of XPNPEP2 promoted or attenuated, respectively, the ability of CC cells to invade and migrate. Moreover, we demonstrated that XPNPEP2 could induce epithelial–mesenchymal transition (EMT) in CC cells. XPNPEP2 overexpression could also promote the metastasis of xenografted tumors in vivo. Our results are the first to describe a novel function of XPNPEP2 in cancers via increasing invasion and metastasis of CC. Thus, XPNPEP2 could serve as an attractive target for metastatic therapy in patients with CC.
Materials and methods
Human tissue specimens
A total of 60 paraffin-embedded cervical tissue samples were obtained from 60 patients at Tongji Hospital (Wuhan, China) between 2011 and 2012, including 20 normal cervical tissue samples, 10 cervical intraepithelial neoplasia (CIN) tissue samples, and 30 CC tissue samples. Normal cervical tissue samples were obtained from patients who had undergone a hysterectomy and had no malignancies. This study was approved by the Ethics Committee of Tongji Medical College of Huazhong University of Science and Technology, and informed written consent was obtained from all subjects prior to the study.
A histopathological diagnosis of cancer was based on the World Health Organization classifications, and the clinical staging was defined according to the FIGO system. Complete clinical and pathological data were available for all the patients, and none of the patients received pre-operative radiotherapy, chemotherapy, or biological therapy. The patients characteristics are summarized in Table 1. The total of 30 CC cases has been followed up during the 5 years. Among them, 6 cases were dead.
Correlation analysis of clinicopathological parameters with expression levels of XPNPEP2 in cervical tissues.
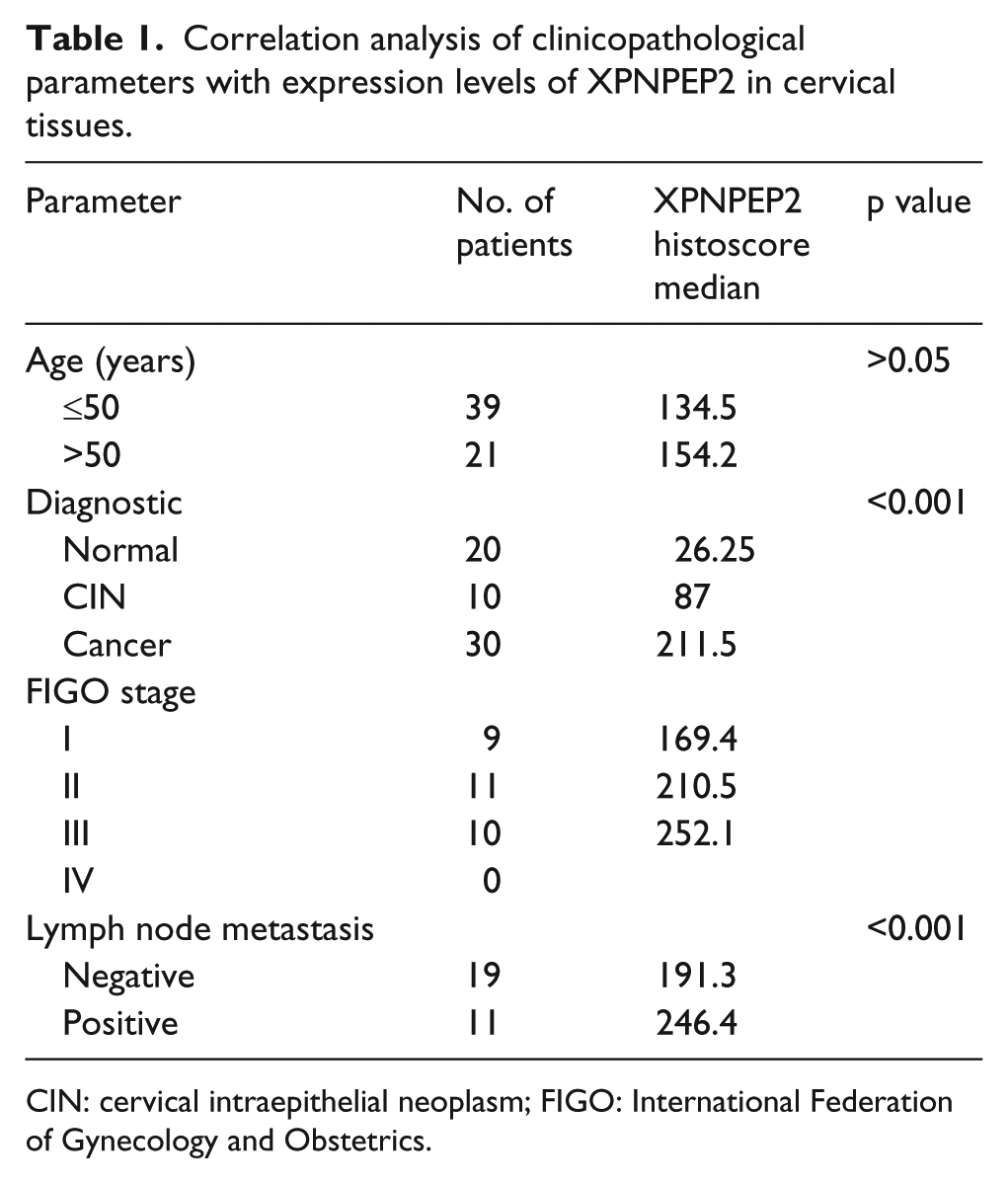
CIN: cervical intraepithelial neoplasm; FIGO: International Federation of Gynecology and Obstetrics.
Cell culture
The CC cell lines C-33A, SiHa, and HeLa and the human cervical epithelial cell line End1 were purchased from the American Type Culture Collection (Manassas, VA, USA). C-33A, SiHa, and HeLa cells were cultured in Dulbecco’s modified Eagle’s medium (DMEM) supplemented with 10% fetal bovine serum (FBS). End1 cells were cultured in keratinocyte serum-free medium supplemented with 0.1 ng/mL human recombinant epithelial growth factor, 0.05 mg/mL bovine pituitary extract, and 44.1 mg/L (final concentration 0.4 mM) calcium chloride. The immortalized human cervical keratinocyte S12 cell line was a generous gift from Kenneth Raj (Centre for Radiation, Chemical and Environmental Hazards, Health Protection Agency, Chilton, Didcot, United Kingdom). S12 cells were cultured in a 1:3 mixture of DMEM and Ham’s F-12 medium supplemented with 5% FBS (Gibco, Thermo Fisher Scientific, Carlsbad, CA, USA), 5 µg/mL insulin (Sigma-Aldrich, St Louis, MO, USA), 8.4 ng/mL cholera toxin (Sigma-Aldrich), 24.3 µg/mL adenine (Sigma-Aldrich), 0.5 µg/mL hydrocortisone (Sigma-Aldrich), and 10 ng/mL epidermal growth factor (PeproTech, Princeton, NJ, USA). All the cell lines were cultivated at 37°C in a humidified atmosphere containing 5% CO2.
Reverse transcription quantitative real-time polymerase chain reaction
Total RNA was extracted from the cells using TRIzol Reagent (Invitrogen, Carlsbad, CA, USA) and reverse transcribed into cDNA using M-MLV reverse transcriptase (Takara, Kusatsu, Japan). Reverse transcription quantitative real-time polymerase chain reaction (RT-qPCR) was performed using a Bio-Rad CFX96 system with SYBR Green. The primer sequences were as follows:
E-cadherin: forward—5′-CGAGAGCTACACGTTCACGG-3′; reverse—5′-GGGTGTCGAGGGAAAAATAGG-3′;
XPNPEP2: forward—5′-GGTCAATACCACAATGTCACTCA-3′; reverse—5′-ACGCCTCTCGTCATGTTGG-3′;
18S: forward—5′-CGTCTGCCCTATCAA-3′; reverse—5′-ATGTGGTAGCCGTTT-3′;
Snail1: forward—5′-TCGGAAGCCTAACTACAGCGA-3′; reverse—5′-AGATGAGCATTGGCAGCGAG-3′;
Vimentin: forward—5′-GACGCCATCAACACCGAGTT-3′; reverse—5′-CTTTGTCGTTGGTTAGCTGGT-3′.
The reactions were amplified as follows: 95°C for 60 s and 40 cycles of 95°C for 15 s, 57°C for 15 s, and 72°C for 45 s.
Transduction with lentiviruses
Lentiviral shRNA targeting XPNPEP2 was used to knock down XPNPEP2 in SiHa and HeLa cells (sh-xp). The lentiviral vector encoding the XPNPEP2 gene was transfected into SiHa and HeLa cells (oe-xp). An empty lentiviral vector was used as a control (negative control). All lentiviruses expressed luciferase tags for subsequent animal imaging experiments. All lentiviruses were obtained from GeneChem (Shanghai, China).
Migration assay
Cells were seeded in uncoated cell culture inserts with 8-µm pores (Corning Incorporated, New York, NY, USA). Cells were harvested and resuspended in 100 µL serum-free medium at a concentration of 5000 cells. The bottom chambers were filled with 400 µL complete medium supplemented with 20% FBS. After 24 h, the migratory cells were fixed for 20 min with 4% paraformaldehyde, stained for 10 min with 0.05% crystal violet, photographed, and counted.
Invasion assay
Invasion assays were carried out using Matrigel-coated cell culture inserts with 8-µm pores (Corning Incorporated). An aliquot of 100 µL serum-free medium containing 5000 cells was added to the inserts, and each insert was placed into a lower chamber containing complete medium supplemented with 20% FBS. After 24 h, invasiveness was evaluated by staining the cells that had migrated through the Matrigel layer to the lower surface of the filter. Invasive cells were photographed and quantified under a microscope.
Scratch assay
Cells were seeded in six-well plates. After the cells reached confluence, the monolayers were scratched with a sterile pipette tip to produce a scratch approximately 0.4–0.5 mm in width, and serum-free medium was added to the culture. After 24 h, the closure of the wound was photographed, and the area was measured using ImageJ software (NIH) and expressed as a percentage of the original area. All scratch assays were performed in triplicate.
Cell viability assay
Cells (2000 cells per well) were seeded into 96-well plates. Cell viability was assayed at 1, 2, 3, 4, and 5 days using the Cell Counting Kit-8 (CCK-8; Dojindo Laboratories, Kumamoto, Japan) according to the manufacturer’s instructions. The absorbance at 450 nm (optical density at 450 nm (OD450)) of each well was measured using an automated microplate reader.
Cell apoptosis assay
Cell apoptosis was detected using a fluorescein isothiocyanate (FITC) Annexin V Apoptosis Detection Kit (BD Biosciences, New York, NY, USA). Cells (1 × 104 cells per well) were seeded into 12-well plates. After 48 h, cells were trypsinized, washed twice with ice-cold phosphate-buffered saline (PBS), and resuspended in 1× binding buffer. Then, the cells were incubated with an FITC-Annexin V solution and propidium iodide (PI) following the manufacturer’s instructions.
In vivo study
All the female Balb/c-null mice in this study were purchased from Beijing HFK Bioscience Co. Ltd. All animal procedures were approved by the Ethics Committee for Animal Experiments of Hubei province. Briefly, 4- to 6-week-old female Balb/c-null mice were housed and maintained in laminar flow cabinets under specific pathogen-free conditions. In the direct metastasis model, stably transfected CC cells (sh-xp, NC, or oe-xp) were injected into the tails of female athymic nude mice (n = 6 per group). Approximately 6 weeks later, mice were anesthetized with 1% sodium pentobarbital and intraperitoneally injected with luciferin. After 10 min, the mice were imaged using an IVIS Spectrum system (Caliper, Hopkindun, MA, USA). Total flux (photons/s) of the metastatic tumor was analyzed using Living Image version 4.3.1 software. Metastatic tumors from each group were collected at the endpoint and subjected to immunohistochemistry (IHC) analysis.
Western blot analysis
Total cell lysates were prepared in radioimmunoprecipitation assay (RIPA) lysis buffer (Beyotime, Shanghai, China) supplemented with a protease inhibitor cocktail (Roche, Mannheirn, Germany). Protein concentrations were determined using a bicinchoninic acid (BCA) assay (Thermo Fisher Scientific). Total lysates (40 µg per sample) were subjected to sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS-PAGE) followed by immunoblotting with primary antibodies against the following proteins: XPNPEP2 (GTX109995, 1:1000; GeneTex, Irvine, CA, USA), glyceraldehyde 3-phosphate dehydrogenase (GAPDH; ab9485, 1:2000; Abcam, Cambridge, MA, USA), E-cadherin (#3195, 1:2000; Cell Signaling Technology), Snail1 (#3879, 1:2000; Cell Signaling Technology), and vimentin (#5741, 1:2000; Cell Signaling Technology). After the membranes were washed, they were incubated with the corresponding horseradish peroxidase-conjugated secondary antibodies (Santa Cruz Biotechnology, Santa Cruz, CA, USA), and the proteins were detected using an enhanced chemiluminescence system (Pierce, Thermo Fisher Scientific). The statistical data of the protein levels in the western blot were analyzed using the analysis tool in Image Lab 4.1 (Bio-Rad, Hercules, California, USA). We selected the weakest protein band in each group as the reference band.
IHC
IHC staining for XPNPEP2 (GTX109995, 1:200), E-cadherin (ab1416, 1:200; Abcam), vimentin (ab92547, 1:200; Abcam), and Snail1 (ab53519, 1:200, Abcam) expression was conducted as described below. Briefly, antigen retrieval was performed in the presence of 0.01 M ethylenediaminetetraacetic acid (EDTA) buffer (pH 9.0). After 30 min of blocking in goat serum, the slides were incubated with primary antibody overnight at 4°C followed by incubation with horseradish peroxidase–linked secondary antibodies. The staining was developed using a DAB kit (BD Biosciences) to optimize the signal intensity.
IHC scoring was evaluated based on the staining intensity (0 = negative, 1 = weak, 2 = moderate, and 3 = strong), and positively stained areas (0–100) were confirmed by three separate individuals. Resulting staining index score >150 was identified as high expression, while score ≤150 was low expression.
Statistics
The data are presented as the mean value ± standard error of mean (SEM) from at least three independent experiments. Statistical analyses were performed with Prism 6.0 GraphPad software. Single comparisons between two groups were done by Student’s t-test. Comparisons between multiple groups were done by one-way analysis of variance (ANOVA) followed by Tukey’s post-test. The Kaplan–Meier method was used for the survival rate. All the tests were considered significant if p value <0.05.
Results
XPNPEP2 correlates with malignant progression in CC
To verify the functions of XPNPEP2 in CC, IHC was utilized to analyze XPNPEP2 expression in a panel of 60 tissue specimens, including 20 normal cervical tissue specimens, 10 CIN tissue specimens, and 30 CC tissue specimens. The results showed that XPNPEP2 was highly expressed in CC tissues compared to normal cervical tissues and CIN tissues (Figure 1(a) and (b)). According to an analysis of the XPNPEP2 IHC scores, we observed that these scores were elevated in either patients with a higher FIGO stage or patients who developed metastasis (Figure 1(c) and (d)). Moreover, high expression of XPNPEP2 was significantly associated with decreased survival in CC patients (Figure 1(e)). Patients in high XPNPEP2 expression group had obviously worse overall survival than those in low-expression group. Additionally, to further validate the expression pattern of XPNPEP2 in CC, we detected its expression in cells, including normal cervical cells (End1), CIN cells (S12), and CC cells (C33A, SiHa, and HeLa). RT-qPCR and western blotting demonstrated that messenger RNA (mRNA) and protein expression of XPNPEP2 was also higher in CC cells (Figure 1(f) and (g)), respectively. These findings indicated that XPNPEP2 was overexpressed in CC and correlated with the malignant progression of CC.

IHC staining of XPNPEP2 in cervical tissues and its clinical relevance. (a) IHC detection of XPNPEP2 in a series of cervical tissues, including normal cervical tissues (n = 20), CIN tissues (n = 10), and CC tissues (n = 30). (b) Scoring of XPNPEP2 expression in IHC-stained tissues. (c) The correlation between the IHC score and FIGO stage. (d) The correlation between the IHC score and metastasis. (e) Kaplan–Meier curves for overall survival (OS) based on XPNPEP2 expression in 30 patients with cervical cancer. Resulting staining index score >150 was identified as high expression, while score ≤150 was low expression. (f) XPNPEP2 mRNA levels were assessed in cell lines using RT-qPCR. (g) XPNPEP2 protein levels were analyzed using western blotting. GAPDH served as an internal loading control. Data are expressed as mean ± SEM (*p < 0.05, **p < 0.01, and ***p < 0.001).
XPNPEP2 facilitates the invasion and migration of CC cells
In accordance with the above findings, XPNPEP2 might exert a crucial role in CC progression. To explore the impact of XPNPEP2 on CC features, CC cells were transfected with XPNPEP2-knockdown (sh-xp) or XPNPEP2-overexpressing (oe-xp) lentivirus to alter XPNPEP2 levels in CC. We confirmed the XPNPEP2 expression by western blotting and RT-qPCR (Figure 2(a) and (b)). Transwell assays revealed that the migration and invasion of CC cells were observably diminished in cells with XPNPEP2 knockdown and significantly increased in cells overexpressing XPNPEP2 (Figure 2(c) and (d)). In addition, scratch assays also confirmed that XPNPEP2 promoted CC cell migration (Figure 2(e) and (f)). Moreover, CCK-8 and apoptosis assays showed that XPNPEP2 did not affect the proliferation and apoptosis of CC cells (Figure 2(g) and (h)). Taken together, these data suggested that XPNPEP2 promoted CC cell invasion and migration without affecting cell proliferation and apoptosis.
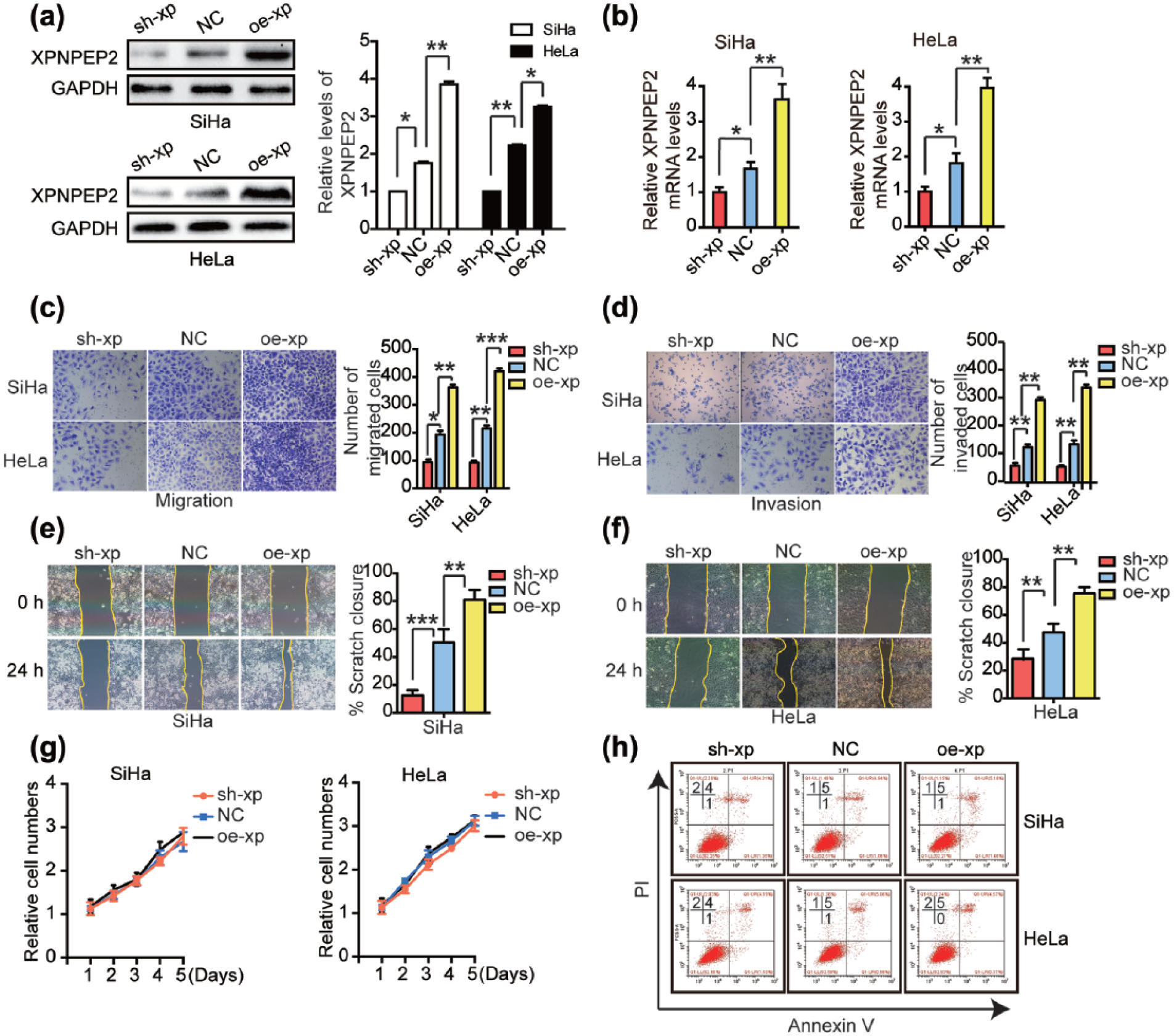
XPNPEP2 is involved in CC cell invasion and migration. (a) and (b) Western blotting and RT-qPCR analysis confirmed the overexpression and knockdown of XPNPEP2 in SiHa and HeLa cells transfected with oe-xp, sh-xp, or NC lentivirus. (c) and (d) Transwell migration and invasion assays were used to assess the migration and invasion of SiHa and HeLa cells transfected with oe-xp, sh-xp, or NC lentivirus. Representative images and statistical analysis of the wound closure capacity of (e) SiHa and (f) HeLa cells transfected with oe-xp, sh-xp, or NC lentivirus at 0 and 24 h after scratching the monolayer. (g) Cell proliferation was determined using the CCK-8 assay. Cell proliferation curves for stably transfected SiHa and HeLa cells at various indicated times were produced. (h) Stably transfected SiHa and HeLa cells were harvested after 48 h for analysis of Annexin V staining. Data are expressed as mean ± SEM (*p < 0.05, **p < 0.01, and ***p < 0.001).
XPNPEP2 induces EMT in CC cells
Next, we investigated the molecular mechanisms of XPNPEP2 that regulated cell invasion and migration. The EMT plays an important role in CC progression and metastasis. 8 EMT is accompanied by the loss of cell–cell contacts and the acquisition of migratory and motile properties that may contribute to cancer progression. 9 RT-qPCR and Western blotting were performed to examine the expression profile of several typical EMT markers in stably transfected SiHa and HeLa cells. Interestingly, we found that silencing XPNPEP2 reduced the expression of the mesenchymal marker vimentin, whereas the epithelial marker E-cadherin was upregulated in the silenced cells. In contrast, enhancing XPNPEP2 elevated vimentin expression and reduced E-cadherin expression. Furthermore, the expression of Snail1, a key transcriptional inducer of EMT, was also increased in cells with overexpressed XPNPEP2 (Figure 3(a) and (b)). Taken together, these results demonstrated that XPNPEP2 promoted the invasion and migration of CC cells by inducing EMT.
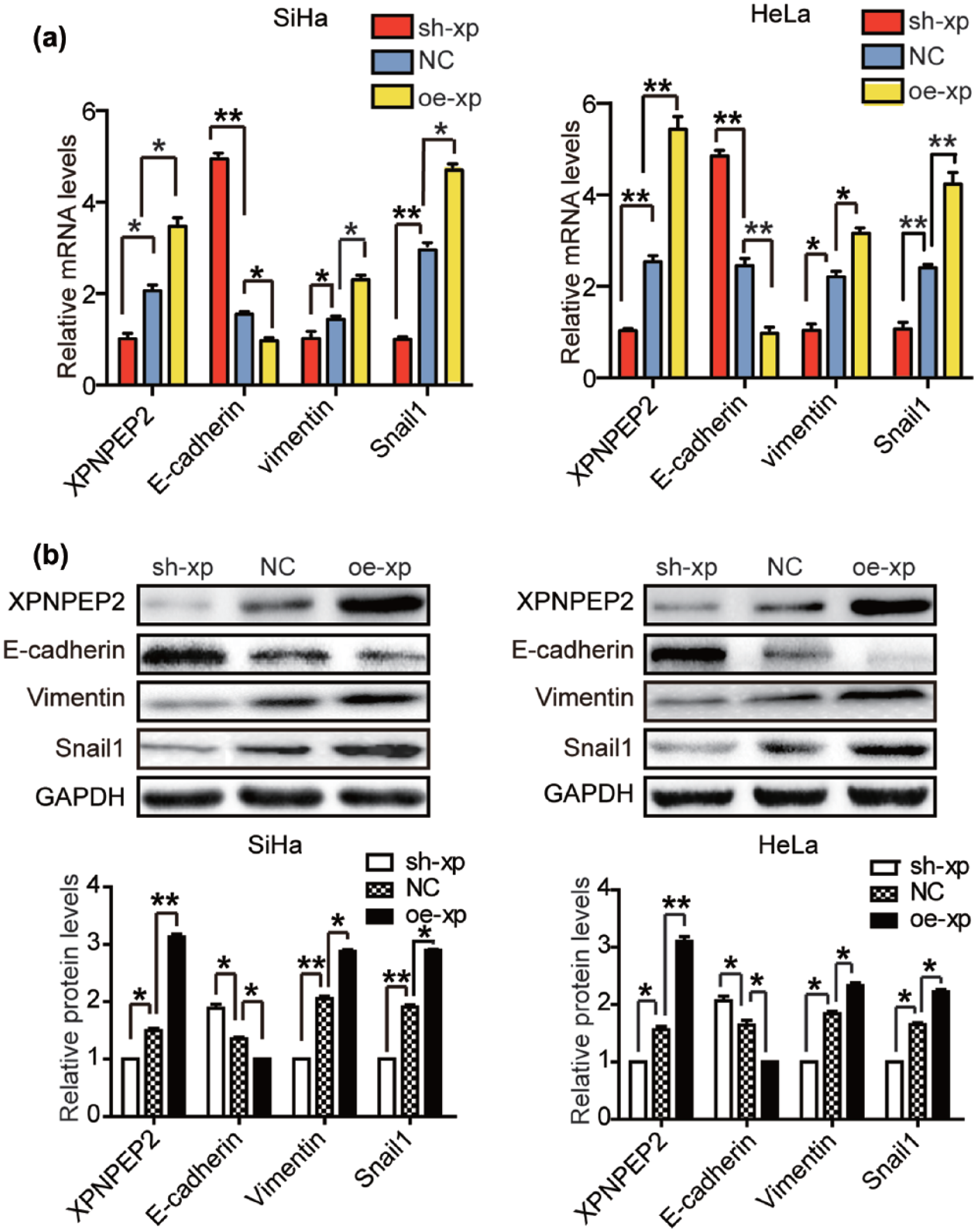
XPNPEP2 induces EMT in CC cells. (a) The mRNA levels of XPNPEP2, E-cadherin, vimentin, and Snail1 in the CC cells stably transfected with oe-xp, sh-xp, or NC lentivirus. (b) The protein levels of XPNPEP2, E-cadherin, vimentin, and Snail1 in the CC cells stably transfected with lentivirus. GAPDH was used as a loading control. Data are expressed as mean ± SEM (*p < 0.05 and **p < 0.01).
Ectopically overexpressed XPNPEP2 facilitates invasion and migration of S12 cells
In order to further demonstrate that XPNPEP2 promotes the invasion and migration of CC cells, we ectopically overexpressed XPNPEP2 in S12 cells that were low in expression of XPNPEP2. The lentiviral vector encoding XPNPEP2 gene was transfected into S12 cells to establish a stable XPNPEP2-overexpressing cell line called S12 oe-xp, and the control lentiviral vector–transfected cell line was called S12 NC (Figure 4(a)). We found that enhancing XPNPEP2 elevated vimentin expression and reduced E-cadherin expression in S12 oe-xp cells. Furthermore, the key transcriptional inducer of EMT Snail1 was also increased with the enhancement of XPNPEP2 in S12 oe-xp cells (Figure 4(b)). Transwell assays revealed that migration and invasion of S12 cells were significantly increased after XPNPEP2 overexpression (Figure 4(c)). In addition, scratch assays also confirmed that XPNPEP2 promoted S12 cell migration (Figure 4(d)). In aggregate, ectopically overexpressed XPNPEP2 promoted S12 cell invasion and migration through inducing EMT.

Ectopically overexpressed XPNPEP2 facilitates invasion and migration of S12 cells. (a) Western blotting analysis of XPNPEP2 in S12 cells stably transfected with either NC or oe-xp lentivirus. (b) Western blot analysis of XPNPEP2, E-cadherin, vimentin, and Snail1 in S12 cells stably transfected with either NC or oe-xp lentivirus. (c) Representative images and statistical analyses of cellular invasion and migration of S12 cells stably transfected with either NC or oe-xp lentivirus. (d) Representative images and statistical analysis of wound closure capacity of S12 cells stably transfected with either NC or oe-xp lentivirus at 0 and 24 h time points. The data are expressed as the mean ± SEM (**p < 0.01).
XPNPEP2 promotes CC cells metastasis in vivo
To further explore the effect of XPNPEP2 on tumor metastasis in vivo, stably transfected SiHa cells (sh-xp, NC, or oe-xp) were injected into the tails of female Balb/c-null mice. Approximately 6 weeks later, we analyzed the number of lung metastases in the mice. In the SiHa oe-xp group, the intensity of the representative bioluminescence images of these mice was obviously stronger than that of the other two groups. The bioluminescence images of mice in the SiHa sh-xp group were the weakest (Figure 5(a) and (b)). The results of HeLa cells in animals were the same as SiHa cells (Figure S1B). We demonstrated that XPNPEP2 promoted CC cell invasion and migration by inducing EMT. To elucidate these results, we excised the metastases at the end of the experiment and examined the histology of the metastatic tissue. IHC analysis of metastatic sections showed reduced staining of E-cadherin and enhanced vimentin staining, as well as increase in the expression of Snail, in the Siha oe-xp group compared with that of the Siha NC group. And IHC showed that the metastatic staining of E-cadherin was increased concomitant with attenuated vimentin expression, and Snail1 expression was also decreased in Siha sh-xp group (Figure 5(c)).

The effect of XPNPEP2 on regulating CC cells metastasis in vivo. (a) Lung metastasis. Representative bioluminescence images of mice (n = 6 each group) bearing stably transfected SiHa cells (sh-xp, NC, and oe-xp) at 6 weeks after intravenous tumor implantation. The bar graph shows the quantification of the normalized total photon counts of the direct lung metastasis model of each group. (b) IHC staining of XPNPEP2, E-cadherin, vimentin, and Snail1 in tumor metastasis sections from each group.
Discussion
This study provided the first evidence that XPNPEP2 participated in CC metastasis and could be a potent molecular marker of tumor metastasis. XPNPEP2 expression in CC was higher than that in normal cervical and CIN samples; furthermore, this expression was correlated with the FIGO stage and lymph node metastasis. In addition, our findings demonstrated that XPNPEP2 promoted migration and invasion of CC cells by inducing EMT. Importantly, our findings demonstrated for the first time the novel function of XPNPEP2 in promoting CC metastasis and suggested that XPNPEP2 could be a promising target for treating CC metastasis.
Membrane-bound aminopeptidase P is encoded by XPNPEP2 and is expressed on the surface of endothelial and epithelial cells in the kidney, liver, and lung.1,10,11 Most studies of XPNPEP2 focused on its effects in the cardiovascular system; for example, XPNPEP2 inactivates bradykinin, and a variant in XPNPEP2 was associated with ACE inhibitor-induced angioedema. 12 However, no studies have explored the relationship between XPNPEP2 and the occurrence and progression of cancer. Our data demonstrated for the first time that XPNPEP2 expression was significantly higher in CC tissue samples than in normal and CIN tissue samples. An analysis of the clinical significance of XPNPEP2 showed that high XPNPEP2 expression correlated with the FIGO stage and lymph node metastasis. These data suggested that XPNPEP2 plays a pivotal role in CC progression. XPNPEP2 is involved in the progression of CC and promoted the migration and invasion of CC cells by inducing EMT. Aminopeptidases are ubiquitously expressed in a variety of human tissues and are named by the amino acid they preferentially cleave (subtypes N, M, A, and L) aside from aminopeptidase P.13,14 Aminopeptidase M has been pharmacologically linked to cell growth, angiogenesis, and tumor progression and has been used to identify pharmacological markers of cancer.15,16 As XPNPEP2 is amino acid-specific hydrolytic enzyme, it was first proven to promote CC progression and could serve as a molecular marker of tumor metastasis. Our discovery broadens the capabilities of XPNPEP2 and shifts our understanding of XPNPEP2.
Tumor progression is involved in cell proliferation, growth, migration, invasion, and anti-apoptosis. In our study, we explored the function of XPNPEP2 on CC cell migration and invasion because of its high expression levels in metastatic tumors. Silencing XPNPEP2 using lentiviral knockdown impaired the migratory and invasive abilities in vitro and in vivo, whereas overexpressing XPNPEP2 facilitated CC cell invasion and migration. We then detected the expression levels of EMT markers in CC cell lines, which had been demonstrated to be associated with cancer progression. Overexpressing XPNPEP2 in SiHa and HeLa cells increased the expression levels of vimentin and Snail1, which are mesenchymal markers and key transcriptional inducers of EMT, and inhibited the expression of the epithelial marker E-cadherin. However, XPNPEP2 did not influence cell proliferation and apoptosis. Taken together, these data suggest that XPNPEP2 promoted tumor progression by increasing tumor cell invasion and migration.
EMT is the process by which cells lose their cell–cell contacts and acquire migratory and motile properties. 17 EMT is regarded as a supplement to tumor invasion and intravasation into the surrounding tissues, blood, and lymphatic circulation. 18 Molecularly, EMT involves the downregulation of epithelial-type proteins (such as E-cadherin) and the acquisition of mesenchymal markers, including the EMT-associated transcription factors Snail and Slug and the intermediate filament protein vimentin. 19 EMT has been reported to be correlated to the progression of cervical carcinoma. However, research on the roles of EMT and the underlying regulatory signals in CC are limited. 20 In our study, overexpression of XPNPEP2 in SiHa and HeLa cells directed the upregulation of vimentin and Snail1 as well as the loss of E-cadherin expression. Moreover, XPNPEP2 overexpression significantly increased the accumulation of CC cells in the lungs of inoculated athymic mice. Furthermore, the IHC of metastasis sections from the lungs of mice indicated that XPNPEP2 and EMT markers were positively correlated, which further confirmed that XPNPEP2 induced EMT-directed progression. These data confirmed the essential role of XPNPEP2 in CC cell metastasis.
The destruction of cervical epithelial extracellular matrix (ECM) is the most important step in the progression of cervical carcinoma in situ to invasive carcinoma. Tumor cells destroy the basement membrane mainly by secreting proteases to degrade the ECM. To date, proteases involved in the degradation of the ECM are primarily found in the matrix metalloproteinase (MMP) and a disintegrin and metalloprotease with thrombospondin motifs (ADAMTS) families; additionally, serine proteases and tyrosine proteases have also been recognized as involved in ECM degradation. 21 New studies have shown that MMPs enhance tumor cell invasion and the metastatic potential via EMT. MMP9 cooperates with the transcription factor Snail to induce EMT in CC. 22 XPNPEP2 acts as a specific proline-specific aminopeptidase capable of degrading polypeptides containing the Xaa motif on the N-terminus, including sex hormones, bradykinins, and neuropeptides. We speculated that XPNPEP2 may degrade adhesion proteins containing the Xaa motif to destroy the basement membrane, which further induces EMT in CC. However, the exact mechanism involved in XPNPEP2-mediated degradation of the basement membrane should be elucidated in future studies.
In conclusion, we showed for the first time that XPNPEP2 was overexpressed in CC tissues and high XPNPEP2 expression was correlated with an advanced FIGO stage, lymph node metastasis, and worse overall survival. Additionally, XPNPEP2 induced EMT and promoted cell migration and invasion of CC cells. Our study enhances our understanding of XPNPEP2 activity and suggests that XPNPEP2 could serve as a potential therapeutic target in metastatic CC.
Footnotes
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported by a grant from National Natural Science Foundation of China (81472444, 81172468, 81472783, 81630060, 81070536, 81403166, and 81230038), the Chinese National Key Plan of Precision Medicine Research (2016YFC0902901), the National Science-technology Supporting Plan Projects (2015BAI13B05), and “973” Program of China (2015CB553903).