
Abstract
The abilities to escape apoptosis induced by anticancer drugs are an essential factor of carcinogenesis and a hallmark of resistance to cancer therapy. In this study, we identified hTERTR-FAM96A (human telomerase reverse transcriptase–family with sequence similarity 96 member A) as a new efficient agent for apoptosome-activating and anti-tumor protein and investigated the potential tumor suppressor function in hepatocellular carcinoma. The hTERTR-FAM96A fusion protein was constructed by genetic engineering and its anticancer function of hTERTR-FAM96A was explored in vitro and in vivo by investigating the possible preclinical outcomes. Effects of hTERTR-FAM96A on improvement of apoptotic sensitivity and inhibition of migration and invasion were examined in cancer cells and tumors. Our results showed that the therapeutic effects of hTERTR-FAM96A were highly effective for inhibiting tumor growth and inducing apoptosis of hepatocellular carcinoma cells in H22-bearing nude mice. The hTERTR-FAM96A fusion protein could specifically bind with Apaf-1 and hTERT, which further induced apoptosis of hepatocellular carcinoma cells and improved apoptosis sensitivity. Our results indicated that hTERTR-FAM96A treatment enhanced cytotoxic effects by upregulation of cytotoxic T lymphocyte responses, interferon-γ release, and T lymphocyte infiltration. In addition, hTERTR-FAM96A led to tumor-specific immunologic cytotoxicity through increasing apoptotic body on hepatocellular tumors. Furthermore, hTERTR-FAM96A dramatically inhibited tumor growth, reduced death rate, and prolonged mice survival in hepatocellular carcinoma mice derived from three independent hepatocellular carcinoma mice cohorts compared to control groups. In summary, our data suggest that hTERTR-FAM96A may serve as an efficient anti-tumor agent for the treatment of hepatocellular carcinoma.
Introduction
Worldwide, hepatocellular carcinoma (HCC) is the second most common cancer (the incidence rank: fifth in men, ninth in women), which accounted for more than 90% among the primary liver cancer cases.1,2 Previous study revealed that HCC was genetically complex, multifactorial, and heterogeneous tumor.3,4 Therefore, several opinions about oncolytic treatments by targeting different signal pathways have been characterized.5,6 These cellular signal pathways presented key signal transduction for various extracellular growth factors and receptors of HCC cells. Inducing apoptosis and death of tumor cells in patients is the ultimate goal. However, the occurrence of drug resistance is a thorny problem in HCC therapy in clinical practice, and tumor cells that escape death appear in a lot of cases that are associated with acquired resistance to apoptosis induced by anticancer drugs.7,8 Tumors acquired resistance to apoptosis though early stage apoptosis or incomplete apoptotic responses to oncolytic treatments of anticancer drugs and late stage apoptosis frequently led to death of tumor cells. Anti-cancer drugs inhibited tumor growth through decreasing apoptosis resistance and inducing apoptosis of tumor cells, which further led to decreasing of neoplasm invasion. 9 Therefore, efficient treatments for HCC are urgently required for inhibition of occurrence and metastasis for patients with HCC.
Human telomerase reverse transcriptase (hTERT) is a potential target for cancer therapy, which has identified as a catalytic enzyme in the action of telomere elongation. 10 Abdul-Ghani et al. 11 reported that hTERT expressed in more than 80% of HCC cells and suggested that telomere elongation is a potential target of gene therapy for HCC. Previous study reported that adenovirus-mediated expression of hTERT COOH-27 terminal polypeptide inhibited HCC growth in mice 12 and overexpression of hTERTC27 also inhibited growth of HeLa cell and tumorigenicity in nude mouse xenografts. 13 In this study, we identified that human telomerase reverse transcriptase receptor (hTERTR) could interact with hTERT and disorder cellular metabolism of HCC cells. hTERTR is a 16-kDa polypeptide that binds to telomere dysfunction and cripples the function of hTERT in hTERT-positive HCC cells. Notably, we found that the anti-tumor efficacy of hTERTR was mediated by perturbing the activities of endogenous telomerase, thereby significantly inhibiting HCC growth in vitro, as well as strongly suppressing tumor cell proliferation and cancer cells metastasis in anti-telomerase therapy mice. In this study, we demonstrated that hTERTR could bind with hTERT and suppress aggressiveness of HCC in xenograph mice.
Family with sequence similarity 96 member A (FAM96A) is a ubiquitous and conserved protein and identified as a member of the cytosolic Fe/S protein in charge of regulator of cellular iron homeostasis and assembly machinery, which possesses apoptosome-activating and an pro-apoptotic tumor suppressor potential in HCC.14–16 FAM96A was described as a novel pro-apoptotic tumor suppressor and has therapeutic effects in gastrointestinal stromal tumors in vitro and in vivo. 17 In addition, the expression of FAM96A was decreased in HCC cells. Notably, our data indicated that the FAM96A protein could enhance apoptosis sensitivity via the mitochondrial apoptosis pathway in vitro and promote immunologic cytotoxicity for apoptotic fragments in vivo. 18
In this study, the hTERTR-FAM96A fusion protein was constructed and confirmed its anti-tumor efficacy in mice bearing HCC. Intriguingly, in contrast to single anticancer agent FAM96A, hTERTR-FAM96A presented synergetic effects on inhibition of HCC aggressiveness both in vitro and in vivo. Oncolytic efficacy has been characterized and primarily discussed by targeting hTERT and Apaf-1. This study provides multi-target fusion protein drug for the treatment of HCC cells and suggests hTERTR-FAM96A may be an efficient agent for HCC treatment.
Materials and methods
Ethics statement
This study was carried out in strict accordance with the recommendations in the Guide for the Care and Use of Laboratory Animals. All experimental protocols and animals were performed in accordance with the National Institutes of Health and approved by Committee on the Ethics of Animal Experiments Defence Research. All surgery and euthanasia were made to minimize suffering.
Construction of recombinant hTERTR-FAM96A protein
Expression system of Apex-X was used to construct the recombinant hTERTR-FAM96A protein. DNA sequence encoding 130 base pairs of FAM96A was amplified by polymerase chain reaction (PCR) from pMD-18-FAM96A and subcloned into the Apex-X plasmids (Apex-FAM96A). Cell-penetrating peptide and hTERTR were cloned into the Apex-FAM96A plasmids and named Apex-hTERTR-FAM96A. PCR and gene sequencing were used to identify plasmid. The recombinant Apex-hTERTR-FAM96A was expressed in Escherichia coli. The hTERTR-FAM96A protein was purified as described in the previous study. 16
Sodium dodecyl sulfate polyacrylamide gel electrophoresis and western blot
The purified hTERTR-FAM96A fusion protein was homogenized and analyzed by sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE) under non-reducing conditions and SDS-PAGE under reducing conditions. The purified hTERTR-FAM96A fusion protein was transferred to nitrocellulose membranes after non-reducing SDS-PAGE was performed. For western blot analysis, hTERT and Apoptin were prepared. The detection of protein was carried out by incubating the membranes with hTERT and Apaf-1 with interferon (IFN)-γ as control. The procedures were referenced from the previous work. 16 All experiments were performed in triplicate and repeated at least three times.
Cells migration and invasion assays
HCC or normal hepatic cells were cultured in Dulbecco’s Modified Eagle’s Medium (DMEM) for 48 h. All cells were suspended at a density of 1 × 105 in 500-µL serum-free DMEM and treated with phosphate-buffered saline (PBS) or hTERTR-FAM96A (0.02–0.50 mg/mL) for 48 h. All cells are seeded on top of the BD BioCoat Migration Matrigel Chambers (BD Biosciences, New York) according to the manufacturer’s instructions. For invasion assay, cells were incubated with hTERTR-FAM96A (0.02–0.50 mg/mL) or PBS for 48 h using a Matrigel Invasion Chamber (BD Biosciences) to analyze the migration of tumors cells. Cell invasion and migration were measured by stain-field microscope.
Lactate dehydrogenase and IFN-γ release assays
The cells without media were washed with PBS three times. Cells were subsequently incubated with 1% Triton X-100 at 37°C for 30 min. Lactate dehydrogenase (LDH) assay was determined using the Promega CytoTox 96 assay kit and recorded at 490 nm. For IFN-γ release assays, splenocytes were removed from the euthanized animals and monoplast suspension was made. Splenocytes were subsequently incubated with inactivated H22 cells. IFN-γ release was measured by the enzyme-linked immunosorbent assay (ELISA) in the supernatants after 72 h. Then, T cells (1 × 106) from the splenocytes were purified 2 and co-cultured with fresh H22 cells for 4 h at the effector:target ratios of 5:1, 15:1, and 40:1. Specific cytotoxic T lymphocyte (CTL) activity to the target cells was determined by LDH cytotoxicity assays. 3
RNA isolation and real-time quantitative PCR
Total RNA was isolated using Trizol reagent. Total RNA of 1 µg was used to generate complementary DNA (cDNA). The cDNA was used for real-time quantitative PCR to analyze Bcl-2, Brg-1, C-myc, Apo-1, Apaf-1, Bax, Fas, and FADD as glyceraldehyde 3-phosphate dehydrogenase (GAPDH) messenger RNA (mRNA) as an endogenous control. The amplified PCR products were quantified by measuring the calculated cycle thresholds (Ct) of sample mRNA and GAPDH mRNA. Relative changes in mRNA expression were calculated by 2−ΔΔCt. The results are expressed as the n-fold difference relative to controls (relative expression levels).
Cell culture and flow cytometry analysis
Tumor cells of SMMC-7721, H22, McA-RH7777, and HepG2 and normal cells of LO2, HHL-5, BRL-3A, and NCTC-1469 cells were purchased from the American Type Culture Collection (ATCC) and cultured in DMEM with 10% fetal bovine serum (FBS). Cell suspensions from tumors of treated mice were prepared for flow cytometric analysis (fluorescence-activated cell sorting). Cell suspensions were filtered through a 100-µm nylon strainer. Tumor cells were labeled with CD3 and CD45 plus CD4 and CD8 staining to get the frequency of CD4 and CD8 cell subsets in total infiltrated immune cells. The stained cells were analyzed by using Becton Dickinson FACScan flow cytometer. For apoptosis assay, flow cytometry was performed to detect apoptosis rates of SMMC-7721, H22, McA-RH7777, and HepG2 cells. After incubation with the 0.25-mg hTERTR-FAM96A, the cells were adjusted to 1 × 106 cells/mL with PBS and labeled with annexin V-fluorescein isothiocyanate (FITC) and propidium iodide (PI) (Annexin V-FITC Kit, BD). FACScan flow cytometer was used to analyze apoptosis rate.
Efficacy/safety assessments
Efficacy assessments included the max toxicity dose in HCC and dose-limiting toxicity in the presence of hTERTR-FAM96A. Safety assessments included the incidence rates (≥10%) of the most frequent treatment-emergent adverse events in a 30-day treatment period in experimental and control groups. The efficacy and safety data included all rats with HCC, the therapeutic drugs, and control.
Animal experiments
Specific pathogen-free (SPF) female BALB/c (6 weeks old) nude mice were purchased from Harbin Veterinary Research Institute (Harbin, China). All animals were fed under pathogen-free conditions. The density of 1 × 106 H22 cells was injected into the right flank of female BALB/c nude mice in a total volume of 200 µL. Therapy for tumor-bearing mice by hTERTR-FAM96A or Sorafenib was initiated when tumor diameters reached 6–8 mm on day 7 after tumor inoculation. Mice with HCC were randomly divided into three groups (n = 30 in each experimental group) and injected intratumorally 0.25 mg of hTERTR-FAM96A, Sorafenib, or PBS, respectively. The treatment was continued seven times at intervals of every 2 days and a total of 14 days. Tumor diameters were recorded once in every 2 days and tumor volume was calculated by using the formula: 0.52 × smallest diameter2 × largest diameter.
Immunohistochemical staining
Immunohistochemical staining was performed by an avidin–biotin–peroxidase technique. Paraffin-embedded tissue sections were prepared and epitope retrieval was performed for further analysis. The paraffin sections were subjected to hydrogen peroxide (3%) for 10–15 min, which were subsequently blocked by a regular blocking solution for 10–15 min at 37°C. Finally, the sections were incubated with anti-NK, anti-KI67, anti-Annexin, or anti-leukomonocyte antibodies at 4°C for 12 h. All sections were washed three times and incubated with secondary antibodies for 1 h at 37°C and were counterstained with hematoxylin or 4′,6-diamidino-2-phenylindole (DAPI).
Statistical analyses
All data are represented as mean ± standard error of mean (SEM). Unpaired data were analyzed by Student’s t test. Comparisons of data between multiple groups were analyzed by analysis of variance (ANOVA). For survival, data were analyzed by the log-rank test. Values of *p < 0.05 and **p < 0.01 were considered statistically significant.
Results
Construction of fusion protein hTERTR-FAM96A and in vitro activity
Figure 1(a) shows the outline of hTERTR-FAM96A, which illustrates a genetic engineering hTERTR-FAM96A fusion protein expressed by E. coli. The result in Figure 1(b) shows that molecular weight of hTERTR-FAM96A was approximately 45 kDa under constant denaturing gel electrophoresis. In Figure 1(c), western blot shows a band with approximately 45 kDa under constant denaturing gel electrophoresis and identified the purified fusion protein of hTERTR-FAM96A specifically bound to both hTERT and Apaf-1 (Figure 1(d)). The experiment of ELISA also showed that hTERTR-FAM96A was able to cross-bind both hTERT and Apaf-1, specifically (Figure 1(d)). Furthermore, the results in Figure 1(e) and (f) demonstrate that hTERT and Apaf-1 expression levels were upregulated in HCC cells, which is consistent with previous studies.17,19 These results indicate that the fusion protein hTERTR-FAM96A was successfully expressed and specifically bound with its target molecules of both hTERT and Apaf-1.
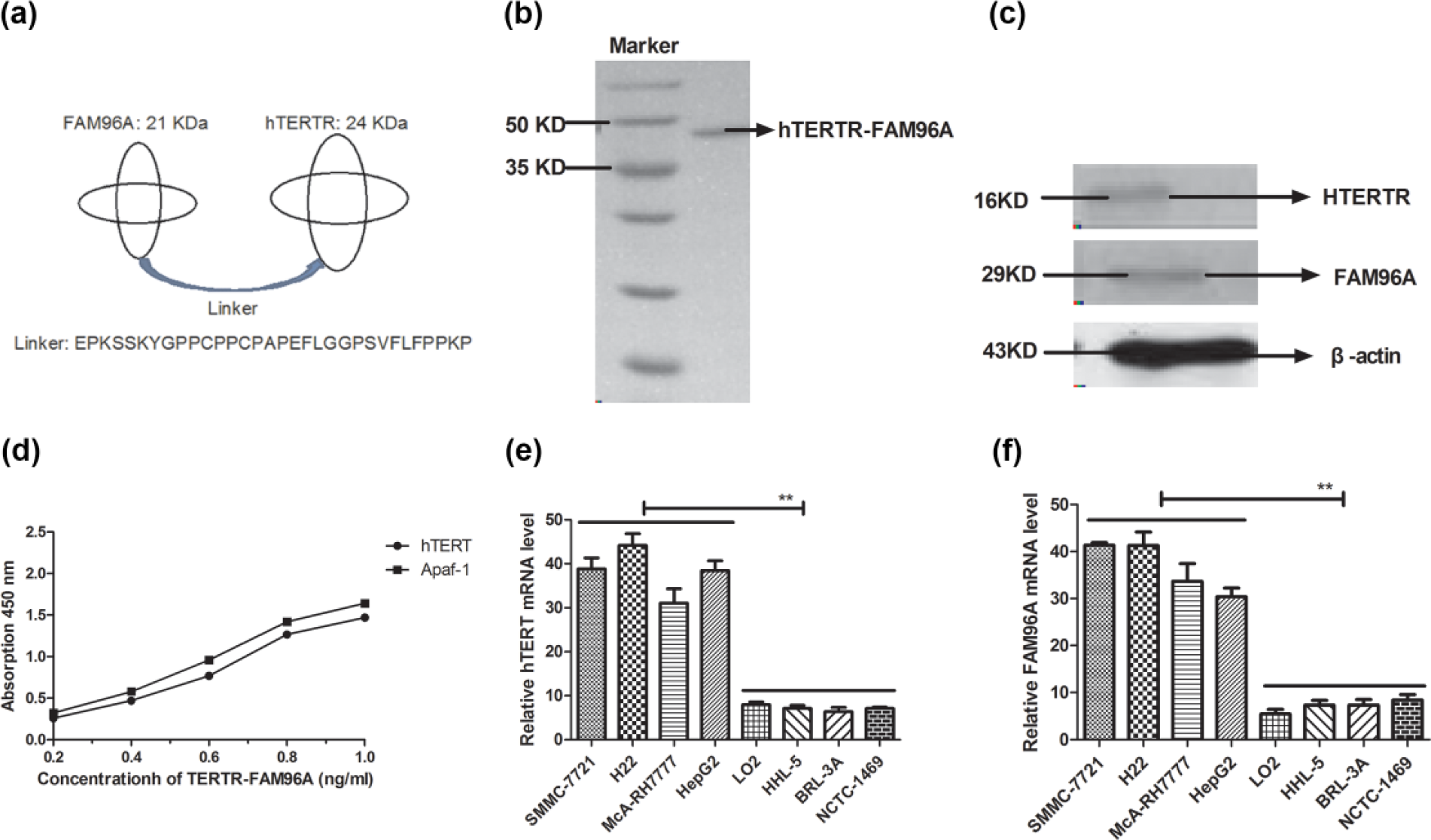
Characteristics of hTERTR-FAM96A and expression of hTERT and FAM96A. (a) Hypothetic structure of the fusion protein of hTERTR-FAM96A. The two proteins are joined together by an interlinker. The interlinker sequence is a 32-amino-acid-residue peptide from the hinge region of human IgG. (b) SDS-PAGE analysis of the purified hTERTR-FAM96A under reducing conditions. Lane 1: Molecular weight protein markers; lane 2: hTERTR-FAM96A. (c) Specificity of hTERTR-FAM96A was analyzed by western blot. (d) Bispecific interaction of hTERTR-FAM96A with both hTERT and Apaf-1. (e) Expression analysis of hTERT in hepatic carcinoma cells and normal hepatic cells. (f) Analysis of FAM96A expression in hepatic carcinoma cells and normal hepatic cells. Student paired two-tailed t test revealed a significant effect. *p < 0.05, **p < 0.01, versus control.
Cytolytic effects of hTERTR-FAM96A on normal cells and tumor cells
In this study, the inhibitory effects of hTERTR-FAM96A on tumor cell lines and normal hepatic cells were investigated to determine the cytotoxic effects of hTERTR-FAM96A by MTT (3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide) assays. The different doses of hTERTR-FAM96A showed different cytotoxic effects on tumor cell lines in a time-dependent manner, but did not affect the growth of normal hepatocytes (Figure 2(a)–(d)). The different doses of hTERTR-FAM96A (0.02–0.50 mg/mL) also demonstrated inhibiting effects for tumor cells in a dose-dependent manner after 72-h treatment. Our data showed that 0.25 mg/mL of hTERTR-FAM96A was enough to inhibit the growth of hepatic tumor cells (Figure 2(e)). Furthermore, 96-h treatment of hTERTR-FAM96A (0.25 mg/mL) showed no or slight cytotoxic effect on human or mouse normal cell lines (Figure 2(f)). In addition, hTERTR-FAM96A promoted apoptosis of tumor cell lines of SMMC-7721, H22, McA-RH7777, and HepG2, which is shown in Table 1. Furthermore, our results indicated that hTERTR-FAM96A significantly inhibited migration (Figure 2(g)) and invasion (Figure 2(h)) of HCC cells. However, no inhibitory effects were observed in normal hepatocytes. These results suggested that hTERTR-FAM96A significantly inhibited hepatocellular tumor cell growth, migration, and invasion.

The fusion protein of hTERTR-FAM96A inhibited tumor cell growth, migration, and invasion. (a)–(d) The hTERTR-FAM96A at different doses (0.02–0.25 mg/mL) showed inhibitory effects on hepatic carcinoma cells and normal hepatocytes in vitro. (e) The fusion protein of hTERTR-FAM96A inhibited tumor cell growth in vitro in a concentration-dependent manner after 72-h treatment. (f) The fusion protein of hTERTR-FAM96A (0.25 mg/mL) changed the viability of hepatic carcinoma cells but not normal hepatic cells. (g) Migration of hepatic carcinoma cells was significantly suppressed after hTERTR-FAM96A (0.25 mg/mL) treatment. (h) Invasion of hepatic carcinoma cells was significantly suppressed after hTERTR-FAM96A (0.25 mg/mL) treatment. Student paired two-tailed t test revealed a significant effect. *p < 0.05, **p < 0.01, versus control.
hTERTR-FAM96A fusion protein induced apoptosis of tumor cells.

hTERTR-FAM96A: human telomerase reverse transcriptase–family with sequence similarity 96 member A.
Therapeutic effects of hTERTR-FAM96A in HCC tumor–bearing mice
Median overall duration of treatment was 7 days across dosing cohort and was 0.02, 0.06, 0.12, 0.25, and 0.50 mg for cohort study (Table 2). Our results indicated that 0.25 mg dose of hTERTR-FAM96A once daily was identified as the maximum tolerated dose (MTD). After the last dose of drugs, the most common treatment-related adverse events were lethargy, vomiting, diarrhea, edema, proteinuria, and fatigue (Table 2). Furthermore, HCC tumor–bearing mice were used as tumor model and treated with hTERTR-FAM96A, Sorafenib, and PBS. Therapeutic effects of hTERTR-FAM96A in HCC tumor–bearing mice were recorded by tumor volume. Also, tumor cellular immunity was analyzed on day 10 after treatment. The result in Figure 3(a) shows that hTERTR-FAM96A exhibited significantly inhibitory effects for HCC tumor growth compared with PBS and Sorafenib group in a 25-day observation. The result in Figure 3(b) and (c) shows that treatment with hTERTR-FAM96A markedly downregulated expression levels of apoptosis-suppressing genes (Bcl-2 and C-myc) and upregulated expression levels of apoptosis-promoting genes (Brg-1, Apo-1, Apaf-1, Bax, Fas, and FADD) in tumors from hTERTR-FAM96A-treated tumor-bearing mice. In addition, the results in Figure 3(d) and (e) show that apoptosis rate of tumor cells was increased after treatment with hTERTR-FAM96A. Furthermore, natural killer (NK) cells were detected and presented that NK cells were increased after hTERTR-FAM96A treatment (Figure 3(f)). These data suggested that hTERTR-FAM96A improved apoptosis of HCC cells and enhanced NK cells for tumor cells on tumor surface.
Treatment-related hypertension and proteinuria by common toxicity criteria grade.

hTERTR-FAM96A: human telomerase reverse transcriptase–family with sequence similarity 96 member A.

The fusion protein of hTERTR-FAM96A effectively suppressed tumor growth and enhanced induce-tumor apoptosis potency. (a) Mean value of tumor volume in each experimental group in HCC model. (b) Analysis of apoptosis-related mRNA expression levels in TERTR-FAM96A-treatd hepatocellular carcinoma cells. (c) Apoptosis-related protein was analyzed by western blot in TERTR-FAM96A-treated tumor cells. (d) Apoptotic cells of H22 cells were determined by PI and Hochest 33258 staining. The red and blue colors represent apoptotic cells (magnification, ×200). (e) Apoptosis rate of H22 cells induced by TERTR-FAM96A was analyzed by FACS. (f) NK cells were analyzed after hTERTR-FAM96A treatment by immunofluorescence. Expression of each gene was calculated relative to the expression of housekeeping gene β-actin and the results are expressed as the n-fold difference relative to β-actin. All the values are the mean and SD of triplicate samples One-way ANOVA revealed a significant effect. *p < 0.05, **p < 0.01, versus control.
Prolongation of survival of tumor-bearing mice treated with hTERTR-FAM96A
The remaining animals with HCC were continued to observe for long-term survival analysis. Over the next 180 days, hTERTR-FAM96A markedly prolonged the survival of HCC-bearing mice (Figure 4(a)). We also determined the toxicity of CD8+ and helper cells of CD4+ in hTERTR-FAM96A-treated tumors. As can be seen from Figure 4(b), tumors from the animals treated with hTERTR-FAM96A exhibited a high degree of both CD4+ and CD8+ cell infiltration, which suggested immune responses to the apoptotic bodies and cellular debris of tumor cells. In addition, CTL responses and IFN-γ release were also assessed in H22-bearing mice. Mice treatment with hTERTR-FAM96A resulted in higher CTL activity and IFN-γ release when compared to control groups (Figure 4(c) and (d)). Meanwhile, the result in Figure 4(e) shows that hTERTR-FAM96A-treated tumors generated more apoptosis bodies analyzed by immunofluorescence compared to Sorafenib and PBS-treated tumors. The result in Figure 4(f) shows that macrophages were also increased in hTERTR-FAM96A-treated tumors after 30-day treatment. Furthermore, immune cell infiltration was analyzed and showed that more immune cell infiltration was generated after hTERTR-FAM96A treatment (Figure 4(g)). Also, immunohistochemical staining for hTERT and FAM96 on tumors showed that hTERTR-FAM96A-treated tumors decreased hTERT and increased FAM96 expression levels in tumor tissues (Figure 4(h)). These results indicated tumor treatment with hTERTR-FAM96A resulted in generation of IFN-γ release, tumor-specific CTL responses, more apoptotic bodies, and lymphocytes, which might contribute to long-term survival and inhibitory efficacy of HCC-bearing mice.

The fusion protein of hTERTR-FAM96A effectively prolonged animal’s survival and enhanced immunologic cytotoxicity. (a) Survival rate was investigated in experimental groups after TERTR-FAM96A Sorafenib, and PBS in 180-day observation. Survival rate of mice was analyzed by the log-rank test. (b) Percentage of the CD4+ and CD8+ cells were analyzed by FACS on tumor surface from HCC mice on day 25. (c) CTL response for H22 cells was analyzed by LDH assay. (d) Tumor-specific IFN-γ released from stimulated splenocytes in HCC was analyzed by ELISA. (e) Apoptosis bodies were analyzed by immunofluorescence in each experimental. (f) Macrophages were detected on tumor after TERTR-FAM96A Sorafenib, and PBS on day 25. (g) The total immune cell infiltration was analyzed in tumor cells after hTERTR-FAM96A treatment. (h) Expressions of hTERT and FAM96A were analyzed on tumors in vivo after hTERTR-FAM96A treatment on day 25. All the values are mean and SD of triplicate samples. One-way ANOVA revealed a significant effect. *p < 0.05, **p < 0.01, versus control.
Discussion
The majorities of patients with HCC are drug resistance and escape apoptosis in cancer-initiating cells. The ability to escape immunocytes-cytotoxity apoptosis is a feature of tumorigenesis and one of the key factors of resistance to cytotoxic therapy of anticancer drugs.20,21 In this study, the therapeutic effect of hTERTR-FAM96A on HCC was studied both in vitro and in vivo. Our results demonstrated that hTERTR-FAM96A not only inhibited expression levels of hTERT but also stimulated FAM96 expression in HCC cells, which contributes to apoptosis of HCC cells, and suppressed tumor growth as well as prolonged the survival of tumor-bearing mice.
In previous studies, the therapeutic effects of hTERTR by targeting hTERT on HCC have been reported and achieved anti-tumor effects in tumor-bearing mice.22–24 The hTERT protein is identified as components of the cytosolic Fe/S protein assembly machinery, and it is evolutionarily conserved and critical for cellular iron homeostasis. 14 However, hTERT expression was promoted and functions of hTERT were not clearly understood in tumor cells. In this study, we identified hTERTR as a novel target-regulating protein by binding with hTERT in human and mouse hepatic carcinoma cells. The expression of hTERT and apoptosis of tumor cells was also examined to confirm its interaction with the hTERTR and encouraging pathway, but single anticancer agent of hTERTR might not achieve the desired therapeutic effects in HCC tumor model.
FAM96A is a 21-kDa (with cell-penetrating peptide) protein and shows low expression in tumor cells. 15 Stehling et al. 14 showed that FAM96A is involved in metabolism, DNA maintenance, and protein translation, facilitating the effective induction of cell death via the mitochondrial apoptosis pathway by binding pro-apoptotic Apaf-1 protein, yet it is poorly expressed in most of tumor cells. FAM96A, as a pro-apoptotic tumor suppressor, presented therapeutic effects for tumor cells. Previous study has showed that reestablishment of FAM96A expression can enhance apoptosis sensitivity and inhibit tumor growth, which has been used to improve apoptosis resistance of cancer cells due to FAM96A loss. 17 In our work, we observed that FAM96A with cell-penetrating peptide efficiently inhibited tumor cell growth and induced apoptosis. Notably, hTERTR-FAM96A treatment markedly prolonged the survival of tumor-bearing mice in long-term survival observation.
Currently, single-agent therapy for improving apoptosis resistance of tumor cells remains several challenges due to the aggressiveness of HCC cells.25–27 Combinations of anticancer agents showed a strong therapeutic effect.28–30 Therefore, a dual function protein hTERTR-FAM96A was constructed with cell-penetrating peptide. Our research indicated the fusion protein of hTERTR-FAM96A was expressed successfully and presented anti-tumor effects both in vitro and in vivo. This kind of protein will give a reference for protein drugs in the treatment of human cancer in library and clinical practice.31,32 In addition, hTERTR-FAM96A significantly promoted CTL responses and IFN-γ release by inducing tumor cells apoptosis in murine HCC models. Among many apoptosis-related molecules involving tumor cells apoptosis, we focused on Bcl-2, Brg-1, C-myc, Apo-1, Apaf-1, and Bax in hepatic carcinoma cells. Expression changes of these molecules play essential role in tumor cells apoptosis. 33 Although the role of C-myc in the regulation of apoptosis has not been elaborated clearly, our study found C-myc was upregulated in hepatic carcinoma cells, but the role of C-myc in other tumor cells remains negligible and needed further investigation. Furthermore, hTERTR-FAM96A treatment significantly stimulated tumor-specific CTL and CD8+ cells, indicating that the formation of apoptotic bodies and debris in hepatic carcinoma cells induced by hTERTR-FAM96A could be regarded as potential cytotoxic toxicity for tumor cells mediated by cellular immunity.
In conclusion, in this study, hTERTR-FAM96A was developed as a TERT-targeting and reestablishment FAM96A-molecular therapy, which has the potential to maximize the inhibition against hepatic carcinoma tumor cells. Importantly, no obvious side effects at dose of MTD were observed with the use of hTERTR-FAM96A therapy, which suggest the beneficial effects of targeted therapy. Moreover, anti-tumor characterization of hTERTR-FAM96A is required to be further investigated to reduce the side effects and to evaluate the therapeutic value of this regimen.
Footnotes
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
This work was supported by the National Science Foundation (81401524 to WXY) of China.