
Abstract
Although radiation therapy is the primary treatment for nasopharyngeal carcinoma, radioresistance remains a major obstacle to successful treatment in many cases, and the exact underlying molecular mechanisms are still ill-defined. EMP2, epithelial membrane protein-2, was a recently identified potential oncogene involved in multiple biological processes including cell migration and cell proliferation. This study was to explore the potential relationship between EMP2 expression, nasopharyngeal carcinoma genesis, and radioresistance. EMP2 expression status in 98 nasopharyngeal carcinoma clinical samples was examined by immunohistochemical staining. As a result, most of the nasopharyngeal carcinoma tumor samples were weakly or negatively stained, while paired adjacent normal tissues were moderately or strongly stained. Moreover, patients with higher expression of EMP2 had significant longer survival times. EMP2 re-expression suppresses cell growth, induces S-phase cell cycle arrest, and promotes radiosensitivity and apoptosis in nasopharyngeal carcinoma cells. These results support that loss of EMP2 is common, and its re-expression may serve as an approach to enhance radiation sensitivity in nasopharyngeal carcinoma.
Introduction
Nasopharyngeal carcinoma (NPC) or nasopharynx cancer is the most common squamous cell carcinoma originating in the nasopharynx, behind the nose where the nasal passages and auditory tubes join the remainder of the upper respiratory tract. 1 NPC has remarkable ethnic and geographic distribution, with a high prevalence in Southeast Asia, Northern Africa, Alaska, and especially in southern China. 2 The annual incidence rate reaches 30–80 per 100,000 people per year in southern China, which is about 30- to 80-fold higher than that in the rest of the world. 3 On the basis of histological classification, NPC was classified by World Health Organization (WHO) into three types: keratinizing squamous cell carcinoma (type I), nonkeratinizing squamous cell carcinoma (type II), and undifferentiated carcinomas (type III). In Southeast Asia, NPC patients usually belong to WHO types II and III. In the West, NPC occurs sporadically and, histologically, usually belongs to WHO type I.1,2 There are various factors that are linked to NPC genesis, such as genetic susceptibility, exposure to chemical carcinogens, papillomavirus, Epstein–Barr virus (EBV) infection, and other geographical environmental factors.1,2,4–6
Radiotherapy alone or combined with chemotherapy is the mainstream treatment for NPC. Types II and III NPC are more sensitive to ionizing radiation than WHO type I. Even in advanced disease, radiotherapy combined with chemotherapy may increase survival. 7 However, radioresistance remains a major obstacle to successful treatment in many NPC cases, and the exact underlying molecular mechanisms are still ill-defined. 8
EMP2, epithelial membrane protein-2, was a recently identified potential oncogene involved in multiple biological processes including cell migration, cell proliferation, cell contraction, and cell adhesion.9–12 EMP2 was found to be highly expressed in >70% of serous and endometrioid ovarian tumors compared with nonmalignant ovarian epithelium. Anti-EMP2 diabodies significantly suppressed tumor growth and induced cell death in OVCAR5 xenografts. 13 EMP2 was also reported to be highly expressed in glioma and breast cancer, and anti-EMP2 antibody significantly inhibited cell growth, invasion, and promoted apoptosis both in vitro and in vivo.14,15 However, when examined in NPC, EMP2 was reported to be negatively or weakly staining.16,17 On the other hand, EMP2 was reported to be able to regulate the expression of integrin and influence integrin/focal adhesion kinase (FAK)-mediated signaling and cellular response.18,19 Integrin/FAK/cortactin signaling is essential for human head and neck cancer resistance to radiotherapy.20–23 So whether EMP2 plays a role in the resistance of radiation therapy of NPC by regulating integrin/FAK pathway is an interesting question.
The exact biology function of EMP2 in NPC cells is unknown. Thinking the significant antitumor effects in other tumor types through targeting EMP2, to understand the exact biology function of EMP2 in NPC is an attractive question. The purpose of this study is to examine EMP2 expression status and its functional roles in tumor cell growth and radiation resistance in NPC.
Materials and methods
Patients and tumor specimens
A total of 98 pathologically confirmed NPC patients’ clinic data were collected from Sichuan Cancer Hospital. All the tissue samples and patient data in this study were obtained with informed written consent. Available paraffin-embedded tissue blocks were retrieved from NPC patients who underwent biopsy from January 2006 to February 2011. The histological subtypes were reappraised according to the current WHO classification, and the tumor staging was re-evaluated with the 7th American Joint Committee on Cancer (AJCC) system by two pathologists, independently. The project was approved by the Scientific and Ethical Committee of Sichuan Cancer Hospital and Institute, China (No. 110563).
Cell culture
Three NPC cell lines CNE-1, CNE-2, and HNE-1 were originally obtained from the Cancer Research Institute of Guangdong Medical University and Southern Medical University, Guangdong, People’s Republic of China. Cells were maintained in Dulbecco’s Modified Eagle Medium (DMEM) or 1640 medium containing 10% fetal bovine serum (FBS), 100 U/mL penicillin, and 100 µg/mL streptomycin at 37°C in a humidified atmosphere of 5% CO2.
Immunohistochemical analysis
Tumor and tumor-adjacent normal tissue microarrays were purchased from Shanghai Xinchao Biotechnology (Shanghai, China). Immunohistochemical (IHC) analyses were performed according to microarray instruction and previously described methods. 24 Briefly, sections were heated in antigen retrieval solution and then blocked with bovine serum albumin for 20 min. The sections were incubated with the primary antibodies against EMP2 (ab105514, 1:200 dilution; Abcam, Cambridge, MA, USA) at 4°C overnight. Secondary antibody incubation and diaminobenzidine (DAB) detection were performed by using a kit from Beijing ZhongShan JinQiao Biotechnology (Beijing, China). The sections were then counterstained with hematoxylin for 2 min at room temperature. Negative control was employed by changing the specific primary antibody with non-immune serum immunoglobulins at 1:200 dilution. IHC staining results were examined and scored by two experience pathologists according to a previously reported method. 25 Total staining of EMP2 was scored as the value of the staining intensity (on a scale of 0–3: negative = 0, weak = 1, moderate = 2, strong = 3) × the percentage of cells stained (positively recorded on an ordered categorical scale: 0 = zero, 1 = 1%–25%, 2 = 26%–50%, 3 = 51%–100%), which resulted in a scale of 0–9. The evaluation was performed by two independent investigators blind to patient’s clinical information and outcome. In correlation analysis, the survival data were classified as weak (0–3), moderate (3.1–6), or strong (6.1–9) staining of EMP2.
Re-expression and RNAi assays
Complementary DNA (cDNA) of EMP2 was synthesized by Genewiz (Beijing, China). EMP2 expression was stably overexpressed using a lentiviral vector with elongation factor-1 alpha (EF-1 alpha) promoter and puromycin selectable marker. Three individual EMP2-specific small interfering RNAs (siRNAs) were purchased from Thermo Scientific. Universal scrambled negative control siRNA was purchased from Ruibo (Guangzhou, China). SiRNA transfection was performed by using X-tremeGENE siRNA reagent (Roche) according to manufacturer’s instructions.
Western blotting analysis
Western blot were performed according to previously reported study. 23 Total cellular proteins were extracted in modified radioimmunoprecipitation assay buffer (RIPA buffer) supplemented with protease inhibitor cocktail (Sigma, St. Louis, MO, USA). Equal amounts of protein were resolved on 12% sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE) gel. After blocking with 5% skimmed milk, membrane was incubated with the primary antibodies against EMP2 (ab105514, 1:1000 dilution; abcam) at 4°C overnight. Second antibody was purchased from Zhengneng Bio (Chengdu, Sichuan, China) and used at a dilution of 1:5000.
Cell growth curve and colony formation assay
A total of 20,000 cells were plated into culture dishes. The viable cells were counted by trypan blue at indicated time points. In colony formation assay, cells were seeded at a density of 600 cells per well. After adhesion, cells were irradiated at indicated doses (0–4 Gy) by an irradiator (RS-2000 biological irradiator, Rad Source Technologies, Inc., Suwanee, GA, USA). After 10–14 days, cells were stained with crystal violet after being fixed with 4% paraformaldehyde (Sigma).
Flow cytometry
For analysis of cell cycle, cells were fixed in cold 70% ethanol overnight at 4°C and then stained with 50 µg/mL propidium iodide (PI) containing 1 mg/mL deoxyribonuclease (DNase)-free ribonuclease A (RNaseA) for a minimum of 10 min before analysis by flow cytometry (BD FACS Calibr; Mod-FitLT V3.1). For analysis of apoptosis, 1 × 106 cells of each group were seeded into culture plates; after treatment with 0 Gy and 4 Gy for 24 h, cells were subjected to flow cytometry analysis using PI/Annexin-V Apoptosis Detection Kit (eBioscience, San Diego, CA, USA) according to the instructions of the manufacturer.
Statistical analysis
Student’s t-test and one-way analysis of variance (ANOVA) was used to analyze differences between groups. Survival curves were generated using Kaplan–Meier method, and the statistical analyses were performed by log-rank test. Relevance analysis of ordinal data was performed by cross χ2 test. After adjustment for age, sex, stage, modality, and the WHO classification, multivariate Cox proportional hazards regression models were used to estimate hazard ratios (HRs) with 95% confidence intervals (CIs). The significance of the other experiments was calculated by two-tailed Student’s t-test and one-way ANOVA. Values of p < 0.05 were considered significant.
Results
Patient characteristics
Totally, 98 patients with newly diagnosed, histologically confirmed, previously untreated NPC were enrolled in Cancer Hospital of Chengdu in Sichuan, China. Table 1 summarizes the demographic and clinical characteristics of NPC patients in this study. Patients were staged according to the 1997 AJCC staging system and were followed up for a median of 50 months.
Clinical pathological features of 98 clinical NPC samples.

EMP2 expression in normal and NPC tumor tissues
Through IHC staining, EMP2 were all moderately or strongly stained in NPC-adjacent normal tissues, while most of the NPC tumor samples were weakly or negatively stained (Figure 1(a)–(d)). Of the NPC tumor samples in stages I–II, 68.75% (22/32) of NPC tumor tissues showed moderate or strong staining, while in stages III–IV, the numbers of moderately/strong staining were 27.3% (18/66) (Figure 1(e)). Moreover, lower expression of EMP2 was observed in samples of T3/T4 and N2/N3 stages, while samples with higher EMP2 expression were mainly observed in the early stages, T1/T2 and N0/N1 (Figure 1(f)–(g)).

EMP2 expression and correlation analysis. (a–c) Representative IHC staining of EMP2: (a) strong EMP2 expression in NPC-adjacent normal tissues; (b) moderate EMP2 expression in NPC tissues of stages I–II; (c) negative EMP2 expression in NPC tissues of stages III–IV. (d) EMP2 expression percentages were statistically analyzed in 98 NPC clinical samples. (e–g) Different NPC stages and types. (h and i) EMP2 expression scores and tumor stages were significantly associated with survival rates of NPC patients. Scale bar, 100 µm. T: primary tumor; N: nodal status. Mod: moderate; Neg: negative.
EMP2 expression scores and NPC stages correlated with survival rates of NPC patients postradiotherapy
As shown in Figure 1(h), EMP2 expression level was significantly correlated with NPC patient’s survival. NPC patients with negative/week EMP2 expression had substantially shorter survival times than patients with moderate/strong EMP2 expression (p < 0.05). Multivariate analyses using Cox proportional hazard model showed that reduced EMP2 expression can serve as an independent prognostic factor for poor survival in patients with NPC. In the analysis of the significance between survival rates and different tumor stages, it was also observed that NPC patients at stages I–II had longer survival times than patients at stages III–IV (Figure 1(i), p < 0.01).
EMP2 re-expression induced S-phase arrest and cell growth inhibition in CNE-2 cells
To characterize the effects of EMP2 expression in NPC cells, we initially checked the expression level of EMP2 in three commonly used NPC cell lines, named CNE-2, CNE-1, and HNE-1. As shown in Figure 2(a) and (b), highest expression of EMP2 was observed in HNE-1, with lowest and moderate expression in CNE-2 and CNE-1, respectively. So we analyzed the downregulation effects of EMP2 in HNE-1 cells (Figure 2(c) and (d)). As a result, we did not observe significant changes in cell growth post RNAi knockdown. On the other hand, through cell number counting, it was found that re-expression of EMP2 significantly suppressed cell growth in CNE-2 cells (Figure 3(a)–(c)). Using flow cytometry, cell cycle analysis indicated that EMP2 re-expression induced cell cycle arrest at S-phase in CNE-2 cells (Figure 3(d) and (e)).

EMP2 expression status and knocking-down effects in NPC cell lines. The expression level of EMP2 were analyzed in three commonly used NPC cell lines by (a) Western blotting assay and subjected to (b) statistical analysis. β-actin was used as an internal control. (c–e) HNE-1 cells were subjected to RNAi knocking-down assay. (c and d) As shown, EMP2-specific siRNA significantly downregulated the protein level of EMP2 in HNE-1 cells after 48 h transfection. (e) However, the growth ability did not change significantly before and after RNAi knocking down.
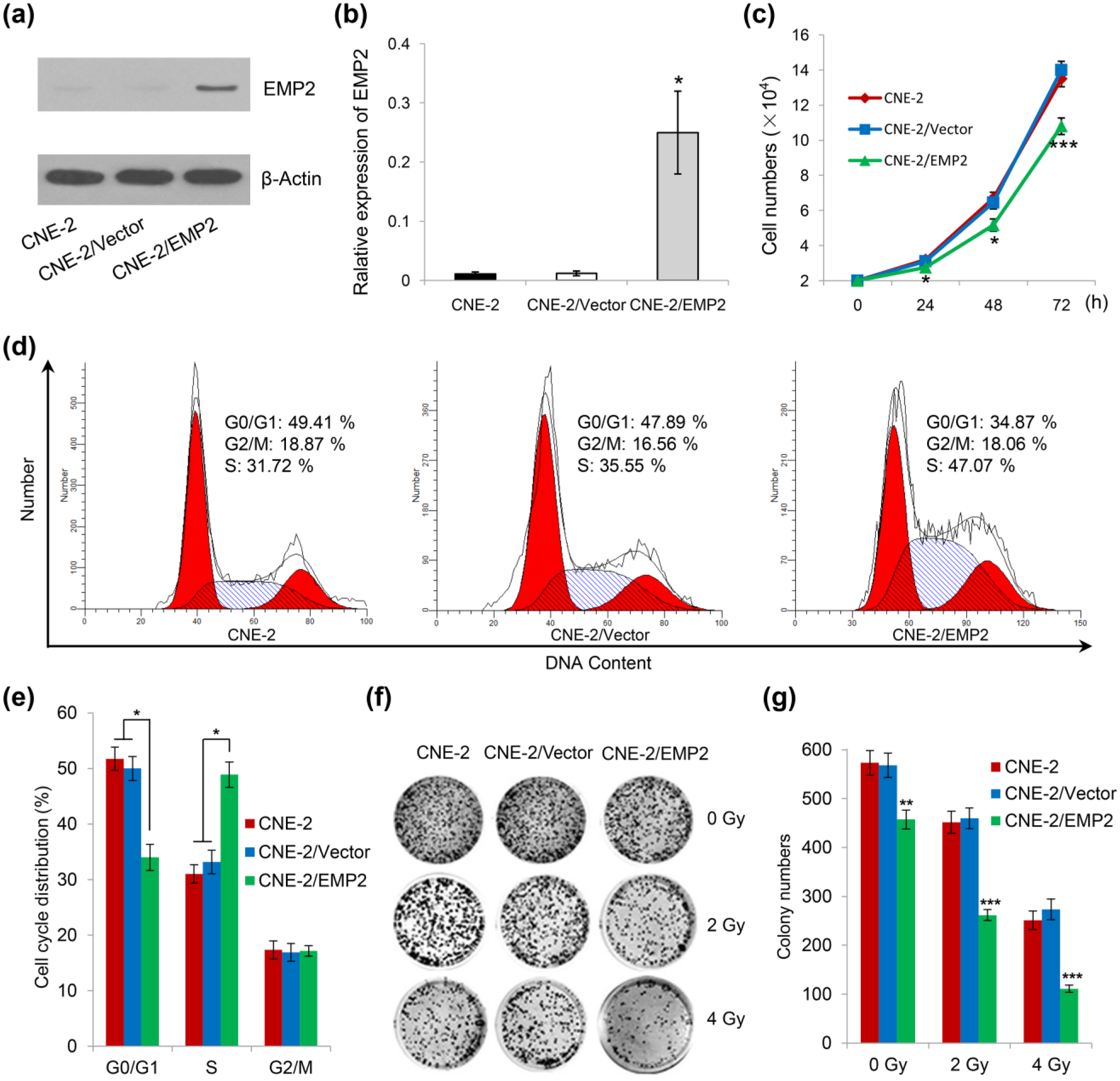
EMP2 re-expression suppresses cell growth, induces S-phase cell cycle arrest, and enhances radiosensitivity in CNE-2 cells. (a and b) EMP2 re-expression was performed by lentivirus transfection and stable cell line selection. (c) EMP2 re-expression significantly inhibits CNE-2 cell growth. (d and e) Cell cycle distribution analysis of CNE-2 cells, EMP2 stable CNE-2 cells, and vector control cells. Cells were fixed in cold 70% ethanol, stained with propidium iodide, and subjected to flow cytometry analysis. After radiation treatment, CNE-2 cells with EMP2 stable re-expression were subjected to (f) colony formation assay and (g) statistical analysis. As shown, a significant reduction in colony number was observed in EMP2 re-expression group under 2 Gy and 4 Gy radiation condition.
Re-expression of EMP2 enhanced radiosensitivity and promoted cell apoptosis in NPC cells
To examine whether EMP2 re-expression can enhance radiosensitivity in NPC cells, we performed colony formation assay (Figure 3(f) and (g)). As shown, a significant reduction in colony number was observed in EMP2 re-expression group under 2 Gy and 4 Gy radiation condition. We further examined whether EMP2 re-expression combined with radiation treatment can induce cell apoptosis by annexin V-FITC (fluorescein isothiocyanate)/PI double staining and flow cytometry assay (Figure 4). As shown, EMP2 re-expression combined with 4 Gy radiation treatment significantly promotes both early-stage and late-stage cell apoptosis in CNE-2 cells.

EMP2 re-expression combined with radiation treatment promotes cell apoptosis in CNE-2 cells. (a) Flow cytometry assays were performed to examine cell apoptosis by annexin V-FITC/propidium iodide (PI) double staining. (b) EMP2 re-expression combined with 4 Gy radiation treatment significantly promotes cell apoptosis in CNE-2 cells. Early apo: early stage apoptosis; Late apo: late stage apoptosis; Total apo: both late and early stage apoptosis.
Discussion
In this study, we examined the expression levels of EMP2 in clinical NPC samples tissues and pared adjacent normal tissues and found that it was weakly/negatively stained in 59% of tumor tissues while all paired normal tissues show moderate/strong staining. Of significance, NPC patients with negative/week EMP2 expression had substantially shorter survival times than patients with moderate/strong EMP2 expression. Moreover, multivariate analyses showed that reduced EMP2 expression can serve as an independent prognostic factor for poor survival in patients with NPC. These results are consistent with previously reported expression status of EMP2 in NPC.16,17 Furthermore, our data indicated that EMP2 re-expression significantly suppressed cell growth, induced S-phase cell cycle arrest, and promoted radiosensitivity and cell apoptosis in CNE-2 cells. Thus, EMP2 is downregulated and serves as a potential tumor suppressor gene in NPC cells. Notably, knockdown of EMP2 by RNAi in HNE-1 did not affect growth of the cells. Due to the fast-growing property of cancer cells, it is possible to fail to detect the growth-promoting effects of knocking down a tumor suppressor.
Although EMP2 has been identified as oncogene in glioma, breast, and ovary cancer types,9–12 overexpression of EMP2 induced significant cell apoptosis in both B lymphoma and NIH3T3 cells in vitro and inhibited lymphoma formation in vivo. 26 In this study, we found re-expression EMP2 alone significantly inhibited NPC cell growth, but no significant apoptosis was detected. When combined with radiation treatment, enhanced radiation sensitivity and cell apoptosis were observed in NPC cells overexpressing EMP2. Thus, EMP2 performs different or even opposite effects in different cell types.
In southern China, NPC is more endemic than any parts of the world. The disease tends to be more sensitive to ionizing radiation therapy than other head and neck cancer types, and thus the primary treatment for NPC is radiotherapy. Nevertheless, some of the NPC patients present local recurrences and distant metastases after radiotherapy due to radioresistance. The underlying mechanisms of radioresistance are largely unknown. Several genes have been suggested to be involved in radioresistance in NPC, such as integrin, MiR-205, MiR-203, RKIP, MALAT1, 14-3-3σ, Maspin, RKIP, and GRP78.20–23,27–31 Though proteomics examination of the differently expressed proteins in radioresistant cell line and its parental cell line CNE-2, Feng et al. 32 identified 34 differential proteins. In this study, we found the expression of EMP2 was almost lost in CNE-2, and its re-expression significantly promoted cell apoptosis when combined with radiation treatment. Further studies are needed to explore the exact molecular mechanisms of EMP2 in NPC genesis and radiation therapy resistance.
In this study, by examination of 98 NPC clinical samples, we found most of the tumor samples were weakly or negatively stained, and patients with higher expression of EMP2 had significant longer survival times. Moreover, EMP2 re-expression significantly suppresses cell growth, induces S-phase cell cycle arrest, and promotes radiosensitivity and apoptosis in NPC cells. Thus, re-expression or upregulation of EMP2 may help to improve clinical radiation therapy of NPC.
Footnotes
Acknowledgements
Mei Tang and Ru-yan Liu contributed equally to this work.
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported by National Natural Science Foundation of China (81102062 and 81272524).