
Abstract
Radiotherapy is a major therapeutic approach in non–small cell lung cancer but is restricted by radioresistance. Although Akt signaling promotes radioresistance in non–small cell lung cancer, it is not well understood how Akt signaling is activated. Since intracellular calcium (Ca2+) could activate Akt in A549 cells, we investigated the relationship between intracellular calcium (Ca2+) and Akt signaling in radioresistant A549 cells by establishing radioresistant non–small cell lung cancer A549 cells. The radioresistant cell line A549 was generated by dose-gradient irradiation of the parental A549 cells. The cell viability, proliferation, and apoptosis were, respectively, assessed using the cell counting kit-8, EdU labeling, and flow cytometry analysis. The phosphorylation of Akt was evaluated by Western blotting, and the intracellular Ca2+ concentration was assessed by Fluo 4-AM. The radioresistant A549 cells displayed mesenchymal morphology. After additional irradiation, the radioresistant A549 cells showed decreased cell viability and proliferation but increased apoptosis. Moreover, the intracellular Ca2+ concentration and the phosphorylation level on the Akt473 site in radioresistant A549 cells were higher than those in original cells, whereas the percentage of apoptosis in radioresistant A549 cells was less. All these results could be reversed by verapamil. In conclusion, our study found that intracellular Ca2+ could promote radioresistance of non–small cell lung cancer cells through phosphorylating of Akt on the 473 site, which contributes to a better understanding on the non–small cell lung cancer radioresistance, and may provide a new target for radioresistance management.
Introduction
Radiotherapy is a major therapeutic weapon in non–small cell lung cancer (NSCLC). It is used not only in those with oligometastases as palliative treatment 1 but also in early stage as adjuvant method. 2 Although modern radiotherapeutic methods have been developed, such as stereotactic body radiation therapy, 3 radioresistance remains a crucial issue in NSCLC treatment and restricts the efficacy of NSCLC radiotherapy. 4
In NSCLC, Akt signaling promotes radioresistance through attenuating radiation-induced apoptosis.5,6 However, it is still poorly understood how Akt is activated and subsequently which sites of Akt protein are phosphorylated. Given a recent report that intracellular calcium (Ca2+) could rapidly activate Akt in epithelial A549 cells, 7 we wonder whether Ca2+ may modulate Akt signaling in NSCLC radioresistance. Thus, we established radioresistant human NSCLC A549 cell line and investigated the potential role of intracellular Ca2+ in Akt activation in this study.
Methods
Cell culture
Human NSCLC A549 cells were purchased from American Type Culture Collection (Manassas, VA, USA). The radioresistant A549 cells were generated by dose-gradient irradiation of the parental cells. All cells were maintained in RPMI 1640 medium (PAA Laboratories, Pasching, Austria) supplemented with 10% fetal bovine serum in a humidified incubator at 37°C with 5% CO2.
Dose-gradient irradiation
Irradiation of the cells was performed with a gamma-ray generator (Philips SL 18). A549 cells were irradiated with gamma ray when they were in exponential growth phase. The cells were trypsinized and passaged after reaching confluence. Finally, the cells received irradiation with two different doses in total, that is, 22 Gy (2, 4, 4, 6, and 6 Gy) and 30 Gy (2, 4, 4, 6, 6, and 8 Gy). After irradiation, the target cells (i.e. radioresistant A549 cells) were passaged until they maintained the morphological stability and proliferation activity. The cells were passaged for five or more generations before being used for other experiments.
Cell viability assay using cell counting kit-8
A549 cells viability was evaluated using the cell counting kit-8 (CCK-8) assay (CK04; Dojindo Laboratories, Shanghai, China). Non-irradiated A549 cells and radioresistant A549 cells were plated at a density of 5000 cells/well with 100 µL culture medium in each well and then incubated in 96-well plates at 37°C with 5% CO2 for 10 h. The cells in 96-well plate were irradiated with five different doses (0, 2, 4, 6, and 8 Gy). The set of experiment for each group was repeated three times. Following that, all cells were replaced by fresh culture medium and further incubated for another 24 h. A volume of 10 µL CCK-8 was added to 100 µL medium and incubated for 2 h. Wells without cells were set as blanks. The absorbance at 450 nm with a reference of 620 nm was measured using a microplate reader (F50; TECAN, Oberdiessbach, Switzerland). Each set of measurement was repeated three times. The relative proliferation rate (%) of the A549 cells was calculated as follows: (CD450 of radioresistant A549 cells − blank)/(OD450 of original A549 cells − blank) × 100.
Cell proliferation analysis using 5-ethynyl-2′-deoxyuridine labeling
The A549 cells were seeded in 24-well plates (2 × 105 cells/well) and the cell proliferation was assessed using Cell-Light™ EdU Apollo®488 In Vitro Imaging Kit (C10310-3; RiboBio, Guangzhou, China). Cells in each well were incubated with 200 µL 5-ethynyl-2′-deoxyuridine (EdU; 50 µM) medium for 2 h. After they were washed twice with phosphate-buffered saline (PBS), the cells were incubated with 200 µL PBS containing 4% paraformaldehyde for 30 min at room temperature, followed by incubation with 200 µL glycine (22 mg/mL) in decolorization shaker for 5 min. After being washed with PBS, cells in each well were incubated with 200 µL 1× Apollo fluorescent dye solution in decolorization shaker at room temperature and away from light for 30 min. The samples were washed with PBS containing 0.5% Triton X-100 twice and incubated with 200 µL 1× Hoechst 33342 in decolorization shaker and away from light for 30 min. After washed with PBS, the samples were imaged using a blue filter (excitation 350/50; emission 480/50 nm) by fluorescence microscopy (LSM 700; Zeiss, Oberkochen, Germany).
Apoptosis detection using flow cytometry
All A549 cells were washed with PBS and then centrifuged (1000 r/min). The cells were resuspended using Annexin-binding buffer. The cells were seeded in 96-well plates (2 × 105–1 × 106 cell/mL) and incubated with 2.5 µL Annexin V-Alexa Fluor (FA101-02; TransGen Biotech, Beijing, China) at 37°C for 20–30 min. After each well was added with 1 µL propidium iodide and 400 µL Annexin-binding buffer, cells were analyzed on a flow cytometer (BD Biosciences, San Jose, CA, USA) with a 488-nm laser. The emissions were captured at 530 nm and 575 nm, respectively.
Ca2+ measurements
Radioresistant A549 cells were cocultured with verapamil for 2 h. Then, original A549 cells, radioresistant A549 cells, and the verapamil-cocultred radioresistant A549 cells were washed with Hank’s balanced salt solution (HBSS; Invitrogen, Carlsbad, CA, USA) and preincubated with 1–5 µM/L Fluo 4-AM (F312; Dojindo Laboratories) at 37°C for 10–60 min. After being washed with HBSS for three times, the cells were treated with HBSS at 37°C for 20–30 min. Fluorescent signals were captured with confocal laser scanning fluorescence microscopy (LSM 700; Zeiss, Oberkochen, Germany) using a 494-nm argon laser excitation with a 516-nm band-pass barrier filter. The images were acquired and analyzed using the ZEN image software.
Western blot
After being collected, the cells were suspended in extraction buffer (1% Triton X-100, 0.5% sodium deoxycholate, 20 mM Tris–HCl, pH 7.5, 12 mM glycero-phosphate, 150 mM NaCl, 5 mM ethylene glycol-bis(β-aminoethyl ether)-N,N,N′,N′-tetraacetic acid (EGTA), 10 mM NaF, 3 mM dithiothreitol, 1 mM sodium orthovanadate,1 mM phenylmethylsulfonyl fluoride, 20 g/mL aprotinin) and then incubated on ice for 3 × 10 min, and lysate was centrifuged (12,000g for 15 min at 4°C) to obtain the supernatant containing protein. Equal amount of proteins was separated on a 10% sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS-PAGE) and transferred to nitrocellulose membranes. A volume of 5% low-fat milk was used to block the membranes for 2 h, then, the membranes were washed thrice with Tris-buffered saline with Tween 20 (TBST) for 10 min. The membranes were probed overnight at 4°C with antibodies recognizing the following antigens: Akt1 (1:1000), p-Akt(308) (1:1000), and p-Akt(473) (1:1000). All the antibodies were obtained from Cell Signal Technology (Danvers, MA, USA). The antibody–antigen complex was detected using horseradish peroxidase–conjugated secondary antibody. Peroxidase labeling was visualized with enhanced chemiluminescence labeling using an ECL Western blotting detection system (Thermo Fisher Scientific, Waltham, MA, USA).
Statistical analysis
All statistical analyses were performed using IBM SPSS, ver. 21.0, software (IBM Co., Armonk, NY, USA). Student’s t test was used to evaluate the difference between two groups. One-way analysis of variance (ANOVA) was used to detect difference among multiple groups, followed by the Student–Newman–Keuls test or the Games–Howell test. Results were showed as mean ± standard deviation. P values less than 0.05 were considered to be statistically significant.
Results
Radioresistant A549 cells displayed the epithelial to mesenchymal transition phenotype
With dose-gradient irradiation of A549 cells to a total dose of 22 Gy and 30 Gy, respectively, we derived the radioresistant cell line A549. After five passages, the A549 cells displayed an enlarged size and mesenchymal morphology indicating to epithelial to mesenchymal transition (EMT; Figure 1), suggesting that A549 cells may undergo EMT after dose-gradient irradiation.
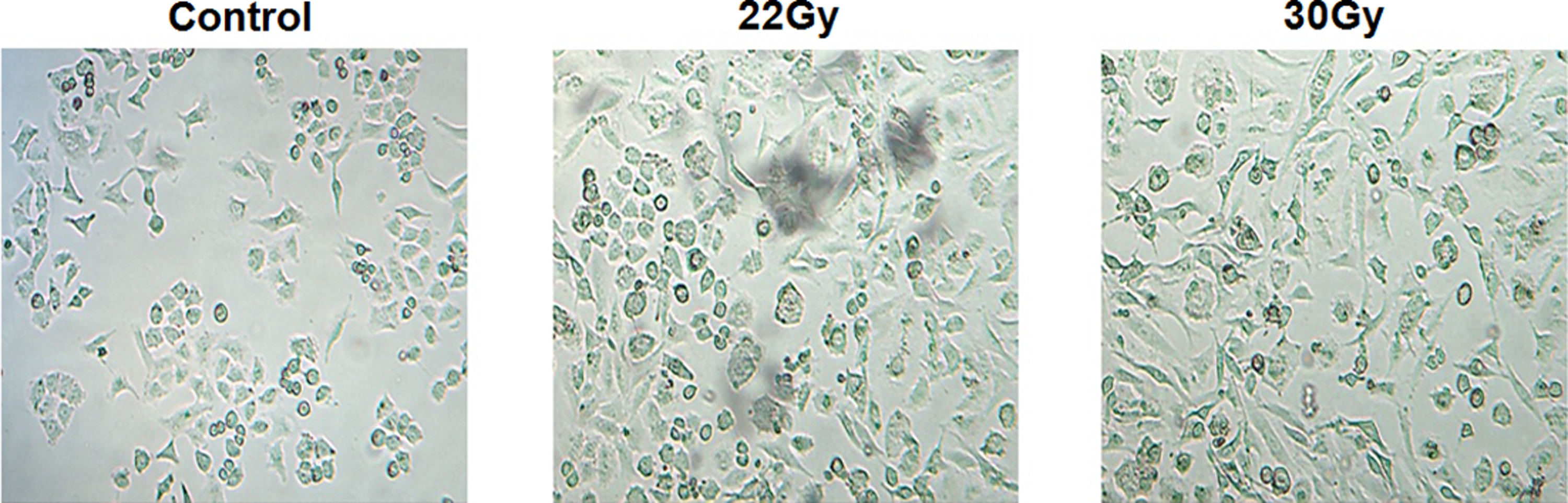
The radioresistant A549 cells displayed the epithelial–mesenchymal transition phenotype.
Radioresistant A549 cells showed downregulated radiosensitivity
We assessed the radiosensitivity of radioresistant A549 cells by assessing cell viability, proliferation, and apoptosis. In the CCK-8 test, both original A549 cells and radioresistant ones were irradiated with four different doses (4, 6, 8, and 10 Gy). The cell viability was decreased after irradiation in both original A549 cells and radioresistant ones, compared with non-irradiated cells. The radioresistant A549 cells after irradiation with four different doses had higher cell viability than original A549 cells, respectively (Figure 2(a)).
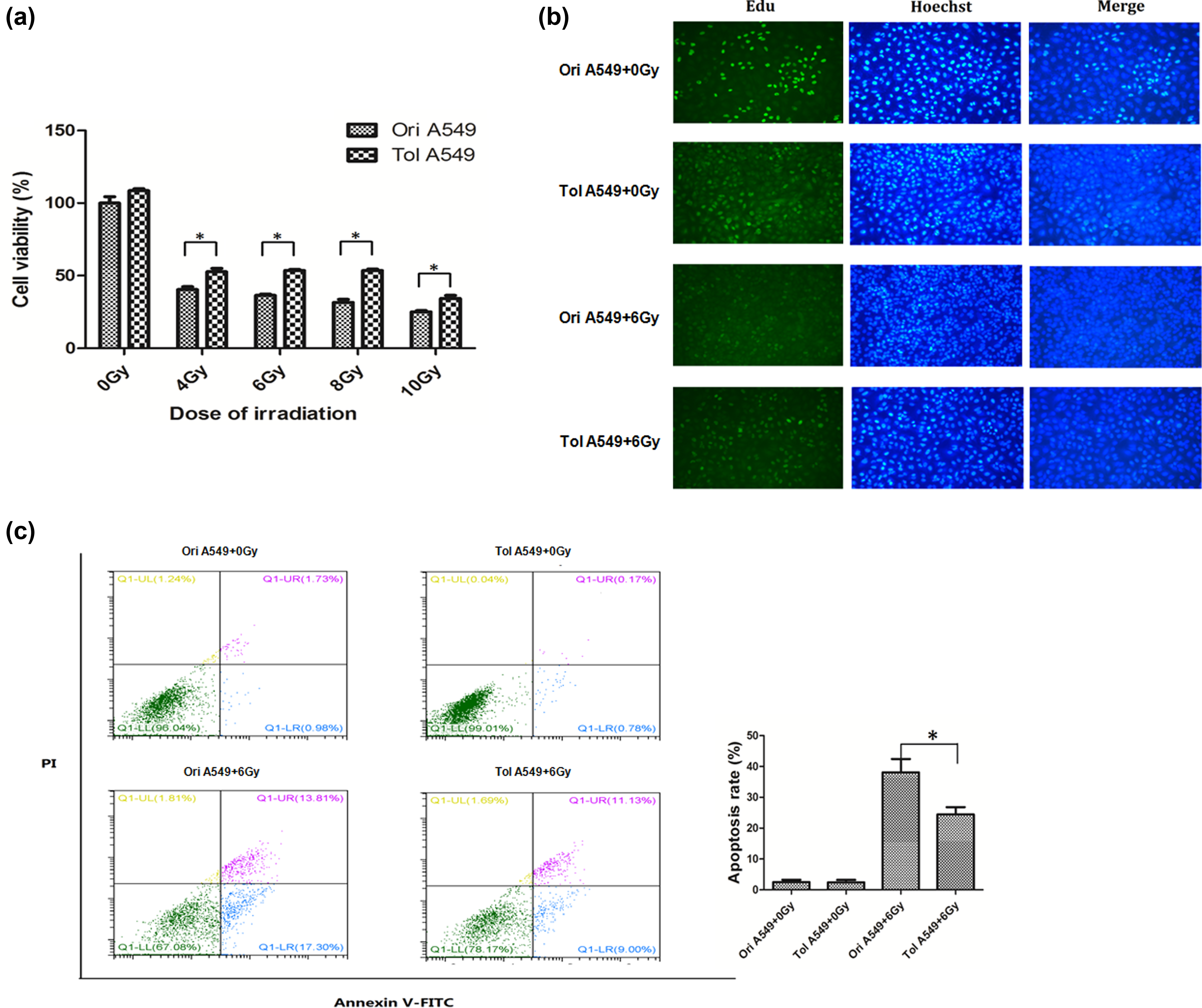
The radioresistance of radioresistant A549 cells. (a) By CCK-8 testing, both original A549 cells and radioresistant ones were irradiated with four different doses (4, 6, 8, and 10 Gy). The cell viability was decreased after irradiation in both original A549 cells and radioresistant ones, compared with non-irradiated cells. The radioresistant A549 cells after irradiation with four different doses had higher cell viability than original A549 cells, respectively. (b) In EdU labeling for cell proliferation analysis, both original A549 cells and radioresistant ones were irradiated with 6 Gy. The proliferation of radioresistant A549 cells was decreased than that of original A549 cells without 6-Gy irradiation, whereas radioresistant A549 cells showed higher proliferation than original A549 cells after 6-Gy irradiation. (c) The flow cytometric analysis showed that percentage of apoptosis in radioresistant A549 cells was less than that in original ones after 6-Gy irradiation.
In EdU labeling for cell proliferation analysis, both original A549 cells and radioresistant ones were irradiated with 6 Gy. The proliferation of radioresistant A549 cells was decreased than that of original A549 cells without 6-Gy irradiation. However, radioresistant A549 cells showed higher proliferation than original A549 cells after 6-Gy irradiation (Figure 2(b)). Moreover, the flow cytometric analysis showed that percentage of apoptosis in radioresistant A549 cells was less than that in original ones after 6-Gy irradiation (20.13% vs 31.11%; Figure 2(c)). These results suggested that the radiosensitivity of radioresistant A549 cells may be downregulated.
Intracellular Ca2+ concentration was increased in radioresistant A549 cells
The Ca2+ measurement showed that radioresistant A549 cells had a higher concentration of intracellular Ca2+ than original cells. When cocultured with calcium channel blocker, that is, verapamil, for 2 h, the intracellular Ca2+ concentration was obviously decreased in radioresistant A549 cells, validating the increased Ca2+ concentration in radioresistant cells (Figure 3).
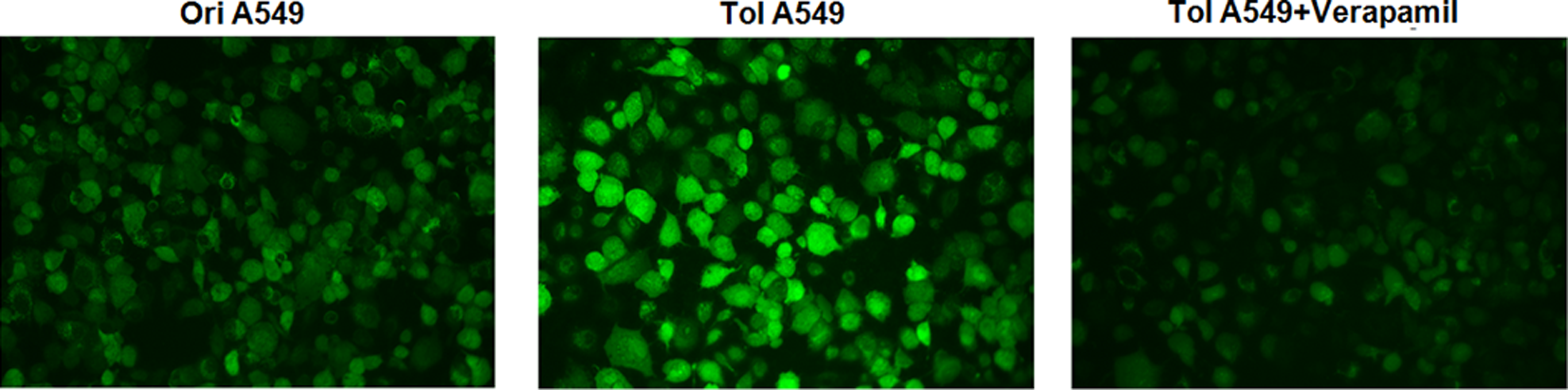
Intracellular Ca2+ concentration was increased in radioresistant A549 cells. The Ca2+ measurement showed that radioresistant A549 cells had a higher concentration of intracellular Ca2+ than original cells, which was inhibited by verapamil.
Intracellular Ca2+ activated Akt signaling in radioresistant A549 cells
The phosphorylation sites of Akt protein in radioresistant A549 cells were assessed. As Figure 4 shows, the p-Akt473 was found in both radioresistant A549 cells and original cells, whereas the site of Akt308 was not phosphorylated. Moreover, the phosphorylation level on the Akt473 site of radioresistant A549 cells was higher than that of original A549 cells. When cocultured with verapamil for 2 h, the phosphorylation level on the Akt473 site in radioresistant A549 cells was significantly decreased. This suggested that the phosphorylation on the Akt473 site in radioresistant A549 cells may be dependent on intracellular Ca2+.

Intracellular Ca2+ activated Akt signaling in radioresistant A549 cells. The phosphorylation on Akt473 site was found in both radioresistant A549 cells and original cells, whereas the site of Akt308 was not phosphorylated. Moreover, the phosphorylation level of Akt473 in radioresistant A549 cells was higher than that of original A549 cells, which was inhibited by verapamil.
Intracellular Ca2+ downregulated apoptosis of radioresistant A549 cells
A549 cells were cocultured with verapamil for 2 h before 6-Gy irradiation. The flow cytometric analysis showed that 6-Gy irradiation induced apoptosis of both radioresistant A549 cells and original cells. After 6-Gy irradiation, the percentage of apoptosis in radioresistant A549 cells was less than that in original ones (12.21% vs 27.38%), whereas coculture with verapamil led to increased apoptosis in radioresistant cells (21.79%; Figure 5). This suggested that intracellular Ca2+ downregulate apoptosis of radioresistant A549 cells.

Intracellular Ca2+ downregulated apoptosis of radioresistant A549 cells. The flow cytometric analysis showed that 6-Gy irradiation induced a lower percentage of apoptosis in radioresistant A549 cells than in original ones. But coculture with verapamil led to an increased apoptosis in radioresistant cells.
Discussion
In our study, we established radioresistant A549 cells by dose-gradient irradiation. The radioresistant cells were insensitive to irradiation as expected and had an increased intracellular Ca2+ concentration when compared with original cells. The increased intracellular Ca2+ could activate Akt via phosphorylating it on the 473 site, which could be reversed by verapamil. Our results suggest that intracellular Ca2+ could upregulate Akt signaling, leading to attenuating apoptosis in NSCLC radioresistance.
The original A549 cells were dose-gradient irradiated with a total dose of 22 Gy and 30 Gy, respectively. The morphology indicated EMT of original cells after irradiation. After additional irradiation, radioresistant A549 cells showed higher viability and proliferation, and lower apoptosis rate than original A549 cells, which is consistent with previous study and further demonstrated the radioresistance of irradiated A549 cells. 8 Thus, the radioresistant A549 cells had been established by dose-gradient irradiation in our study.
This study found that the intracellular Ca2+ concentration in radioresistant A549 cells was higher than in original cells. Without irradiation, an increased concentration of intracellular Ca2+ was involved into apoptosis of A549 cell, 9 whereas radioresistant A549 cells showed a higher Ca2+ concentration but a lower apoptosis rate than original A549 cells in our study. This finding demonstrated the radioresistance of A549 cells after dose-gradient irradiation. Both radioresistant A549 cells and original cells were damaged by additional irradiation, as our study showed the decreased cell viability and proliferation as well as increased apoptosis after additional 6-Gy irradiation. But radioresistant A549 cells were not so injured as original cells after additional irradiation. Moreover, the apoptosis rate of radioresistant A549 cells was reversed by calcium channel blocker (i.e. verapamil) before additional 6-Gy irradiation, further confirming that intracellular Ca2+ may contribute to radioresistance of A549 cells. We acknowledge that we did not coculture original A549 cells with verapamil. But the difference of intracellular Ca2+ and apoptosis between radioresistant A549 cells cocultured with and without verapamil has demonstrated the role of intracellular Ca2+ in the radioresistant cells.
Although Akt signaling involves in radioresistance of A549 cells,5,6 the intracellular modulator of Akt signaling was only microRNA-21 reported. 10 In our study, we found that an increased concentration of intracellular Ca2+ could induce phosphorylation of Akt protein. Since Akt phosphorylation on Thr(p-Akt308) and Ser(p-Akt473) was involved into apoptosis of endothelial cells,11,12 we detected whether Akt308 and Akt473 sites were phosphorylated in radioresistant A549 cells. Our study showed an increased phosphorylation on Akt473 site of radioresistant A549 cells, compared with original A549 cells. Moreover, the p-Akt473 was inhibited by calcium channel blocker, confirming the effect of intracellular Ca2+ on the phosphorylation on Akt473 site of radioresistant A549 cells. Thus, these findings suggested that intracellular Ca2+ activated Akt signaling through phosphorylating Akt473 site in radioresistant A549 cells. Combined the findings that calcium channel blocker could promote apoptosis in radioresistant A549 cells after irradiation, we suggested that intracellular Ca2+ may activate Akt signaling through phosphorylating Akt473 site, inhibiting apoptosis of radioresistant A549 cells under irradiation.
Conclusion
In conclusion, our study found that intracellular Ca2+ could promote radioresistance of NSCLC cells through phosphorylating Akt on the 473 site. The results of this study contribute to a better understanding on the NSCLC radioresistance and may provide a new target for radioresistance management.
Footnotes
Acknowledgements
Y.W. and J.H. are equal contributors and co-first authors.
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) received no financial support for the research, authorship, and/or publication of this article.