
Abstract
MicroRNA signature is altered in different disease states including cancer, and some microRNAs act as oncogenes or tumor suppressors. MiR-590-3p has been shown to be involved in human cancer progression. However, its role in hepatocellular carcinoma remains unknown. In this study, miR-590-3p level was measured, and clinicopathological features were determined in hepatocellular carcinoma tissues. The function of miR-590-3p was examined in vitro and in vivo. Real-time reverse transcription polymerase chain reaction analysis demonstrated downregulation of miR-590-3p in hepatocellular carcinoma tissues, and its downregulation was associated with a poor overall survival of hepatocellular carcinoma patients. Ectopic expression of miR-590-3p promoted growth of hepatocellular carcinoma cells, whereas its depletion inhibited cell growth. Transcriptional enhancer activator domain 1 was identified as a validated miR-590-3p target. Upregulation of transcriptional enhancer activator domain 1 was found in hepatocellular carcinoma tissues and inversely correlated with miR-590-3p. Our results indicate a tumor suppressor role of miR-590-3p in hepatocellular carcinoma through targeting transcriptional enhancer activator domain 1 and suggest its use in the diagnosis and prognosis of liver cancer.
Keywords
Introduction
Hepatocellular carcinoma (HCC) is the fifth most prevalent malignancy and the second leading cause of cancer-related deaths in the world. 1 HCC is one of the most difficult-to-treat malignancies. Surgical resection and liver transplantation are thought to be curative therapies for HCC. However, fewer than 30% of all patients are eligible candidates due to poor liver function, advanced disease at the time of diagnosis, and limited donor livers. 2 To achieve the most from available therapies, the most promising strategy is to diagnose and treat early in an early curable stage and result in long-term survival.
MicroRNAs (miRNAs) are small, non-coding RNAs with 18–22 nucleotides in length. 3 Mature miRNAs have the ability to regulate many target genes’ expression through the translational repression and degradation. 3 MiRNAs constitute a large gene regulatory network that plays important roles in various developmental and physiological processes, including cancer. 4 MiRNA profiling analysis has identified hundreds of miRNAs that are dysregulated in almost all types of human cancers. 4 Like other protein coding genes, miRNAs can also function as tumor suppressor or oncogenes. 4
Until now, miRNA profiling analysis has identified a series of miRNAs that are altered in HCC. Some of these miRNAs have been shown to be downregulated in HCC and served as a tumor suppressor, including miR-139, 5 miR-384, 6 miR-122, 7 and miR-188-5p. 8 In contrast, some miRNAs are commonly upregulated and play oncogenic role in HCC, such as miR-517a, 9 miR-151, 10 and miR-18a. 11 Recently, a report supports that miR-590-3p may function as a tumor suppressor in glioblastoma multiforme (GBM) by targeting ZEB1 and ZEB2 genes. 12 However, little is known about the role of miR-590-3p in the pathogenesis of HCC.
In this study, we first measured the expression of miR-590-3p in HCC tissues and cell lines and evaluated its expression with clinical factors and HCC patients’ overall survival. Moreover, we performed in vitro and in vivo experiments to determine the effect of miR-590-3p on the tumor growth of HCC cells. We also identified the potential target of miR-590-3p by bioinformatics analysis coupled with luciferase assay.
Materials and methods
HCC tumor specimens and cell lines
Sixty pairs of primary human hepatocellular tumor and adjacent normal tissue samples were obtained from the Departments of Oncology, Xiangya Hospital Central South University, between November 2009 and November 2011. Tissue specimens were procured in accordance with the Research Ethics Committee of Xiangya Hospital Central South University. The inclusion and exclusion criteria of HCC patients were: (a) confirmed pathologic diagnosis of HCC; (b) underwent a primary and curative liver resection; (c) not receiving preoperative chemotherapy or radiotherapy, and extrahepatic metastases; (d) no previous treatment for HCC; (e) with complete clinical information and follow-up data. The patients’ characteristics are summarized in Table 1. Four HCC cell lines (HepG2, Huh7, SK-Hep1, Hep3B) and one normal liver cell line (L02) were obtained from ATCC (Rockville, MD, USA) and were cultured as recommended by the supplier.
Correlation between miR-590-3p level and clinical factors in 60 HCC patients.
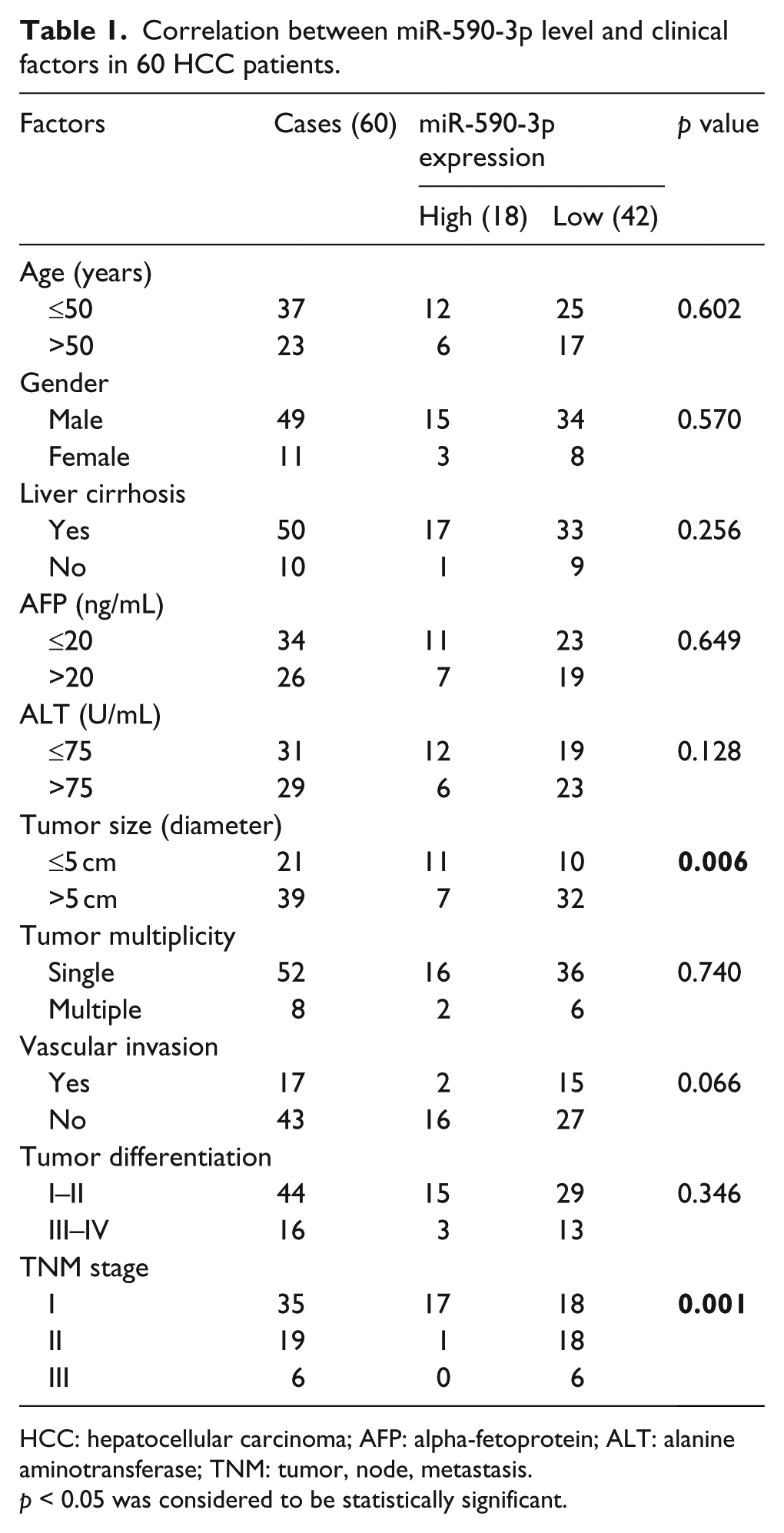
HCC: hepatocellular carcinoma; AFP: alpha-fetoprotein; ALT: alanine aminotransferase; TNM: tumor, node, metastasis.
p < 0.05 was considered to be statistically significant.
RNA extraction and real-time quantitative PCR
Total RNA was isolated from liver tissues and cell lines by Trizol (Invitrogen Carlsbad, CA, USA.), according to the manufacturer’s protocol. For mature miRNA expression, reverse transcription (RT) was performed using the PrimeScript RT reagent kit (Invitrogen Carlsbad, CA, USA). The stem-loop RT primer was designed according to Chen et al. 13 For each reaction, 100–200 ng of total RNA was mixed with 1 µL of stem-loop primer (10 µM), 10 µL 2 × miRNA RT solution mix, 1.5 µL RT enzyme mix, and made up to 20 µL of RNAse free water. The solution was incubated for 30 min at 16°C, then for 30 min at 37°C. The complementary DNA (cDNA) obtained was stored at −20°C until use. Polymerase chain reaction (PCR) reaction was performed on ABI 7500 PCR system and using SYBR Green PCR Master Mix (Applied Biosystems, Foster City, CA, USA). Each well contained in a total reaction volume of 25 µL diluted in RNAse free water: 2 µL of cDNA, prepared as described above, 0.75 µL of forward and 0.75 µL of reverse primer (see sequences detailed in Table S1), and 12.5 µL of SYBR Green PCR Master Mix. Cycling parameters were as follows: 95°C for 10 min followed by 40 cycles at 95°C (15 s) and annealing/extension at 60°C (1 min). Primers used in this study are listed in Table S1. Relative expression was calculated using the comparative delta CT (2−ΔΔCt) method.
Cell transfection
For miR precursor or anti-miR transfection, cells were plated in 60 mm dishes and transiently transfected with 50 nM pre-miR-590-3p, negative control RNA, 60 nM anti-miR-590-3p, or control anti-sense RNA (Applied Biosystems, Foster City, CA, USA).
Cell proliferation assay
Cell proliferation was monitored using a Cell Counting Kit-8 (CCK-8; Dojindo Laboratories, Kumamoto, Japan). HCC cells (5000 cells/well) transfected with pre-miR-590-3p or anti-miR-590-3p were allowed to grow in 96-well plates. Cell proliferation was documented every 24 hr following the manufacturer’s protocol. To measure cell proliferation, 10 µL of CCK-8 solution was added to each well and incubated at 37°C for 1 hr. Absorbance was measured at 450 nm using a microplate reader (Bio-Tek, Winooski, VT, USA).
Colony formation assay
HCC cells (500 cells/well) transfected with pre-miR-590-3p or anti-miR-590-3p were allowed to grow in 24-well plates. The plates were precoated with a bottom layer of 0.5% noble agar (Sigma-Aldrich St. Louis, Mo., USA). The cells were maintained at 37°C in a humidified 5% CO2 atmosphere for 2 weeks. Fresh media was added every 3–4 days to prevent the plates from drying out. Resulting colonies were stained with 1 mL 0.05% crystal violet. Colony formation was counted under the microscope. The colony was counted only if it contained >50 cells.
Cell migration and invasion assays
Cell migration and invasion were measured using 24-well, 8-µm pore Transwell inserts (Corning, NY, USA). HCC cells (1 × 105 cells/well) transfected with pre-miR-590-3p were plated onto 8-µm Transwell filters; 600 µL of complete Dulbecco’s Modified Eagle Medium (DMEM) were plated into the lower chambers. For the invasion assays, membrane insert were precoated with 45 µL Matrigel (BD Biosciences, NJ, USA). After an overnight incubation at 37°C and 5% CO2, cells in the upper membrane were removed carefully with cotton wool. Cells that had migrated or invaded through the membrane were fixed with 4% paraformaldehyde and stained with 0.5% crystal violet for 10 min. After 3–4 washes, the transwells were mounted under a coverslip, and the migrating cells were counted and photographed.
Animal experiments
All animal experiments were carried out under protocols approved by the Institute Research Ethics Committee of Xiangya School of Medicine, Central South University. BALB/c nu/nu mice from Charles River Laboratories (Beijing, China) were maintained in a specific pathogen-free condition with a 12 h light–dark cycle and provided food and water ad libitum. HepG2 cells or Huh7 cells (5 × 106) transfected with ago-miR-590-3p or agomir-negative control (GenePharma, Shanghai, China) were injected subcutaneously into the flank region of nude mice (n = 6 per group). Tumor growth was monitored every 3 days. Mice were sacrificed and tumor weighs were measured 4 weeks later.
Plasmid construction and luciferase reporter assay
For luciferase reporter gene assay, wild type (WT) or mutation of 3′ untranslated region (UTR) of transcriptional enhancer activator domain 1 (TEAD1) was cloned into the luciferase reporter plasmid pMIR-REPORT vector (Promega, Madison, WI, USA). HCC cells were transfected with 50 ng/well TEAD1-3′UTR-wt or TEAD1,3′UTR-mut, and 50 nM/well pre-miR-590-3p mimic or control RNA using Lipofectamine 2000 reagent (Invitrogen). The preparation of cell lysates and the dual-luciferase assays, using the dual-luciferase reporter assay system (Promega), were performed 24 h after transfection. The firefly luciferase activity was normalized for transfection efficiency based on the sea pansy luciferase activity.
Western blotting
Whole cells were lysed in radioimmunoprecipitation assay buffer (RIPA) buffer with protease inhibitor. Total protein (50–200 µg) was resolved by sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE), transferred onto the polyvinylidene fluoride membrane, and subjected to immunoblot analysis. The rabbit anti human TEAD1 (1:500; Cell Signaling Technology, Danvers, MA, USA) and β-actin (1:2000, Cell Signaling Technology) were incubated with the blot overnight at 4°C. The bands were visualized using the ECL system (GE Healthcare, Piscataway, USA) after incubation with secondary antibody.
Statistical analysis
Statistical significance of differences between groups was analyzed by unpaired Student’s t test, and p < 0.05 was considered to be statistically significant. The correlation between miR-590-3p and TEAD1 messenger RNA (mRNA) levels was analyzed by two-tailed Pearson correlation test. The survival rate of overall survival (OS) patients was calculated using Kaplan–Meier survival analysis. All real-time reverse transcription polymerase chain reaction (RT-PCR, assayed in triplicate), western blotting, and transfection experiments were repeated twice, and reproducible results were obtained. SPSS 19.0 was used for statistical analyses.
Results
MiR-590-3p was downregulated in human HCC tissues
We first measured miR-590-3p level in primary human HCCs and pair-matched normal liver tissues by quantitative RT-PCR (qRT-PCR). Among the 60 HCC samples analyzed, miR-590-3p level was downregulated in 42 HCC samples (Figure 1(a)). We then evaluate the clinical significance of miR-590-3p in HCC. The result showed that miR-590-3p level was inversely correlated with tumor size (p = 0.006) and tumor, node, metastasis (TNM) stage (p = 0.001) (Table 1). In addition, patients with lower miR-590-3p level had a shorter overall survival than patients with high miR-590-3p levels (Figure 1(b)).
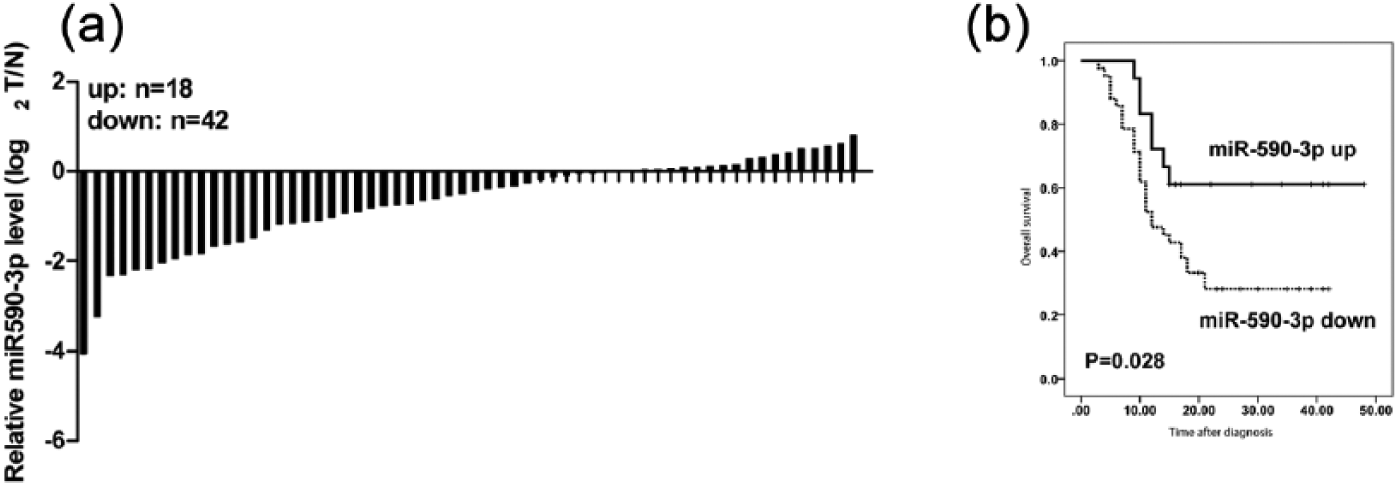
MiR-590-3p was downregulated in human HCC tissues and corrected with poor overall survival. (a) The expression of miR-590-3p in 60 paired of HCC tissues and adjacent normal liver tissues was detected by qRT-PCR. (b) Kaplan–Meier curve for HCC patients classified as high or low miR-590-3p expression; the p value was calculated using the log-rank test.
MiR-590-3p modulated growth of HCC cells
We, therefore, investigated its growth regulatory property of miR-590-3p in HCC cells. We first measured the miR-590-3p level in four HCC cell lines and one normal liver cell line by qRT-PCR. MiR-590-3p level was downregulated in four HCC cell lines (HepG2, Huh7, SK-Hep1, Hep3B); compared with normal liver cell line L02 (Figure S1(A)), these data suggest that the expression level of miR-590-3p is downregulated in human HCC. Next, we explored the effect of miR-590-3p on HCC cell growth in HepG2 and Huh7 cells. As shown in Figure 2(a) and (b), overexpression of miR-590-3p by precursor transfection reduced growth in both HepG2 and Huh7 cells. In contrast, depletion of endogenous miR-590-3p by transfecting anti-miR-590-3p resulted in accelerated growth of HCC cells (Figure S1(B) and (C)). In addition, overexpression of miR-590-3p also inhibited colony formation in both HepG2 and Huh7 cells (Figure 2(c) and (d)), whereas inhibition of miR-590-3p in HCC cells promoted their colony formation (Figure S1(D) and (E)). These results demonstrate growth regulatory property of miR-590-3p in vitro.
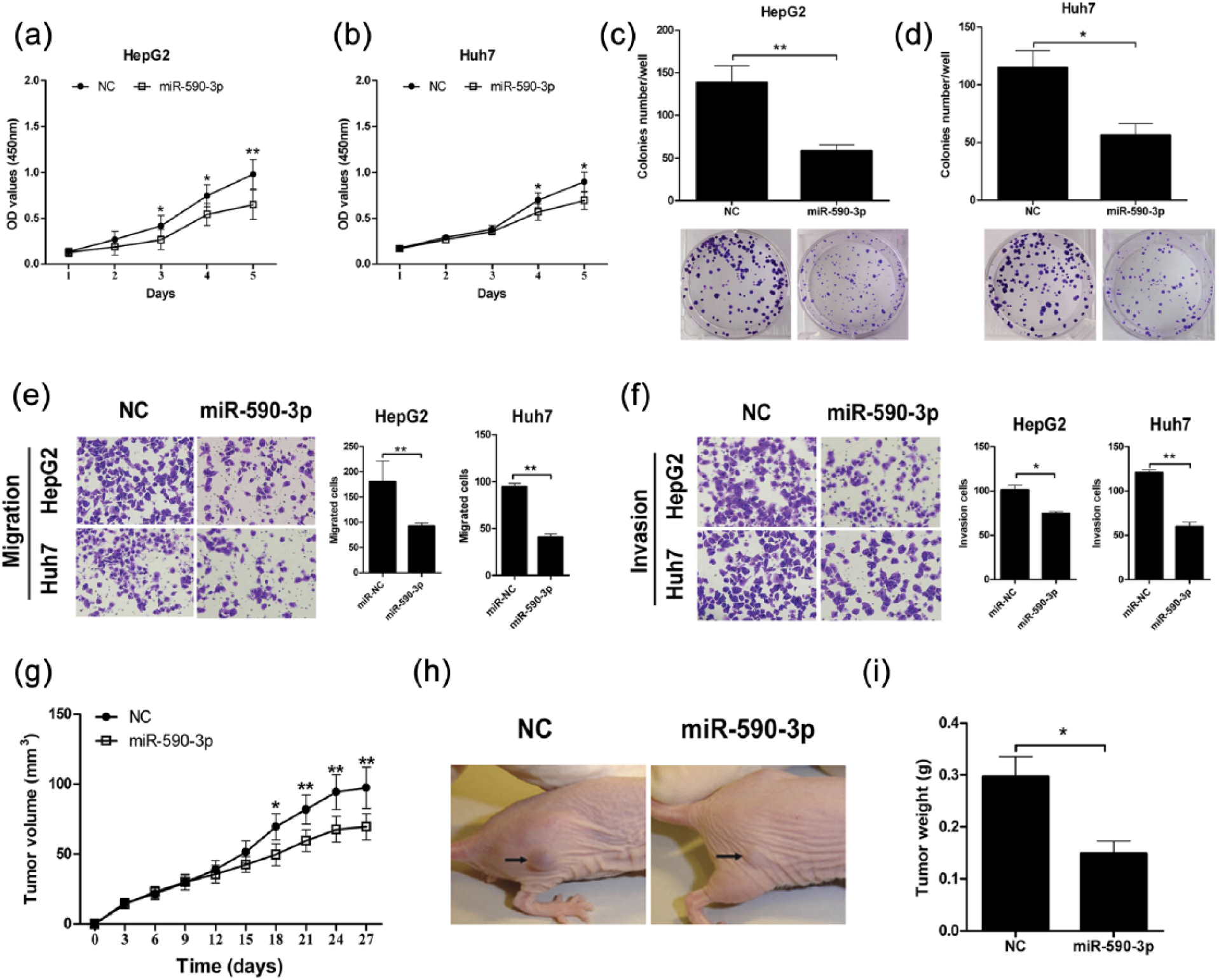
MiR-590-3p suppressed HCC growth in vitro and in vivo. HepG2 and Huh7 cells were transfected with pre-hsa-miR-590-3p or negative control (NC) RNA, respectively. (a and b) Overexpression of miR-590-3p inhibited HepG2 and Huh7 proliferation measured by a CCK-8 assay at the indicated time. (c and d) Overexpression of miR-590-3p inhibited colony formation in HepG2 and Huh7 cells. (e and f) Representative images of and quantitative data on the migration and invasion of HepG2 and Huh7 cells after pre-hsa-miR-590-3p transfection. (g) Overexpression of miR-590-3p in HepG2 cells inhibited tumor growth in a xenograft model. (h) Representative image of tumors 21 days after cell injection were shown. (i) Tumor weights were measured on day 27. Data shown are mean ± SD from three independent experiments.
We also performed cell invasion and migration assay to identify the miR-590-3p functions in HCC cell lines. The transwell assay showed that overexpression of miR-590-3p significantly decreased HepG2 and Huh7 cells migration and invasion (Figure 2(e) and (f)). We further confirmed in vitro findings using an in vivo tumor model. As shown in Figure 2(g)–(i), overexpression of miR-590-3p in HepG2 cells significantly inhibited xenograft tumor growth compared with their controls. Similar results were obtained for Huh7 cells (data not shown). Collectively, our data showed that miR-590-3p suppresses HCC cell growth in vitro and in vivo.
TEAD1 was a direct functional target of miR-590-3p
Next, we identified the possible target of miR-590-3p that may potentially be involved in the suppressive effects on HCC cell growth. Among the numerous genes predicted by TargetScan and/or miRanda, TEAD1, a transcription factor belongs to the TEAD family, which plays an oncogenic role in tumor progression, was a validated target of miR-590-3p (Figure 3(a)). TEAD1 harbors a conserved miR-590-3p site in its 3′UTR. Dual-luciferase reporter assay showed that overexpression of miR-590-3p remarkably suppressed the luciferase signaling of wt 3′UTR, but not mutant TEAD1 3′UTR in HepG2 and Huh7 cells (Figure 3(b) and (c)). Western blot analysis showed reduced expression of TEAD1 in HepG2 and Huh7 cells transfected with pre-miR-590-3p (Figure 3(d)), indicating miR-590-3p can target TEAD1 in HCC cells.
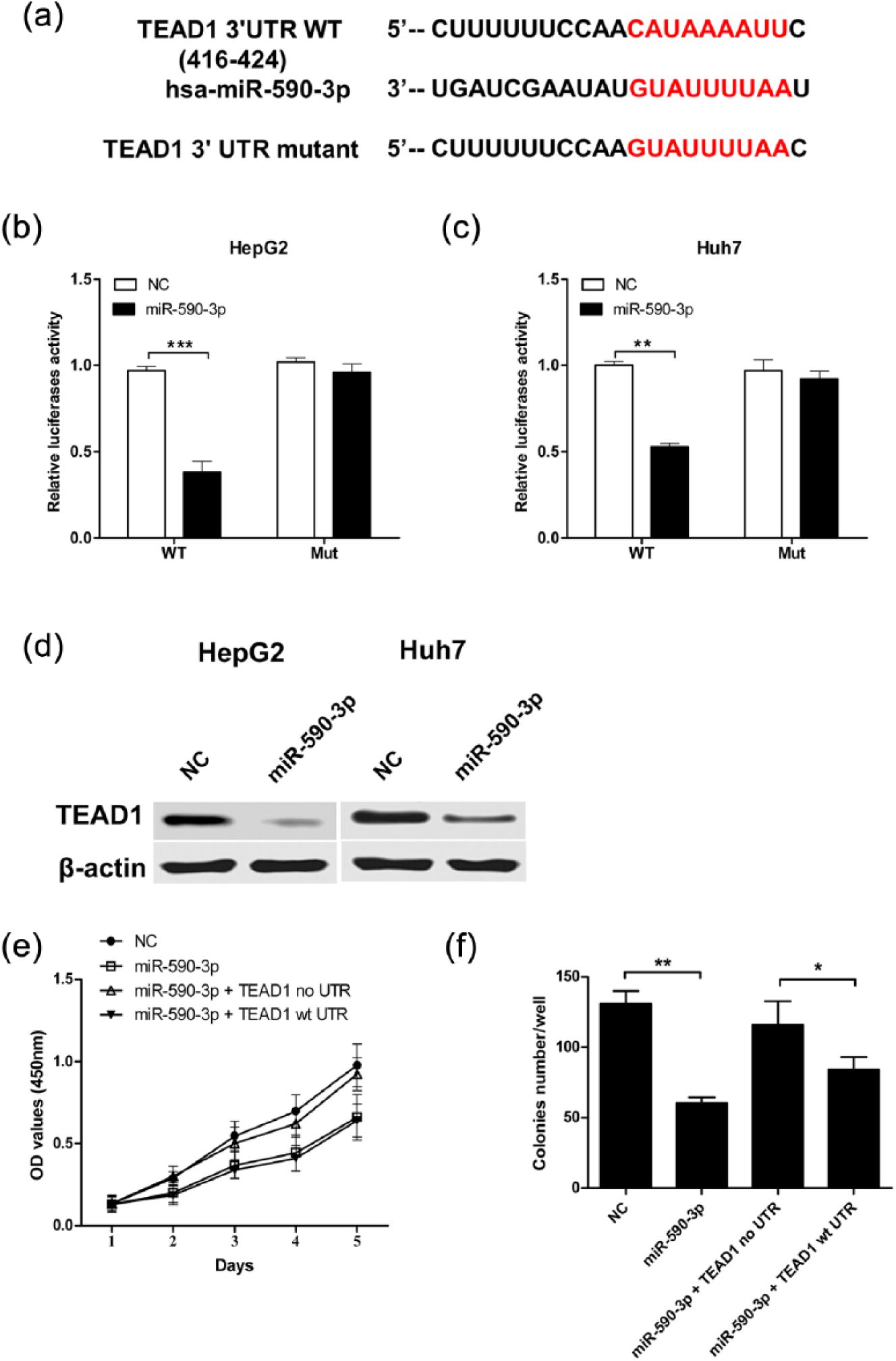
MiR-590-3p directly targeted TEAD1 in HCC cells. (a) Sequence alignment of wild-type (WT) and mutated (Mut) putative miR-590-3p-binding sites in the 3′UTR of TEAD1. (b and c) Relative luciferase activities of plasmids carrying WT or mutant TEAD1 3′UTR in HepG2 and Huh7 cells co-transfected with pre-hsa-miR-590-3p or miR-NC. (d and e) The protein levels of TEAD1 in HepG2 and Huh7 cells after co-transfection were examined by western blot. (e and f) HepG2 cells were co-transfected with pre-hsa-miR-590-3p or miR-NC and TEAD1 plasmid with or without 3′UTR. (b) The effect of miR-590-3p on cell proliferation. (c) Colony formation was rescued under the condition of overexpression of TEAD1 without 3′UTR. Data shown are mean ± SD from three independent experiments.
To further check whether TEAD1 is a direct target of miR-590-3p, HepG2 cells were co-transfected with pre-miR-590-3p and TEAD1 with or without 3′UTR. Overexpression of TEAD1 without 3′UTR has no inhibitory effect on cell growth and colony formation (Figure 3(e) and (f)). Thus, TEAD1 is a direct and functional target of miR-590-3p in HCC cells.
High TEAD1 level was inversely correlated with miR-590-3p in HCC tissues
Next, we investigated whether the level of TEAD1 was correlated with dysregulation of miR-590-3p in HCC tissues. The results showed high level of TEAD1 in HCC tissues (Figure 4(a). The survival curve showed that patients with high TEAD1 level had a significantly shorter overall survival (p = 0.002) than those with low TEAD1 expression (Figure 4(b)). Comparison of miR-590-3p and its target showed inverse correlation between the two (r = −0.31, p = 0.015) (Figure 4(c)). These data showed reciprocal regulation of suppressor miR-590-3p and its tumor oncogenic target TEAD1 in human HCC.

TEAD1 was upregulated and inversely correlated with miR-590-3p in HCC tissues. (a) The expression of TEAD1 in 60 pairs of HCC tissues and adjacent normal liver tissues was detected by qRT-PCR. (b) Kaplan–Meier curve for HCC patients classified as high or low TEAD1 expression; the p value was calculated using the log-rank test. (c) The correlation between miR-590-3p and TEAD1 mRNA levels in HCC tissues analyzed using Pearson correlation test.
Discussion
It is now well established that microRNAs and their key targets are either elevated or declined in almost all types of cancer. 4 Although a few altered microRNAs exhibit oncogenic or tumor suppressor properties, the biological functions of most microRNAs remain to be elucidated. To our knowledge, the present study is the first comprehensive analysis of the biological role of miR-590-3p in the progression of HCC. Significant downregulation of miR-590-3p in HCC tissues was found, and its downregulation was associated with a poor overall survival of HCC patients. Moreover, ectopic expression of miR-590-3p regulated HCC cell proliferation, inhibited the migration and invasion in vitro, and inhibited tumor growth in vivo through targeting TEAD1. Upregulation of TEAD1 was found in HCC tissues and inversely correlated with miR-590-3p. Therefore, our results suggest a tumor suppressor role of miR-590-3p in HCC through targeting TEAD1.
In our study, the diagnosis of HCC was made at an early stage of the disease. In total, 58.3% were in stage I; only 10% were stage III. This may have produced a shift toward lower stages of HCC. Thus, our HCC population may be skewed toward higher median survival. Therefore, larger sample size will be needed to confirm and extend our findings in future.
A significant observation of the present study is the dysregulation of miR-590-3p and its target TEAD1 in HCC. MiR-590-3p has been reported to be downregulated in GBM tissues and all of the GBM cell lines. 12 Notably, the expression of ZEB1 and ZEB2, a tumor suppressor target of miR-590-3p, was significantly increased in HCC tissues.14–16 Our study revealed that TEAD1, the target of miR-590-3p, also increased in HCC tissues. Thus, it is conceivable that downregulation of miR-590-3p with concomitant upregulation of its tumor oncogene target TEAD1 plays a causal role in HCC pathogenesis.
By using bioinformatics analysis, TEAD1 was selected as a potential target of miR-590-3p. The transcription factor TEAD (TEA/ATTS domain) was first identified as a nuclear protein that binds and activates transcription from the SV40 enhancer. 17 There are four members of the TEAD family named TEAD1 to TEAD4. These proteins share in common the TEA DNA binding domain. With the help of coactivators, such as YAP1 (Yes-associated protein), TAZ (transcriptional coactivator with PDZ-binding motif), and vgll proteins, the TEAD family plays a crucial role in some physiological processes as well as in human cancers. 18 Increased TEAD expression was seen in breast cancers, 19 renal cell carcinoma, 20 medulloblastoma, 21 Kaposi sarcoma, 22 and gastric cancer. 23 In this study, TEAD1 was identified to be a direct target of miR-590-3p by bioinformatics analyses and dual-luciferase reporter assay. TEAD1 harbors a conserved miR-590-3p site in its 3′UTR. Reduced expression of TEAD1 in HCC cells was observed after transfection with pre-miR-590-3p. Furthermore, re-expression of TEAD1 partially rescued the miR-590-3p-induced inhibition of cell proliferation, suggesting that TEAD1 is a direct and functional target of miR-590-3p in HCC cells. Finally, it has been reported that TEAD1 showed a high expression level in prostate cancers and correlated with poor clinical outcome in prostate cancer patients. 24 Overexpression of TEAD1 promoted colorectal cancer cells proliferation, 25 while TEAD1 knockdown resulted in decrease of proliferation and invasion of gastric cancer cells. 23 In our current study, in HCC tissues, TEAD1 also showed a high expression level, which was correlated with poor prognosis in HCC patients.
In conclusion, using human HCC tissues, we have shown that downregulation of tumor suppressor miR-590-3p with concomitant upregulation of their target TEAD1 is an impotent molecular event that could play a causal role in hepatocarcinogenesis. Therefore, rescuing miR-590-3p level may be a novel strategy for the treatment of HCC.
Footnotes
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) received no financial support for the research, authorship, and/or publication of this article.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.