
Abstract
RBM38, a member of RNA recognition motif family of RNA-binding proteins, can regulate the expression of diverse targets by influencing their messenger RNA stability and play a vital role in cancer development. RBM38 may act as an oncogene or suppressor gene in several human tumors. However, its role in human renal cell carcinoma remains unclear. In this study, we found that the expression of RBM38 was lower in renal cell carcinoma tissues and cell lines. Moreover, overexpression of RBM38 could reduce, whereas knockdown of RBM38 could accelerate renal cell carcinoma cell lines growth rate and number of colonies formation of renal cell carcinoma cell lines. Furthermore, RBM38 inhibited renal cell carcinoma cell lines migration and invasion through epithelial–mesenchymal transition suppression by up-regulating E-cadherin and down-regulating β-catenin and vimentin. For in vivo assays, we found that the RBM38-positive group CAKI-1-RBM38 formed smaller tumors in nude mice compared with the control group. Kaplan–Meier analysis showed that renal cell carcinoma patients with lower expression of RBM38 had a significantly shorter survival time than those with higher expression of RBM38 (p = 0.028). All these suggested that RBM38 acts as a tumor suppressor in renal cell carcinoma, which has the potential value for the prediction of renal cell carcinoma prognosis.
Introduction
Renal cell carcinoma (RCC) is the most common type of kidney neoplasms in adult patients, which accounts for approximately 3% of all adult malignancies, and about 30% of the patients who have metastasis on first presentation. 1 Although surgical resection is the current standard of treatment for localized RCC, about 50% of patients existed with the risk of postoperative recurrence and metastasis, which has an extremely poor prognosis. 2 Study on the prognostic factors of RCC and its available clinical molecular biomarkers as well as target therapy methods is vital to improve the survival rate, and contributes to the clinical practice. 3 Accordingly, establishing multiple biomarkers of RCC models in clinical practice and searching for the related sensitive tumor markers is of great significance to early detection of RCC and its prognosis after operation. At present, several molecular biomarkers, which were confirmed relative with the prognosis of RCC, have been previously reported, such as interleukin 33 (IL33), 4 cullin1, 5 thioredoxin domain–containing protein 5 (TXNDC5), 6 and so on. Moreover, a better identification and characterization of the mechanisms in underlying RCC pathogenesis may allow for developing new drugs to cure RCC. 7
RNA-binding proteins (RBPs), which contain one or more RNA-binding motifs, such as the RNA recognition motif (RRM), the human heterogeneous nuclear ribonucleoprotein (hnRNP) K homology motif, the RGG box, and the double-stranded RNA-binding domain (dsRBD) motif,8,9 are thought to be important in regulating gene post-transcription and all aspects of RNA metabolism and function. These regulations are critical for gene regulation in mammalian cells,9,10 and unexpected mutations or the aberrant production of RBPs can cause cancer progression. 11
The RNA-binding motif protein 38 (RBM38, also known as RNPC1) is a member of the RRM-containing RBP family, which is known to regulate messenger RNA (mRNA) translation of p53 and mRNA stability of p21, 12 Hu antigen R (HuR), 13 p63, 14 p73, 15 mouse double minute 2 homolog (MDM2), 16 macrophage inhibitory cytokine 1 (MIC-1), 17 estrogen receptors (ERs), 18 progesterone receptors (PRs), 19 and E2F1 transcripts. 20 RBM38 is frequently overexpressed in cancers, including dog lymphoma, 21 esophageal cancer,22,23 prostate,24,25 chronic lymphocytic leukemia, 26 and breast cancer, 18 suggesting that RBM38 may function as an oncogene. But new evidences suggested that RBM38 acts as a tumor suppressor in breast cancer, 27 ovarian cancer, 20 colorectal cancer (CRC), 28 and acute myeloid leukemia. 29 These data raise a possibility that RBM38 have both positive and negative impacts on tumorigenesis by regulating different signaling pathways. Consistently, Zhang et al. 21 confirmed that RBM38 repressed p53 mRNA translation in both wild-type (WT) and mutant cells and displayed two opposed functions on p53-dependent tumorigenesis.30,31 Recently, it has also found that the expression of RBM38 varied in different types of cancers by bioinformatics analysis. 23 However, expression and biologic functions of RBM38 in human RCC remain obscure. Moreover, there were few studies describing the relationship between the RBM38 expression and prognosis in patients who suffered from cancer.
In this study, we detected the expression of RBM38 in normal renal and RCC tissues, and analyzed the relationship of RBM38 expression and prognosis in renal cancer patients with tissue microarray (TMA). The relationship between RBM38 expression and clinicopathological features of RCC patients was also evaluated. We also investigated the effect of RBM38 on cell viability, apoptosis, migration, and invasion ability. Both the clinical data and experimental results suggested that RBM38 might act as a tumor suppressor in RCC, which has the potential for the prediction of RCC prognosis.
Materials and methods
Cell lines and culture
Human RCC cell lines CAKI-1 and CAKI-2 were purchased from the Institute of Biochemistry and Cell Biology of the Chinese Academy of Sciences (Shanghai, China). The cells were maintained in M5A medium containing 10% fetal bovine serum (FBS). All cells were cultured in an incubator with humidified 5% CO2 at 37°C.
Lentivirus transfection
Lentiviral constructs of RBM38 overexpression and knockdown were generated as previously described. 18 For RBM38 overexpression, the human RCC cell lines CAKI-1 and CAKI-2 were stably transfected with RBM38 overexpression lentivirus (named as RBM38) and a negative control (named as NC). For RBM38 knockdown, CAKI-1 and CAKI-2 cells were stably transfected with an NC (named as SCR) and RBM38 knockdown lentivirus (named as sh1, sh2, and sh3). For RBM38 knockdown, one construct (sh3), named as shRBM38, with ⩾85% knockdown efficiency was used for further studies. Briefly, CAKI-1 or CAKI-2 cells were seeded in six-well plates and cultured to 30%–50% confluence before being used for transfection with the retroviruses. In addition, polybrene (1 µg/mL) was added with the retroviruses to enhance the target cells infection efficiency.
Quantitative real-time polymerase chain reaction
Total RNA was extracted from cultured cells with TRIzol reagent (TaKaRa, Dalian, China) following the manufacturer’s protocols. For quantitative real-time polymerase chain reaction (qRT-PCR), RNA was reverse transcribed to complementary DNA (cDNA) using a Reverse Transcription Kit (TaKaRa). Real-time polymerase chain reaction (RT-PCR) analyses were performed using a standard protocol from fluorescent SYBR Green I methodology (TaKaRa) on an ABI StepOnePlus instrument (Applied Biosystems, Carlsbad, CA, USA). Here, β-actin was used as the internal control. Each experiment was performed in triplicates and repeated three times. These data were analyzed by the 2−ΔΔCT method. The primers sequences were as follows: β-actin: 5′-GCTGTGCTATCCCTGTACGC-3′ (forward) and 5′-TGCCTCAGGGCAGCGGAACC-3′ (reverse); RBM38: 5′-ACGCCTCGCTCAGGAAGTA-3′ (forward) and 5′-GTCTTTGCAAGCCCTCTCAG-3′ (reverse).
Cell viability assays
Cell proliferation assay was performed using cell counting kit-8 (CCK-8; Dojindo, Japan) according to the manufacturer’s instructions or protocols. For the cell viability assay, cells were seeded in a 96-well plate at a density of 3000 cells per well. The CCK-8 method was utilized to determine cell viability at 2, 24, 48, 72, 96, and 120 h after seeding cells. The absorbance was measured at 450 nm using a microplate reader (BioTek, Winooski, VT, USA).
Colony formation assays
For the colony formation assay, 400 cells were plated into six-well plate and maintained in media containing 10% FBS to allow colony formation for 15 days. The colonies were imaged and counted after fixing with methanol and staining with 0.1% crystal violet.
Wound healing assay
Cells were seeded into six-well plates and allowed to grow until 95% confluence. Then, the cell layer was gently scratched through the central axis using a sterile plastic tip and loose cells were washed away. Quantification of cell motility by measuring the distance between the invading fronts of cells in three random selected microscopic fields (100×) for each condition and time point (0 and 24 h).
Cell migration and invasion assay
Cells (3.0 × 104) were suspended by 200 µL serum-free medium and seeded on the top chamber of the transparent 8-µm pore polycarbonate transwell (Millicell, Durham, USA). Culture medium containing 10% FBS was added to the bottom chamber. Cells were incubated for 48 h or 72 h at 37°C, and the cells that did not migrate through the pores in the membrane were removed by scraping the membrane with a cotton swab. Cells which passed through the filter were fixed and stained using 0.1% crystal violet. Images of five random fields (100×) were captured from each membrane. Finally, we counted the number of migratory or invasive cells and calculated the average values of them. Each experiment was conducted in triplicate.
Cytometry
Apoptosis was assayed as reported. 32 Briefly, the cells were harvested, washed with phosphate-buffered saline (PBS) for two times, fixed by 95% ethyl alcohol for at least one night, then washed twice with cold PBS, and stained with propidium iodide (PI; Key Biotech, Nanjing, China). The cell cycle analysis was performed using a BD FACS Canto II (BD Biosciences, NJ, USA) according to the manufacturer’s protocol. The data were analyzed by FlowJo version 7.2.2 (Ashland, OR, USA).
Western blotting analysis
Cells were washed twice with cold PBS and then scraped off in 300 µL of radio immunoprecipitation assay lysis buffer (Beyotime, China) and incubated on ice for 30 min. Then, cell lysates were centrifuged at 4°C, 12,000g for 15 min, and the supernatants were stored at −80°C. Protein concentrations were measured by using the bicinchoninic acid (BCA) protein assay kit (Beyotime). The prepared proteins were added equal parts of 2 × loading buffer, then boiled for 5 min, and separated by 12% sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE), and then transferred to polyvinylidene fluoride (PVDF) membranes (Millipore, USA), after that, blocked with 5% skim milk in TBST buffer (10 mM Tris–HCl, 50 mM NaCl, and 0.1% Tween-20) for 1–2 h at room temperature. The membranes were immunoblotted with the primary antibodies for one night at 4°C: anti-rabbit vimentin (Cell Signaling Technology, MA, USA), β-catenin (Cell Signaling Technology), P21 (Santa Cruz, CA, USA), RBM38 (Santa Cruz), anti-mouse E-cadherin (Cell Signaling Technology), or β-actin (Santa Cruz). β-actin was used as the loading control. Horseradish peroxidase–conjugated goat anti-mouse-IgG and anti-rabbit-IgG (Bioworld, MN, USA) were used as secondary antibodies. The antibodies E-cadherin, vimentin, β-catenin, P53, P21, and RBM38 were used at dilution 1:2000, and the secondary antibodies were used at dilution 1:5000. The intensity of the bands was determined using densitometric analysis. All experiments were performed three times.
Tumorigenesis in nude mice
Mouse studies were conducted according to the Guide for the Care and Use of Laboratory Animals and approved by the Animal Care and Use Committee of Nanjing Medical University (Nanjing, China). BALB/C female nude mice (4–6 weeks old, 18–22 g in body weight) were randomly divided into two groups (each containing five mice). RBM38-positive group CAKI-1-RBM38 and matched control group CAKI-1-NC (1 × 106 cells) in 150 µL PBS of a suspension mixture with Matrigel (BD Biosciences) were subcutaneously injected into the mice. After 6 weeks, mice were sacrificed and the final tumor was weighted.
Clinical sample collection
Human kidney specimens were obtained from 140 patients who underwent radical nephrectomy for clear cell RCC at the First Affiliated Hospital of Nanjing Medical University (Nanjing, China) between 2008 and 2011. Fresh adjacent tissue to carcinoma tissues and the carcinoma specimen of RCC were collected from 25 patients and confirmed by histopathology. The collection and use of the samples were reviewed and approved by the Institutional Ethics Committee of the First Affiliated Hospital of Nanjing Medical University (Nanjing, China), and expedited pathological diagnosis and staging of these specimens were performed prior to sampling and preserving them for research. Histological diagnosis was established according to the guidelines of the World Health Organization (2003). 33 Clinical follow-up data was retrieved for all patients, but the patients in our follow-up studies most have undergone partial nephrectomies, who were hardly followed by tumor necrosis, vascular invasion, capsular invasion, which was not available for statistical analysis.
Immunohistochemical staining
Serial formalin-fixed and paraffin-embedded tissues were sectioned at a 4-µm thickness, deparaffinized, and rehydrated in gradients of high-percentage ethanol to distilled water. For quenching endogenous peroxidase activity, sections were immersed in 3% hydrogen peroxide for 15 min at room temperature. Antigen retrieval involved boiling in 10 mM sodium citrate buffer (pH 6.0) for 3 min in a pressure cooker, followed by cooling to room temperature. Sections were then incubated with the RBM38 antibody (diluted 1:350; Life Span Biosciences, USA) at 4°C overnight, After washing in TBST, slides were incubated with the anti-RBM38 rabbit IgG antibody (Vector Laboratories, Burlingame, CA, USA) for 30 min at room temperature, followed by 3,3′-diaminobenzidine tetrahydrochloride (DAB) staining. Sections were lightly counterstained with hematoxylin.
Evaluation of immunostaining
Two pathologists examined the evaluation of RBM38 independently and blindly. Positive RBM38 staining as cytoplasmic with or without nuclear staining was defined, and it was graded according to both the intensity and percentage of cells with positive staining. The percentage of cells positively stained was scored 1–4: 1 (0%–10%), 2 (11%–50%), 3 (51%–80%), and 4 (81%–100%). The intensity of RBM38 staining was scored 0–3 (0 = negative, 1 = weak, 2 = moderate, and 3 = strong). Immunoreactive score (IRS) was used to evaluate the level of RBM38 staining, which is calculated by multiplying the scores of staining intensity and the percentage of positive cells. According to the IRS, RBM38 staining pattern was defined as negative (IRS: 0–3) and positive (IRS: 4–12).
Statistical analysis
All data were expressed as the mean ± standard deviation (SD), and analyses were performed using SPSS version 17.0 software. Statistical significance was tested by a Student t-test, or a chi-square test as appropriate. Patient survival was evaluated using the Kaplan–Meier method and compared using the log-rank test. Univariate and multivariate Cox regression analyses were performed to analyze the survival data. Values of p < 0.05 were considered statistically significant.
Results
RBM38 was lower expressed in human renal cancer tissues and cells
qRT-PCR and western blot were used to examine RBM38 expression in human renal cancer tissues and cells. The results demonstrated that RBM38 mRNA was significantly down-regulated in RCC tissues compared with adjacent renal tissues (Figure 1(a) and (b), p < 0.05). It was also found that RBM38 protein down-regulated in RCC tissues compared with adjacent renal tissues in most cases (9/11; Figure 1(c)). RBM38 was found lower expressed in renal cancer cells lines compared with normal renal epithelial HK-2 cells both in mRNA and protein levels (Figure 1(d) and (e)). Among renal cancer cells lines, HK-2 cells expressed relatively higher levels of RBM38, while lower expression or barely detectable levels were found in CAKI-1, CAKI-2, 786-O, 769-P, and ACHN cells.

RBM38 expression in RCC cell lines and tissues: (a) and (b) Relative expression of RBM38 mRNA in RCC tissues compared with corresponding adjacent renal tissues. Adjacent renal tissues had higher expression of RBM38, where the RCC tissues showed the lower level of expression (*p < 0.05). (c) RBM38 protein expression in eight pairs of RCC tissues and corresponding adjacent renal tissues. The RBM38/actin ratio was markedly lower in tumors compared with adjacent normal tissues in nine among 11 cases. “T” means RCC tissues; “A” means adjacent renal tissues. (d) Relative expression of RBM38 mRNA in five RCC cell lines, compared with renal tubular epithelial cell line HK-2. The renal tubular epithelial cell line HK-2 showed higher expression of RBM38 mRNA than other cell lines (*p < 0.05). (e) RBM38 protein expression in five renal cell lines, compared with renal tubular epithelial cell line HK-2. RBM38 protein was found expressed lower in renal cancer cells lines compared with normal renal epithelial HK-2 cells.
Expression of RBM38 in transfected RCC cell lines
To further address the functions of RBM38 in renal cancer cells, we infected CAKI-1 cells and CAKI-2 cells and selected stably infected cells. The RBM38 overexpression RCC cell lines were named as CAKI-1-RBM38 and CAKI-2-RBM38, while the matched control cell lines were named as CAKI-1-NC and CAKI-2-NC, respectively. The RBM38 knockdown RCC cell lines were named as CAKI-1-shRBM38 or CAKI-2-shRBM38, while the matched control cell lines were named as CAKI-1-SCR or CAKI-2-SCR, respectively. We confirmed the expression levels of RBM38 by western blot (Figure 2(a) and (b)) and qRT-PCR (Figure 2(c) and (d), both p < 0.05). As shown in Figure 2, RBM38 expression was significantly down-regulated or up-regulated in CAKI-1 and CAKI-2 cells transfected with the shRBM38 or RBM38, respectively, compared with RBM38-SCR or RBM38-NC transfected cells, respectively.

Expression of RBM38 in transfected RCC cell lines. (a) and (b) Western blot was used to verify the efficiency of RBM38 overexpression, knockdown, and p21 protein expression level in RBM38-transfected CAKI-1 and CAKI-2 cells. The expression of RBM38 was increased in the RBM38 overexpression RCC cell lines, while decreased in the RBM38 knockdown RCC cell lines, and RBM38 positively regulated p21. (c) and (d) qRT-PCR was used to verify the efficiency of RBM38 overexpression and knockdown in CAKI-1 and CAKI-2 cell lines (*p < 0.05).
RBM38 inhibited the proliferation and growth in human RCC cell lines
As shown in Figure 3(a) and (b), RBM38 knockdown RCC cell lines led to significantly increasing cell proliferation (p < 0.05), while RBM38 overexpression RCC cell lines led to significantly decreasing cell proliferation (Figure 3(c) and (d), p < 0.05). The ability of CAKI-1 or CAKI-2 cell lines to form colonies was better when RBM38 was knockdown (Figure 3(e) and (f), p < 0.05). The ability of CAKI-1 or CAKI-2 cell lines to form colonies was poorer when RBM38 was overexpressed (Figure 3(g) and (h), p < 0.05). The growth of the stable transfected human RCC cell lines over 5 days was determined using CCK-8 assay. As shown in Figure 4(a)–(c), CAKI-1-shRBM38 cells showed a delayed G1 phase compared with CAKI-1-SCR cells (68.47 ± 1.144 vs 80.397 ± 1.105, p < 0.05), while, as shown in Figure 4(d)–(f), CAKI-1-RBM38 cells showed an expedited G1 phase compared with CAKI-1-NC cells (76.33 ± 1.276 vs 67.25 ± 1.138, p < 0.05). Also, as shown in Figure 4(g)–(i), CAKI-2-shRBM38 cells showed a delayed G1 phase compared with CAKI-2-SCR cells (72.80 ± 1.516 vs 81.49 ± 1.591, p < 0.05), while, as shown in Figure 4(j)–(l), CAKI-2-RBM38 cells showed an expedited G1 phase compared with CAKI-2-NC cells (80.41 ± 1.537 vs 72.04 ± 1.076, p < 0.05). These results demonstrated that the RBM38 knockdown RCC cell lines significantly decreased G0/G1 phase and increased the S and G2/M phases in cell cycle, while RBM38 overexpression RCC cell lines significantly increased the G0/G1 phase and decreased the S and G2/M phases in cell cycle. It was found that human RCC cells were arrested primarily in the G1 phase by RBM38 in cell cycle.
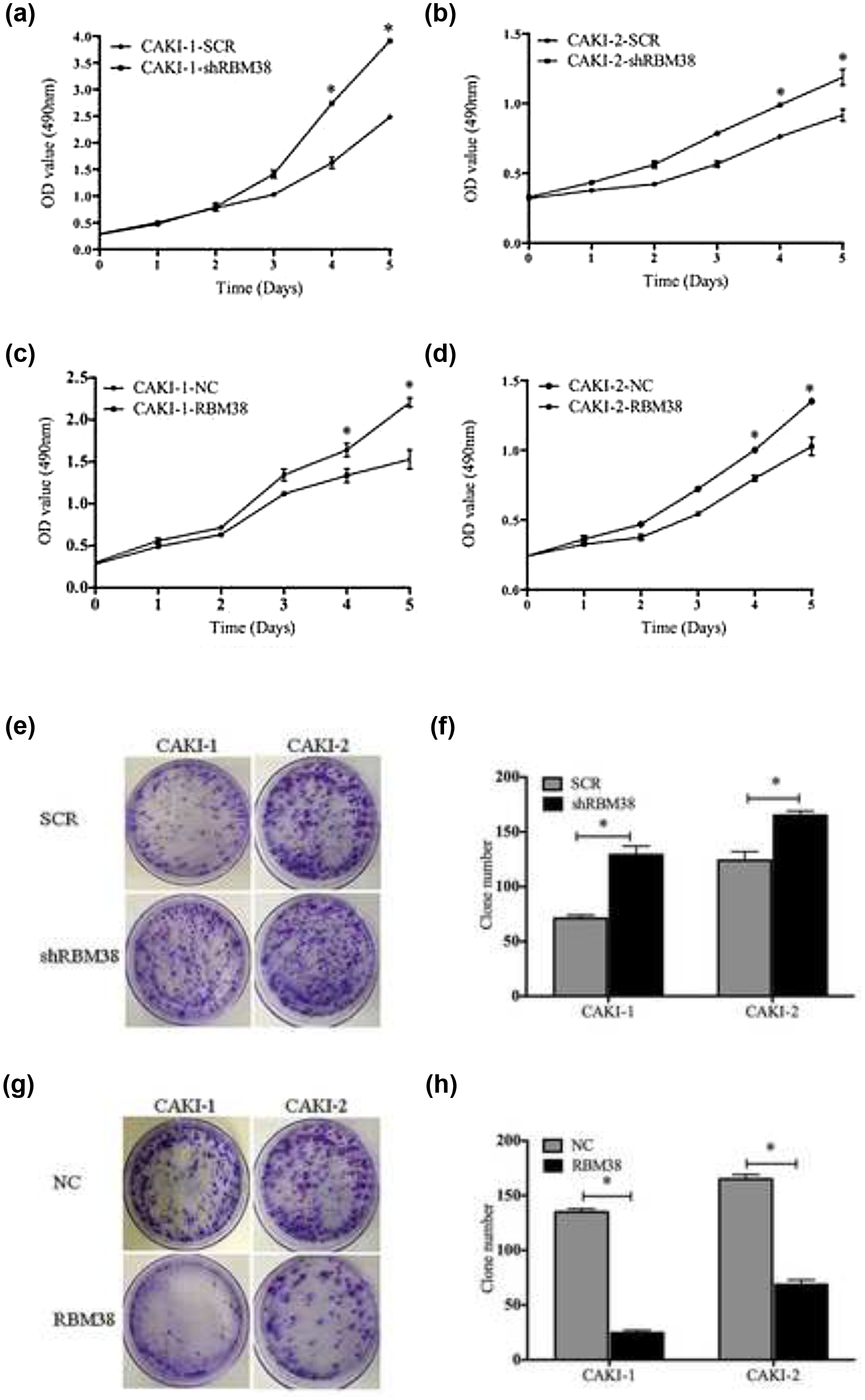
RBM38 inhibits the proliferation and growth of RCC cell lines. (a) and (b) The growth of RCC cell line over 5 days was measured by CCK-8 assays. The cell proliferation rate of (a) CAKI-1-shRBM38 and (b) CAKI-2-shRBM38 was significantly increased compared with control cells, respectively (*p < 0.05). (c) and (d) The cell proliferation rate of (c) CAKI-1-RBM38 and (d) CAKI-2-RBM38 was significantly decreased compared with control cells, respectively (*p < 0.05). (e) and (f) The growth of RCC cell lines over 15 days was measured using colony formation assays. Anchorage-dependent growth assays were shown at the bottom. The number of CAKI-1-shRBM38 or CAKI-2-shRBM38 was significantly increased compared with control cells, respectively. (e) Representative photographs and (f) quantification are shown (*p < 0.05). (g) and (h): The number of CAKI-1-RBM38 or CAKI-2-RBM38 was significantly decreased compared with control cells, respectively. Anchorage-dependent growth assays were shown at the bottom. (g) Representative photographs and (h) quantification are shown (*p < 0.05).

Cell cycle progression was measured by flow cytometry. The progression of (a)–(c) CAKI-1-shRBM38 cells and (g)–(i) CAKI-2-shRBM38 cells was decreased in the G1 phase compared with control cells (SCR), respectively. Representative photographs and quantification are shown. The progression of (d)–(f) CAKI-1-RBM38 cells and (j)–(l) CAKI-2-RBM38 cells was increased in the G1 phase compared with control cells (NC), respectively. Representative photographs and quantification are shown (*p < 0.05).
RBM38 suppressed the migration and invasion of human RCC cell lines
As shown in Figure 5(c) and (d), determined by their migration in the wound gap after 24 h, migrated distance of CAKI-2-shRBM38 increased by 92 µm (p < 0.05), while CAKI-2-RBM38 decreased by 149 µm (Figure 5(g) and (h), p < 0.05) compared with the control cells, respectively. Moreover, we obtained the similar results in CAKI-1 cells (Figure 5(a), (b), (e), and (f)). We conducted three-dimensional cell migration assay using transwell chambers and invasion assay with Matrigel-precoated transwell chambers. We found that CAKI-2-shRBM38 exhibited significantly increasing the ability of migration and invasion (Figure 5(k) and (l), p < 0.05), while CAKI-2-RBM38 exhibited significantly decreasing the ability of migration and invasion (Figure 5(o) and (p), p < 0.05); the similar results in CAKI-1 cells were obtained (Figure 5(i), (j), (m), and (n)).
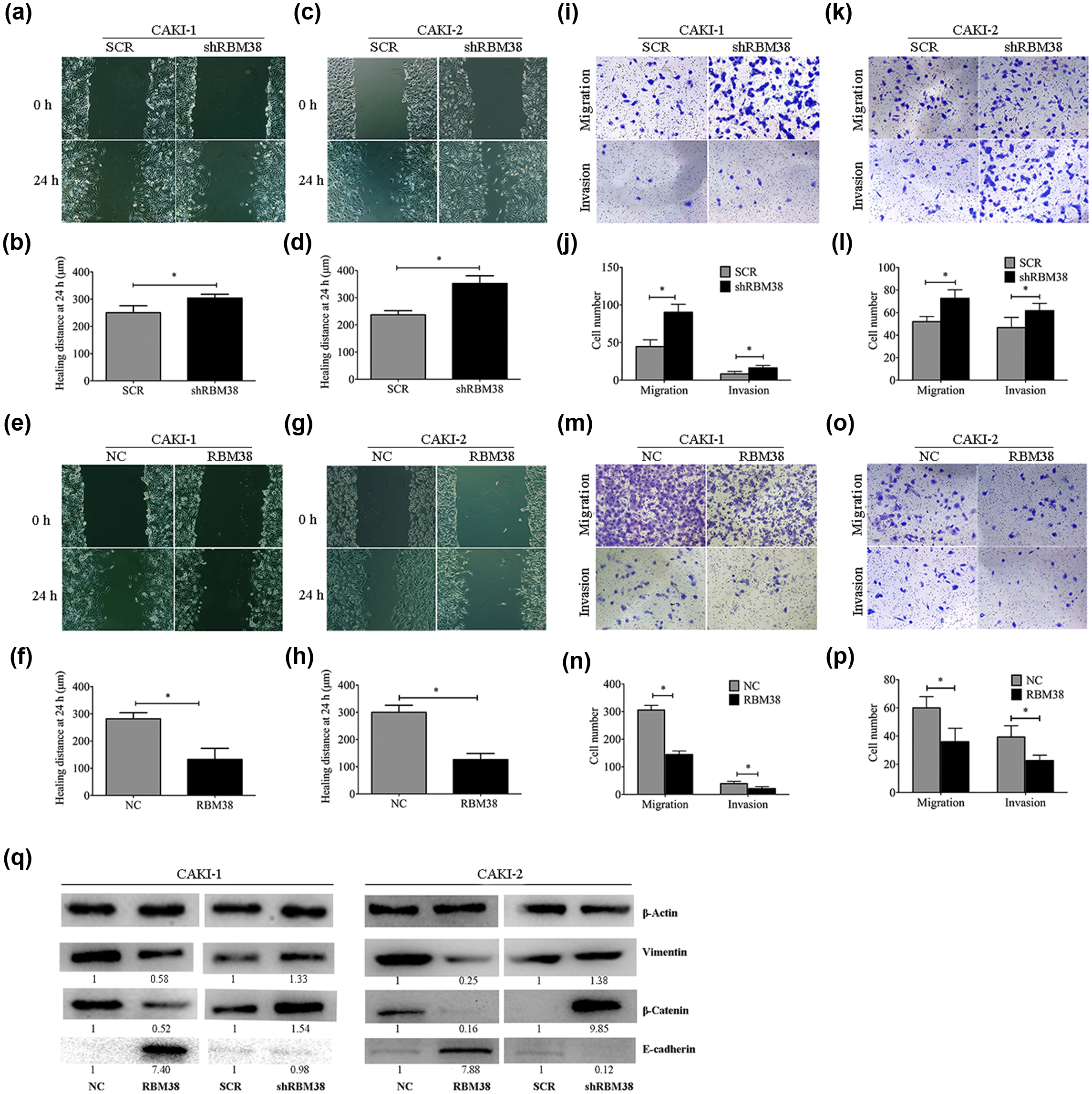
RBM38 significantly decreased the migration and invasion of RCC cell lines. (a) and (b) Wound healing assay of RBM38 knockdown for CAKI-1 cells. Images of wound repair were taken at 0 and 24 h after wound. The distance of wound closure is shown by area at 24 h. (a) Representative photographs and (b) quantification are shown (original magnification: ×100). (c) and (d) Wound healing assay of RBM38 knockdown for CAKI-2 cells. (e) and (f) Wound healing assay of RBM38 overexpression for CAKI-1 cells. Images of wound repair were taken at 0 and 24 h after wound. The distance of wound closure is shown by area at 24 h. (e) Representative photographs and (f) quantification are shown (original magnification: ×100). (g) and (h) Wound healing assay of RBM38 overexpression for CAKI-2 cells. (i) and (j) Transwell migration assay and Matrigel invasion assay of RBM38 knockdown for CAKI-1 cells. (i) Representative photographs and (j) quantification are shown (original magnification: ×100). (k) and (l) Transwell migration assay and Matrigel invasion assay of RBM38 knockdown for CAKI-2 cells. (m) and (n) Transwell migration assay and Matrigel invasion assay of RBM38 overexpression for CAKI-1 cells. (m) Representative photographs and (n) quantification are shown (original magnification: ×100). (o) and (p) Transwell migration assay and Matrigel invasion assay of RBM38 overexpression for CAKI-2 cells (*p < 0.05). (q) Western bolt analysis of vimentin, β-catenin, and E-cadherin protein expression level in RBM38-transfected CAKI-1 and CAKI-2 cells and respective control cells. RBM38 negatively regulated vimentin and β-catenin, while positively regulated E-cadherin in CAKI-1 and CAKI-2 cells. The fold change of RBM38 is shown below each lane.
We observed that the RBM38 knockdown RCC cell lines led to a spindle-shaped fibroblastic morphology. In addition, RBM38 overexpression RCC cell lines lost their fibroblast-like morphology, which was accompanied by a cobblestone-like epithelial morphology (data not shown). This morphological change might suggest the phenotypic change of epithelial–mesenchymal transition (EMT). As shown in Figure 5(q), the expression of E-cadherin was increased in the RBM38-positive group, while decreased in the RBM38-negative group. The expressions of β-catenin and vimentin were increased in the RBM38-silenced cell lines, while decreased in the RBM38-overexpressed cells.
RBM38 suppressed tumorigenesis in nude mice
To evaluate in vivo tumor-suppressive functions of RBM38, tumorigenicity of CAKI-1 cells expressing RBM38 was evaluated in nude mice. CAKI-1-RBM38 and CAKI-1-NC were injected subcutaneously with Matrigel into athymic nude mice. The CAKI-1-NC group formed tumors after 3 weeks, while tumors derived from the CAKI-1-RBM38 group were found after 4 weeks. After 6 weeks, mice were sacrificed and checked for final tumor weight (Figure 6(a)). The CAKI-1-RBM38 group formed smaller tumor volume and weight compared with the CAKI-1-NC group (Figure 6(b), p < 0.05).

Overexpression of RBM38 suppressed tumor formation. (a) A representative picture of tumor growth in nude mice subcutaneously inoculated with RBM38-overexpressed cell lines CAKI-1-RBM38 (marked as RBM38 in the figure) and matched control cell lines CAKI-1-NC (marked as NC in the figure). The CAKI-1-RBM38 group showed a retarded tumor growth compared with the CAKI-1-NC group. (b) The mean tumor weights in nude mice subcutaneously inoculated with CAKI-1-NC and CAKI-1-RBM38 (*p < 0.05).
Relationship between RBM38 expression and RCC patients’ clinicopathological features
Patients with RCC were classified into RBM38-positive and RBM38-negative groups based on the relative RBM38 expression (Figure 7). Clinicopathological factors were also compared between the two groups (Table 1); compared with the RBM38-negative group, it was demonstrated that the RBM38-positive group was younger (p = 0.004). The RBM38-positive group was more tend to be no diabetes mellitus patients (p = 0.002) compared with the RBM38-negative group. However, RBM38 expression was not associated with other parameters such as gender (p = 0.939), tumor size (p = 0.466), histological grade (p = 0.462), and tumor stage (p = 0.793).

Immunohistochemical expression of RBM38 in RCC patients by tissue microarray. The percentage of cells positively stained was scored 1–4: 1 (0%–10%), 2 (11%–50%), 3 (51%–80%), and 4 (81%–100%). The intensity of RBM38 staining was scored 0–3 (0 = negative, 1 = weak, 2 = moderate, and 3 = strong). IRS was used to evaluate the level of RBM38 staining, which is calculated by multiplying the scores of staining intensity and the percentage of positive cells. According to the IRS, RBM38 staining pattern was defined as negative (IRS: 0–4) and positive (IRS: 5–12). (a) RBM38 expression is completely negative. (b) RBM38 expression is weak. (c) RBM38 expression is moderate. (d) RBM38 expression is strong. (a) and (b) were defined as the RBM38-negative group; (c) and (d) were defined as the RBM38-positive group.
Correlation between RBM38 expression and clinicopathological characteristics of RCC.

RBM38: RNA-binding motif protein 38; RCC: renal cell carcinoma; DM: diabetes mellitus.
The RBM38 expression was correlated with age (p = 0.004) and DM (p = 0.002). The p value is calculated by chi-square test or Fisher‘s exact probability method.
RBM38 expression has correlation with the overall survival of patients
We further examined whether the RBM38 expression level correlated with the prognosis of RCC patients after treatment. Kaplan–Meier analysis showed that the RBM38-negative group had a significantly shorter survival time than the RBM38-positive group (log-rank test, p < 0.05; Figure 8); median follow-up was 66.5 months. As shown in Table 2, RBM38 expression level, histological grade, and tumor stage were significantly correlated with the survival rate in patients with RCC (p < 0.05). A multivariate analysis showed that relative expression of RBM38, histological grade, and tumor stage were independent prognostic markers for survival of RCC patients. Taken together, these data indicated that RBM38 expression was a powerful prognostic factor for RCC patients’ overall survival.

Kaplan–Meier survival curves of RCC patients based on RBM38 expression.
Univariate and multivariate analyses for recurrence-free survival (Cox proportional hazards regression model).

HR: hazards ratio; CI: confidence interval; RBM38: RNA-binding motif protein 38; RCC: renal cell carcinoma.
The relative expression of RBM38 (p = 0.023), histological grade (p = 0.004), and tumor stage (p = 0.014) were independent prognostic markers for surviving of RCC patients.
Bold value signifies that the p value is calculated by chi-square test or Fisher‘s exact probability method.
Kaplan–Meier analysis showed that the RBM38-negative group had a significantly shorter survival time than the RBM38-positive group (log-rank test, p < 0.05).
Discussion
RCC is a malignant tumor originated from renal tubular epithelial cells, which occupies the second place of incidence rates among the urinary system tumors. 7 RBM38 is an RBP that contains one RRM domain. It can bind and stabilize the mRNA of the p21, thereby inducing cell cycle arrest in the G1 phase, 13 which was proved to play pivotal roles in regulating cell proliferation, cell cycle arrest, and cell myogenic differentiation. 12 To our knowledge, this is the first study on investigating the role of RBM38 in RCC. Although RBM38 acts as oncogene or tumor suppressor in different cancer, there are few studies describing the relationship between the RBM38 expression and prognosis in patients who suffered from cancer. In this study, we first found that RBM38 expression showed a predictive value for the prognosis in RCC patients after surgery.
For in vitro assays, we found that the expression of RBM38 was lower in RCC cell lines (CAKI-1, CAKI-2, 786-O, 769-P, and ACHN) than in normal cell line HK-2. Moreover, overexpression of RBM38 could reduce, whereas knockdown of RBM38 could accelerate RCC cell lines growth rate and number of colonies formation of RCC cell lines. It implied a suppressive function of RBM38 in RCC cell lines. Cell proliferation is controlled by cell cycle progression, which is regulated by cyclins, cyclin-dependent kinases (Cdks), and Cdk inhibitors.34,35 It was found that RBM38 showed inhibitory effect on the growth of RCC cells via a delay in cell cycle progression by cell cycle arrest at the G1 phase. Consistent with this, RBM38 was reported to cause cell cycle arrest in G1 in colon cancer cell line RKO, osteosarcoma cell line U2OS, and breast cancers cell lines MCF-7 and MDA-MB-231. p21 belongs to the Cdk inhibitors involved in the cell cycle control.34–36 RBM38 was reported to regulate p21 stability by binding to adenylate-uridylate-rich (AU-rich) elements in its 3′-untranslated regions (UTRs). 12 p21 was changed concomitantly with RBM38 in RCC cell lines, suggesting that RBM38 in part induces cell cycle arrest in G1 via binding to and at least partly stabilizing p21 transcription in RCC cell lines. For in vivo assays, we found that the RBM38-positive group CAKI-1-RBM38 formed smaller tumor in nude mice compared with the control group. These results were well consisted with experimental data both in vitro and in vivo, suggesting that RBM38 is a tumor suppressor in RCC.
RCCs are not only proliferative but also readily metastatic to other places. Cell migration and invasion are the key steps of RCC metastasis. Our data revealed that RBM38 knockdown could increase RCC cell lines migration and invasion. We observed that RBM38 knockdown led to the phenotypic change of EMT in RCC cell lines. EMT is a complex molecular program that regulates changes in cell morphology and function during embryogenesis and tissue development, and also contributes to tumor progression and metastasis, which leads to the loss of epithelial features and to the acquisition of a mesenchymal phenotype and plays a crucial role in RCC development and progression.37,38 Moreover, RBM38 regulated E-cadherin expression positively, while regulated vimentin and β-catenin expression negatively. Our data were consistent with the characteristics of EMT protein changes, 39 suggesting that up-regulation of RBM38 suppresses EMT phenomenon of RCC cells in vitro. Taken together, we speculated that RBM38 inhibited RCC cell lines migration and invasion through EMT suppression, which may occur not only by up-regulating E-cadherin but also by down-regulating mesenchymal genes, such as β-catenin. 40 However, additional studies are required to fully understand the detailed function of RBM38 in EMT.
By searching the database, Feldstein et al. 20 found that higher expression of RBM38 exhibited significantly increased survival in ovarian cancer, breast cancer, and glioblastoma. However, Ding et al. 23 found that the relationship between expression of RBM38 and prognosis varied in different cancer tissues by bioinformatics analysis. In RCC clinical samples, we found that RBM38 was significantly down-regulated in RCC samples compared with adjacent non-tumor samples. Moreover, RBM38 expression was significantly correlated with depth of invasion, ability of cell proliferation, and tumor size. Kaplan–Meier analysis showed that the relative expression of RBM38 was an independent prognostic marker for survival of RCC patients.
In conclusion, RBM38 can significantly decrease the RCC cells proliferation and metastasis capability. Its expression was lower in RCC tumor tissues and cell lines, acting as a tumor repressor in RCC. Moreover, the higher RBM38 expression was correlated with a higher survival rate of RCC, suggesting that RBM38 was an independent prognostic marker for survival of RCC patients.
Footnotes
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Ethical approval
All experiments involving human subjects were performed in accordance with relevant guidelines and regulations of the Institutional Ethics Committee of the First Affiliated Hospital of Nanjing Medical University, and all examinations were performed after obtained informed consents. Mouse studies were conducted according to the Guide for the Care and Use of Laboratory Animals and approved by the Animal Care and Use Committee of Nanjing Medical University.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This study was sponsored by Chinese National Natural Science Foundation (81001329), the Six Talents Peak projects of Jiangsu Province (to H.Y., WSW-025), and a project founded by the Priority Academic Program Development of Jiangsu Higher Education Institutions (PAPD).