
Abstract
Wnt pathway has been implicated in the process of human carcinogenesis. Axis inhibition protein2 (Axin2), a major scaffold protein is an antagonist of Wnt pathway and is potent to act as a tumor suppressor gene in various human cancers. Therefore, the seven polymorphic sites of Axin2 gene were analyzed, in relation to lung cancer susceptibility in North Indians. A total of 608 subjects were genotyped using PCR-RFLP technique for each polymorphic site including 303 cases and 305 controls. Further association analysis was carried out using logistic regression approach to obtain adjusted odds ratio and statistical significance. MDR and CART analysis were applied to evaluate high order interactions between the SNP’s. Three out of seven studied polymorphic sites showed a strong protective effect in subjects having mutant genotype for Axin2 148 C >T and heterozygous genotype for 1365 G > A and 1712 + 19 G > T towards lung cancer risk. The other important finding was the significant association of Axin2 148 C >T in SQCC patients having variant (TT) genotype. Axin2 1712 + 19 G >T showed a decreased risk for all the histological subtypes in patients with heterozygous (GT) genotype. MDR analysis predicted a best interaction model (Axin2 148, Axin2 2062 and Axin2 1712 +19) with maximum CVC (10/10) and minimum prediction error (0.38) along with significant permutation p-value. CART analysis gave a wide spectrum of interactive combinations which exhibited a major contribution in modulating lung cancer susceptibility. Axin2 148 and Axin2 1712 + 19 were found to play a major role in modulating lung cancer risk.
Introduction
Wnt signaling is one of the primary pathways which form the basic framework of cellular development and differentiation in organisms. Cellular polarity in embryos and tissue maintenance in adults are some of the major dimensions of this intermeshing pathway. 1 This cascade directs the expression of all the primary genes involved in the processes of cell polarity, cellular adhesion, apoptosis, and tissue differentiation. 2 Canonical Wnt pathway is a β-catenin-mediated signaling network which is switched on upon the binding of the Wnt ligands to the cell surface–frizzled receptors and low-density lipoprotein receptor–related protein 5/6 (LRP5/6) complex. 3 This is the primary interaction which disrupts the destruction complex comprising disheveled (Dsh), adenomatous polyposis coli (APC), and Axin2 proteins which is formed for the ubiquitination of β-catenin.4,5 But, due to Wnt ligand–surface receptor binding, the phosphorylation of β-catenin by glycogen synthase kinase-β (GSK-3β) in this destruction complex is inhibited, and then, consequently, β-catenin is translocated to the nucleus where it binds to the several transcription factors such as T-cell factor/lymphoid enhancer factor (TCF/LEF) which regulate the expression of target genes including matrix metalloproteinases, cox-2, cyclin D1, vascular endothelial growth factor (VEGF), and c-myc. 3
Wnt pathway can be regulated by various proteins including Wnt antagonists such as sFRP and DKK and other proteins, for example, Axin2, which forms the part of the destruction complex that inhibits the translocation of β-catenin into the nucleus. 3 Axin2 is an architectural protein which forms the scaffold of this destruction complex. In a way, Axin2 negatively controls the Wnt signaling pathway, and it has been established as one of the potent tumor suppressor gene. Axin2 is located on chromosome 17q24 in humans and codes for 843 amino acid–long protein. Axin2 gene comprises 10 exons. 6 Dysregulation of Wnt pathway may lead to its constitutive activation which enhances the unregulated division of cells,7,8 and henceforth, this augments the process of carcinogenesis in various tissues such as skin, breast, liver, ovary, and lung. 9 Overexpression of target genes is evident in various tumor tissues. 10 Lung cancer is a multifactor-driven cellular abnormality which arises because of certain heterogeneous and unknown genetic and molecular anomalies, 11 and recently, some reports have established the role of this developmental pathway in the oncogenesis of lung tissue. 12 Several types of cancers have evidently reported the modifications in Axin2 protein. 13 The variations in the Axin2 gene may cause certain modulations in the protein structure, and as it forms the scaffold of the destruction complex, its binding with APC may be affected and the process of addition of phosphate moiety to β-catenin may be altered, which is the primary step for its ubiquitination. 14 Various findings have illustrated the loss of heterozygosity and also certain mutations in various cancers. 15 Axin2 gene has been found to be dysregulated in various tumors such as breast, colorectal, ovarian, and hepatocellular. 16 Elevated levels of β-catenin were certainly attributed to the decreased expression of Axin2 in lung cancer tissue, and in case of non-small-cell lung cancer (NSCLC), accumulation of β-catenin in nucleus is attributed to the epigenetic silencing of Axin2 gene. 17 There are no findings about the specific mutations in Axin2 gene in relation to lung cancer.
Polymorphic sites in the gene sequence do greatly influence the orchestration of the protein structure and function. Several studies have been carried out exploring the effect of certain polymorphic sequence differences on the disease susceptibility. 18 Several genetic factors intricately influence the individual’s predisposition to acquire lung cancer. There have been a few reports where the sequence variation within the Axin2 gene was analyzed to find out its effect on lung cancer risk. 19 One of the commonly studied single-nucleotide polymorphism (SNP) includes rs22040308, which has been assessed in different populations such as Turkish, 20 Japanese, 21 Polish, 22 and Chinese. 23 These studies have been carried out in different carcinomas including ovarian, 22 lung, 20 prostate, 24 and astrocytoma. 25 A few studies have also evaluated several other intronic and exonic polymorphic sites including Axin2 148 C/T, Axin2 1365 G/A, Axin2 432 T/C, Axin2 956 + 16 A/G, Axin2 1712 + 19 G/T, Axin2 1386 C/T, and Axin2 2062 C/T in Turkish lung cancer patients 20 and astrocytoma patients. 25 However, the sample size was very low and further studies are required to establish the role of these genetic variants within Axin2 gene in modulating toward lung cancer susceptibility.
Taking into account the above-mentioned facts, this study was designed to find the association of seven Axin2 SNPs, including 148 C > T, 1365 G > A, 432 T > C, 956+16 A > G, 1712+19 G > T, 1386 C > T, and 2062 C > T, with lung cancer susceptibility in North Indian population. An attempt was made to identify the high-risk subgroups by stratifying the subjects on the basis of histology and smoking parameters. The interaction of these SNPs was analyzed using novel computational algorithms, so as to accomplish a comprehensive appraisal of lung cancer risk. Multifactor dimensionality reduction (MDR) approach and Classification and Regression Tree (CART) analysis yielded some interesting patterns and combinations of these polymorphic sites that contribute evidently in modulating lung cancer risk.
Material and methods
Study population
A total of 608 subjects have been enrolled in this study, comprising 303 cases and 305 controls. The patients have been recruited from Lung Cancer Clinic of Department of Pulmonary Medicine, Postgraduate Institute of Medical Education and Research (PGIMER), Chandigarh. The study was approved by the Institutional Ethics Committee of PGIMER. Subjects were explained about this study and an informed written consent was also obtained from each participant. A volume of 5 mL of peripheral blood was taken from each volunteer, and a detailed questionnaire was filled by the volunteers with the help of a trained personnel in order to obtain all the epidemiological details such as age, sex, smoking history, and region they belong from. The clinical details of the patients were obtained from the medical records which included histology of the tumor, stage of the disease, and other specific details. The selection criteria for cases were only that the patient should not have past history of any other carcinoma; otherwise, there were no age, gender, smoking, histology, TNM, and familial history restrictions. The controls were the subjects who visited the hospital for normal health checkups. Controls were selected on the basis of the fact that they did not have any record of previous tumors; however, their familial history did not pose any restriction. The sampling was carried in such a manner so that there is age (±10 years), gender, and smoking matching among cases and controls. This attempt was done in order to avoid sampling bias. In order to obtain a clear idea about the total smoking exposure, pack-years were obtained using the following equation: ((cigarettes or beedis per day/20) × number of years smoked).
DNA extraction
The isolation of genomic DNA was done using the phenol chloroform extraction method with certain modifications as detailed by Sobti et al. 26
Genotyping of Axin2 variants using polymerase chain reaction–restriction fragment length polymorphism
The genotyping of the seven polymorphic variants of Axin2 gene was carried out by polymerase chain reaction–restriction fragment length polymorphism (PCR-RFLP). The primers and restriction enzymes used in this protocol have been previously detailed by Gunes et al. 20 The PCR protocol used for the amplification of these genotypic variants is as follows. The 25 µL reaction mixture comprised 1 × PCR buffer, 1.5 mM MgCl2 with 0.5 µM of both forward and reverse primers, 200 µM of each deoxynucleotide triphosphates (dNTPs), 100 µg/mL bovine serum albumin (BSA), 1 U Taq polymerase (DNAzyme II DNA Polymerase; Thermo Scientific, Waltham, MA, USA), and approximately 200 ng DNA. The annealing temperature was optimized for each variant and their respective restriction enzymes have been mentioned in Table 1. The amplified product was digested with the respective restriction enzymes (5U) and was incubated at desired temperature overnight. RFLP products were resolved on the agarose gel electrophoresis/polyacrylamide gel electrophoresis depending on the difference in the size of fragments to be separated. The gels were visualized using either ethidium bromide or silver staining. Ethidium bromide can be replaced by some safe staining agent as it is toxic, and if it is used, proper precautions have to be taken. The restriction patterns were studied and scoring was done accordingly as mentioned in Table 1. The genotyping of 15% samples was repeated twice in order to check the reproducibility of the genotyping process, and the results were found to be 100% concordant.
List of primers and restriction enzymes used.

Statistical analysis
Difference in the demographic variables such as age, gender, and smoking status was evaluated using the test of significance according to the type of variable. For the categorical variables such as gender and smoking status, chi-square statistics was used. However, the continuous variables such as age and pack-years were compared using t-test among cases and controls. The genotypic frequencies of the control group were evaluated using chi-square goodness of fit in order to find whether they were in accordance with Hardy–Weinberg equilibrium (HWE; p2 + 2pq + q2 = 1). The difference in the genotypic frequencies among cases and controls was evaluated using Pearson’s χ2 test. Unconditional multivariate logistic regression approach was used to find the association between the polymorphic variants and lung cancer predisposition. This analytical approach gives an adjusted odds ratio (AOR) and 95% confidence interval (CI) which is adjusted for various confounding factors such as age, gender, and smoking status. The same approach was used for evaluating the risk associated with these polymorphic sites in the various subgroups formed on the basis of histology and smoking. All p values were two-sided, and a p value of <0.05 was considered statistically significant. 27 All the statistical analyses were performed with MedCalc version 16.8.1 (MedCalc Software, Ostend, Belgium) and SPSS Version 20.0. Haplotype and linkage disequilibrium (LD) analysis was carried out using the SHEsis software 28 (“http://analysis.bio-x.cn/myAnalysis.php”). For analyzing the inter SNP interaction, MDR approach was used to overcome the limitations of the traditional logistic regression approach. It is a non-parametric method where no hypothesis is made, and hence, no genetic inheritance model is applied. It basically analyzes all the data by reducing the data in a multidimensional manner and pooling the genotypes into high-risk and low-risk groups. It can easily access multiple loci at the same time. It finds the best interaction model on the basis of cross-validation consistency (CVC) which is number of times a model is identified as the best model and testing accuracy. The permutation testing is the method which is used by this approach to find the significance of the hypothesis generated. MDR analysis was performed using the version 0.5.1 of the open-source MDR software package that is available online (http://www.epistasis.org). 29 Another high-order interaction between all the polymorphic variants was analyzed using CART. CART was applied using the CART software (6.0; Salford Systems, San Diego, CA, USA). The CART software uses the data mining approach to create a decision tree which depicts the interaction of all the variables under study through a binary recursive partitioning method to identify subgroups at higher risk, which are not easily revealed by the traditional logistic regression models. The analysis is conducted in such a manner where the most significant predictor is used to split the sample into subgroups and continues until the differences are not significant. The tree obtained has various nodes and sub nodes depicting their role in modulating the disease susceptibility. The node with a lower case rate is used as a reference to find out the odds ratio (OR) and 95% CI for all other genotypes depicted in other nodes. 30 Multiple testing corrections were not used for any of the statistical analyses carried out in this study.
Results
Demographic characteristics
Table 2 summarizes the various demographic details of the study subjects who were recruited for this study. The study comprised 303 cases and 305 controls. The mean age of both the groups has been illustrated in Table 2 which came out to be 57.38 ± 10.74 years for cases and 53.23 ± 10.44 years for controls. The numbers of males and females were also compared between the two groups, and it was observed that there was no significant difference in both cohorts (p = 0.60). The percentage of males in cases and controls was 86% and 88%, respectively. The next important factor under consideration was smoking; an adequate attempt was made to match the number of smokers and non-smokers in both groups. It was seen that number of smokers was slightly higher in the case group (78.8% vs 73.1%). However, the difference was not found to be significant (p = 0.11). The mean pack-years were also calculated for both the study groups. The mean pack-years were higher in the cases as compared to controls (28.09 ± 34.09 years vs 19.4 ± 19.56 years). A significant difference (p = 0.0003) was found between the pack-years of cases and controls pertaining to the fact that smoking came out to be a significant risk factor for lung cancer which is an already established fact. Furthermore, considering the clinical parameters of the patients, it was found that the major percentage of patients were diagnosed with squamous cell carcinoma (SQCC; 43.23%) followed by adenocarcinoma (ADCC; 32%), and the least percentage of patients were found to suffer from small-cell lung cancer (SCLC; 23.76%). The other clinical factors under study include the TNM staging where the primary tumor extension is classified from T0 to T4, the presence and extent of lymph node involvement are classified from N0 to N3, and distant metastasis is classified into M0 and M1a/1b.
Demographic characteristics among cases and controls.

SD: standard deviation; ADCC: adenocarcinoma; SQCC: squamous cell carcinoma; SCLC: small-cell lung carcinoma; n: total number of case patients or control subjects.
p values were derived from Pearson chi-square test except age; Student’s t-test was used for age. All p values are two-sided. p < 0.05 was considered statistically significant.
Allelic and genotypic frequencies of the Axin2 variants and their association with overall lung cancer risk on the basis of histological subtypes
The genotypic and allelic frequencies of the seven different variants of Axin2 gene have been tabulated in Table 3. The seven polymorphic sites which have been evaluated in this study include Axin2 148 C > T, 1365 G > A, 432 T > C, 956 + 16 A > G, 1712 + 19 G > T, 1386 C > T, and 2062 C > T. The genotypic and allelic frequencies have been illustrated for each variant along with the minor allele frequencies (MAFs) in both the study groups. Table 3 also shows the HWE values for control group of each polymorphic site. It can be observed that the following variants were in accordance to HWE, namely, Axin2 148 (χ2 = 0.94, df = 1, and p = 0.33), 1365 (χ2 = 1.53, df = 1, and p = 0.21), 956 + 16 (χ2 = 0.31, df = 1, and p = 0.57), Axin2 432 (χ2 = 0.22, df = 1, and p = 0.63), and 2062 (χ2 = 1.84, df = 1, and p = 0.17). Only two variants, namely, Axin2 1386 (χ2 = 8.79, df = 1, and p = 0.003) and 1712 + 19 (χ2 = 175, df = 1, and p = 0.000), showed a certain deviation from HWE.
Genotypic distribution of the Axin2 genetic variants and their association with risk of lung cancer along with the stratified association analysis based on histology.

ADCC: adenocarcinoma; SQCC: squamous cell carcinoma; SCLC: small-cell lung carcinoma; AOR: adjusted odds ratio; CI: confidence interval; MAF: minor allele frequency; HWE: Hardy–Weinberg equilibrium.
AOR, 95% CI, and their corresponding p values were calculated by unconditional logistic analysis after adjusting for age, gender, and smoking status.
Two-sided χ2 test for either genotype distribution or allelic frequencies between the cases and controls.
p values in boldface depict statistically significant values.
In case of Axin2 148 C > T, the frequency of the heterozygous (CT) genotype was consistent among the cases (49.5%) and controls (47.2%). However, there lies a significant difference in between the genotypic distribution of the cases and controls (χ2 = 6.96, df = 2, and p = 0.03). MAF was found to be in 0.42 in cases and 0.49 in controls. Using CC genotype as reference, the multivariate logistic regression showed that subjects carrying the variant (TT) genotype were exhibiting a protective effect toward lung cancer risk (OR = 0.55, 95% CI: 0.34–0.87, and p = 0.01). Out of the three histological subgroups, a decreased frequency of patients carrying variant (TT) genotype was observed only in SQCC subgroup (OR = 0.51, 95% CI: 0.28–0.94, and p = 0.03). No such association was observed in ADCC and SCLC.
In case of Axin2 1365 G > A polymorphic site, when a comparison between the genotypic frequencies of cases and controls was done, a significant difference was observed (χ2 = 6.83, df = 2, and p = 0.03). The MAF was found to be 0.1 and 0.06 in controls and cases, respectively. Taking GG as a reference genotype, logistic regression analysis revealed a protective effect in patients with heterozygous (GA) genotype (OR = 0.45, 95% CI: 0.27–0.75, and p = 0.002). However, patients with variant (AA) genotype did not show any such association with risk of lung cancer. Furthermore, stratifying the subjects on the basis of their histological subtypes, it was evident that lung cancer patients carrying the heterozygous and mutant genotypes did not exhibit any significant association in any of the histological subgroups; however, significant protective effect for heterozygous genotype was found in SCLC patients (OR = 0.24, 95% CI: 0.08–0.72, and p = 0.01).
The next intronic variant of Axin2 gene under study was 956+16 A > G. The genotypic frequencies of cases and controls were compared, and it was found that there was no significant difference between the two study groups (χ2 = 0.89, df = 2, and p = 0.63). The proportion of subjects carrying the variant (GG) genotype was similar in both cases (0.6%) and controls (0.6%). MAF was comparable in cases (0.08) and controls (0.09). Association analysis was carried out using AA (wild) as a reference genotype, and it was visualized that neither of the variant genotypes, that is, AG and GG, showed any significant role in modulating the risk toward lung cancer as shown in Table 3. This same trend was also evident in subjects within every histological subtype. None of the histological subgroups showed any significant inclination toward affecting the risk associated with lung cancer in individuals carrying the combined variant (AG + GG) genotype for this polymorphic site.
For Axin2 432 T > C, the proportion of subjects with wild (TT) genotype was considerably over-represented in both cases (97.3%) and controls (94.7%). However, chi-square analysis revealed no significant difference between the genotypic frequencies among cases and controls (χ2 = 2.07, df = 1, and p = 0.14). The MAF was found to be 0.02 in controls and 0.01 in cases. We also report in this study a lack of subjects having the variant (CC) genotype both in cases and controls. Taking TT genotype as reference, the individuals carrying the heterozygous (TC) genotype did not show any significant variation in their frequencies among cases and controls. Furthermore, when patients were classified on the basis of their histological subtype, none of the subgroups revealed any association with lung cancer risk. Therefore, this polymorphic site did not confer any role toward modulating lung cancer susceptibility.
The next variant under study was Axin2 1712+19 G > T. The proportion of subjects carrying the GG genotype was negligible in the control group and 3.6% in cases. The MAF was found to be 0.43 in controls and 0.37 in cases. The genotypic frequencies were significantly different between cases and controls (χ2 = 33.8, df = 2, and p < 0.001). When OR was calculated using TT genotype as a reference, a strong protective association was observed in patients having the heterozygous (GT) genotype which was found to be highly significant (OR = 0.34, 95% CI: 0.22–0.52, and p < 0.0001). This trend continued among even in the different histological subgroups where a decreased risk of lung cancer was observed in patients carrying the GT genotype within the SQCC (OR = 0.29, 95% CI: 0.16–0.52, and p < 0.0001), ADCC (OR = 0.41, 95% CI: 0.23–0.75, and p = 0.004), and SCLC (OR = 0.16, 95% CI: 0.08–0.35, and p < 0.0001).
Another variant lying in exon 7 of the Axin2 gene, namely, 2062 C > T polymorphism, was also analyzed in this study. The frequency of wild (CC; 84.8% vs 85.5%) and heterozygous (CT; 15.1% vs 14.4%) was same in between cases and controls, respectively. No significant difference was observed in genotypic distribution of cases and controls (χ2 = 0.02, df = 1, and p = 0.88). MAF of 0.07 was observed in both cases and controls. The frequency of the variant (TT) genotype was null in both the groups. When OR was calculated using CC (wild) as a reference genotype, subjects carrying the heterozygous (CT) genotype did not exhibit any significant risk of developing lung cancer (OR = 0.96, 95%CI: 0.60–1.52, p = 0.86). When further stratification was carried out on the basis of histological subtype, this polymorphic variant did not pose any significant modulation for any of the histological subtype.
The next variant analyzed is Axin2 1386 C > T, the genotypic frequencies among cases and controls did not differ significantly (χ2 = 0.66, df = 2, and p = 0.71). The presence of subjects possessing variant (TT) genotype was almost similar in both cases and controls, respectively (12.2% vs 10.8%). MAF was found to be 0.40 in cases and 0.38 in controls. When logistic regression was applied by using CC as the reference genotype, no significant association was observed in subjects having the variant (TT) or heterozygous (CT) genotype for this polymorphic site. In the different subgroups, based on the histological subtype, only SCLC patients carrying the variant (TT) genotype showed a significant 2.8 fold risk of developing lung cancer (OR = 2.82, 95% CI: 1.18–6.73, and p = 0.01). None of the other subgroups showed any such association.
Effects of smoking status and its association between Axin2 gene polymorphisms and lung cancer risk
In order to evaluate the role of smoking in modulating the association between the Axin2 gene polymorphisms and lung cancer risk, the subjects were classified on the basis of their smoking status into smokers and non-smokers. The association analysis was carried out for all the seven polymorphic sites of Axin2 gene, and the genotypic frequencies along with the AOR with 95% CI within both the subgroups have been tabulated in Table 4. In case of Axin2 148 C > T, smokers harboring both the susceptible alleles were found to show a protective effect toward lung cancer as compared to the wild genotype patients (OR = 0.50, 95% CI: 0.29–0.85, and p = 0.01). However, no such association was observed in case of non-smokers. A similar finding was observed in case of smokers having the heterozygous GA genotype for Axin2 1365 G > A (OR = 0.51, 95% CI: 0.28–0.91, and p = 0.02). Also, the non-smokers having the same genotype as above exhibited a decreased risk toward lung cancer (OR = 0.32, 95% CI: 0.10–1.10, and p = 0.05). Patients in both smoker and non-smoker groups, possessing the combined variant genotype (GA + AA) reported a decreased risk (OR = 0.57, 95% CI: 0.33–0.98; OR = 0.33, 95% CI: 0.11–0.99) toward lung cancer with a significant p value of 0.04. In case of Axin2 1712+19 G > A, it was observed that the frequency of GT genotype was under-represented in the cases as compared to the control group. Furthermore, subjects who were smokers and carry single copy of variant allele (GT) were at a decreased risk of lung cancer as compared to controls. On the contrary, in case of the following genetic variants, namely, Axin2 432 T > C, 956+16 A > G, Axin2 2062 C > T, and 1386 C > T, no significant association was observed in any of the groups classified on the basis of smoking.
Genotypic distribution of the Axin2 variants based on smoking status and its association with risk of lung cancer.

AOR: adjusted odds ratio; CI: confidence interval.
AORs, 95% CIs, and their corresponding p values were calculated by unconditional logistic analysis after adjusting for age, gender, and smoking status.
Two-sided χ2 test for either genotype distribution or allelic frequencies between the cases and controls.
Combinatorial analysis of the risk associated with different polymorphic sites of Axin2 gene
An attempt was made to analyze the SNP to SNP interaction to evaluate the combined effect of these polymorphic sites on lung cancer risk. The genotypic frequencies of these different combinations and the AOR and 95% CIs were summarized in Table 5. Various double combinations were analyzed for finding whether there exists any significant difference between the genotypic frequencies among cases and controls. We observed that subjects who carried the single copy of variant allele for the genotypic combination Axin2 148 C > T and 1365 G > A showed a protective effect (OR = 0.42, 95% CI: 0.20–0.86, and p = 0.017). Likewise, the patients having the heterozygous (CT + CT) genotype for Axin2 148 C > T and Axin2 2062 C > T also exhibited a protective effect (OR = 0.38, 95% CI: 0.17–0.83, and p = 0.01). A strong protective effect was observed in patients carrying the heterozygous (CT + GT) genotype for Axin2 148 and Axin2 1712+19 G > T (OR = 0.21, 95% CI: 0.09–0.48, and p = 0.003). A similar effect was seen for the genotypic combination Axin2 1365 G > A and Axin2 1712+19 G < T in patients having heterozygous (GA + GT) genotype. We also found out that the subjects carrying a single copy of variant allele for Axin2 1712+19 G > T along with another single copy of variant allele of either 432 T > C (OR = 0.15, 95% CI: 0.05–0.45, and p = 0.007), 956+16 T > C (OR = 0.23 95% CI: 0.12–0.46, and p < 0.001), and (OR = 0.19, 95% CI: 0.09–0.38, and p < 0.001) showed a decreased risk of lung cancer. However, this trend was not observed only for one genotypic combination, namely, Axin2 1386 C > T and 1712+19 G > T, where no significant association was assessed between the genotypic combination and lung cancer susceptibility.
Genotypic distribution based on different genotypic combinations and their association with lung cancer risk.

OR: odds ratio; CI: confidence interval; 0: wild genotype; 1: heterozygote genotype; 2: mutant genotype; 3: combined hetero and mutant genotype.
Adjusted OR, 95% CI, and their corresponding p values were calculated by unconditional logistic analysis after adjusting for age, gender, and smoking status.
Two-sided χ2 test for either genotype distribution or allelic frequencies between the cases and controls.
Haplotype and LD analysis and their association with lung cancer susceptibility
Table 6 summarizes the frequencies for various haplotypes which were obtained on analyzing the seven polymorphic sites of the Axin2 gene. Haplotype and LD analysis was done using sHEis software. Haplotype frequencies were classified into different haplotype blocks. Only those haplotype blocks were evaluated where the case and control frequency was more than 0.03, and rest of the haplotypes were not considered for finding their correlation with lung cancer predisposition. Out of the 70 possible haplotype combinations, only seven Hap blocks showed frequency more than 0.03 and hence could be evaluated for association. Global chi-square was also calculated for comparing the frequencies among cases and controls (χ2 = 252.77, df = 7, and p = 1.33e−015). Hap blocks 1 and 5 showed an extremely low frequency of patients with the respective haplotypes among cases and hence conferring a strong protective effect toward lung cancer with highly significant p values. These Hap blocks have either variant allele for Axin2 148 C > T or 1712+19 G > T, and allele from other polymorphic sites was wild type. On the contrary, Hap3, Hap 6, and Hap7 showed an increased risk toward lung cancer as shown in Table 6. Hap block 2 consists of variant allele of Axin2 1386 C > T, and all other alleles were wild type. Similarly, Hap3 comprised variant alleles from both Axin2 1386 C > T and 1712+19 G > T and showed the maximum fourfold increased risk toward lung cancer (OR = 4.05, 95% CI: 2.95–5.57, and p = 3.68e−019).
Analyzed haplotype frequencies and risk toward lung cancer due to Axin2 haplotypes.
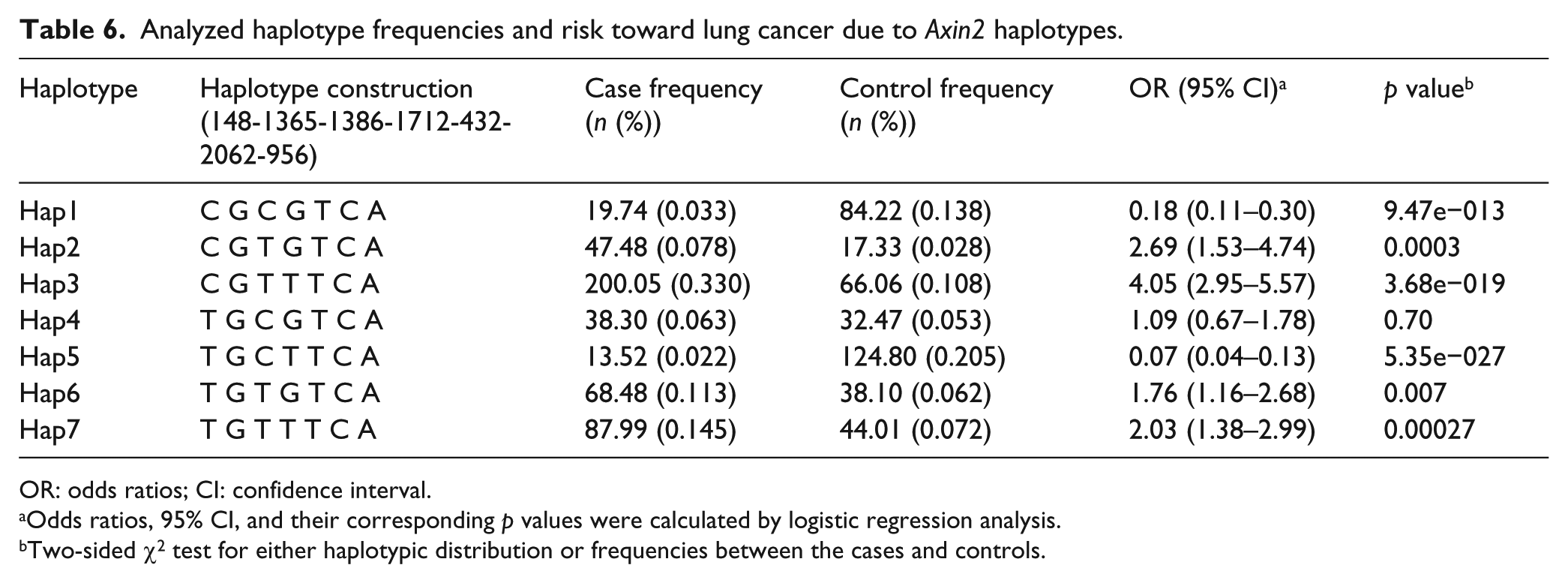
OR: odds ratios; CI: confidence interval.
Odds ratios, 95% CI, and their corresponding p values were calculated by logistic regression analysis.
Two-sided χ2 test for either haplotypic distribution or frequencies between the cases and controls.
Table 7 depicts the LD (D′) and r2 values for all the seven polymorphic sites. A strong LD was observed in case of Axin2 432 and 1365 (D′ = 0.89 and r2 = 0.001). Also, the LD was eminently strong in case of Axin2 148 and 432 (D = 0.628 and r2 = 0.007) and Axin2 956 and 432 (D = 0.243 and r2 = 0.012). The pairwise LD is illustrated in Figure 1. The blocks represent the D′ values. The red color shows the maximum LD between the two variants and white shows the least LD values.
Showing pairwise D′ (r2) values for the seven polymorphic sites among cases and controls.

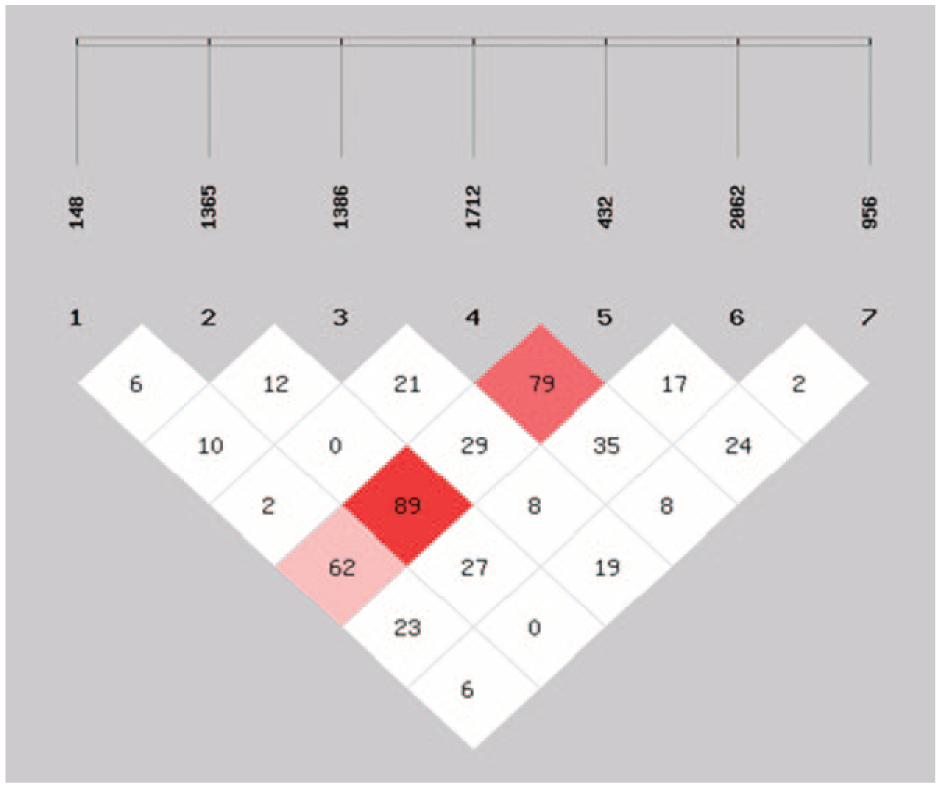
Showing pairwise linkage disequilibrium (D′) between the seven variants of Axin2 gene. red shows the maximum LD and white depicts the minimum LD between the polymorphic sites.
MDR analysis
MDR uses a computational algorithm approach to investigate all the possible interactions between these susceptibility contributing factors which might include different SNPs, other genes, and some environmental determinants. The results obtained from MDR analysis have been tabulated in Table 8. The interpretation and selection of the best model are done on the basis of the CVC and average prediction error (1-testing accuracy). The best model can be determined if it has maximum CVC and minimum prediction error or maximum testing accuracy. In this study, the best interaction model (Axin2 148, Axin2 2062, and Axin2 1712+19) has CVC 10/10 and prediction error of 0.38 with a permutation p value of 0.02. Out of the three models given by the software, this interaction model gave the minimum prediction error, maximum CVC, and significant p value. It is clear from these results that these three SNPs are evidently contributing to disease susceptibility. However, in this study, the data have not been adjusted for covariates. So, further advanced analysis can reveal some interesting findings.
Multifactor dimensionality reduction (MDR) analysis showing association of high-order interactions with lung cancer.

The entropy dendrogram obtained from this analysis is illustrated in Figure 2. Figure 2 suggests the degree and type of interaction taking place in between the SNPs which contribute in modulating the lung cancer susceptibility. As the variant Axin2 1712+19 G > T is linked with the other SNPs with a green-colored line, it shows that it has a redundant or very weak interaction with the other two SNPs, namely, Axin2 148 C > T and 2062 C > T. However, these two variants were found to be closely related as they were lined with a red-colored line which shows synergy between the two. Also, the length of the line connecting these two SNPs is shorter, and hence, it is evident that they have a strong interaction.

The multifactor dimensionality reduction (MDR) interaction dendrogram.
CART analysis
CART analysis was used to find out the complex interactive patterns of all the polymorphic sites in this study. This analysis did reveal all the possible high-order interactions in between these variants of Axin2 gene. For this interactive and complex algorithm, the mutant and heterozygous genotypes were combined as a single-variant genotype for the polymorphic sites. The decision tree gave 13 terminal nodes as shown in Figure 3. The splitting of the tree started from the Axin2 1712+19 G > T, a similar result was also observed in case of MDR result where Axin2 1712+19 G > T came out to be a single-risk factor and was the initiator of the best interaction model. Terminal node 13 (Axin2 956 + 16 (W)/Axin2 432 (M)/Axin2 1712 + 19 (M)) was selected as the reference as it has lowest case rate of 14.2. All the other genotypes were compared with the genotype of this terminal node so as to obtain the OR associated with them. The OR and 95% CI associated with all the terminal nodes have been enlisted in Table 9. The subjects having the genotype as mentioned in terminal node 9 were found to possess maximum risk of developing lung cancer (OR = 24, 95% CI: 1.68–34.08, and p = 0.01). Certain other interactive combinations reported in terminal node 1 (OR = 15.60, 95% CI: 3.29–73.84, and p = 0.0005), terminal node 5 (OR = 19.50, 95% CI: 3.00–126.48, and p = 0.001), and terminal node 7 (OR = 21, 95% CI: 2.39–183.94, and p = 0.005) also exhibited a very high risk of acquiring lung cancer with highly significant p values as illustrated in Table 9. The OR obtained from this highly complex regression analysis was not adjusted for the various confounders such as age, sex, and smoking.

CART analysis for Axin2 SNPs.
Risk estimates based on CART analysis.

OR: odds ratio; CI: confidence interval.
Case rate is percentage LC cases among all individuals in each terminal node {case/(case + control) × 100}.
p < 0.05 is considered statistically significant.
Reference group.
Discussion
In this study, the role of seven polymorphic sites, which includes five variants located in different exons of the Axin2 gene, namely, Pro50Ser in exon1, Ile144Ile in exon 1, Pro455Pro in exon5, Pro462Pro in exon 5, and Leu688Leu in exon 7, and two other intronic variants Axin2 1712 + 19 and 956+16, has been evaluated in association toward lung cancer predisposition. The loss of heterozygosity in the region where genomic locus of Axin2 lies has been reported in various cancers including ovarian, hepatocellular, and colorectal. 15 There have been reports in ovarian and colon cancers which have cited that sequence variations in the functional domains of Axin2 gene may affect its interaction with β-catenin and APC. 31 The experiments involving the truncation of APC where the Axin2 binding sites have been removed have reported its role in different human cancers. 32 There are various domains present in Axin2 protein, and the APC binding domain has significant homology with regulators of G-protein signaling (RGS); hence, it is referred as RGS domain. 33 In one of the most commonly studied SNP, Axin2 148 C > T (Pro50Ser in exon1), there is substitution of proline with serine at codon 50 which lies in close proximity to RGS domain. Its SNP might play a role in modulating Axin2 binding efficiency. 21
It was keen to observe a protective effect in subjects who carried the variant (TT) genotype for Axin2 148 C > T (p = 0.01). Many studies have evidently reported the same findings in case of various cancers. A study carried in Turkish population in lung cancer patients also gave the same insight about this polymorphic site where in the TT genotype conferred a decreased risk of lung cancer. 20 The similar results have been reported in Chinese 23 and Japanese lung cancer patients. 21 Also, similar reports are available in case of other type of cancers including breast, 33 prostate, 24 astrocytoma, 25 and ovarian. 22 This suggests that being a potent tumor suppressor gene, the variations in the genetic sequence might compensate on its function of regulating Wnt cascade as it is an indispensable part of the control regime of Wnt signalling. 34 Another variant lying in exon 5, Axin2 1365 G > A, has revealed a protective effect in subjects who were carrying a single copy of the susceptible allele. However, no other study has reported such an association in case of lung cancer, perhaps only one study has evaluated this SNP in lung cancer where no association was reported. 20 The reasons behind this difference might pertain to very low sample size and also the differences in ethnicity. Although, this variant is also studied in case of Turkish astrocytoma patients where no association was observed for this polymorphic site in regard of disease susceptibility. Another polymorphic site which was found to have a very pronounced protective effect in our study was Axin2 1712+19 G > T, which is located in intron 5 of the Axin2 gene. Axin2 1386 and 1712 + 19 showed a certain deviation from HWE. The reasons behind this slight deviation might be population stratification, random mating, and sample bias for these two variants. The intronic variants may have been examined to identify their role in influencing the m-RNA formation. These variations may lead to differential splicing pattern, and henceforth, it affects the primary structure of the protein. 35 However, the studies done in Turkish lung cancer 20 and astrocytoma patients 25 are not concordant with the findings of our study.
In case of two polymorphic sites, namely, Axin2 956+16 A > G and 2062 C > T, no effect was observed on lung cancer risk in this study. However, these two variants are correlated with the tooth agenesis in Brazilian population. 36 Genetic variant 956+16 A > G lies in intron2 and has been reported to influence the splicing process at exon2, although this interaction is weak. 37 Similarly, 2062 C > T, which is although a synonymous substitution of leucine with leucine at 688 codon has been proposed to effect the splicing process by disrupting the enhancer sequences present just before the splice site in exon7. 37 In line with our findings, these genetic variants did not confer any modulation in Turkish lung cancer patients. 20
Furthermore, in this study, lung cancer risk was also evaluated in various subgroups made on the basis of histological subtype. Being a diverse type of carcinoma, these subtypes of lung cancer have been predicted to have different disease etiology and pathogenesis. 38 In our study, the Axin2 148 C > T was found to harbor a protective effect in SQCC patients (p = 0.03). These results are concordant with those observed in Turkish lung cancer patients. 20 In case of 1365 G > A, the Turkish population did not exhibit any significant association in any of the subgroups, 20 and our study also revealed similar results for SQCC patients, but a slight association was observed in SCLC patients. For all other five variants, the results from the study carried on Turkish lung cancer population suggest that they do not contribute in modulating the risk of the disease particularly in any histological subgroup. 20 Our findings are in line with this study except in the case of Axin2 1712 + 19 G > T where a decreased risk was observed in all the three subgroups.
Smoking has always been considered as one of the major factors contributing toward the risk of developing lung cancer. This factor may interact with other genetic variabilities differently to modulate the individual’s predisposition toward the oncogenesis of lung tissue. This study evaluated the risk associated with these polymorphic sites in smokers and non-smokers separately. A previous study done in Turkey has reported a lower risk of lung cancer in case of smokers having mutant (TT) genotype of Axin2 148 C > T. Our findings are also in accordance to the results of this study. This previous study has examined the risk of all the seven polymorphic sites under study in smokers and non-smokers. No effect has been observed for any of the category in case of Axin2 432 T > C, 956+16 A > G, 2062 C > T, 1386 C > T, and 1712+19 G > A. 20 Our study also revealed no significant correlation in smokers and non-smokers for above-mentioned sites except for 1712+19 G > T, where smokers possessing GT genotype showed a protective association.
Haplotype and LD analysis has revealed the status of the linkage of these seven genetic variants present on Axin2 gene. This is probably the first attempt to find out the haplotype blocks comprising the seven alleles which significantly contribute to lung cancer risk in North Indians. The Hap block which showed the maximum risk toward lung cancer comprised the variant allele from both Axin2 1386 C > T and 1712+19 G > T, and all other alleles were wild type. Recent study done in Chinese NSCLC patients has examined LD and risk associated with the different haplotypes of Axin2 gene (rs2240308, rs22040307) and MMP7 (rs4791169). 39 A similar kind of study was also carried out in ovarian cancer patients of Poland, where haplotype analysis was done for four different SNPs of Axin2 gene. 22
MDR approach was also applied to find the various non-parametric interactions taking place in between these polymorphic variants. None of the previous studies have analyzed such high-order interactions for all the seven polymorphic sites of Axin2 gene. This analysis revealed the best interaction model (Axin2 148, Axin2 2062, and Axin2 1712+19) with maximum CVC (10/10) and minimum prediction error (0.38) along with significant permutation p value. Very few reports are available where MDR was employed to find out these interactions; one such recent study was done in Chinese population where Axin2 gene (rs2240308 and rs22040307) and MMP7 (rs4791169) and smoking were taken as a parameter to find out the best interaction model contributing toward lung cancer risk. 39 Another study also used this exhaustive MDR method in ovarian cancer to find out the interactions between variants of Axin2, β-catenin, and APC. 22
So far, this is the first study where high-order regression analysis was carried out to obtain the decision tree and odds associated with combinations of various genotypes using CART analysis. This was the first attempt to look into the SNP–SNP interaction taking place between the Axin2 genetic variants and their contribution in modulating lung cancer predisposition as Axin2 is a potent tumor suppressor gene. This was evident from the results of this study that Axin2 148 C > T and Axin2 1712+19 G > T are the major role-playing SNPs in the interactive models obtained both from MDR and CART. These kinds of novel innovative findings can help in defining the role of these sequence variants within the tumor suppressor genes in the process of carcinogenesis. This study has given an insight about the genetic variants of Axin2 protein, which is an indispensable participant of the destruction complex maintaining β-catenin levels and hence regulating Wnt pathway. However, the functional significance of these variations is yet unclear. Studies in near future with larger sample size and in different ethnicities can support these findings.
Conclusion
Evidences from this study have exhibited an interesting correlation of these seven genetic variants with lung cancer susceptibility. The data obtained from this study reveals a strong association of these SNPs with lung cancer risk in North Indians. This study has employed certain innovative high-order computational algorithms such as MDR and CART to find the interactions between these polymorphic sites and their contribution in lung cancer predisposition.
Footnotes
Acknowledgements
We would like to express our gratitude to all the subjects who participated in this current study. Dr Digamber Behra and Dr Navneet Singh have provided access to data, helped in the approval of study, and guided in the analysis of data. Dr Siddharth Sharma has designed the study, supervised, and edited the manuscript. Charu Bahl has carried out all the experimental work and statistical analysis and drafted the manuscript under the esteemed supervision of Dr Siddharth Sharma, who is the corresponding author for this research work and also the principal investigator.
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
This work was supported by grant from the Indian Council of Medical Research, New Delhi, India (Grant No. 5/13/126/2011/NCD-III).