
Abstract
Arsenic trioxide (As2O3) has been used clinically as an anti-tumor agent. Its mechanisms are mostly considered to be the induction of apoptosis and cell cycle arrest. However, the detailed molecular mechanisms of its anti-cancer action through cell cycle arrest are poorly known. Furthermore, As2O3 has been shown to be a potential DNA methylation inhibitor, inducing DNA hypomethylation. We hypothesize that As2O3 may affect the expression of cell cycle regulatory genes by interfering with DNA methylation patterns. To explore this, we examined promoter methylation status of 24 cell cycle genes in breast cancer cell lines and in a normal breast tissue sample by methylation-specific polymerase chain reaction and/or restriction enzyme–based methods. Gene expression level and cell cycle distribution were quantified by real-time polymerase chain reaction and flow cytometric analyses, respectively. Our methylation analysis indicates that only promoters of RBL1 (p107), RASSF1A, and cyclin D2 were aberrantly methylated in studied breast cancer cell lines. As2O3 induced CpG island demethylation in promoter regions of these genes and restores their expression correlated with DNA methyltransferase inhibition. As2O3 also induced alterations in messenger RNA expression of several cell cycle–related genes independent of demethylation. Flow cytometric analysis revealed that the cell cycle arrest induced by As2O3 varied depending on cell lines, MCF-7 at G1 phase and both MDA-MB-231 and MDA-MB-468 cells at G2/M phase. These changes at transcriptional level of the cell cycle genes by the molecular mechanisms dependent and independent of demethylation are likely to represent the mechanisms of cell cycle redistribution in breast cancer cells, in response to As2O3 treatment.
Introduction
The cell cycle is an intricate and highly controlled process implicated in cell growth and division, DNA-damage response, and diseases including cancer.1,2 Abnormalities in the expression of cell cycle regulatory genes resulting in higher proliferative capacity have been found in almost all human tumors, such as breast cancer.3,4 The cell’s progression through G1, S, G2, and M phases of the cell cycle occurs in an orderly fashion and is controlled by the cooperative activity of a set of cell cycle regulatory genes.5,6 These genes consist of those that encode cyclin-dependent kinases (CDKs) and the proteins that modulate their activity, cyclins, and cyclin-dependent kinase inhibitors (CKIs). 6
The mechanisms governing cancer initiation and progression are the consequence not only of genetic alterations but also of epigenetic perturbations.7–9 Aberrant changes in the promoter methylation pattern are the most common mechanism for the transcriptional inactivation of many genes and are found in different types of human tumors. 10 A number of genes such as tumor suppressor genes, cell cycle control genes, DNA repair genes, and apoptosis-related genes are regulated by methylation of their promoter regions. 11 In proliferating cells, DNA methylation is laid down by DNA methyltransferases (DNMTs; DNMT1, DNMT3A, and DNMT3B). Inhibition of the expression of these proteins by DNA methylation inhibitors would lead to the progressive reduction in DNA methylation in newly divided cells. 12 DNA methylation inhibitors cause re-expression of methylation-silenced tumor suppressor genes resulting in the arrest of tumor growth and have emerged as an effective strategy against cancer. 13 One of the recently identified DNA methylation inhibitors is arsenic trioxide (As2O3).14,15 This multi-target drug represents an efficient treatment option for both new and relapsed cases of acute promyelocytic leukemia (APL)16–19 and also has potential therapeutic value for the treatment of other malignant hematopoietic diseases and several solid tumors including breast cancer.20–22 Its mechanisms have been intensively studied and are mostly considered to be the induction of cell apoptosis and cell cycle arrest.23,24 Furthermore, As2O3 exposure has been shown to cause hypomethylation around the promoter region of several tumor suppressor genes leading to their re-expression in various cancer cells.14,25 The mechanism of DNA hypomethylation after exposure to As2O3 is not clear. However, the arsenic metabolism may be associated with hypomethylation of promoter CpG islands in As2O3-treated cells. In a recent study using an APL cell line, we have shown that As2O3 by biotransformation into the intracellular methylated metabolites through arsenic methyltransferase (AS3MT) catalysis and by the suppression of DNMTs (DNMT1, DNMT3a, and DNMT3b) messenger RNA (mRNA) expression causes the consumption of methyl donors. The depletion of methyl groups led to the inability to maintain methylated cytosines in DNA, resulting in DNA demethylation. 15
One of the mechanisms that have been proposed through which As2O3 eliminates APL and other cancer cells is the accumulation of cells in the phase of cell cycle and the subsequent mitotic arrest–associated apoptosis or mitotic catastrophe.26–30 However, the cell cycle regulation mechanism of As2O3 has not yet been clarified, especially in solid tumors. 31 These reports led us to hypothesize that the induction of DNA hypomethylation is one of the molecular mechanisms underlying the As2O3-promoted cell cycle arrest in breast cancer cells mediated by changes in the expression level of cell cycle–related genes. To provide preliminary support of this hypothesis, we first analyzed the promoter methylation status of 24 genes key to cell cycle regulation in different types of breast cancer cell lines and in a normal breast tissue sample, by methylation-specific polymerase chain reaction (MSP) and/or restriction enzyme–based methods, and then examined whether As2O3 could demethylate and restore the expression of hypermethylated genes.
Materials and methods
Cell lines
In this study, we used three different types of human breast cancer cell lines, which may mirror some of the substantial genomic and transcriptional abnormalities and biological heterogeneity found in breast tumors: MCF-7 cells (ER+, PR±, and HER2−) exhibits low levels of invasion and belongs to the luminal subtype of human breast cancer cell lines; MDA-MB-231 cell line (ER−, PR−, and HER2−) is highly invasive and placed in the basal-B subtype; and MDA-MB-468 cells (ER−, PR−, and HER2−) have an intermediate invasive activity and belong to the basal-A subtype.32–34
These cell lines were obtained from the National Cell Bank of Iran, Pasteur Institute of Iran (NCBI, Tehran, Iran). All cell lines were cultured in RPMI 1640 medium (Invitrogen, Auckland, New Zealand); supplemented with 10% fetal bovine serum (FBS; Invitrogen), 2 g/L sodium bicarbonate (Sigma, St. Louis, MO, USA), and 1% penicillin/streptomycin (Biosera, Ringmer, East Sussex, UK) in a humidified incubator at 37°C in 5% CO2; and harvested with 0.25% trypsin–0.03% EDTA.
As2O3 treatment and 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide assay
As2O3 was provided by Sina Darou (Tehran, Iran). For As2O3 treatment, the relevant amounts of stock solution (5 mM) of As2O3 were added to a culture medium to attain the desired concentrations. The cells were cultured in the absence and presence of varying concentrations of As2O3, ranging from 1 to 5 µM concentrations.
3-(4,5-Dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) assay was used to determine the inhibitory effect of As2O3 on proliferation and cell viability. The cells at a density of 5000 cells/100 µL/well were seeded into 96-well plates (SPL Life Sciences, Pocheon, Korea) overnight and then treated with the desired concentrations (1, 2, 3, 4, and 5 µM) of As2O3 for 24, 48, and 72 h. After treatment, the media were replaced with 100 µL of MTT solution (0.5 mg/mL) (Sigma) and the cells were further incubated at 37°C for 3 h. Precipitated formazan was solubilized with 100 µL of dimethyl sulfoxide (DMSO), and the optical densitometry was measured at a wavelength of 570 nm in an enzyme-linked immunosorbent assay (ELISA) reader. The percentage of cell viability was calculated as (%) = (ODexp/ODcon) × 100, where ODexp and ODcon are the optical densities of treated and untreated cells, respectively.
Flow cytometric analysis of cell cycle distribution
Cellular DNA content and cell cycle distribution were determined by flow cytometric analyses before and after treatment of cultured cells with different concentrations of As2O3 for 48 h. Briefly, 1 × 106 trypsinized cells from control (untreated) and treated cells were harvested, washed with cold phosphate-buffered saline (PBS), and then fixed in cold 70% ethanol to store at 4°C overnight. Next, fixed cells were pelleted by centrifugation and were resuspended in stain solution containing PBS, propidium iodide 50 µg/mL (Invitrogen), RNase A 100 µg/mL (Sigma), and 0.05% Triton X-100 at 37°C. After 30-min incubation, samples were analyzed by ParTec PAS III flow cytometer (ParTec, Munich, Germany), and data were interpreted using the Windows™ FloMax® software.
Genomic DNA isolation
The total genomic DNA was isolated from cell lines, before and after As2O3 treatment, by a standard salting out procedure. The normal breast tissue sample (reduction mammoplasty) was obtained after surgical resection from Shaghayegh Clinic in Tehran, Iran. Genomic DNA was extracted from the frozen tissue using QIAamp® DNA Mini Kit (Qiagen, Valencia, CA, USA), according to the manufacturer’s instructions, and used as a negative control for methylation. In addition, human peripheral blood samples from normal donors were provided by Hematology, Oncology and Stem Cell Transplantation Research Center, Shariati Hospital, Tehran, Iran. Normal lymphocytes (NL) (mononuclear cells) were isolated from human peripheral blood by density-gradient centrifugation using Ficoll-Paque™ (GE Healthcare Life Sciences, Parramatta, Australia). Then, their total genomic DNA was also used as a positive control for unmethylated alleles of each gene. The DNA samples were used immediately or stored frozen until analysis at −20°C.
Promoter methylation analysis (restriction enzyme–based method)
The Human Cell Cycle EpiTect Methyl II Signature qPCR Array (SABiosciences, a QIAGEN company, Mainz, Germany) was used for bisulfite-free determination of the promoter methylation status of a panel of 22 genes that positively or negatively control the cell cycle. According to the manufacturer’s procedure, the isolated genomic DNAs were digested with two different restriction endonucleases, a methylation-sensitive restriction enzyme (MSRE) and/or a methylation-dependent restriction enzyme (MDRE). Following digestion, the relative amount of remaining DNA in each individual enzyme reaction was quantified by real-time polymerase chain reaction (PCR) using primers that flank a promoter region of gene of interest. The relative fractions of methylated and unmethylated DNA were subsequently determined by comparing the amount in each digest with that of a mock (no enzymes added) using a ΔCT method. The product of the mock represents the total amount of input DNA for real-time PCR detection. Therefore, the methylation status of individual genes as percentages of unmethylated and methylated fraction of input DNA was determined using a data analysis template which was provided by the manufacturer (SABiosciences).
Sodium bisulfite modification of extracted DNA and methylation-specific PCR
A total of 1–2 µg of genomic DNA was treated with sodium bisulfite using the EpiTect Bisulfite Kit (Qiagen, Hilden, Germany) according to the manufacturer’s protocol.
Based on the CpG-rich islands in the promoter sequence of RBL1 and cyclin D2, methylation- and unmethylation-specific primers were designed using MethPrimer program (www.urogene.org/methprimer/). Primer sequences used for MSP of RASSF1A are from previously published study. 35 The sequences of PCR primers, annealing temperatures, and the expected sizes of PCR products are summarized in Table 1. The PCR reaction was performed by FastStart Taq DNA Polymerase, dNTPack (Roche Applied Science, Mannheim, Germany), according to the supplier’s recommendations and included forward and reverse primers (10 pmol/µL of each primer pair per reaction), and bisulfite-modified DNA (30–100 ng) in a final volume of 25 µL. Taq DNA Polymerase 2× Master Mix RED (Ampliqon, Odense, Denmark), consisting of 1.5 mM MgCl2 final concentration, was also used; in this case, reactions were hot-started at 95°C for 5 min prior to PCR cycling and the addition of Ampliqon Master Mix. Amplification of three genes were performed with a thermal cycler instrument (Applied Biosystems Veriti™, Foster City, CA, USA) under following conditions: 1 cycle of 95°C for 5 min; 40 cycles of 95°C for 30 s, at specific annealing temperature as listed in Table 1 for 25 s, and 72°C for 25 s; and a final extension of 7 min at 72°C. A reaction without DNA (water blank) was used as negative control reaction for contamination checking in each set of PCR reactions; EpiTect® Control DNA (human), methylated and bisulfite converted (Qiagen, Venlo, Limburg, Netherlands), containing in vitro-methylated and bisulfite-converted DNAs, was used as positive control for methylated alleles; NL DNA and normal breast tissue sample DNA used as negative controls for methylation show unmethylated DNAs. After amplification, 10 µL of the PCR products was resolved by electrophoresis in 3% agarose gels (UltraPure™ Agarose; Invitrogen, Carlsbad, CA, USA), stained with SYBR® Safe DNA Gel Stain (Invitrogen, Carlsbad, CA, USA), and visualized under ultraviolet (UV) illumination (Bio-Rad’s Gel Doc™ XR+ System, Hercules, CA, USA). All MSP assays were repeated at least three times.
PCR primer sequences used for promoter methylation analysis, the expected sizes of PCR products, and annealing temperatures.

M: methylated-specific primer; U: unmethylated-specific primer; F: forward primer; R: reverse primer.
Analysis of gene expression by quantitative real-time PCR
Total RNA from cultured cells was extracted by TriPure Isolation Reagent (Roche Applied Science) according to manufacturer’s instructions and quantified by NanoDrop ND-1000 (NanoDrop Technologies, Wilmington, Delaware, USA). Then, 1 µg of RNA from each sample was applied to synthesize complementary DNA (cDNA) using the PrimeScript RT Reagent Kit (Takara Bio Inc., Otsu, Japan) according to the manufacture’s specifications. Real-time PCR assay was performed with a light cycler instrument (Roche Diagnostics, Mannheim, Germany) using SYBR Premix Ex Taq (Takara Bio Inc.). In a final volume of 20 µL in a capillary tube, 2 µL of cDNA samples, 10 µL SYBR Green Master Mix, 0.5 µL of each forward and reverse primers (10 pmol), and 7 µL of nuclease-free water (Qiagen, Hilden, Germany) were added to conduct real-time PCR. Thermal cycling conditions involved an initial activation step of 30 s at 95°C followed by 40 cycles including a denaturation step of 5 s at 95°C and a combined annealing/extension step of 20 s at 60°C. Melting curves were analyzed to validate single PCR product of each primer. All primer pairs amplified a single fragment without primer dimer. Hypoxanthine phosphoribosyltransferase 1 (HPRT1) was amplified as a housekeeping gene, and the fold change in expression of each target mRNA relative to HPRT1 was calculated based on 2−ΔΔCt relative expression formula. The sequences of forward and reverse primers and the relevant products sizes are shown in Table 2.
Nucleotide sequences of the primers used for quantitative real-time RT-PCR.

Statistical analysis
All experiments were carried out in duplicate or triplicate, and the results have been expressed as mean ± standard error (SE). Statistical significances of difference were determined using paired two-tailed Student’s t-tests. Statistically different values were considered significant at p values <0.05 compared to corresponding control.
Results
DNA methylation pattern of cell cycle regulatory genes in breast cell lines
In this study, we analyzed the promoter methylation pattern of 24 different genes (22 genes by a restriction enzyme–based PCR method and 2 additional genes by MSP method) in two breast cancer cell lines (MCF-7 and MDA-MB-231) and in a normal breast tissue sample. After identifying methylated genes, the effect of As2O3 on the promoter methylation pattern of the methylated genes was analyzed by methylation-specific PCR (MSP) method in three cell lines (including MDA-MB-468 cell line).
The functional gene groupings are listed in Table 3; all these genes are involved in the cell cycle regulation such as phase transitions and checkpoints, DNA replication, and cell cycle arrest. The promoter methylation status of a panel of 22 genes (from SABiosciences) is listed in Table 4 as percentage of unmethylated and methylated fraction of input DNA in MCF-7 and MDA-MB-231 cell lines and also in normal breast tissue. According to the manufacturer, methylated represents the fraction of input genomic DNA containing two or more methylated CpG sites in the targeted region of a gene and unmethylated represents the fraction of input genomic DNA containing no methylated CpG sites in the amplified region of a gene. The real-time PCR results revealed that 55.8% and 55.1% of total input DNA had methylated RBL1 (p107) gene promoter in MCF-7 and MDA-MB-231 cell lines, respectively, compared to 2.8% in normal breast tissue. Additionally, the methylation frequencies of GADD45A were 10.9% and 4.6% in MCF-7 and MDA-MB-231, respectively, compared to 0.3% in normal breast tissue. However, the promoter region of the remaining genes did not show significant methylation in human breast cancer cell lines.
24 cell cycle regulatory genes: functional gene groupings.

The promoter methylation status of a panel of 22 genes key to cell cycle regulation in MCF-7 and MDA-MB-231 breast cancer cell lines and in a normal human breast tissue.

UM: unmethylated; M: methylated.
Methylation status of the CpG islands of two more cell cycle–related genes, RASSF1A and cyclin D2, was analysed by MSP in MCF-7, MDA-MB-231, and MDA-MB-468 cell lines. Selection of RASSF1A and cyclin D2 was based on PubMeth (www.pubmeth.org); among the most analyzed genes for DNA methylation in breast cancer, two well-known cell cycle genes, RASSF1A and cyclin D2, are mostly hypermethylated in the majority of human breast cancers. Our MSP results show that RASSF1A and cyclin D2 were almost fully methylated in all three cell lines.
Inhibitory effect of As2O3 on survival of breast cancer cells
MTT assay was used to determine the suppressive effect of various concentrations of As2O3 at different time intervals (24, 48, and 72 h) on proliferation and viability of MCF-7, MDA-MB-231, and MDA-MB-468 cell lines. As shown in Figure 1, As2O3 induced a remarkable reduction of viability in a dose- and time-dependent manner in three breast cancer cell lines. For example, at 48-h treatment with 1, 2, 3, and 4 µM As2O3, viability of MCF-7 cells was decreased by 7.84%, 25.57%, 43.7%, and 60.2% and MDA-MB-231 cells by 8.31%, 43.97%, 66.14, and 72.02%; and viability of MDA-MB-468 cells was reduced by 40.5%, 59.15%, 72.25%, and 80.5%, respectively (p < 0.05).

Evaluation of As2O3 effect on the viability of MCF-7, MDA-MB-231, and MDA-MB-468 breast cancer cell lines. The cells were treated with indicated concentrations of As2O3 and their viability was assessed by MTT assay after 24, 48, and 72 h. Values are represented as percentage of viability compared to the untreated control cells and are given as mean ± SE from three independent experiments (p < 0.05 by one-way variance analysis).
As2O3 causes redistribution of cell cycle in breast cancer cells
Cellular DNA content and cell cycle changes were determined by flow cytometric analyses after 48 h of incubation with various concentrations of As2O3. As shown in Figure 2, As2O3 increased the number of MCF-7 cells accumulated in G1 phase and reduced the number of MCF-7 cells in S and G2/M phases of the cell cycle. On the contrary, As2O3 induced G2/M arrest and markedly decreased the proportion of cells in G1 phase in MDA-MB-231 cell line. In MDA-MB-468 cells, As2O3 induced both S and G2/M arrest and markedly reduced the proportion of cells in G1 phase. Collectively, these results suggest that As2O3 induces the cell cycle arrest at G1, S, and/or G2/M phases depending on the breast cancer cell lines.
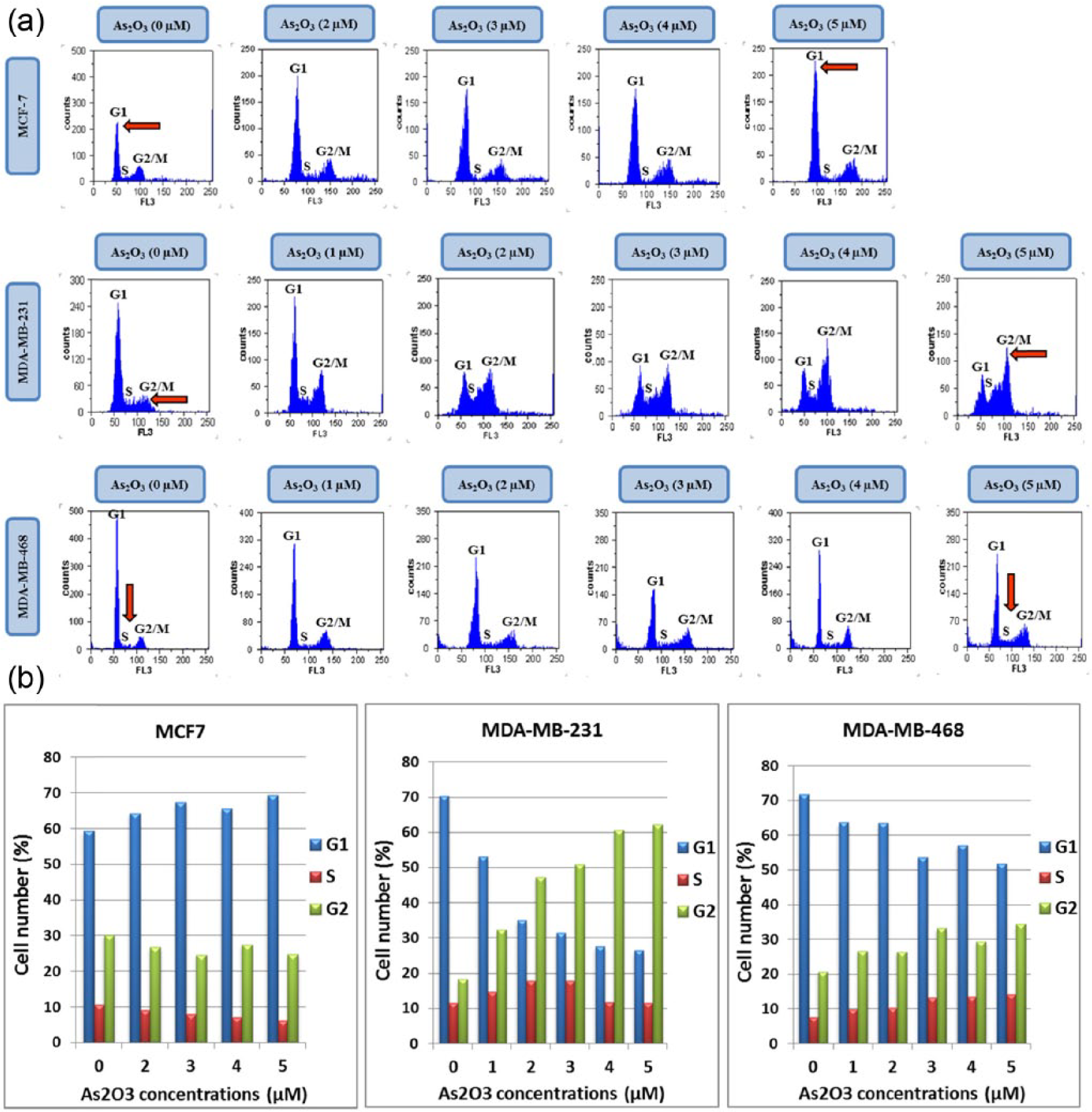
As2O3-induced cell cycle arrest at different phases in MCF-7, MDA-MB-231, and MDA-MB-468 cell lines: (a) flow cytometric analyses of cell cycle distribution in As2O3-treated breast cancer cells stained with propidium iodide (PI) and (b) percentage of cells accumulated at G1, S, and G2/M phases of the cell cycle depending on the cell type after 48 h of incubation with different concentrations of As2O3. Columns indicate means of three independent experiments.
As2O3-induced demethylation of promoter-associated CpG islands of cell cycle–related genes in breast cancer cell lines
To determine whether As2O3 can act as a demethylating agent, the methylation status of the CpG islands of three genes (RBL1, RASSF1A, and cyclin D2), which were highly methylated, was examined by MSP method in MCF-7, MDA-MB-231, and MDA-MB-468 cell lines before and after treatment with As2O3.
Based on MTT results, all three breast cancer cell lines were treated with As2O3 at concentration of 3 µM for 48 h for performing bisulfite conversion and MSP reactions. As shown in Figure 3, after treatment with As2O3, the RBL1, RASSF1A, and cyclin D2 genes were converted into partially demethylated genes in MCF-7, MDA-MB-231, and MDA-MB-468 cell lines. We showed a significant decrease in the methylated-specific bands and an increase in the unmethylated-specific bands of RBL1, RASSF1A, and cyclin D2 genes in cells treated with As2O3. It is useful to note that the analysis of DNA methylation pattern of the RBL1 promoter by MSP yielded results almost similar to the restriction endonuclease–based method. Collectively, our study suggests that As2O3 treatment could result in demethylation of the hypermethylated CpG islands of the cell cycle–related genes in different human breast cancer cell lines.
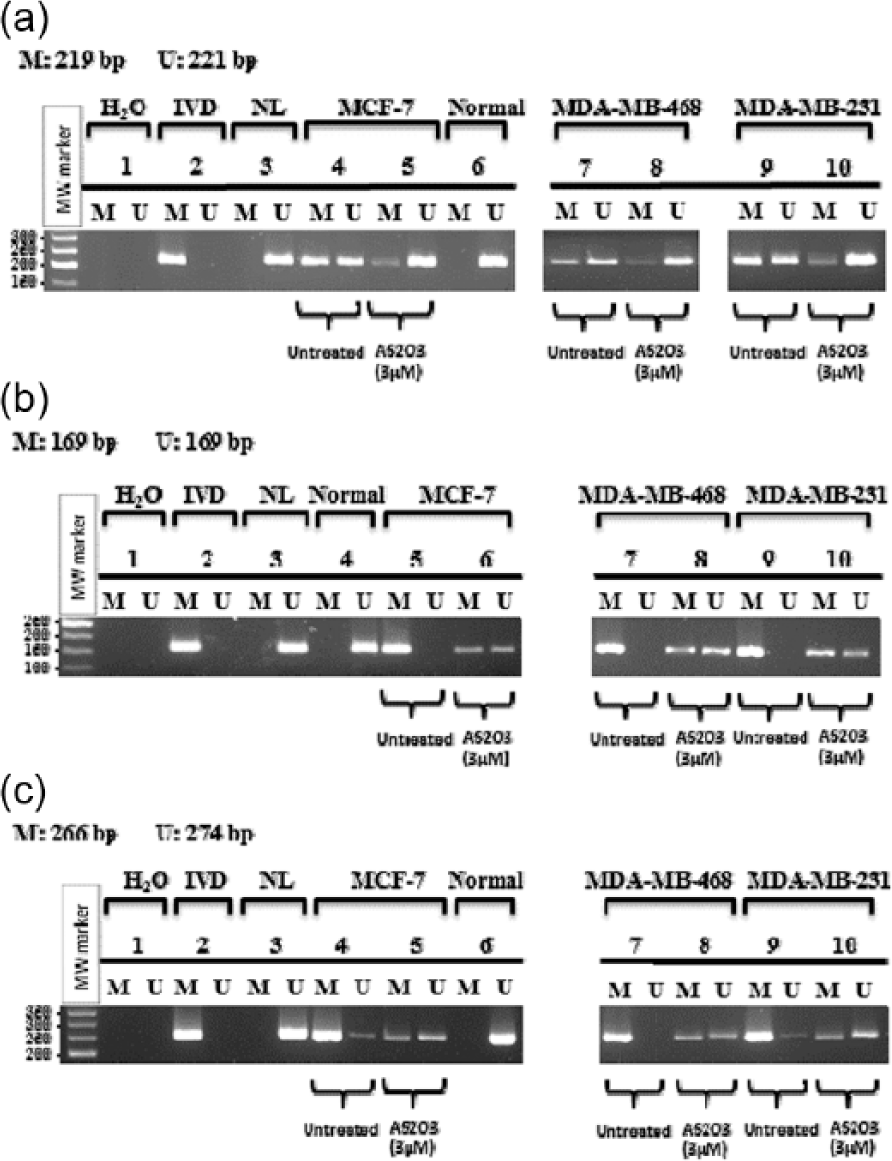
Methylation analysis of the three cell cycle–related gene promoters, (a) RBL1, (b) RASSF1A, and (c) cyclin D2, by MSP in three breast cancer cell lines. Cell lines, MCF-7, MDA-MB-468, and MDA-MB-231, were treated with 3 µM of As2O3 for 48 h, and CpG island promoter methylation status of these cell cycle genes was determined using methylation-specific PCR. H2O served as a negative control reaction without DNA (water blank) for control of contamination in the PCR reaction. In vitro–methylated DNA (IVD) and normal lymphocytes (NL) were used as positive and negative controls for methylation, respectively. Normal breast tissue (reduction mammoplasty) was used as another negative control for methylation. Untreated and As2O3-treated cell lines were amplified with methylated (M) and unmethylated (U) primers after bisulfite-treated genomic DNA.
Alterations in mRNA expression levels of DNMTs and AS3MT in As2O3-treated breast cancer cells
To investigate the mechanism of As2O3-induced DNA hypomethylation, we used quantitative real-time PCR to determine whether DNMTs and AS3MT mRNA levels would be changed in breast cell lines after treatment with different concentrations of As2O3 for 48 h. As presented in Figure 4, As2O3 inhibited DNMT1 (maintenance DNMT) mRNA levels in all three breast cancer cell lines (p < 0.05). The mRNA expression of DNMT3a as a de novo DNMT was decreased in MCF-7 and MDA-MB-468 cell lines, but not in MDA-MB-231 cells. Interestingly, the mRNA expression of DNMT3b (de novo DNMT) was increased in three breast cancer cells after As2O3 treatment. The mRNA level of AS3MT, a key enzyme that catalyzes the conversion of inorganic As2O3 to methylated products, was increased steadily in As2O3-treated MCF-7 and MDA-MB-468 cells (p < 0.05; Figure 4(a) and (c)); however, its expression was reduced in MDA-MB-231 cells (Figure 4(b)).

The effect of As2O3 on transcriptional levels of DNMT1, DNMT3a, DNMT3b, and AS3MT in breast cancer cell lines: (a) MCF-7, (b) MDA-MB-231, and (c) MDA-MB-468 cells were treated with various concentrations of As2O3 for 48 h. Data are shown as fold change in relative expression levels of transcripts determined by quantitative real-time PCR. Values are normalized using the expression of the housekeeping HPRT1 on the basis of comparative Ct (2−ΔΔCt) method. Values are given as mean ± SE of three independent experiments (p < 0.05).
Correlation between DNA methylation pattern and basal expression of RBL1, RASSF1A and cyclin D2 genes in breast cancer cell lines
In order to determine whether a relationship exists between promoter DNA methylation status and basal expression level of RBL1, RASSF1A, and cyclin D2 genes in the untreated MCF-7, MDA-MB-231, and MDA-MB-468 cell lines, the percentage of methylated bands (as compared to the unmethylated) was estimated from the intensity of bands in the gel image using Quantity One and Multi-Analysis softwares (Bio-Rad Laboratories, Hercules, CA, USA). The basal gene expression level (expression level in the untreated cells) was measured by quantitative reverse transcription PCR (QRT-PCR), as described before.
As shown in Figure 5, the results show that the expression of RBL1 was partially silenced, whereas expression of RASSF1A and cyclin D2 was almost fully silenced in all three untreated cell lines, suggesting that the downregulation of RBL1 mRNA levels and the loss of RASSF1A and cyclin D2 expressions in the studied breast cancer cell lines were probably as the result of promoter methylation.

Relation between percentage of (a) promoter methylation and (b) basal gene expression level of RBL1, RASSF1A, and cyclin D2 in untreated MCF-7, MDA-MB-231, and MDA-MB-468 cell lines. Values are given as mean ± SE of three independent experiments.
As2O3-induced re-expression or upregulation of demethylated cell cycle genes mRNA in breast cancer cells
Figure 6 shows increases in mRNA expression levels of RBL1, RASSF1A, and cyclin D2 genes in breast cancer cells treated with different concentrations (1, 2, 3, and 4 µM) of As2O3 for 48 h, relative to the untreated cells. The mRNA level of RBL1 was markedly enhanced and mRNA expression of RASSF1A and cyclin D2 was indeed restored by As2O3 treatment in all three cell lines. These results are consistent with the concept that demethylation of the methylation-silenced cell cycle genes is linked to the As2O3-mediated re-expression or upregulation of these genes.

Effects of As2O3 on mRNA expression level of genes involved in cell cycle regulation in breast cancer cell lines. The effects of different concentrations of As2O3 on the mRNA levels of methylated genes (RBL1, RASSF1A, and cyclin D2) and unmethylated genes (p21, GADD45A, cyclin D1, cyclin D1, and cyclin D1) involved in cell cycle regulation were analyzed after treatment for 48 h in (a) MCF-7, (b) MDA-MB-231, and (c) MDA-MB-468 cells. Data are shown as fold change in expression level of transcripts relative to the untreated controls as determined by QRT-PCR. Values were normalized using the expression of the housekeeping HPRT1 on the basis of comparative Ct (2−ΔΔCt) method. Values are given as mean ± SE of three independent experiments (p < 0.05).
As2O3 changes transcriptional level of unmethylated cell cycle genes in breast cancer cells
To gain further insights into the effect of As2O3 on cell cycle, we examined the mRNA expression of several other genes key to the cell cycle regulation in breast cancer cells exposed to various concentrations of As2O3 for 48 h. Based on the results of the restriction enzyme assays, these cell cycle–related genes were unmethylated in MCF-7 and MDA-MB-231 cell lines.
As indicated in Figure 6, the expression levels of cell cycle negative regulatory genes p21, p53, and GADD45A were significantly increased in MCF-7 cells with wild-type p53 gene (p < 0.05). While, the mRNA level of p53 was not measured in the MDA-MB-231 and MDA-MB-468 cell lines with mutant p53. As2O3 reduced the mRNA levels of cyclin D1 and cyclin E1 in MCF-7 cells (p < 0.05), while the mRNA level of these genes was increased or not changed in MDA-MB-231 and MDA-MB-468 cells. The expression of cyclin B1, a positive regulator of cell cycle, was noticeably decreased (p < 0.05) in all three breast cancer cell lines, more significantly in MDA-MB-231 and MDA-MB-468 cell lines.
Discussion
It is now well known that the deregulation of cell cycle progression is one of the most important changes during cancer development. Understanding the molecular details of the cell cycle control and checkpoint aberrations in cancer and manipulation of these mechanisms may provide insights into the discovery of effective treatment strategies.36,37
In this study, we aim to determine promoter methylation status of 24 different cell cycle regulatory genes in different subtypes of breast cancer cell lines and in a normal breast tissue sample by MSP and/or restriction enzyme–based methods. The function of these cell cycle regulatory genes is listed briefly in Table 5. Our methylation analysis indicates that only promoters of RBL1 (p107), RASSF1A, and cyclin D2 genes were aberrantly methylated in all three cell lines. A low level of methylation was also found at the promoter of GADD45A gene in MCF-7 and MDA-MB-231 cells lines. Promoter methylation of these genes was probably tumor-specific phenomenon, confined to tumors and did not occur in the normal breast tissue. While our results revealed that a large number of important genes involved in controlling cell cycle checkpoints and DNA repair were unmethylated in breast cancer cells, it seems that the DNA methylation alterations are not implicated as a regulator of abnormal expression of many cell cycle genes in these breast cancer cells.
24 cell cycle genes studied for methylation.

PCNA: proliferating cell nuclear antigen; Mre11-Rad50-Nbs1: MRN.
It has been shown that As2O3 can inhibit cancer cell proliferation by induction of cell cycle arrest in many cancer cells, including breast cancer. However, the molecular mechanisms by which As2O3 regulates various genes that cause redistribution of cell cycle remain to be elucidated. Recent studies have suggested novel chemo-preventative properties of As2O3 as DNMTs inhibitor.14,15,38 In this study, we also found that As2O3 inhibits DNMT1 expression in all three breast cancer cells lines, but not DNMT3a or DNMT3b. The results are similar to those that reports that As2O3 inhibits DNMT1 protein and mRNA in the colon 39 and liver cancer cells. 14 The prototypic DNMT, DNMT1 (maintenance DNMT), has far higher catalytic activity than DNMT3a and DNMT3b; 9 it is essential and sufficient for maintenance of global DNA methylation and aberrant CpG island methylation patterns in human tumor cells.14,40,41 It has been shown in colon, breast, bladder, and lung cancer cells that reduction of DNMT1 levels is sufficient to induce demethylation and the resultant re-expression of methylation-silenced genes.40,42–45 We also found that As2O3 increased AS3MT expression level in MCF-7 and MDA-MB-468 cell lines but reduced in MDA-MB-231. These findings suggest that although AS3MT may be the predominant enzyme in the pathway for As2O3 methylation, there could be AS3MT-independent mechanisms and unidentified methyltransferases involved in alternative As2O3 methylation pathways.46–50
In order to demonstrate that As2O3 as a DNA methylation inhibitor can induce hypomethylation of aberrantly methylated cell cycle regulatory genes in breast cancer cells, we examined the methylation status of promoter-associated CpG islands of the RBL1, RASSF1A, and cyclin D2, before and after treatment with As2O3 by MSP in MCF-7, MDA-MB-231, and MDA-MB-468 cell lines. Treatment with 3 µM of As2O3 for 48 h induced hypomethylation of the partially or fully hypermethylated CpG islands around the promoter regions of the RBL1, RASSF1A, and cyclin D2 genes in all three cell lines. Interestingly, the detection of DNA methylation pattern at the RBL1 promoter by MSP yielded similar results to restriction enzyme–based method. These results suggest that the ability of As2O3 to alter DNA methylation of the cell cycle genes may be an important mechanism leading to cell cycle arrest and apoptosis in breast cancer cells.
Furthermore, as it was expected, our results showed that there was an inverse correlation between promoter DNA methylation status and basal expression level of RBL1, RASSF1A, and cyclin D2 genes in the untreated MCF-7, MDA-MB-231, and MDA-MB-468 cell lines (Figure 5). The mRNA levels of RASSF1A and cyclin D2 genes were undetectable in all three cell lines before the As2O3 treatment where the promoters of these genes were fully hypermethylated. Whereas, RBL1 gene had some detectable expression in all three cell lines where the promoter of this gene was partially methylated or possibly was hemimethylated.
We show for the first time that the RBL1 (p107) gene promoter was aberrantly partially methylated in all three breast cell lines compared to the normal breast tissue. Like its family members (Rb and p130), RBL1 gene product is worthily involved in transcription repression and tumor suppression and plays a key role in negative control of cell cycle progression.51,52 RBL1 is most highly expressed during the G1 to S transition of the cell cycle in actively dividing cells and stably associated with cyclin E/Cdk2 and cyclin A/Cdk2 complexes and inhibited their kinase activity in late G1 phase and during the S phase.51–53 In our study, a methylation frequency of RBL1 in MCF-7 was 55.8% and in MDA-MB-231 was 55.1% compared to only 2.8% in normal breast tissue. As2O3 treatment induced demethylation of the RBL1 gene in all three cell lines up to 50%. At the same time, As2O3 induced significant upregulation of its mRNA (about five-fold) in MCF-7 and a slight increase in MDA-MB-231 and MDA-MB-468 cell lines.
RASSF1A is an important human tumor suppressor protein acting as a negative regulator of cell cycle progression through inhibition of G1/S. It can engage the Rb family cell cycle checkpoint and inhibit cyclin D1 accumulation probably through inhibition of its mRNA translation, which likely prevents RASSF1A-expressing cells from entering S phase.54–56 Epigenetic inactivation of tumor suppressor gene RASSF1A by hypermethylation of CpG islands in the promoter region is an extremely common event in a considerable proportion of human cancers (at least 37 tumor types), including 49%–62% of breast cancers.54,55,57 In our study, the promoter region of RASSF1A was completely methylated in all three cell lines and unmethylated in normal breast cancer tissue. As2O3 treatment induced hypomethylation of the RASSF1A genes in all three cell lines. The expression level of this gene was undetectable; As2O3 indeed restored mRNA expression level of RASSF1A in all three cell lines (45-, 13- and 100-fold increase in MCF-7, MDA-MB-231, and MDA-MB-468 cell lines, respectively).
Cyclin D2, a major regulator of cell cycle, is implicated in regulation of G1- to S-phase transition. Expression of cyclin D2 is unique among the D-type cyclins, being upregulated in a variety of normal human tissues, including breast tissue.6,58 Whereas, loss of cyclin D2 expression by hypermethylation of its promoter region was found to be associated with tumor development in breast, lung, gastric, and pancreatic cancer, indicating that cyclin D2 might be involved in a vital tumor suppressor function in a cancer-type dependent manner.10,58–60 Our results also showed aberrant methylation of the cyclin D2 gene in all three breast cancer cell lines and correlated with silencing of the gene expression. When the cells were exposed to As2O3, the cyclin D2 promoter was partially demethylated, and cyclin D2 mRNA expression was restored.
To gain further insights into the anti-cancer effect of As2O3 on cell cycle, we investigated the effect of As2O3 treatment on the mRNA expression level of several other important unmethylated or slightly methylated cell cycle regulatory genes, including p21, p53, and GADD45A which are known as negative regulators of cell cycle progression and cyclin D1, cyclin E1, and cyclin B1 as positive modulators of the cell division. These cyclins are key components of the cell cycle machinery that are responsible for the progression through the G1/S and G2/M phases of the cell cycle.61–64 In our experiments, differential expression of cyclins was observed after As2O3 treatment in all three breast cancer cell lines, by a mechanism independent of demethylation.
The mRNA expression of GADD45A was increased in a dose-dependent manner in three studied breast cancer cell lines after As2O3 treatment (p < 0.05). It has been reported that GADD45A protein plays a role in the G2/M checkpoint in response to the DNA damage, and its most prominent action at the G2/M transition is disrupting the CDK1/cyclin B1 complex. However, GADD45A can also interact with p21 resulting in cell cycle arrest at both G1/S and G2/M checkpoints. 5 GADD45A gene is mostly downregulated in many cancer cells including breast cancer; 65 the decrease in expression level of this gene is often, but not always, associated with its promoter methylation. It has been reported that As2O3 induces the GADD45A expression in cancer cells; however, the mechanism of induction is not known. 66 In this study, even though the methylation frequencies of GADD45A were 10.9% and 4.6% in MCF-7 and MDA-MB-231 cells, respectively, compared to 0.3% in normal breast tissue, the As2O3 treatment caused a significant upregulation of GADD45A expression level (about 16- and 11-fold increase, respectively). In this regard, the upregulation of GADD45A may have some significant roles in As2O3-induced cell cycle arrest in breast cancer cells.
In a previous study, we have shown that As2O3 is able to induce apoptosis and block cell cycle progression in the G2 phase in human APL NB4 cells. 23 Also, it has been demonstrated that As2O3 can cause the cell cycle arrest at different phases, G1/S and G2/M transitions,21,23,67–69 and even in S phase, 70 in cancer cells. Liu et al. 69 reported that the As2O3-induced G1 or G2/M cell cycle arrest in myeloma cells was dependent on p53 status. Here, our study demonstrated that As2O3 is capable of perturbing the cell cycle of the human breast cancer cells at various phases. Flow cytometric analysis revealed that As2O3 induced G1 arrest and inhibited cell cycle progression to the S and G2/M phases in MCF-7 cells. The growth arrest was associated with an inhibition of G1 cyclin (cyclin D1 and cyclin E1) expression. As2O3 also induced upregulation of p53, p21, GADD45A, RBL1, RASSF1A, and cyclin D2 by demethylation-independent and/or dependent mechanisms. p21 is a CKI required for the p53-mediated G1 arrest leading to apoptosis in cancer cells. 71 In the presence of functional p53, As2O3 might act like a DNA damaging agent 69 that triggers p53-dependent DNA repair apparatus involving upregulation of p21 and GADD45A and blocking of cyclin D1 and cyclin E1 followed by the G1 arrest and eventually leading to apoptosis in MCF-7 cells with p53 wild-type.
On the contrary, As2O3 induced G2/M arrest in MDA-MB-231 cells as well as S and G2/M arrests in MDA-MB-468 cells. Cyclin D1 and cyclin E1 levels were increased or remained unchanged, and alterations in p21 expression were not significant in these two cell lines. It has been reported that in the lack of functional p53 and G1 arrest, DNA damage can lead to a p53-independent G2/M arrest but involving other DNA damage–responsive proteins such as ATR and ATM; this can block the formation of the mitotic cyclin B1/CDK1 complex and induce G2/M cell cycle arrest.61,69 Moreover, GADD45A may also be involved in the G2/M checkpoint through several mechanisms such as interacting with p21 or disrupting the interaction between CDK1 and cyclin B1.63,64 In this study, As2O3 clearly decreased the mRNA expression of cyclin B1 and upregulated the GADD45A mRNA level in these cell lines.
In conclusion, no single mechanism has been able to elucidate all of the effects seen with As2O3; yet, it most likely acts at multiple levels, with different modes of actions. However, we provided new evidence that the induction of DNA hypomethylation probably is one of the molecular mechanisms underlying the As2O3-promoted cell cycle arrest in breast cancer cells. As2O3 as a DNA methylation inhibitor via inhibition of DNMT1 mRNA level induced demethylation of the promoter-associated CpG islands resulting in upregulation of several cell cycle–related genes such as RBL1, RASSF1A, and cyclin D2 in breast cancer cell lines. As2O3 depending on the cell type induced the cell cycle arrest at different phases of breast cancer cell lines; it also induced alterations in the mRNA expression of several cell cycle regulatory genes, such as p21, GADD45A, p53, cyclin D1, cyclin E1, and cyclin B1 independent of demethylation in breast cancer cells. Overall, As2O3 induces cell cycle arrest at G1, S, or G2 phases, depending on the cell type, and modulates cell cycle regulatory gene expression by the molecular mechanisms dependent and/or independent of demethylation in breast cancer cells. Promoter methylation and mRNA expression of more cell cycle–related genes need to be analyzed in response to As2O3 effect in order to explore exact As2O3 mechanisms in induction of cell cycle arrest.
Footnotes
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
This study was funded by Hematology, Oncology and Stem Cell Transplantation Research Center, Shariati Hospital, Tehran University of Medical Sciences, Tehran, Iran.