
Abstract
Curcumin, a major yellow pigment and spice in turmeric and curry, is a powerful anti-cancer agent. The anti-tumor activities of curcumin include inhibition of tumor proliferation, angiogenesis, invasion and metastasis, induction of tumor apoptosis, increase of chemotherapy sensitivity, and regulation of cell cycle and cancer stem cell, indicating that curcumin maybe a strong therapeutic potential through modulating various cancer progression. It has been reported that microRNAs as small noncoding RNA molecules are related to cancer progression, which can be regulated by curcumin. Dysregulated microRNAs play vital roles in tumor biology via regulating expressions of target genes and then influencing multiple cancer-related signaling pathways. In this review, we focused on the inhibition effect of curcumin on various cancer progression by regulating expression of multiple microRNAs. Curcumin-induced dysregulation of microRNAs may activate or inactivate a set of signaling pathways, such as Akt, Bcl-2, PTEN, p53, Notch, and Erbb signaling pathways. A better understanding of the relation between curcumin and microRNAs may provide a potential therapeutic target for various cancers.
Introduction
Curcumin, extracted from the rhizomes of Curcuma longa, has been widely used in China and India for medicinal purposes for thousands of years, in view of its biological and pharmacological properties1–3 (Figure 1). The β-diketone chain of curcumin leads to the existence of keto and enol tautomeric conformations in equilibrium which plays a vital role in anti-oxidant activities of curcumin. 4 The anti-cancer activities of curcumin were based on its anti-oxidant potential, and many targets were identified, such as COX (cyclooxygenase) and NF-κB.5,6 Curcumin has a wide spectrum of biological functions such as anti-inflammatory, anti-oxidant, anti-coagulation, anti-hyperlipidemia, anti-atherosclerosis, anti-mutagen, as well as anti-neoplastic. 7 Although curcumin has been reported to have variable effects in many diseases, the potential anti-cancer properties of curcumin have attracted more interest in recent years, and its mechanism of anti-cancer has been studied the most.8,9 Curcumin has shown anti-tumor effects on several cancers, including blood system tumor, respiratory and digestive tract cancer, genitourinary tumors, gynecological tumors, head and neck squamous cell carcinoma, melanoma, and leukemia.10,11 In addition, curcumin could serve as a “chemosensitizer” and even reverse the multiple drug resistance of malignancies. However, the anti-cancer mechanism of curcumin has not been fully understood yet. Recent studies have shown that curcumin exerts its anti-cancer effect through the regulation of epigenetic changes, including regulation of histone modification, inhibition of DNA methyltransferases, and regulation of microRNA (miRNA) expressions.12,13 Studies showed that curcumin could regulate the expression of miRNAs in various cancers through which curcumin may exert its anti-cancer effect.14,15 MiRNAs are a broad class of small, noncoding, endogenous RNAs that play a vital function in regulating gene expression by messenger RNA (mRNA) degradation or translation repression.16–18 MiRNAs as one of the largest class of gene regulators are abundance in many human cell types and regulate the expression of about 60% of mammalian genes.3,19,20 A large number of studies demonstrated that miRNAs are first transcribed as long primary miRNA molecules (pri-miRNAs) by RNA polymerase II and then pri-miRNAs are processed into pre-miRNAs by DGCR8 (DiGeorge syndrome critical region 8) and Drosha (RNase III enzyme). Pre-miRNAs are transported into the cytoplasm and cleaved to miRNA duplex. Mature miRNAs are critically involved in the pathogenesis, evolution, and progression of cancers21,22 (Figure 2). According to their role on the development of tumor, miRNAs could be classified into two groups: onco-miRNAs and tumor suppressor miRNAs. 23 Dysregulated miRNAs play vital roles in tumor biology via regulating expression of target genes and then influencing multiple cancer-related signaling pathways.24,25 For instance, it was reported that miRNA-188-5p could suppress hepatocellular carcinoma cell proliferation and metastasis by directly targeting FGF5 (fibroblast growth factor 5). 26 Increased miR-152 sensitized cisplatin-resistant ovarian cancer cells through suppressing cisplatin-induced autophagy, increasing cisplatin-induced apoptosis, and restricting cell proliferation. 27 Another study showed that miR-205 acted as a tumor radiosensitizer in breast cancer (BC) cells by targeting ZEB1 (zinc finger E-box binding homeobox 1) and Ubc13 (the ubiquitin-conjugating enzyme). 28 The ectopic miRNAs were associated with curcumin anti-cancer effects, including induction of tumor cell apoptosis, inhibition of tumor proliferation, restraint of cell invasion and metastasis, regulation of cell cycle and cancer stem cell (CSC), and increase of chemoradiotherapy sensitivity. In this review, we will summarize available evidence on the anti-cancer effects of curcumin through regulating expression of miRNAs.

Chemical structure of curcumin (1,7-bis-4(hydroxy-3-methoxyphenyl)-1,6-heptadiene-3,5-dione). Curcumin is the yellow spice extracted from the rhizome of Curcuma longa.

MiRNAs are a class of short highly conserved noncoding RNAs, which undergo sequential processing by various enzymes such as RNA pol II, Drosha, DGCR8, Exportin-5, and Dicer from transcription of miRNA genes. Mature miRNAs regulate multiple gene expressions through binding to the 3′-untranslated region (UTR) of target mRNAs, leading to inhibition of mRNA translation or mRNA cleavage.
The mechanisms of curcumin regulate expression of miRNAs
Extensive studies have shown that curcumin has potent anti-cancer effect through regulating the expression of miRNAs. MiR-21 is upregulated in various human cancers, wherein accumulating evidence indicates that it acts as an oncogene. In colorectal cancer (CRC), curcumin could restrain pri-miR-21 transcriptional regulation to decrease expression of miR-21 through transcription factor AP-1 (activator protein-1). AP-1 was reported to bind with the pri-miR-21 promoter which could increase the promoter activity and then reinforce the expression of miR-21. Curcumin treatment not only decreased the binding of c-Fos/AP-1 and p-c-Jun/AP-1 to the promoter but also inhibited the activation of c-Jun. 15 In addition, Yang et al. found that in prostate cancer (PCa), p65 NF-κB subunit could increase miR-21 expression by binding to the miR-21 promoter. Curcumin could function as a selective regulator of the NF-κB signaling to block the activation of the NF-κB which resulted in the inhibition of miR-21 expression.29,30 Methylation as a fundamental regulator of gene transcription may be another mechanism by which curcumin regulates miRNA expression. For example, curcumin could cause epigenetic modulation of miR-203 expression through demethylating the miR-203 promoter in bladder cancer. Curcumin treatment led to reversing methylation of the miR-203 promoter which suggested that curcumin could elevate the expression of miR-203. 14 EZH2 (enhancer of zeste homolog 2) as a histone methyltransferase enhancer was reported to associate with depressions of miRNAs (let-7 family, miR-26a, miR-101, and miR-200b) in pancreatic ductal adenocarcinoma (PDAC) cells. Curcumin could significantly inhibit the expression of EZH2 mRNA in tumor cells, leading to re-expression of miR-200c, miR-101, miR-26a, and let-7 family in pancreatic cancer cells. Interestingly, the re-expression of miR-101 could also lead to decreased mRNA expression of EZH2. Taken together, curcumin inhibited pancreatic cancer progression through regulating an EZH2-miRNA regulatory circuit. 31 Tumor hypoxia results in deregulated expression of hypoxia inducing factor (HIF) which could increase expression of EZH2 and also increase the expression of oncogenic miR-21 in PCa cells. Curcumin therapy showed anti-tumor activity through restraining the expression of HIF, EZH2, as well as miR-21 under hypoxic condition. 32 P53 as a tumor-suppressive transcription factor was associated with the expression of miR-215 and miR-192-5p (miR-215/192-5p). Curcumin could increase miR-215/192-5p expression via p53-dependent mechanisms in non-small cell lung cancer (NSCLC) cells. 33 In oral squamous cell carcinoma (OSCC), epidermal growth factor (EGF) was reported to elevate the expression of miR-31 (an oncogenic miRNA). EGF treatment led to miR-31 upregulation through the activation of Akt. Curcumin exhibited anti-cancer potential through attenuating Akt activation which then resulted in the downregulation of miR-31 34 (Table 1). These findings suggested that the mechanisms of curcumin-regulated expression of miRNAs are complicated, including the activation/inactivation of miRNAs transcription, the regulation of associated signaling pathway, the methylation/demethylation of miRNAs promoter, and the inhibition of some proteins such as EZH2 and HIF.
The mechanisms of curcumin regulate expression of miRNAs.

Curcumin inhibits cell proliferation and promotes apoptosis by regulating miRNAs
Curcumin could suppress proliferation and induce apoptosis of cancer cells through PI3K-Akt signaling pathway by regulating expression of miRNAs. The PI3K/Akt signaling pathway has been found to play an important role in the survival, proliferation, and apoptosis of tumor cells. It has been shown that curcumin inhibited cell proliferation and induced apoptosis of NSCLC cells through the upregulation of miR-192-5p and suppression of the PI3K/Akt signaling pathway. 35 The tumor suppressor PTEN (phosphatase and tensin homolog) was originally identified as a negative regulator of PI3K/Akt pathway. Curcumin was reported to increase the expression of PTEN through producing suppression effect on miR-21. 36 P53 is another well-studied tumor suppressor gene and can be negatively regulated by PI3K/Akt pathway. 37 For example, a study showed that curcumin modulated miR-19/PTEN/Akt/p53 axis to exhibit its tumor promotor effects against bisphenol A–associated BC promotion. 38 Curcumin was also reported to increase transcriptional activity of p53 directly in NSCLC, and then, the upregulation of p53 induces the expression of miR-192-5p and miR-215, which decreased expression of XIAP (X-linked inhibitor of apoptosis protein) to serve as tumor promotors. 33 Several downstream targets of the PI3K/Akt signaling pathway, such as p21 and Bcl-2, were also regulated by curcumin. P21 is an important factor in the control of the cell cycle. Curcumin could promote the expression of p21 via inhibiting the expression of miR-208 in PCa cells. 39 MiR-34a reinforced by curcumin could inhibit BC cells proliferation and induce BC cells apoptosis through decreasing expression of Bcl-2 and Bmi-1. 40 Curcumin also exerted the same effect on BC cells through downregulation of Bcl-2 and survivin/BIRC5 via upregulating miR-181b. 41 Besides, miR-203/Akt2/Src axis also could be regulated by curcumin through which curcumin decreased proliferation and increased apoptosis of bladder cancer cells. 14 Taken together, PI3K/Akt signaling pathway is one of important pathways associated with cell proliferation and apoptosis and regulated by curcumin (Figure 3).

Curcumin regulates expressions of miRNAs to exert its inhibitory effects on suppressing proliferation and inducing apoptosis of cancer cells by inactivation of PI3K/Akt signaling pathway. Curcumin can activate PTEN, a negative regulator of PI3K/Akt pathway, through regulating expression of miR-19 and miR-21. Curcumin also regulates the downstream targets of PI3K/Akt pathway such as Akt, XIAP, p53, p21, and Bcl-2 to suppress the proliferation of tumor through upregulating miR-192-5p, miR-203, miR-181b, and miR-34a or downregulating miR-19, miR-125-5p, and miR-208. Moreover, curcumin can directly activate p53 and then increase expression of miR-192-5p and miR-215 which decreases the level of XIAP.
In addition to regulate PI3K-Akt signaling pathway, curcumin regulates a number of other genes or signaling pathways by mediating miRNAs to suppress proliferation and induce apoptosis of cancer cells. Curcumin-induced miR-145 restrained proliferation and triggered apoptosis of glioblastoma (GBM) cells through inhibition of genes implicated in pluripotency, including OCT4 (octamer-binding protein 4) and SRY (sex determining region Y)-box 2 (SOX-2). 42 Curcumin upregulated the expression of miR-186 (an anti-tumor miRNA) which mediated inhibitory effect of curcumin on lung adenocarcinoma (LA) progression through targeting caspase-10. 43 Curcumin also decreased the WT1 (Wilms’ tumor 1) expression and then restrained proliferation partly by upregulating miR-15a and miR-16-1 in leukemic cells. 44
Curcumin provides a potential therapeutic strategy for cancer treatment, since numerous miRNAs have been linked to cancer cell proliferation and apoptosis could be dysregulated by them. Curcumin inhibits cancer cell proliferation and/or promotes apoptosis through upregulating a set of tumor suppressor miRNAs, including miR-15a, miR-34a, miR-181b, miR-186, miR-192-5p, and miR-215, or downregulating numerous onco-miRNAs, like miR-19, miR-21, and miR-208. The dysregulated miRNAs mediate the inhibitory effect of curcumin on cancer cell proliferation by activating or inactivating multiple signaling pathways, including Akt, p53, PTEN, and Bcl-2 pathways.
Curcumin regulates cell cycle via miRNAs
Curcumin has recently been linked to regulate cell cycle in various cancers. Increasing evidence has shown that curcumin modulated cell cycle to exert its anti-cancer activities through regulating multiple miRNAs such as miR-21, miR-34a, miR-19, and miR-145. In CRC cells, treatment of RKO and HCT116 cells with curcumin increased the fraction of cells with 4N DNA which represents the G2/M phase. Curcumin resulted in G2/M phase arrest possibly through decreasing the expression of miR-34a and miR-2.15,45 Moreover, miR-21 as an onco-miRNA significantly increased the number of viable cells and reduced cell cycle arrest at G2/M phase in GBM cells. 46 Further study confirmed that treatment with curcumin in GBM cells increased the SubG1 phase of cells, while decreased the cells in the G1 or S phase. Curcumin-induced miR-145 may account for the increase in SubG1 phase through targeting OCT4 and SOX-2. 47 In BC, treatment of MCF-7 cells with bisphenol-A (BPA) accelerated G1 to S phase progression. Curcumin could reverse BPA-elicited increase in the percentage of cells in S phase partly through miR-19/PTEN/AKT/p53 axis to prevent BPA-triggered cell cycle progression. 38 MiRNAs modulate cell cycle progression in cancer and specifically contribute to the death or proliferation of the cancer cells. Moreover, curcumin inhibitory effects are mediated by miRNAs suggesting that curcumin and miRNAs utilize distinctive signaling pathways to regulate cancer cell cycle.
Curcumin modulates miRNAs to restrain cancer invasion and metastasis
It is generally recognized that cancer invasion and migration contribute to the high risk for the relapse of cancer patients. Invasion as a critical step in metastasis is currently the focus of many researches.48,49 While growing evidences show that curcumin may serve as a stronger therapeutic agent and regulate multiple signaling pathways through miRNAs to exert its anti-tumor activities. For example, curcumin upregulated miR-22, a well-known tumor suppressor miRNA, which inhibited expression of Erbb3 (erythroblastic leukemia viral oncogene homolog 3). Erbb3 is an activator of PI3K/AKT signaling pathway which plays an important role in the migration of red blood (RB) cells. 50 P53, Bcl-2, and p27kip1 are downstream targets of PI3K/AKT pathway which can also be regulated by curcumin. Curcumin exerted its inhibitory effect on nasopharyngeal carcinoma (NPC) metastasis through inhibiting expression of miR-125a-5p and subsequently increasing expression of P53. 50 The expression of p53 was associated with cancer aggressiveness correlating with increased grading of invasive cancer front and cancer depth. 51 Bcl-2 was originally identified as a key regulator of apoptosis, but there are increasing evidences that show that Bcl-2 could also promote cancer cell migration and invasion. 52 Curcumin-induced miR-34a inhibited cell invasion through restraining expression of Bcl-2. 53 MiR-221 as an oncogenic miRNA could be decreased by curcumin in pancreatic cancer. Decreased miR-221 subsequently increased expression of p27kip1 and PTEN which led to the inhibition of cell proliferation and migration. 54 In addition, PTEN as a negative regulator of PI3K/Akt pathway could also be regulated by miR-21. Curcumin downregulated miR-21 to inhibit metastasis of PC through increasing PTEN. 30 Decreasing miR-21 by curcumin also stabilized the expression of the tumor suppressor PDCD4 (programmed cell death protein 4) which plays an anti-metastatic role in CRC. 15 The PI3K-Akt pathway was also implied to be involved in the regulation of PDCD4 degradation. Increasing PDCD4 by curcumin could inhibit cancer cell proliferation as well as restrain cell invasion and metastasis.55,56 Some studies have confirmed that PDCD4 inhibited invasion and metastasis partly through PDCD4/MMPs (matrix metalloproteinases) and PDCD4/AP-1 pathways in various cancers.57,58 MMPs play an essential role in cancer metastasis because of its ability to degrade extracellular matrix. Curcumin inhibited MMP-2 expression through upregulating miR-874 in NSCLC cells. 59 In metastatic BC cells, miR-181b induced by curcumin inhibited expression of MMPs which are prerequisite in tumor progression of invasion and metastasis. 41 Curcumin treatment of pancreatic cancer cells targeting the re-expression of miR-200b/c could decrease MMPs expression and cause re-expression of PTEN. 60 The miR-200 family is well known to inhibit metastatic biology. 61 Curcumin attenuated the activation of cell migratory machinery via increasing expression of miR-33b in melanoma cells. Upregulated miR-33b directly targeted HMGA2 (high-mobility group AT-hook 2) to suppress melanoma migration. HMGA2 played an important role in enhancing cancer metastasis via downregulating miR-200b expression62,63 (Figure 4). Curcumin also increased miR-7 which led to the inhibition of cell migration and invasion in pancreatic cancer. The upregulated miR-7 and subsequently downregulated SET8, a target of miR-7, markedly reduced invasive capacity of pancreatic cancer cells. 64
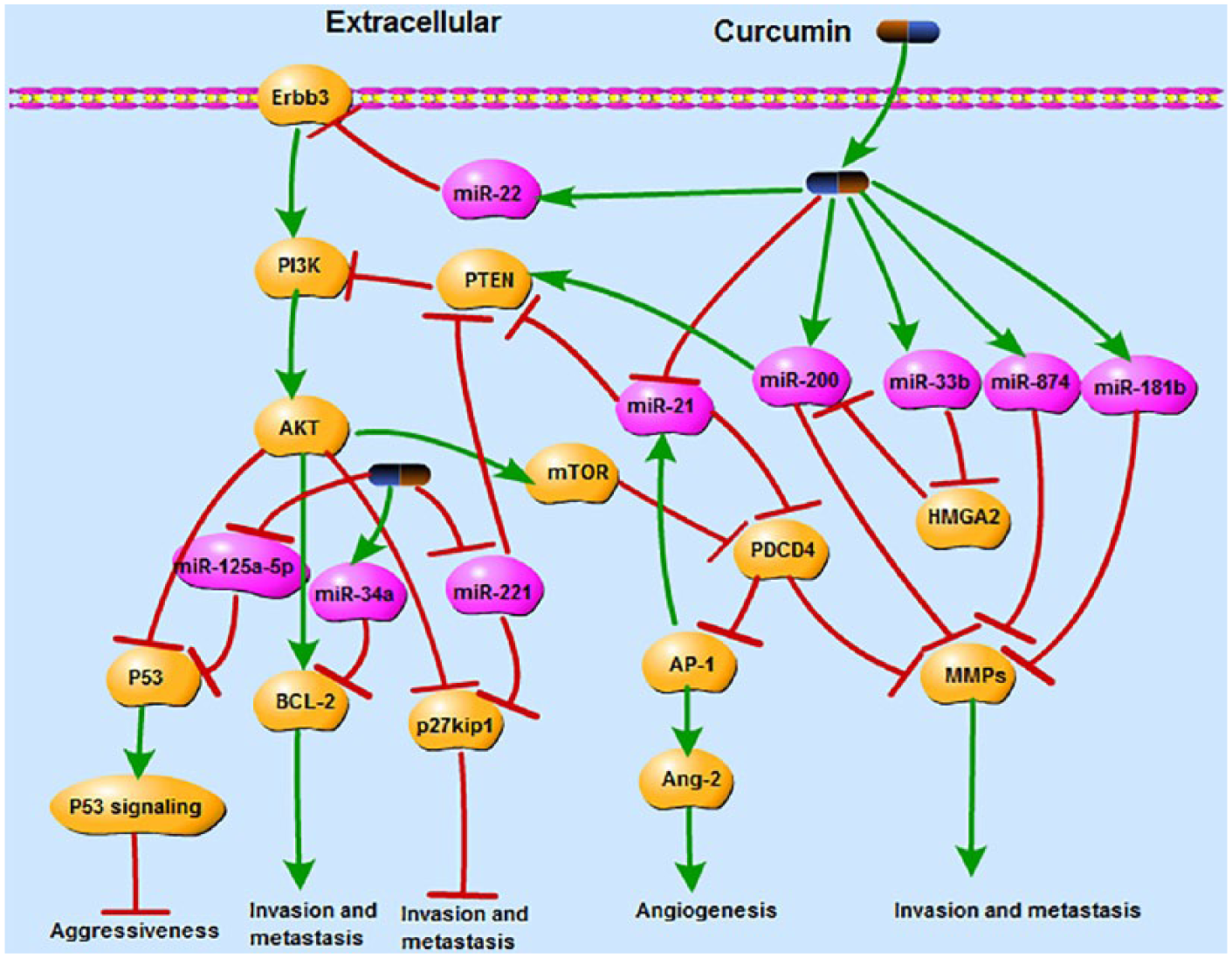
Curcumin significantly activates/inhibits multiple signaling pathways through miRNAs to exert its anti-tumor activities of invasion and metastasis such as PI3K/AKT signaling pathway, PDCD4/MMPs signaling pathway, and PDCD4/AP-1 signaling pathway. A series of miRNAs such as miR-22, miR-125a-5p, miR-34a, miR-221, miR-21, miR-874, miR-181b, and miR-200b/c play important roles in the process of invasion and metastasis by targeting downstream genes such as Erbb3, P53, Bcl-2, p27kip1, AP-1, PDCD4, MMPs, and HMGA2.
Curcumin regulates CSCs via miRNAs
Increasing evidences suggest that cancer cells are driven by a subset of self-renewing cells, termed tumor initiating cells (TICs) or CSCs, which are essential for cancer progression.65,66 CSCs play important role in tumor progression due to their tumorigenic ability, differentiation potential, self-renewal capacity, and resistance to chemotherapy, all of which contribute to cancer metastasis, relapse, chemotherapy resistance, as well as poor prognosis.67,68 Curcumin and its analogs have recently been shown to be effective in inhibiting tumor progression by targeting the CSCs. 69 Additionally, it has reported that miRNAs play important roles in mediating inhibitory effect of curcumin on CSCs self-renewal capacity, tumorigenic ability, and sphere formation.70,71 MiRNAs such as miR-200, let-7, miR-146a, miR-101, and miR-26a are typically lost in pancreatic cancer. Curcumin was reported to decrease formation of pancreatospheres and CSC function through increasing expression of these miRNAs. 31 Curcumin also decreased the formation of pancreatic cancer sphere cells consistent with inhibition of CSC cell surface markers EpCAM and CD44 expression partly through inhibition of miR-21 and increase of miR-200.72,73 Curcumin treatment decreased expression of Notch-1-specific miRNAs (miR-34a, miR-21) and increased tumor suppressor let-7a level to inhibit esophagosphere formation. 74 Curcumin could regulate multiple miRNAs by the CSC self-renewal and stemness abilities. Therefore, combination therapy of curcumin in the future treatment of cancer has important clinical implications for the future cancer treatment. Curcumin increases tumor suppressor miRNAs such as miR-34a, miR-200, miR-203, miR-192-5P, miR-22, miR-33b, miR-874, miR-145, miR-146a, miR-101, and miR-26a or downregulates onco-miRNAs such as miR-21, miR-221, miR-19, miR-208, and miR-125a-5p in cancer cells to regulate cancer biology including cancer cell apoptosis, proliferation, invasion, metastasis, cell cycle, and CSC (Table 2).
Curcumin regulates expressions of miRNAs to influence cancer progression.

Curcumin increases chemotherapy sensitivity via miRNAs
In recent years, chemotherapy is the most effective therapeutic strategy for the treatment of malignant tumors. However, development of chemoresistance has become a major therapeutic obstacle in human cancer treatment, and the mechanisms of drug resistance remain unclear.75,76 Extensive studies have revealed that curcumin sensitized various chemotherapeutic drugs in cancers, including human breast, colon, pancreas, gastric, liver, brain, and hematological malignant. 77 For example, there are studies showing that curcumin helped increase sensitization to cisplatin-mediated killing of ovarian cancer cells and played a critical role in reversing cisplatin resistance in human LA cells which could help patients reach 5 years survival.78,79 Additionally, growing evidences suggested that when combined with some traditional chemotherapy drugs, curcumin could increase chemotherapy sensitivity and reduce chemoresistance. Curcumin in combination with 5-fluorouracil (5-FU), docetaxel, or paclitaxel significantly caused chemotherapy sensitivity, brought out significant biological improvements, and overcame the high resistance to anti-neoplastic agents. 80 That the mechanism of curcumin has synergistic effect of increasing chemotherapy sensitivity or reversing chemotherapy resistance remains unclear. MiRNAs which can be regulated by curcumin have been found to be implicated in cancer therapeutic response, including altering chemoresistance by regulating the expressions of resistance-associated genes. 81 For example, curcumin increased sensitivity of cisplatin to LA cells via downregulating miR-186, which indicated that curcumin could induce inhibitory activity mainly through an miRNA signaling pathway. 82 Moreover, Twist1 (twist family bHLH transcription factor 1) and ABCB1 genes (ATP-binding cassette, sub-family B (MDR/TAP), member 1) were reported to be targeted by miR-186 which may account for overcoming chemoresistance of cisplatin in epithelial ovarian cancer.83,84 Another research reported that curcumin suppressed the growth of resistant colon cancer cells treated with 5-FU and oxaliplatin through downregulating miR-21 and restoring PTEN levels with subsequent reduction in Akt phosphorylation. 85 In addition, further study revealed that the anti-cancer viability of the chemotherapeutic drug 5-FU in colon cancer was enhanced by curcumin due to the suppression of miR-27a. Curcumin downregulated miR-27a with concomitant induction of ZBTB10, a target of miR-27a, which could enhance the therapeutic efficacy in those patients who have developed drug resistance. 86 In CRC, curcumin sensitized 5-FU to overcome chemoresistance via upregulating epithelial to mesenchymal transition (EMT)-suppressive miR-200c which inhibited the expression of ZEB1 and BMI1. 87 In pancreatic cancer, gemcitabine combined with curcumin treatment is much more effective in restraining NF-κB, COX-2, PGE2, and VEGF through modulation of miR-200 and miR-21 expressions, and the inactivation of miR-21 may in turn reactivate PTEN, leading to the inactivation of phosphorylated Akt 88 (Table 3). In the future, more convincing research should be carried out about curcumin increasing chemotherapy sensitivity and reversing chemotherapy resistance through miRNAs pathway which can provide a more effective strategy for cancer treatment.
Curcumin increases chemotherapy sensitivity via modulating miRNAs.

Questions and prospects
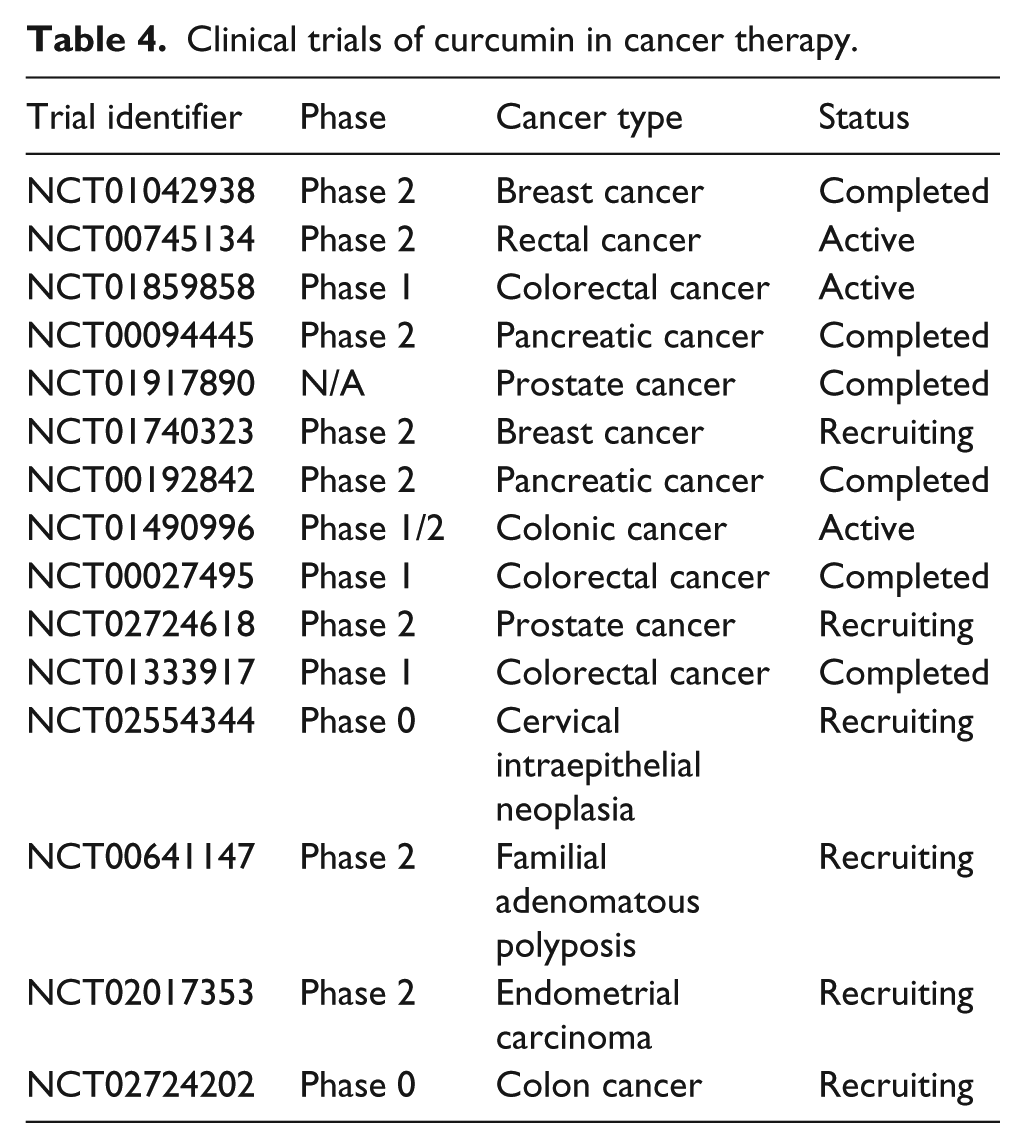
The natural compound curcumin is a cheap, non-toxic, and easily available agent with excellent anti-inflammatory, anti-oxidant, anti-coagulation, anti-hyperlipidemia, anti-atherosclerosis, anti-mutagen, as well as anti-neoplastic clinical effects.7,89 Curcumin exerts anti-cancer effects on tumor progression through modulating the expression of tumor suppressor miRNAs and onco-miRNAs in different cancer cells both in vitro and in vivo. The functional links between miRNAs and their target genes affect tumor progression through tumor-associated pathways in a variety of tumors. Curcumin regulates miRNAs to act as a promising anti-cancer and therapeutic target through the activation of tumor suppressor pathways or inactivation of tumor promotion pathways. 90 Recently, extensive research has revealed that tumor chemotherapy combined with curcumin can increase the curative effect, inhibit or reverse the resistance responses, as well as reduce chemotherapy side effects. There are more than one signaling pathway regulating the transformation and progression of cancers. The use of curcumin is promising because it could target numerous signaling pathways through regulating multiple miRNAs, while the monomodal therapy often targets only a single gene in a signaling pathway which results in typical failure in cancer therapy. Therefore, curcumin could be more efficient on cancer therapy, as it could be used to overcome drug resistance or to improve therapeutic efficacy. A large number of studies have illustrated the anti-cancer role of curcumin, and mechanistic studies have demonstrated that curcumin exerts anti-cancer effects by targeting numerous key signaling pathways involved in cancers through regulating expressions of miRNAs. Because of the promising results from these studies, the efficacies of curcumin in cancer therapies are being investigated in clinical trials (http://www.clinicaltrials.gov/; Table 4). However, there is a major concern that curcumin has poor solubility and limited bioavailability which restrict its clinical application.91,92 Its analogs, such as difluorinated curcumin (CDF), 3,5-bis(2-flurobenzylidene)piperidin-4-one (EF24), as well as curcumin nanoparticle formulation, provide effective ways to deliver curcumin into tumors and have more bioavailability than curcumin. Unlike usual curcumin, new technologies are planned to deliver therapeutic agents especially to tumors and improve anti-cancerous effects, thus avoiding systemic and unpleasant effects.93,94 Besides, most of the point of views above were dated from in vitro and limited in vivo research including some clinical trials. There is a long way to go before curcumin can be utilized in clinical cancer treatment strategy. Further well-designed randomized clinical trials need to be established to verify curcumin as a multi-targeted therapeutic regimen for the management of malignant tumors. In addition, the studies we reviewed above on mechanism of curcumin mediated by miRNAs are only a branch of curcumin’s complex regulated network, and further studies are needed to focus on this valuable natural compound. In the future research, further studies are required to better understand the functional links between curcumin and miRNAs in different types of cancers.
Clinical trials of curcumin in cancer therapy.
Conclusion
In this review, we focused on the inhibitory effect of curcumin on cancer progression by regulating expressions of multiple miRNAs. Curcumin-mediated inhibition or enhancement of miRNAs regulate activation/inactivation of a series of complicated signaling pathways, such as Akt, PTEN, Bcl-2, p53, Notch, and Erbb signaling pathways which participate in cancer biology. Through a clear understanding of the relation between curcumin and miRNAs, we may provide a more effective therapeutic strategy against cancers. It is nice to see that synthesis of curcumin derivatives, use of new drug delivery systems based on nanoscience, and technology of nanoparticles and liposome have improved the basis for the curcumin clinical treatment. 95 Despite limitations of this study, we still believe curcumin could bring a new insight into molecular targeting treatment of malignant tumors.
Footnotes
Acknowledgements
The authors thank Shan-Liang Zhong, MD, for his discussion and help in the revision. Siying Zhou and Sijie Zhang contributed equally to this work.
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Ethical approval
This article does not contain any studies with human participants or animals performed by any of the authors.
Funding
This study was funded by National High Technology Research and Development Program of China (No. 2014AA020604) and the National Natural Science Foundation of China (No. 81272470).