
Abstract
MicroRNAs are small nonprotein-encoding RNAs ranging from 18 to 25 nucleotides in size and regulate multiple biological pathways via directly targeting a variety of associated genes in cancers. MicroRNA-27b is a highly conserved MicroRNA throughout vertebrates and there are two homologs (hsa-miR-27a and hsa-miR-27b) in humans. MicroRNA-27b is an intragenic microRNA located on chromosome 9q22.1 within the C9orf3 gene, clustering with miR-23b and miR-24-1 in human. As a frequently dysregulated microRNA in human cancers, microRNA-27b could function as a tumor suppressor or an oncogenic microRNA. More and more studies indicate that microRNA-27b is involved in affecting various biological processes, such as angiogenesis, proliferation, metastasis, and drug resistance, and thus may act as a promising therapeutic target in human cancers. In this review, we discuss the role of microRNA-27b in detail and offer novel insights into molecular targeting therapy for cancers.
Introduction
Cancer is one of the leading causes of death human diseases. Based on GLOBOCAN estimates, approximately 14.1 million new cancer patients and 8.2 million deaths occurred in 2012 worldwide. Cancer leads to a tremendous burden on society in both developed and developing countries alike, particularly in the latter whose population accounts for 82% of the world. The occurrence of cancer is increasing due to the growing and aging population, as well as established risk factors containing smoking and overweight. 1 To the contrary, thanks to the advances in early diagnose and newly treatment, the number of survivors continues to grow. 2 Therefore, it is essential to search for innovative treatments and biomarkers for cancers.
MicroRNAs (miRNAs), small nonprotein-encoding and ranging from 18- to 25-nucleotide RNAs in size, are predicted to regulate the expression of over 90% genes at the post-transcriptional level, thereby affecting diverse cellular and molecular processes, such as tumorigenesis, tumor growth, apoptosis, cellular migration/invasion, angiogenesis, metabolic pathways, and signal transduction.3–5 MiRNAs modulate messenger RNAs (mRNAs) translation through canonical base pairing between the seed sequence (nucleotides 2–8) of the miRNAs and the complementary seed match sequences of target mRNAs, which are typically located in the 3′ untranslated region (3′UTR). Interaction between miRNAs and mRNAs is not specific, that is, every miRNA can target multiple mRNAs, and correspondingly each mRNA can be regulated by a great deal of miRNAs.6,7 An RNA-induced silencing complex-independent manner by directly binding to DNA on the transcriptional level is also always observed when miRNA regulated mRNAs. 8 More than 50% of known human miRNA host genes are located in fragile chromosomal regions that are susceptible to amplification, deletion, or translocation during the process of tumor progression. 9
MiR-27b is a familiarly dysregulated miRNA in human cancers, for example it is always up-regulated in glioma, 10 cervical cancer, 11 breast cancer (BRC) 12 and down-regulated in lung adenocarcinoma,13,14 prostate cancer (PCa), 15 colorectal cancer (CRC), 16 acute myeloid leukemia (AML), 17 gastric cancer (GC), 18 and bladder cancer (BC). 19 Through the suppression of multiple targets, miR-27b has recently emerged as a key suppressor or an oncogene in cancers. Here, we review the role of miR-27b in human cancers and partly reveal its functions in tumor progression, therapy, prognosis, as well as its prospect of clinical application based on molecular targeting therapeutic strategies.
The miR-23-27-24 cluster
The miR-23-27-24 family contains two paralogs and several members: miR-23a-27a-24-2 (called the miR-23a cluster) on chromosome 19 and miR-23b-27b-24-1 (named the miR-23b cluster) on chromosome 9 in human. While, in mouse, the miR-23a and miR-23b clusters are localized on chromosome 8 and chromosome 13, respectively. 20 In humans, the miR-23a cluster encodes a primary miRNA (pri-miRNA) transcript composes of three miRNAs: hsa-miR-23a, hsa-miR-27a, and hsa-miR-24–2, the miR-23b cluster containing hsa-miR-23b, hsa-miR-27b, and hsa-miR-24–1 analogously. MiR-23a differs from its paralog miR-23b by only one nucleotide near their 3′ ends, so is the miR-27a and miR-27b. However, miR-24-1 and miR-24-2 can yield the same mature miR-24 sequence. 21 Similar evolution and diverse expression patterns are observed among these miRNAs in this cluster which was mainly found and highly conserved in vertebrate species. Members of homologous miRNAs in this cluster might be diverse across animal species, for example, in fishes it is predicted that more homologous miRNAs (such as multiple pre-miRNAs for miR-23a and miR-24), and novel homologous miRNAs (such as miR-27c, 27d, 27e, and 24b) exist. 22 Simultaneous loss of miR-23b-27b-24-1 gene cluster was observed and its encoded genes C9orf3 and Fanconi Anemia Complementation Group C (FANCC) were also decreased in liver and kidney cancers. According to the DNA methylation data in TCGA, the methylation status of the miR-27b gene promoter is increased in kidney cancers while unchanged in liver cancers. Consequently, genetic deletion and DNA methylation may account for the reduced miR-27b level in kidney cancers and only genetic deletion for the reduced miR-27b level in liver cancers. Although miR-27a and miR-27b are forecasted to have the largely identical target genes, the same targets that experimentally validated are less than 15% according to the data in miRTarBase. 23
Regulators of miR-27b expression
MiR-27b is an intragenic miRNA located on chromosome 9q22.1 within the C9orf3 gene, clustering with miR-23b and miR-24-1 in human. Although the mechanism underlying miR-27b dysregulation in human cancer is not yet fully elucidated, much evidence suggests that the regulation of miR-27b is mainly by genomic loss, epigenetic changes (including DNA methylation and histone modification), transcriptional regulation, multiple molecules, and signaling pathways.
Loss of the miR-27b coding gene leading to reduced miR-27b level has been reported in liver and kidney cancers, indicating that genetic deletion plays a significant role in the regulation of miR-27b. 23 It has been proved that promoter hypermethylation of miRNA is very common in the vast majority of tumors. Transcriptions silencing by CpG methylation was reported to play critical roles in the inactivation of tumor-suppressive genes. Yan et al. 24 and Ye et al. 16 have revealed that hypermethylation of CpG islands (chr9: 96887100–96887300) in the miR-27b gene promoter region correlated with attenuated expression of miR-27b in colon cancer cells and in several CRC cell lines, separately. FB1, which is a common mycotoxin contaminant of corn, down-regulates miR-27b in HepG2 cells, probably through inducing chromatin instability, followed by global DNA hypomethylation and histone demethylation. 25 Cytokines have been proved to be tightly associated with the regulation of miR-27b, such as interleukin-6 (IL6), (C-C motif), ligand 18 (CCL18), and IFN-γ. IL6 is a pro-inflammatory cytokine and can activate AKT pathways resulting in DNA methyltransferase 1 (DNMT1) phosphorylation and DNA methylation, which account for the reduced level of miR-27b in CRC. 26 Besides, chemokine CCL18 reduces miR-27b expression in BRC cells and IFN-γ has also been shown to decrease miR-27b expression in liver epithelial cells.27,28 Nuclear factor-kappa B (NF-κB) inflammatory pathways may be quite an indispensable mechanism on the regulation of miR-27b, which has been shown by several reports. Overexpression of miR-223 decreases the level of miR-27b through suppressing IKKα which is a component of the IKK complex that phosphorylate IκBα in C6 astrocytoma cells. IκBα negatively regulates transcriptional activation of NF-κB in the canonical pathway; hence, it is a reasonable conjecture that NF-κB pathway is implicated in the regulation of miR-27b by miR-223. 29 Her2/neu(ERBB2) increases miR-27b expression through the PI3K/AKT/NF-κB signaling cascade. MiR-27b expression is also up-regulated via NF-κB and IKK/IkBα as a result of tumor necrosis factor alpha (TNFα) and epidermal growth factor (EGF) treatment in BRC. Nischarin regulates miR-27b expression through a feedback loop by directly inhibiting NF-κB-p65 phosphorylation. Altogether, above findings suggest that during the progression of BRC, miR-27b becomes up-regulated, followed by down-regulated Nischarin and elevated NF-κB, and miR-27b sustained increasing in turn, a positive feedback loop. 30 In addition to these two mentioned above, Zhou et al. 31 have shown that NF-κB p65 trans-activated miR-27b cluster genes in human biliary epithelial cells. Ionizing Radiation (IR) induced miR-27b down-regulated in a time- and dose-dependent manner in leukemia cell lines CCL119 and TIB152. B-estradiol/ERa signaling may contribute to this appearance through activating the ERK1/2 pathway. Interestingly, miR-27b expression could be elevated by low-dose IR in both female and male leukemia patients at some certain time points, so it is supposed that other molecular mechanisms may be involved. For example, up-regulation of Dicer may be one of these. 32 Furthermore, there are two poly-positive feedback mechanisms reported. Followed by activation of the c-Src/AKT oncogenic pathway, miR-27b is repressed and down-regulated in human colon cancer cells. MiR-27b targets and directly reduces ARFGEF1 and paxillin, thus elevated ARFGEF1 and paxillin activating AKT and c-Src in turn, resulting in AKT/miR-27b/ARFGEF1 and c-Src/miR-27b/paxillin positive feedback circuits 33 (Figure 1).

The figure summarizes how miR-27b expression is regulated. Epigenetic changes such as DNA methylation and multiple signaling pathways including NF-κB, PI3K/AKT, ERa/ERK1/2 pathways are the underlying mechanisms. Some regulated factors such as TNFα, EGF, MIR-223, HER2, and IR are involved in these pathways. There are three feedback circuits in the regulation of miR-27b, namely, AKT/miR-27b/ARFGEF1, c-Src/miR-27b/paxillin, and Nischarin/miR-27b/NF-κB. Besides, cytokines such as IL6, CCL18, and IFN-γ have been proved to be tightly associated with the regulation of miR-27b.
Angiogenesis
Excessive vascular leakage and angiogenesis are the signs of cancer. For tumor progression, angiogenesis is the rate-limiting factor. Tumor angiogenesis plays a critical role for tumor maintenance and growth, and angiogenesis inhibitors could offer an important therapeutic advantage which has been demonstrated by many studies.34,35 It is generally accepted that angiogenesis is a balance between the anti- and proangiogenic genes. This balance could be disturbed by a variety of miRNAs, which suppress multiple target mRNAs and inhibit their translation or transcription. 36 MiR-27b integrates two vascular processes, regression and expansion, which are opposite and controlled by anti- and proangiogenic factors, respectively. Attraction and repulsion are integral elements to establish functional vascular networks. Some studies have confirmed that miR-27b functions as an angiogenesis promoter by targeting multiple antiangiogenic genes. MiR-27b promotes angiogenesis, endothelia tip cell fate, branching (sprouting), as well as appropriate arterial-venous segregation and inhibits endothelial cell (EC) repulsion in vitro. While, in vivo, miR-27b increases the number of perfused blood vessels and contributes to the maintenance of embryonic blood vessel formation.
The growth of new vessels is a process that delicately controlled involving the activation, proliferation, migration, and maturation of ECs. 36 It has been reported that miR-27b is highly expressed in ECs, as well as lung and heart, which are characteristic as highly vascularized tissues, suggesting a potential role in EC function and the vasculature. MiR-27 enhances angiogenesis by promoting EC proliferation and migration via targeting and directly inhibiting Sprouty2 (Spry2), Semaphorin6A (Sema6A), and Semaphorin6D (Sema6D) in response to vascular endothelial growth factor (VEGF). 21 The most important pathway VEGF/VEGFR2 axis activates two downstream Ras/Raf/ERK and PI3K/AKT signaling pathways, both of which act crucial roles in cancer angiogenesis. Moreover, the targets of miR-27b mediated directly are Sprouty2, Sema6A, Sema6D, and Dll4/Notch which act as inhibitors of angiogenesis demonstrated by numerous reports.37,38 Sprouty2 protein belongs to the Sprouty family that encodes multiple proteins including Spry1 to Spry 4 and Sprouty-related domain 1(SPRED1) and SPRED2 and negatively modulate the Ras/Raf/ERK pathway. 39 It has been confirmed that miR-27b targets the 3′UTR of Spry2 mRNA and protein. MiR-27b significantly represses the expression of Spry2; thus, the Ras/Raf/ERK pathway is activated.21,40 The semaphorin family comprises of more than 20 members and a variety of semaphorins can promote or inhibit tumor angiogenesis and development. 41 As members of the semaphorin family, Sema6A and possible Sema6D significantly repressed the VEGF/VEGFR2 axis, two downstream signaling pathways Ras/Raf/ERK and PI3K/AKT are also reduced correspondingly.21,42 MiR-27b significantly increased vascularization and growth of subcutaneous tumors. By increasing micro-vessel density (MVD), miR-27b promotes cancer growth and angiogenesis in lung carcinoma. At least to a certain extent, these effects are due to the reduced expression of Dll4/Notch 43 (Figure 2).
MiR-27b significantly promotes angiogenesis by directly suppressing Sprouty2, Sema6A, Sema6D, and Dll4/Notch which function as inhibitors of angiogenesis. As a result, the RAF/MEK/ERK pathway is activated mediated by Sprouty2, and the VEGF-C/VEGFR2 signaling is activated in response to VEGF through Sema6A and Sema6D, as well as two downstream pathways Ras/Raf/ MEK/ERK and PI3K/AKT. While miR-27b also could function as an inhibitor role via directly targeting VEGF-C/VEGFR2 axis.
In contrast to these studies above, miR-27b is also found to be an inhibitor of angiogenesis. It is universally known that the VEGF families are important mediators of angiogenesis and lymphatic vessel formation that promote angiogenesis and lymphangiogenesis during development of tumor. 44 VEGF-C is identified as a direct target gene of miR-27b in CRC and in GC. Through suppressing the VEGF-C/ VEGFR2 signaling, as well as its two downstream pathways, miR-27b inhibits CRC and GC angiogenesis. Moreover, overexpression of miR-27b inhibits proliferation, migration, and tube formation in human umbilical vein endothelial cells (HUVECs) by affecting the expression of VEGF-C16,45 (Figure 2).
Although the function of miR-27b in angiogenesis seems to be opposite, it is not contradictory to analyze them from the mechanism. MiR-27b represses multiple antiangiogenic genes including Sprouty2, Semaphorin6A (Sema6A), Semaphorin6D (Sema6D), and Dll4/Notch, consequently activating both the Ras/Raf/ERK pathway and the more important VEGF/VEGFR2 pathway; therefore, miR-27b functions as an angiogenesis promoter. In the latter two experiments, miR-27b depresses the VEGF/VEGFR2 pathway, therefore acting as an angiogenesis inhibitor. As a result, the factors that determining whether miR-27b is an angiogenesis promoter or an inhibitor include not only different cell phenotypes but downstream pathways which are more important. On the whole, VEGF/VEGFR2 axis makes great contribution to angiogenesis mediated by miR-27b and may be the most important determinant for angiogenic role of miR-27b.
It has been indicated that the VEGF molecules and their receptors (VEGFRs) are the main targets for the treatment of pathological blood vessels and lymphatic vessels. Furthermore, recent studies suggest that VEGF-C may be a better target than VEGF-A, not only in inhibiting the lymphatic vessel formation but also in terms of angiogenesis. 46 If the antiangiogenic therapy for VEGF is used, the tumor vasculature can be depleted and the self- renewal of the tumor stem cells can be prevented. MiR-27b inhibitors or analogues may be potential new treatments.
MiR-27b affects cell proliferation in tumor
It is universally known that the cell cycle progression is a finely regulated process, which is controlled by both positive and negative cell cycle regulatory molecules, such as cyclins, cyclin-dependent kinases (CDK), and CDK inhibitors. 47 Retinoblastoma (Rb) protein could be phosphorylated in the late G1 phase by the CDK4-CDK6-cyclin D1 complexes and CDK2–cyclin E complexes, which are characterized as G1 phase CDKs-cyclin and then factors that induce the S phase entrance released. 48 Through regulating multiple genes and proteins involved in cell cycle progression, miR-27b has a positive or negative effect on cell proliferation. MiR-27b promotes BRC cells proliferation via accelerating the G1–S phase transition to enhance the cell cycle progression through negative regulation of suppression of tumorigenicity 14 (ST14), a transmembrane serine protease. ST14 down-regulates cyclin E and CDK2 complexes, up-regulates p27Kip1, an important CDK inhibitor, and consequently keeps cells from G1 phase entering into S phase, resulting in excessive cells accumulation in the G1 phase. MiR-27b suppresses ST14 expression and inhibits G1/S arrest of BRC cells. 12 Consistent with this discovery, Jin et al. 30 also showed that by targeting Nischarin, miR-27b induced cell proliferation and anchorage-independent growth of BRC. It is reported that in cervical cancer the overexpression of miR-27b mediates down-regulation of polo-like kinase 2 (PLK2) which belongs to a serine/threonine protease family and is a crucial factor for response to DNA damage and the regulation of cell cycle progression. Thus, the overexpression of miR-27b can promote the proliferation of cervical cancer cells and inhibited cell apoptosis induced by paclitaxel through decreasing the expression of PLK2. 11 Up-regulation of miR-27b in cervical cancer tissues and cells suppresses cadherin 11 (CDH11) and induces the proliferation through promoting cell cycle transition from G 1 to S phase. 49 MiR-27b could control cell arrest in the G0/G1 phase, expedite the progression of cell cycle, and promote glioma cell proliferation. Furthermore, miR-27b increases the activity of downstream molecules in β-catenin/Tcf-4 pathway, including STAT3/c-myc/Cyclin D1, while the direct target is unknown yet. 10 In addition to promoting proliferation, miR-27b is also crucial for proliferation inhibition in multiple tumors. Different from glioma, in GC cell miR-27b-3p suppress oncogene ROR1 and the downstream pathway c-Src/STAT3 signaling, as well as the related proteins p-c-Src, p-STAT3, c-Myc, and cyclin D1. Accordingly, miR-27b-3p decreases the proliferation ability and colony formation of GC cell via inducing G0/G1 phase cell arrest. 18 Similar inhibition effect of miR-27b on GC cell proliferation is found by Geng et al. 50 and the target Frizzled7 (FZD7), an important co-receptor in the WNT signaling pathway may be the underlying mechanism (Figure 3).
MiR-27b regulates cell proliferation via various target genes including Nischarin, FZD7, PPARγ, RET, VEGF-C, ARFGEF1, Sp1, LIMK1, EGFR, and c-Met and then WNT, NF-κB, and Akt signaling pathways participate in the proliferation regulation. In addition, ST14, PLK2, CDH11, ROR1, and cyclin A2 which are associated with cell cycle progression are also the direct targets of miR-27b. ST14 regulates the cyclin E and CDK2 complexes and p27Kip1, and ROR1 influences the downstream proteins containing p-c-Src, p-STAT3, c-Myc and cyclin D1, which ultimately lead to cell cycle progression alter.
Except for molecules associated with cell cycle, there are other mechanisms for miR-27b effect on cell proliferation. MiR-27b reduces the expression of peroxisome proliferator-activated receptor gamma (PPARγ) via targeting its mRNA and protein. Either up-regulated miR-27b or down-regulated PPARγ gene expression could reduce cancer cell growth in vitro and tumor growth in mouse xenografts in neuroblastoma. Reduced expression of oncogene NHE1 which is a downstream protein of PPARγ and decreased NF-κB activity are supposed to contribute to this growth inhibition. 51 6-Gingerol (6G)-induced increased miR-27b suppresses tumor cell proliferation and triggers cell apoptosis of myeloid leukemia through inhibiting PPARγ expression, followed by the suppression of the inflammatory cytokine gene which related to the oncogenic NF-κB pathway. 52 RET is a proto-oncogene that encodes one of the tyrosine kinase receptors which is involved in signal transduction for hemopoietic cell growth and differentiation. In t(8;16) AML, miR-27b negatively regulates RET expression, thus its inhibition of AML cell growth and differentiation could be expected. 17 Wang et al. 32 further confirmed the inhibitory effect of miR-27b on the proliferation of leukemia cell lines CCL119 and TIB152 through inducing cell arrest in the S phase of the cell cycle via suppressing cyclin A2. It has been demonstrated that VEGF-C/VEGFRs axis is a vital inducer of tumor cell proliferation, metastasis, and invasion. In CRC, through decreasing the expression level of VEGF-C, miR-27b significantly inhibits cell proliferation, colony formation, and self-renewal in vitro and tumor growth in vivo. 16 In human colon cancer HCT116 cells, miR-27b down-regulates ARFGEF1 and activates the ARFGEF1/Akt pathway, leading to tumor growth suppression. 33 MiR-27b mediated proliferation, migration, and invasion suppressed was also found in non-small-cell lung cancer (NSCLC) cell lines, the targets including Sp1 and LIMK1, separately.13,14 Decreased epidermal growth factor receptors (EGFRs) and c-Met signaling pathways are responsible for the miR-27b inhibition effect on BC proliferation, invasion, and /or metastasis. EGFR is the cell-surface receptor of the extracellular ligands-EGF family and c-Met is a hepatocyte growth factor (HGF) receptor 19 (Figure 3).
MiR-27b regulates epithelial mesenchymal transition and influences tumor cell migration and invasion
Metastasis, a process that the tumor cells migrate from the primary tumor site to distant organs through the way of blood or lymphatic system, is a vital feature of malignant tumors. The original tumor cells accumulate a series of changes, including genetic and epigenetic aspects, such as the abnormal expression pattern of miRNA to conduct this metastatic cascade. 53
Epithelial mesenchymal transition (EMT) is a process that the epithelial cells lose typical epithelial traits and transform into mesenchymal cells, thereby close cell connections are lost, and enhanced migration capacity is obtained. 54 Through positively or negatively regulating EMT, miR-27b affects the process of migration and invasion in several cancers. HGF activation decreases the miR-27b expression and up-regulates the expression of ST14/matriptase which is an essential enzyme for extracellular matrix (ECM) degradation and a target for miR-27b. MiR-27b significantly inhibits cell invasion of human head and neck squamous carcinoma (HNSCC). Both decreased EMT and enhanced ECM may be the underlying mechanisms. 55 CCL18-mediated miR-27b reduction enhances the EMT, and promotes both MDA-MB-231 and MCF-7 cells migration and invasion, indicating the metastasis suppressor role of miR-27b in BRCs. 27 MiR-27b is observably up-regulated in cervical cancer cells and tissues and evidently triggers migration and invasion of C33A cells via inhibiting CDH11. Mechanically, miR-27b promotes EMT through affecting E-cadherin, vimentin, and N-cadherin, in which CDH11 is involved. 49 Enhanced level of miR-27 is also observed in GC tissues and cells, as well as increased transfer capability of the human gastric cancer AGS cells induced by miR-27. Zhang et al. confirmed tumor suppressor gene adenomatous polyposis coli (APC) as a target of miR-27 and through suppressing APC, Wnt/b-catenin pathway was triggered, resulting in EMT. In addition, miR-27 increased the expression of ZEB1, ZEB2, Slug, and Vimentin which are EMT associated genes and simultaneously decreased the levels of E-cadherin. Loss of E-cadherin contributes to enhanced tumor cell motility and is a sign of the EMT 56 (Figure 4).
EMT plays a significant role in miR-27b mediated invasion and metastasis. CDH11 and APC are two direct targets of miR-27b. MiR-27b promotes EMT via affecting E-cadherin, vimentin, and N-cadherin, in which CDH11 is involved. Wnt/b-catenin pathway is activated by miR-27b through APC, accompanied by increased expression of ZEB1, ZEB2, Slug, and Vimentin and decreased E-cadherin, resulting in EMT. By targeting VEGF-C, miR-27b inhibits lymphangiogenesis through CaMKII, AMPK, and p38 pathways. MiR-27b influencing Nischarin, along with α5 PAK1, cofilin, and LIMK expression is another important mechanism. Besides, Spry2, paxillin, and ST14 are also significant targets of miR-27b for invasion and metastasis.
Except for regulating EMT, miRNA-27b plays a vital role in the transfer via inhibiting multiple genes that are associated with motility and migration at the post-transcriptional level. SPRY2-mediated ERK signaling activation may be another mechanism for miR-27b inducing the invasion and of migration of GC cells proved by Jiang et al. 57 MiR-27b is decreased in metastatic, castration-resistant PCa tissue and cells, and suppresses migration and invasion which are key metastatic processes in PCa cells ALVA31 and PC3-ML. These inhibitory effects on migration and invasion are concomitant with decreased activity of Rac1 without affecting total Rac1 levels and enhanced level of tumor suppressor E-cadherin. Rac1, the Rho GTPase, regulates cytoskeleton rearrangements which is essential for cell migration and metastatic phenotype. The process of invasion and migration may occur when epithelial cell–cell contacts are disrupted and cell adhesion molecules such as E-cadherin are absence, suggesting that E-cadherin function as a metastasis suppressor in cancers. 58 MiR-27b inhibits the formation of focal adhesions and cell motility abilities including invasion and migration in colon cancer HCT116 cells through targeting paxillin and silencing paxillin gene expression accompanied by reduced activation of c-Src signaling. 33 MiR-27b-induced cell invasive/metastatic potential of both glioma and hepatocellular carcinoma (HCC) is directly mediated by Spry2 suppression, thereby the miR-27b/Spry2 axis may as a promising molecular target for glioma and HCC metastasis.59,60 The up-regulated miR-27b in BRC cells alters cell migration and invasion abilities by influencing Nischarin expression and inducing PAK signaling activation. Nischarin is an α5β1 integrin binding partner, inhibiting α5 expression, along with PAK1, cofilin, and LIMK phosphorylation, and accordingly suppressing the metastatic behavior of BRC cells. 30 Tumor cells spreading to the sentinel lymph node represents the first step of metastasis. Lymphangiogenesis, the process that lymph tube comes into being, makes crucial contributions to tumor metastasis development.61,62 The formation of lymphatic vessels can be actively promoted by lymphangiogenic factors secreted by tumors, such as VEGF-C, the best-characterized lymphangiogenic factor. MiR-27b suppresses lymphangiogenesis in chondrosarcoma cells via the VEGF-C/VEGFR-3 axis through CaMKII, AMPK, and p38 pathways 63 (Figure 4).
MiR-27b influences tumorigenesis
Solid tumor is an ecological system composed of tumor cells, non- tumor cells infiltrating around, and nearby molecular proteins. The entire ecosystem is described as a tumor microenvironment, which is an important marker of cancer. It is currently believed that microenvironment significantly contributes to tumorigenesis. 64 Tumorigenesis is a complex process, in which normal cells accumulate gene and apparent genetic changes, as well as malignant transformation, and ultimately obtain some characteristics, such as resistance to apoptosis, independent growth, tissue invasion, and /or transfer. 65 The up-regulation or down-regulation of miR-27b in different types of tumors supports its dual role in the malignant transformation as either an onco-miRNA or a tumor inhibitor. By directly targeting and negative regulation of oncogenes or tumor suppressor genes, miR-27b may act as a tumor inhibitor or a tumor promoter, respectively. Altered expression of miR-27b plays an important role in cancer initiation and progression.
MiR-27b is highly expressed in glioma tissues and cells, and the abnormal expression of miR-27b promotes the growth, inhibits the apoptosis, and induces the invasion of glioma cells, suggesting that miR-27b functions as a tumor promoter in glioma tumorigenesis. 10 RHOH and AIM2 which function as oncogenes are identified as targets of miR-27b in lymphoma, suggesting a probable tumor inhibitory effect of miR-27b. There is a conjecture that the absence of miR-27b may be one of the causes of this lymphoma phenotype. 66 MiR-27 acts as a tumor-suppressive role in NSCLC tumorigenesis through suppressing cells growth, migration, and invasion by targeting and silencing Sp1 which is identified as an oncogene and LIMK1 which is indispensable for malignant transformation.13,14 Restoration of mature miR-27b significantly suppresses BC cell proliferation, migration, and invasion via directly regulating EGFR and c-Met signaling pathways, characteristics of a tumor suppressor. 19 Parallel effects of miR-27b on PCa cell (PC3 and DU145) are also observed by direct regulation of GOLM1. 15 Expression of the miR-27b cluster is frequently decreased in clear cell renal cell carcinoma tissue (p < 0.0001). Restoration of miR-27b markedly inhibits cancer cell proliferation, migration, and invasion. Thus, miR-27b functions as a tumor suppressor, presumably targeting several oncogenes in clear cell renal cell carcinoma cells. 67 Adenoid cystic cancers (ACCs) are the tumors which could arise from both the salivary glands and the breast. The salivary glands ACC (sACC) shows a poor prognosis, while the breast ACC (bACC) has good clinical expectations. Meanwhile, miR-27b is down-regulated in sACC cases and up-regulated in bACC tissues. Tumor suppressive role of miR-27b in ACC may be responsible for this phenomenon. 68 MiR-27b is significantly decreased in the plasma and neoplastic tissue of oral squamous cell carcinoma (OSCC) patients, 69 while up-regulated in the saliva of OSCC patients. 70 This appearance may be due to the dual role of miRNA-27b in cancer initiation and progression. It is supposed that OSCC cells release miRNA-27b into the saliva, resulting in a significant increase in miRNA-27b levels of salivary (Table 1).
Function of miR-27b target genes involved in tumorigenesis and drug therapy.
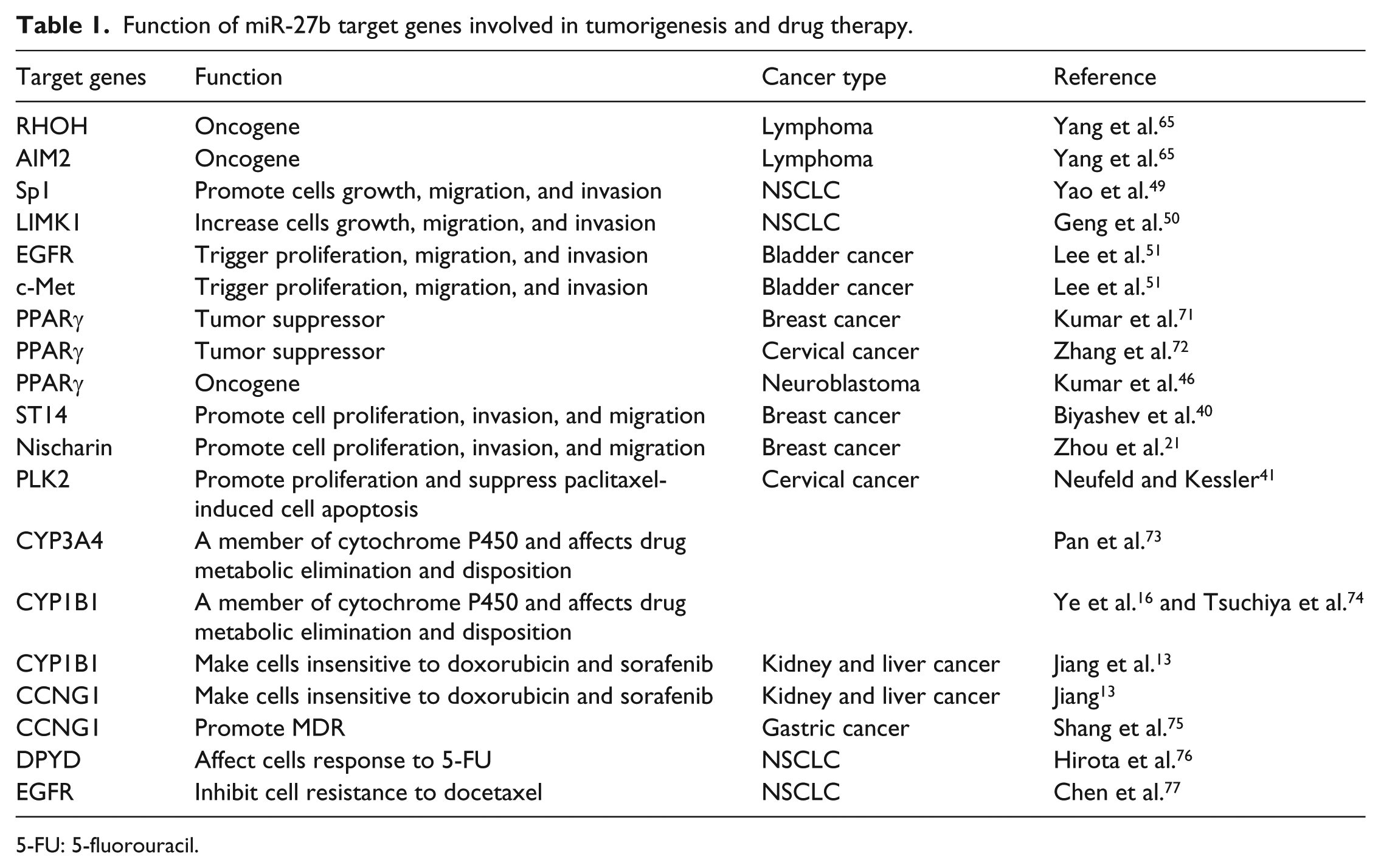
5-FU: 5-fluorouracil.
The role of miR-27b in tumorigenesis may be completely opposite despite via targeting the same gene in diverse cancers. PPAR family which comprises of three isoforms, PPARα, PPARβ/δ, and PPARγ,is vital member of the nuclear receptor superfamily. 78 The PPARγ nuclear receptor pathway plays an important role in cancer, but it appears to act as either tumor suppressor or oncogenic functions in different cancers. NHE1 is an oncogenic factor and a downstream molecule of PPARγ. PPARγ reduces NHE1 expression in breast 71 and cervical cancer, 72 while activates NHE1 expression in neuroblastoma. 51 Hence, PPARγ functions as a tumor suppressor in breast and cervical cancer and an oncogene in neuroblastomas. PPARγ is a direct target of miR-27b and its mRNA and protein expression is inhibited by miR-27b through binding to its 3′UTR. Consequently, miR-27b promotes BRC and cervical cancer tumorigenesis and suppresses tumorigenesis of neuroblastomas. In cancers but diverse sub-types, the function of miR-27b may still be different. Wang et al. 12 and Jin et al. 30 have proved that miR-27b is characteristic as an oncogene through promoting breast cell proliferation, migration, invasion via negative regulation of ST14 and Nischarin, separately. Similarly, miR-27b is up-regulated in human BRC tissue compared to non-cancerous tissues. To the contrary, there is a report that miR-27b acts as a tumor suppressor in Japanese BRC patients. 74 The cause of this phenomenon may be different sample sources: the first two studies from the United States patient samples and the latter from Japanese. This may suggest that the phenotype of miR-27b is different between Europeans and Asians. Similar phenomena occur in cervical cancer. It is well known that human papilloma virus (HPVs) highly correlated with the occurrence of cervical cancer. Consistent with the report by Zhang et al. 72 that HPV16 E7 up-regulates miR-27b to act as a cancer-promoting role, Liu et al. 11 also revealed that HPV16 E7 could increase the generation of miR-27b, promote cell proliferation, and suppress paclitaxel-induced cell apoptosis in cervical cancer via PLK2, a member of PLKs family. It is also accordance with the convention that oncogene HPV induces carcinogenesis through up-regulation of an onco-miRNA. Interestingly, there is a report that E6/E7 silencing increased the expression of miR-27b which is supposed to act as an antitumorigenic role in HPV18-positive HeLa cells. 79 The use of different HPV-positive cervical cancer cells (HPV16-positive CaSki and SiHa cells vs HPV18-positive HeLa cells) may contribute to the difference. There is also no wonder that miR-27b plays a contradictory role in a variety of neoplasms (Table 1).
MiR-27b regulates P450 and influences drug therapy
Although the success rate of cancer treatment has been improved, the intrinsic or acquired resistance is still the primary obstacle to successful treatment and ultimately leads to cancer death. 80 Various mechanisms may be involved in the drug resistance of cancer, among which miRNA plays an important role in the acquisition of drug resistance indicated by more and more studies. As an emerging modulator in cellular pathways, particularly in drug metabolic via regulating the P450, miR-27b makes significant contribution to cancer therapy.
Drug metabolism enzyme-cytochrome P450 participate in the metabolism of endogenous substances and exogenous substances including drugs and then could regulate the detoxification of anticancer drugs. As members of cytochrome P450, CYP1B1,25,74 and CYP3A4 73 expression could be regulated by miR-27b post-transcriptionally. Therefore, miR-27b may play a significant role in drug metabolic elimination and disposition. MiR-27b is identified to make cells sensitive to a variety of anticancer drugs including doxorubicin and sorafenib in the kidney and liver cancer, which have been notorious for resistance to multidrug. Mechanically, miR-27b cooperates with anticancer drugs via both activation of p53-dependent apoptosis through targeting CCNG1 (also called cyclin G1) and suppression of CYP1B1-mediated drug detoxification, which ultimately leading to the two “escape route” for cancer cells greatly cutoff. Adriamycin (ADM) induces tumor cells arrest in S phase and miR-27b promotes cells high sensitivity to doxorubicin, resulting in a more serious replication defect. Combined application of miR-27b and doxorubicin exerts a great synergistic anticancer effect on reduction in hepatocellular cancer cell growth and enhanced cell death than miR-27b or doxorubicin alone. Furthermore, after combination therapy, liver damage is unable to be detected, indicating that combination therapy specifically attacks HCC cells. Sorafenib is the only medicine for systemic late-stage liver cancer treatment approved by Food and Drug Administration (FDA), while to which <5% patients respond. Interestingly, on the basis of TCGA data, about 60%–75% patients with high miR-27b levels show a longer survival period after the use of sorafenib. MiR-27b and anticancer drugs may be applied as a package “cocktail” in order to achieve maximum synergistic effect and minimum side-effect in the future. 23 It was proved that extract and some ingredients of Ginkgo biloba leaves showed anticancer activity; however, the risk of carcinoma was not reduced when often taking Ginkgo biloba leaves. Changes in the expression of miRNA such as miR-27b may affect the metabolism of bilobalide in vivo, resulting in this phenomenon. 81 Dihydropyrimidine dehydrogenase (DPD), the rate-limiting enzyme in the process of pyrimidine catabolism, related to 5-fluorouracil (5-FU) sensitivity closely in cancers. Accumulated 5-FU is rapidly transformed into inactivated metabolites in cancer cells mediated by DPD. Low DPD expression leads to a high risk of the 5-FU toxic accumulation, while increased DPD activity results in decreased 5-FU activity, both of which contribute to the failure in the 5-FU therapy extremely common in NSCLCs. Altogether, appropriate DPD activity is the key to the success of 5-FU treatment. 82 MiR-27b reverses multiple drug resistance (MDR) of GC and promotes apoptosis induced by drugs by directly targeting CCNG1 which is well known to negatively regulate P53 stability. Interestingly, miR-27b up-regulation resulted in increased expression of miR-508-5p, which is mediated by CCNG1 and P53. The two miRNAs synergistically sensitize tumor cells to multiple chemotherapy, and patients accompanied with high expression of both miR-27b and miR-508-5p are more sensitive to chemotherapy. Consequently, the miR-27b/CCNG1/P53/miR-508-5p axis makes important contribution to GC-associated MDR, and restoring the miR-27b and miR-508-5p might reverse MDR of GC in the future clinical practice. 75 DPD gene (DPYD) is confirmed to be a target of miR-27b and regulated by miR-27b at the post-transcriptional level in NSCLCs, so the expression of miR-27b is associated with 5-FU-based chemotherapy tightly. Monitoring the expressions of miR-27b and DPYD in patient tissues of lung cancer, as well as individualizing 5-FU dosages to improve therapeutic efficacy and reduce the risk of toxicity may have the prospect of clinical application in NSCLC. 76 MiR-27b is shown to probably enhance cell viability and promote cell resistance to docetaxel treatment of NSCLC through directly targeting EGFR expression. Therefore, miR-27b may be a promising target for improving the effectiveness of docetaxel 77 (Table 1). Esophageal cancer patients accompanied with high level of miR-27b exert resistance to cisplatin and significantly correlate with shorter survival after treatment with cisplatin. MiR-27b triggers normal fibroblasts converted into cancer-associated fibroblasts in esophageal cancer, resulting in the poor response. 83 MiR-27b is down-regulated in the multidrug resistant Ehrlich Ascites Tumor (EAT) cells, and up-regulated in oxaliplatin resistant colon cancer HCT116 cell lines, suggesting that it may be implicated in chemotherapy resistance in these two cancers.84,85 ADM, vincristine (VCR), carboplatin (CBP), mitomycin C (MMC), 5-FU, and cisplatin are commonly used drugs in the treatment of HCC. Up-regulation of miR-27b expression is observed in the five drug-resistant cell lines. 86
As a biomarker for clinical outcome and prognosis
Late diagnosis, recurrence, and regional lymph node metastasis are the main reasons for the poor prognosis. Circulating miRNAs have been demonstrated to be promising non-invasive biomarkers for detection and prediction of a variety of human cancers. Among these, serum miRNAs are especially ideal biomarkers for prediction, which is readily available, and more convenient to monitor the response to treatment. A miRNA pair comprise of miR-15b and 27b in serum is able to distinguish NSCLC from healthy groups with a sensitivity of 100% (95% confidence interval (CI): 0.93–1.0), specificity of 84% (95% CI: 0.73–0.91), positive predictive value (PPV) of 82%, and negative predictive value (NPV) of 100% (p = 3.70E−25), indicating that the early detection of NSCLC could be achieved by monitoring serum level of the miR-27b and 15b. 87 MiR-27b is significantly decreased in the plasma and neoplastic tissue of OSCC patients compared with the normal group (p = 0.00019), accompanied by higher expression of TPT1 gene, suggesting that miR-27b may be used as an oral cancer early detection biomarker. 69 It has been shown that miR-27b level is remarkably higher and Nischarin is significantly lower in BRC tissues compared with carcinoma adjacent tissues (p < 0.0001). Highly aggressive BRC cells show a high level of miR-27b expression than less invasive cancer cells. Patients with high expression of miR-27b exert an evidently shorter duration of disease-free survival (DFS) and poorer clinical outcomes compared with low miR-27b groups. Therefore, the expression of miR-27b functions as a biomarker for the prognosis and metastasis of BRC. 30 High level of miR-27b in plasma related to shorter progression-free survival (PFS) in metastatic CRC patients treated with 5-Fu/oxaliplatin (95% CI: 1.1–1.8, p < 0.004). 88 Compared with the normal group, the expression levels of miR-27b-3p in the metastasis groups are significantly higher (p = 0.008). Patients with elevated miR-27b-3p level are shown to be associated with higher risk of distant metastasis compared with low miR-27b-3p expression group (p = 0.004). MiR-27b-3p could be as an independent vital predictor for distant metastasis–free (DMF) survival and distant metastasis of triple-negative breast cancer (TNBC) indicated by the multivariate analysis. TNBC patients could be stratified into different prognostic subgroups based on miR-27b-3p level for potentially individualized therapy. 89 Consistent with this, Liu et al. 90 confirmed that miR-27b-3p was positively associated with tumor node metastasis (TNM) stage and negatively with DFS and overall survival (OS) of TNBC, an independent biomarker for prognosis. Up-regulated miR-27b in HCC promotes cell migration and invasion through inhibiting Spry 2 expression. Furthermore, miR-27b is significantly associated with HCC patients TNM stage, vessel invasion, as well as tumor differentiation (p < 0.05). 60 Gastric carcinoma patients with high levels of miR-27b were reported to be more sensitive to chemotherapy and tend to exit longer survival duration and better prognosis than those with lower level. Thus, miR-27b may be a potential marker of the chemotherapy response and a prognostic factor for GC patients. 75 Significantly shorter OS was observed in clear cell renal cell carcinoma patients with lower expression of the miR-27b. Reduced expression of miR-27b increased the risk of disease progression and predicted poor survival. In patients of clear cell renal cell carcinoma with more advanced pathological T stage (T3 or more) and grade 3 tumors, miR-27b levels were significantly down-regulated (p = 0.0233). OS was significantly shorter in the low than in the high miR-27b expression group (p = 0.0253). 67
Conclusion and prospects
This review focused on the role of miR-27b in a broad range of cancers with regard to diverse cancer processes including angiogenesis, proliferation, cell cycle, EMT, invasion, and migration through regulating a variety of genes and pathways. MiR-27b levels are always shown to be down-regulated in CRC, AML, GC, and BC, and up-regulated in glioma, cervical cancer, BRC, and thus lead to the recognition of its dual role as either an onco-miRNA or a tumor inhibitor. The relation between ectopic expression of miR-27b and tumor development suggests that miR-27b is an underlying biomarker for molecular targeted therapy. The multiple factors that are shown to be involved in the regulation of miR-27b further highlight its importance. In addition, miR-27b could regulate cytochrome P450 and then affect detoxification of anticancer drugs. Synergistic effect of miR-27b and multiple anticancer drugs has been demonstrated, thus applying them as a package “cocktail” has the prospect of clinical application. Despite the effective delivery of synthetic therapeutic miRNAs to the target organizations will be a challenge, one miRNA, miR-34a, has already entered the first phase of clinical trials in human liver cancer. With a better understanding of miR-27b and its targets, along with its associated pathways, we can explore a better target for cancer treatment.
Footnotes
Acknowledgements
Li Ding and Jie Ni contributed equally to this work.
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
This work was supported by the National High Technology Research and Development Program of China (no. 2014AA020604), the National Natural Science Foundation of China (no. 81272470), the National Key Clinical Specialist Construction Programs of China (no. 2013[544]), the Major Program of Natural Science Foundation of Jiangsu Province (no. BL2014090), and the Natural Science Foundation of Jiangsu Province (no. BK20151579).