
Abstract
Background
Breast cancer is the most common cancer type in women globally, constituting a substantial portion of all documented female malignancies. It is the second major cause of cancer-associated mortality in women, with over one million new diagnoses each year.
Purpose
The present work evaluated the anti-cancer activity of amarogentin by assessing its growth-inhibition and apoptosis-triggering capabilities against breast cancer cells.
Materials and Methods
The cytotoxic effects of amarogentin at various dosages on MDA-MB-231 cells were evaluated using the MTT test. The incidences of apoptosis in amarogentin-induced cells were evaluated using a dual staining technique. The concentrations of oxidative stress markers and PI3K/AKT/mTOR proteins were evaluated in amarogentin-treated cells utilizing kits.
Results
The current findings demonstrate that the MDA-MB-231 cell viability was diminished after treatment with diverse dosages of amarogentin. The amarogentin treatment significantly enhanced apoptotic incidences in MDA-MB-231 cells, as seen by the dual staining assay results. The amarogentin treatment elevated thiobarbituric acid reactive substances level and subsequently diminished antioxidants in the MDA-MB-231 cells. Furthermore, amarogentin suppressed PI3K/AKT/mTOR proteins in the breast cancer cells.
Conclusion
The current findings indicate that amarogentin treatment significantly reduces cell viability and promotes oxidative stress-dependent apoptosis in breast cancer cells. Therefore, it has the potential to serve as an anti-cancer candidate for breast cancer treatment.
Introduction
Breast cancer continues to be the most prevalent malignancy among women globally, accounting for a significant proportion of all recorded female cancers. Breast cancer is the second most common cause of cancer-associated mortality in women, with over a million new diagnoses occurring each year. The global cases of breast cancer reached 2.3 million new incidences in 2020 alone, contributing to 685,000 mortalities and making it the major cause of cancer-associated disability in women globally (Bray et al., 2024). This underscores its profound public health burden, with breast cancer responsible for 11.6% of all cancer incidences. The disease is defined by the uninhibited growth of aberrant cells within the breast tissue, which can originate in the milk ducts or lobules. These heterogeneous diseases, varying in their biological and molecular characteristics, can also develop in the surrounding breast tissue (Xu et al., 2023). The worldwide burden of breast cancer is projected to intensify, with predictions indicating an upsurge to over 3 million new diagnoses and 1 million mortalities by 2040. This escalating trajectory necessitates comprehensive research into its multifaceted etiology, ranging from genetic predispositions and hormonal influences to environmental exposures and lifestyle factors, to devise more effective prevention and treatment strategies. This extensive burden highlights the critical need for advanced diagnostic and therapeutic approaches to mitigate the disease’s impact and improve patient outcomes (Sha et al., 2024). This global increase in incidence is particularly pronounced in high-income countries, reflecting a strong association with socioeconomic development and globalization. However, the low and low-middle income countries have experienced the most significant upsurge in breast cancer burden, indicating a complex interplay of socioeconomic factors and disease progression (Sun et al., 2024).
Cancer development is intricately linked to the subversion of programmed cell death, a critical homeostatic mechanism. Specifically, the apoptotic dysregulation is a hallmark of nearly all cancer types, enabling uncontrolled cellular proliferation and tumor progression (Goldar et al., 2015). This dysregulation often manifests as an acquired resistance to apoptotic signals, allowing aberrant cells to evade elimination and accumulate. This resistance allows abnormal cells to continuously multiply, forming squamous cells that are characteristic of cancerous growths (Giménez-Bonafé et al., 2009). Furthermore, the evasion of programmed cell death allows cancerous cells to develop resistance to various chemotherapeutic agents, thereby complicating treatment strategies and leading to disease relapse. Therefore, a profound understanding of apoptotic signaling pathways is fundamental for developing targeted therapeutics, particularly inhibitors of anti-apoptotic proteins and promoters of pro-apoptotic proteins (Pistritto et al., 2016).
Current standard therapies for breast cancer, such as surgery, radiotherapy, and chemotherapy, often result in significant adverse effects and financial burdens, prompting the urgent need for more effective and less toxic alternatives. Despite advancements in screening and treatment modalities like radiation therapy, surgical interventions, and various pharmacological treatments, breast cancer continues to pose a substantial health issue worldwide (Mishra et al., 2023). This alarming trend highlights the critical need for safer alternative therapies, particularly given that conventional chemotherapies frequently induce severe systemic toxicities such as fatigue, nausea, and myelosuppression, which can significantly compromise patients’ quality of life. These concerns underscore the pressing need for the development of alternative therapeutic modalities that are not only more efficacious but also possess improved safety profiles (Sharma & Saripilli, 2025). Amarogentin is a bioactive secoiridoid glycoside found in several plants, like Gentiana rigescens, Swertia chirata, and Gentiana lutea. Several previous studies have already been reported that amarogentin ameliorated osteoporosis (Li et al., 2023), protected sepsis-induced brain injury (Song & Zhou, 2022), decreased inflammatory response in atopic dermatitis condition (Zhang et al., 2023), and protected tetrachloride-induced liver fibrosis (Zhang et al., 2017) in animal models. In addition, amarogentin showed anti-cancer activity against various cancers, including colorectal cancer (Wang et al., 2025), liver cancer (Zhang et al., 2020), and gastric cancer (Tan et al., 2022) cells. However, there are no previous reports to claim its anti-cancer activity against breast cancer. Consequently, the present work evaluated the anti-cancer activity of amarogentin by assessing its growth-inhibitory and apoptosis-inducing capabilities against breast cancer cells.
Materials and Methods
Chemicals
Amarogentin, dimethyl sulfoxide (DMSO), Dulbecco’s Modified Eagle Medium (DMEM), and other reagents were procured from Sigma–Aldrich, USA. The test kits for measuring biochemical markers were obtained from Elabscience and MyBioSource, USA, respectively.
Cell Line Collection and Maintenance
Breast cancer MDA-MB-231 cells were purchased commercially from ATCC, USA, and grown in DMEM with FBS (10%) and antibiotics (1%) in a 5% CO2 chamber. The grown cells were obtained after reaching 80% confluency and employed for further tests. The experiments used in this study were approved by the ethical committee of the Affiliated Hospital of Hebei University.
MTT Assay
The influences of amarogentin on the growth of MDA-MB-231 cells were evaluated using the MTT test. The cells were cultured on a 96-well plate and exposed to amarogentin for 24 h at different dosages of 5, 10, 20, 30, 40, and 50 µM, respectively. Following the amarogentin treatment, 20 µL of MTT reagent was added with 100 µL of DMEM in each well and incubated for 4 h. After dissolving formazan deposits in the wells with DMSO (100 µL), the absorbance was measured at 570 nm.
Dual Staining
The dual staining technique was conducted on both untreated and treated breast cancer cells to evaluate the apoptosis level. The MDA-MB-231 cells were grown in a 24-well plate and treated with 30 µM of amarogentin and/or 2 µg of doxorubicin (DOX) for an additional 24 h. After that, mixtures of 100 µg/mL AO/EB stains were applied to the wells for 5 min in a shady place. Lastly, the apoptosis level was evaluated using a fluorescent microscope.
Determination of Oxidative Stress Markers
The untreated and amarogentin-treated cells were collected, and the cell lysate was obtained by lysing the cells with cell lysis buffer. Then, the suspension was centrifuged at 6,000 rpm for 10 min, and subsequently, the resultant supernatant was employed to assess oxidative stress biomarker levels. The concentrations of thiobarbituric acid reactive substances (TBARS), glutathione (GSH), superoxide dismutase (SOD), and catalase (CAT) were assessed in the untreated and amarogentin-treated cells. The tests were done with kits by adhering to the protocols outlined by the manufacturer (Elabscience, USA).
Determination of PI3K/AKT/mTOR Protein Levels
The levels of PI3K, AKT, and mTOR proteins were assessed in the cell lysate of both control and amarogentin-treated MDA-MB-231 cells. The tests were performed in triplicate using assay kits following the manufacturer’s suggested procedures (MyBioSource, USA).
Statistical Analysis
All statistical tests were done using Prism (GraphPad) software. The findings are depicted as mean ± SD of triplicates. The values were evaluated using one-way ANOVA and Tukey’s post hoc test, with p < .05 considered significant.
Results
Effect of Amarogentin on Breast Cancer MDA-MB-231 Cell Viability
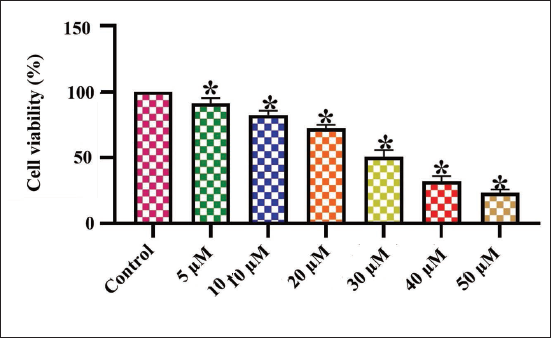
Figure 1 depicts the outcomes of the MTT assay, indicating the cytotoxicity of amarogentin on the MDA-MB-231 cell viability. The administration of amarogentin at different concentrations (5–50 µM) notably diminished the MDA-MB-231 cell viability. The amarogentin treatment demonstrated a substantial reduction in MDA-MB-231 cell growth. Subsequently, the IC50 dosages of amarogentin against MDA-MB-231 cells were established at 30 µM, and this dose was utilized for further studies.
Effect of Amarogentin on the Viability of Breast Cancer MDA-MB-231 Cells. The Treatment with Diverse Concentrations (5–50 µM) of Amarogentin Decreased the Viability of Breast Cancer MDA-MB-231 Cells in a Dose-dependent Manner. The Results Were Illustrated as the Mean ± Standard Deviation (SD) of Triplicates. The Values Underwent One-way Analysis of Variance (ANOVA) and Tukey’s Post Hoc Tests using Prism (GraphPad) Software. * Denotes Statistical Significance at a p < .05 in Comparison to the Control Group.
Effect of Amarogentin on Apoptotic Level in MDA-MB-231 Cells
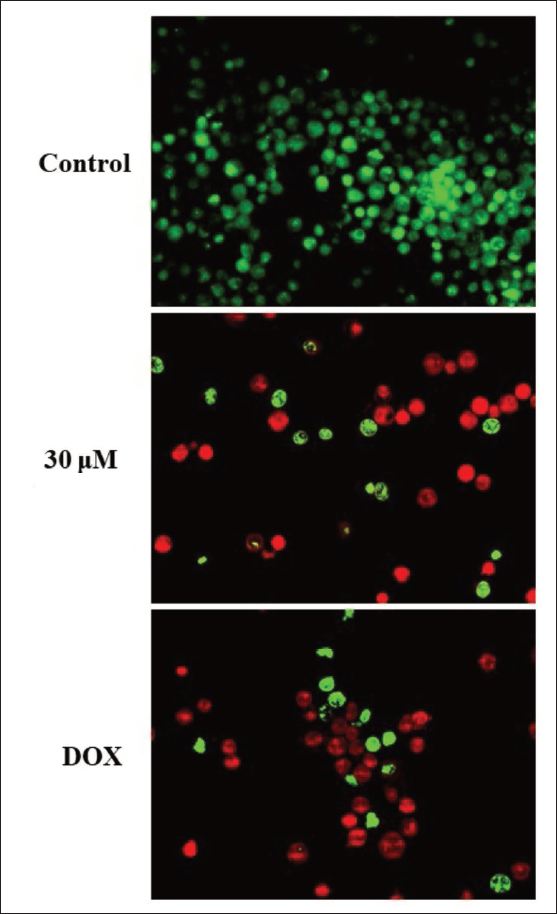
The levels of apoptosis in both untreated and amarogentin-treated MDA-MB-231 cells were evaluated by dual staining assay, with results illustrated in Figure 2. The MDA-MB-231 cells exposed to amarogentin at a concentration of 30 µM displayed intense red fluorescence, indicating a higher incidence of apoptotic cell death. Similar findings were also observed in the DOX treatment, which supports the apoptotic-inducing effects of amarogentin on breast cancer cells.
Effect of Amarogentin on the Apoptotic Cell Death in Breast Cancer, the MDA-MB-231 Cells. The Treatment with 30 µM of Amarogentin Resulted in a Substantial Increase in Intense Red Fluorescence in the Breast Cancer MDA-MB-231 Cells, Which Indicates the Onset of Apoptotic Cell Death.
Effect of Amarogentin on Oxidative Stress Markers in MDA-MB-231 Cells
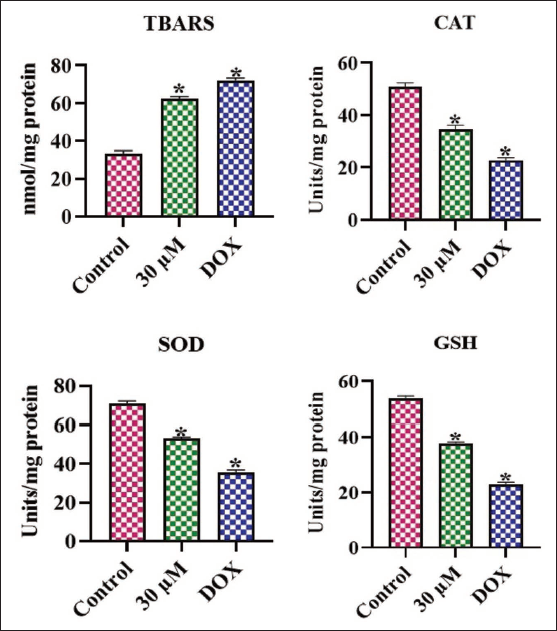
Figure 3 depicts the oxidative stress markers in the untreated and amarogentin-treated cells. The untreated cells revealed reduced TBARS levels alongside increased amounts of SOD, CAT, and GSH. However, the treatment with 30 µM of amarogentin resulted in a notable increase in TBARS and a subsequent diminution in antioxidant concentrations in the MDA-MB-231. These findings demonstrate that amarogentin treatment promotes oxidative stress in the breast cancer cells.
Effect of Amarogentin on the Oxidative Stress Marker Levels in the MDA-MB-231 cells. The Results Were Illustrated as the Mean ± Standard Deviation (SD) of Triplicates. The Values Underwent One-way Analysis of Variance (ANOVA) and Tukey’s Post Hoc Tests using Prism (GraphPad) Software. * Denotes Statistical Significance at a p < .05 in Comparison to the Control Group.
Effect of Amarogentin on PI3K/AKT/mTOR Pathway in MDA-MB-231 Cells
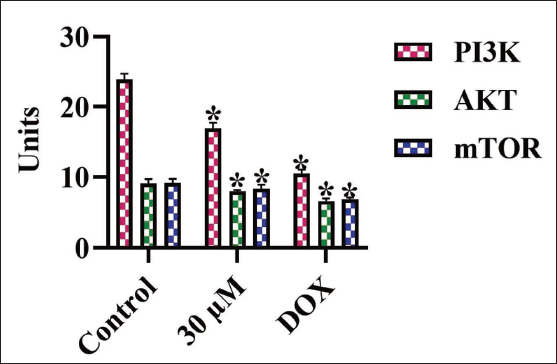
The concentrations of PI3K, AKT, and mTOR proteins in the cells were evaluated, with results illustrated in Figure 4. In untreated cells, the increased PI3K, AKT, and mTOR levels were observed. Whereas the 30 µM of amarogentin treatment markedly reduced the concentrations of these proteins in the MDA-MB-231 cells. Moreover, the DOX treatment also decreased these proteins, which supports the results of amarogentin treatment. These findings suggest that amarogentin may suppress uncontrolled cell proliferation in MDA-MB-231 cells by blocking the PI3K/AKT/mTOR pathway.
Effect of Amarogentin on the PI3K/AKT/mTOR Levels in the MDA-MB-231 Cells. The Results Were Illustrated as the Mean ± Standard Deviation (SD) of Triplicates. The Values Underwent One-way Analysis of Variance (ANOVA) and Tukey’s Post Hoc Tests using Prism (GraphPad) Software. * Denotes Statistical Significance at a p < .05 in Comparison to the Control Group.
Discussion
Breast cancer, a multifaceted disease with significant heterogeneity, continues to pose a substantial global health burden worldwide. Despite advancements in medical technology, including targeted therapies, the breast cancer cases continue to rise worldwide (Siegel et al., 2024). This alarming trend underscores the urgent need for safer and potential therapies to combat the increasing prevalence of the disease. The 5-year survival rate for metastatic breast cancer, for instance, remains below 30% even with aggressive adjuvant chemotherapy, highlighting persistent challenges in advanced disease management (Nolan et al., 2023). This stark reality emphasizes the critical need for novel therapeutic approaches that can overwhelm drug resistance, prevent metastasis, and enhance long-term outcomes for patients. Moreover, the adverse effects related to standard therapies like chemotherapy, radiation, and surgery often lead to debilitating side effects, further diminishing patients’ quality of life and contributing to treatment non-adherence (Giaquinto et al., 2022). Indeed, conventional cancer therapies are frequently associated with adverse effects, including fatigue, nausea, anemia, and emotional distress. These significant challenges underscore the urgent need for the exploration of safer and potentially alternative therapies that can mitigate the severe side effects of current treatments while enhancing patient outcomes and quality of life (Wu et al., 2022).
Apoptosis, a finely regulated physiological mechanism of cell death, is fundamental for regulating tissue homeostasis and eliminating damaged or superfluous cells within multicellular organisms. This intricate process is orchestrated by a complex signaling network involving various protein families, including caspases, Bcl-2 family proteins, and apoptotic protein inhibitors, which collectively govern the cellular fate between survival and programmed demise (Pistritto et al., 2016). Dysregulation of these apoptotic mechanisms can profoundly influence oncogenesis, as cancer cells often acquire strategies to evade programmed cell death, thus enhancing uncontrolled growth and tumor development. Specifically, the inactivation of apoptosis represents a critical dysregulatory event observed across the majority of cancer types, highlighting its central role in oncogenic transformation. This evasion confers a survival advantage, leading to unchecked cellular growth and often participates in the onset of multidrug resistance in various malignancies (Carneiro & El-Deiry, 2020). This capacity of tumor cells to escape apoptosis is now recognized as a hallmark of cancer, enabling their sustained proliferation and contributing to resistance against diverse chemotherapeutic agents. Consequently, understanding the molecular underpinnings of apoptotic deregulation is crucial for developing novel therapies aimed at re-establishing apoptosis in malignant cells (Wu et al., 2019).
The evaluation of novel compounds for their anti-cancer potential critically hinges on their capacity to selectively induce programmed cell death, specifically apoptosis, in malignant cells. This mechanism of action is central to many existing chemotherapeutic and targeted therapies, underscoring its importance in identifying effective anti-tumor agents (Zhou et al., 2017). Distinguishing between true apoptotic induction and mere proliferation arrest or necrotic cell death is paramount for understanding drug mechanisms and predicting clinical efficacy. Therefore, precise and reliable methodologies for quantifying apoptosis in drug-treated cancer cell lines are indispensable for advancing anti-cancer drug discovery (Hu et al., 2020). Apoptosis, a genetically determined elimination process, is a desirable outcome for anti-cancer therapies because it removes cancer cells without inducing inflammation, unlike necrosis. This distinction is crucial, as drug-induced necrosis can lead to systemic inflammation and adverse side effects, whereas apoptosis is a controlled process that minimizes damage to surrounding healthy tissues (Thumpati et al., 2025). Furthermore, the capacity of drugs to selectively trigger apoptosis in tumor cells while sparing non-malignant cells is a key determinant of their therapeutic index, minimizing adverse side effects that often limit the utility of conventional chemotherapies. The intricate mechanisms governing apoptosis involve a cascade of molecular events, making its rigorous analysis an essential step in validating the therapeutic potential of new anti-cancer drug candidates (Song et al., 2022). In this work, the apoptosis-inducing potential of amarogentin was evaluated against breast cancer cells using a dual staining method. The results showed that the amarogentin treatment notably induced apoptosis in MDA-MB-231 cells. Therefore, it was evident that amarogentin can suppress breast cancer development via promoting apoptosis in breast cancer cells.
Oxidative stress is recognized as a pivotal factor in the onset and progression of various cancers. This imbalance, favoring oxidants, disrupts redox signaling and can lead to molecular damage, making it a key mechanism in oncogenesis (Hayes et al., 2020). The relationship between oxidative stress and tumors is complex, often presenting a dual role where reactive oxygen species (ROS) can either promote tumorigenesis or induce cytotoxic effects depending on their concentration and cellular context (Gorrini et al., 2013). Specifically, while low to modest ROS levels can enhance cell growth, genetic instability, and activate pro-tumorigenic signaling, elevated ROS concentrations can trigger anti-tumorigenic responses, leading to oxidative stress-induced cell death. This paradoxical nature positions ROS manipulation as a hopeful therapeutic method for cancer therapy, particularly given the altered redox balance observed in malignant cells compared to their normal counterparts (Akladios et al., 2015). Consequently, strategies designed to selectively elevate ROS levels to a toxic threshold within cancer cells are gaining traction as a promising avenue for inducing apoptotic cell death, thereby exploiting this inherent vulnerability. Indeed, cancer cells often develop adaptive mechanisms to cope with this heightened oxidative stress, including upregulation of antioxidant pathways, yet these adaptations can also render them uniquely vulnerable to therapeutic interventions that further tip the redox balance (Tang et al., 2019).
The induction of oxidative stress-mediated apoptotic cell death in tumor cells represents a promising therapeutic approach, wherein carefully controlled generation of ROS can overwhelm these adaptive mechanisms. This approach targets the delicate redox homeostasis within malignant cells, pushing them beyond their compensatory capacity to a point where uncontrolled oxidative damage initiates programmed cell death. Such strategies exploit the inherent sensitivity of cancer cells to oxidative stimuli due to their already elevated basal ROS levels and often compromised antioxidant defenses (Victorino et al., 2014). This heightened susceptibility allows for therapeutic interventions that specifically aim to increase intracellular ROS above a lethal threshold, thereby triggering apoptosis. The analysis of novel drug compounds in tumor cells involves a comprehensive assessment of lipid peroxidation, as indicated by TBARS levels, alongside the levels of key antioxidants, including SOD, CAT, and GSH (Mishra et al., 2023). Such analyses are critical for elucidating the precise mechanisms by which novel therapeutic compounds modulate cellular redox balance, thereby influencing apoptotic pathways in cancer cells. This mechanistic understanding is crucial for optimizing drug efficacy and minimizing off-target effects, guiding the advancement of more targeted and potent anti-cancer therapies. Specifically, evaluating these biomarkers helps to determine if a drug induces oxidative stress leading to apoptosis (Mishra et al., 2023). In this study, the oxidative stress-inducing potential of amarogentin on breast cancer cells was evaluated. The results showed a considerable increase in TBARS levels and subsequent diminution in SOD, CAT, and GSH in the amarogentin-treated cells. These findings suggest that amarogentin can induce oxidative stress, thereby promoting apoptosis in breast cancer cells.
The PI3K/AKT/mTOR pathway is frequently dysregulated in numerous cancers, including breast cancer, impacting cell growth, survival, and motility. Aberrant activation of this cascade, often initiated by ligand binding to receptor tyrosine kinases, results in the phosphorylation of the PI3K regulatory subunit and subsequent activation of its catalytic p110 subunit. Activated AKT, in turn, phosphorylates various downstream targets, including mTORC1, thereby regulating protein synthesis, proliferation, and metabolism (Khan et al., 2019). Dysregulation of this cascade is a common feature in breast cancer pathogenesis and is intricately linked to treatment resistance, particularly to endocrine and anti-HER2 therapies. Given its central role in tumorigenesis and therapeutic resistance, the PI3K/AKT/mTOR signaling has become a significant focus for targeted drug development in breast cancer. Several inhibitors focusing on the individual components of this signaling have demonstrated promising therapeutic effects in clinical trials, offering novel strategies for improving patient outcomes (Xiao et al., 2021).
The over-activation of the PI3K/AKT/mTOR signaling significantly influences tumorigenesis, aggressiveness, the immune microenvironment, and cellular responses to various therapies. Genetic alterations within this pathway are frequently found in breast cancer and contribute significantly to its progression and resistance to conventional treatments (Wylaź et al., 2023). Constitutive activation of PI3K/AKT/mTOR signaling can arise from various genetic aberrations and deletions or mutations in the tumor suppressor “PTEN”, all of which participate in unchecked cell proliferation and survival. The PI3K enzyme functions as a key upstream regulator of AKT through its intricate control over mTOR complex 2 and phosphatidylinositol (3,4,5)-trisphosphate (PIP3), which are responsible for phosphorylating AKT. The tumor suppressor PTEN is often lost or inactivated, further contributing to this aberrant signaling and leading to more aggressive tumor behavior and treatment resistance (Hu et al., 2021). Consequently, a significant portion of luminal breast cancers exhibit increased PIK3CA mutations and other PI3K/AKT/mTOR pathway alterations, particularly in patients demonstrating unresponsiveness to endocrine therapies. In this context, the advancement of targeted therapies against the PI3K/AKT/mTOR cascade is pivotal to overwhelm the resistance to standard therapies and enhance patient outcomes in breast cancer (Coussy et al., 2020). Encouragingly, the present findings exhibited that the treatment with amarogentin notably decreased the PI3K, AKT, and mTOR proteins in the MDA-MB-231 cells. These findings support the hypothesis that amarogentin can inhibit the progression of breast cancer via down-regulating the PI3K/AKT/mTOR pathway.
In addition to these findings, the present study has several limitations. Firstly, it is an in vitro-only approach, which restricts the generalizability of the findings. Additionally, the study lacks dose-response kinetics, limiting our understanding of amarogentin’s optimal therapeutic window. Furthermore, the absence of in vivo validation and mechanistic studies hinders the translation of these findings to clinical settings. The study also lacks an assessment of amarogentin’s selectivity toward cancer cells. Moreover, the PI3K/AKT/mTOR analysis needs further validation, and mRNA level studies were not conducted. These limitations highlight the need for further studies to comprehensively evaluate amarogentin’s anti-cancer potential. Further studies are needed to address these limitations, and these will be addressed in our future studies, incorporating in vivo models, dose-response kinetics, and mechanistic validation to strengthen the findings and explore amarogentin’s therapeutic potential.
Conclusion
In conclusion, the present findings demonstrate that amarogentin treatment significantly suppresses cell viability and triggers apoptosis in breast cancer cells. The amarogentin treatment also remarkably increased oxidative stress response in MDA-MB-231 cells. Furthermore, the amarogentin treatment also inhibited the PI3K/AKT/mTOR pathway. Consequently, amarogentin has the capacity to serve as an anti-cancer candidate to treat breast cancer. Nevertheless, further studies are necessary to clarify the underlying molecular mechanisms involved in amarogentin-induced apoptosis in breast cancer cells.
Footnotes
Abbreviations
AKT: Protein kinase B; ANOVA: Analysis of variance; CAT: Catalase; DMEM: Dulbecco’s Modified Eagle Medium; DMSO: Dimethyl sulfoxide; DOX: Doxorubicin; FBS: Fetal bovine serum; GSH: Glutathione; IC50: Half-maximal inhibitory concentration; MDA-MB-231 cells: Human breast cancer cell line; mTOR: Mammalian target of rapamycin; MTT: 3-(4,5-Dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide; PI3K: Phosphoinositide 3-kinase; ROS: Reactive oxygen species; SD: Standard deviation; SOD: Superoxide dismutase; TBARS: Thiobarbituric acid reactive substances.
Authors’ Contribution
Minxiang Wei and Yujie Gu: Methodology, experiments, and preparation of original draft.
Cuiyu Han and Yuanyuan Liu: Data validation, review and editing, and conceptualization.
Lu Yan and Rui Gao: Data curation, supervision, conceptualization, and project administration.
Data Availability
Data will be made available on request.
Declaration of Conflicting Interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Ethical Approval
The experiment was approved by the ethical committee of the Affiliated Hospital of Hebei University.
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: The project is supported by the Health Commission of Hebei Province (Grant No. 20210407).
Informed Consent
NA.