
Abstract
In bone tissue engineering, tailored biomaterials mimicking mesenchymal stem cells (MSCs) niche could regulate cell behavior and fate decision. The mechanisms, however, remain obscure. Recently, increasing evidence has shown that non-coding RNAs (ncRNAs) are critical modulators of the mechano-induced MSCs’ responses. Mechanosensitive ncRNAs could convert various physical forces into biochemical signals, and orchestrate signaling networks that regulate the osteogenic differentiation of MSCs in their unique microenvironment. In this review, we focus on the mechanosensitive ncRNAs which could interpret mechanical stimuli during the osteogenesis of MSCs, summarize the signaling pathway networks by which these ncRNAs drive MSCs fate, and point out the limitations and the areas waiting for further exploration.
Introduction
Mesenchymal stem cells (MSCs) are able to differentiate into osteogenic cell lineage, which are promising candidates for regenerative medicine, especially for bone tissue engineering. It is known that MSCs reside in a complex microenvironment, where extracellular biophysical cues play a pivotal role in the regulation of cell behavior and fate decision 1 . Variation of mechanical load makes microenvironment changing continuously, and modulates MSCs function. Additionally, contrary to the single cell, a collective of cells is showed to contact with extracellular matrix (ECM) and the neighboring cells, and extracellular stiffness could mediate the cell-cell communication by changes in gap junction tunnels 2 . Cells could perceive the substrate elasticity and the surface patterns varying from 10 nm to 100 μm 3,4 . Specific topography and stiffness of substrate are suggested to initiate the intracellular biochemical factors to maintain the undifferentiated state of MSCs, 5 or drive MSCs to osteogenic, chondrogenic, or adipogenic lineages, which might be directed by cell shape and cytoskeletal tension 6 –8 . The interaction between MSCs and substrate is one of the most critical aspects of bone tissue engineering. Tailored biomaterials with specific topographical features, including grooves, ridges, pits, and pillars, could mimic the topographic landscape of the MSCs niche 9 . It is suggested that micro- and nano-grooved patterns could impact the osteogenic differentiation 10,11 . Polyisocyanopeptide-based three-dimensional (3D) matrices could mimic the stiffness of adult MSCs niche and show biopolymer-like stress stiffening. It could direct the fates of MSCs from adipogenesis to osteogenesis by simply altering the material’s polymer length 12 . Osteogenic differentiation under MSCs-substrate interaction is tightly modulated by multiple signaling molecules which can stimulate osteoblast-specific transcription factors. It is reported that several signaling pathways, such as bone morphogenetic proteins (BMPs) signaling, are coupled to cellular structures or mechano-sensitive proteins directly 13 . For example, localization in the structures which participate in sensation and modification of cell shape indicate that BMPs could sense and transform mechanical cues into cell fate changes 14 . The regulatory networks at the post-transcriptional level, however, are poorly characterized.
Non-coding RNAs (ncRNAs) profiling analysis and marker genes expression provide new insights into the modulation network of the mechano-induced MSCs’ responses, including adhesion, proliferation, differentiation, and apoptosis 15 . The classification of ncRNAs is based on their length. Keeping the cutoff at the length of 200 nucleotide (nt), these ncRNAs shorter than 200 nt are termed as short ncRNAs, including microRNAs (miRNAs), small interfering Ribonucleic Acids (siRNAs), piwi-interacting RNAs (piRNAs), small nucleolar RNAs (snoRNAs), small nuclear RNAs (snRNAs), and tRNA-derived fragments (tRFs). Among these short ncRNAs, miRNAs are one of the “star molecules,” which could recognize the conserved miRNA responsive elements (MREs) within mRNAs 16 . The Argonaute-2 (AGO2) protein complex could identify the miRNA-MRE pairs, contributing to the destabilization of mRNAs and the downregulation of protein expression 17 . MiRNAs are thus considered as the negative regulators of the expression of genes related to the extracellular factors. It is reported that 122 AGO2-associated miRNA families are identified to target 73 mRNAs, which could encode cytoskeletal, contractile, adhesive, and extracellular matrix proteins 18 . In addition, those ncRNAs longer than 200 nt are designated as long ncRNAs (lncRNAs). LncRNAs are showed to sponge the miRNAs to cross-talk with miRNAs and regulate their expression levels, which are considered as another important modulators of gene expression.
In this review, we focus on the mechanosensitive ncRNAs which convert various physical forces into biochemical signals during the osteogenesis of MSCs, summarize the signaling pathway networks by which these ncRNAs drive MSCs fates, and point out the limitations and the areas waiting for further exploration.
Mitogen-Activated Protein Kinase (MAPK) Signaling Pathway
Various extracellular stimuli could activate MAPK signaling pathway to regulate cell proliferation and differentiation 19 . It is demonstrated that MAPK signaling pathway plays a pivotal role in the regulation of many aspects of osteoblast differentiation 20 .
Electrospun poly (l-lactic acid) (PLLA) membranes with a random arrangement of nanofibrous could mimic the collagen distribution of ECM and regulate the proliferation and differentiation of stem cells 21 . It is reported that PLLA nanofibers could promote the osteogenic differentiation of human bone marrow-derived stem cells (hBMSCs), suggesting that topographical cues in the microenvironment could regulate the functions and fates of MSCs 22 . MiR-193a-3p is sensitive to characteristics of substrate topography, and the expression of miR-193a-3p is downregulated during the topographical feature-induced process of osteogenic differentiation. It is showed that inhibition of miR-193a-3p could promote osteogenic differentiation only in the absence of chemical-factors-induced osteogenic medium and antagomiR-193a-3p-loaded PLLA nanofiber scaffolds could enhance the regeneration of critical-sized bone defects markedly. Loss- and gain-of-function experiments demonstrate that miR-193a-3p could bind to MAP3k3 to regulate MAPK signaling pathway 22 . It presents a strong genetic evidence of direct mechanotransduction function of ncRNAs, which could be an effective tool to direct MSCs fates and differentiation commitment in bone tissue engineering.
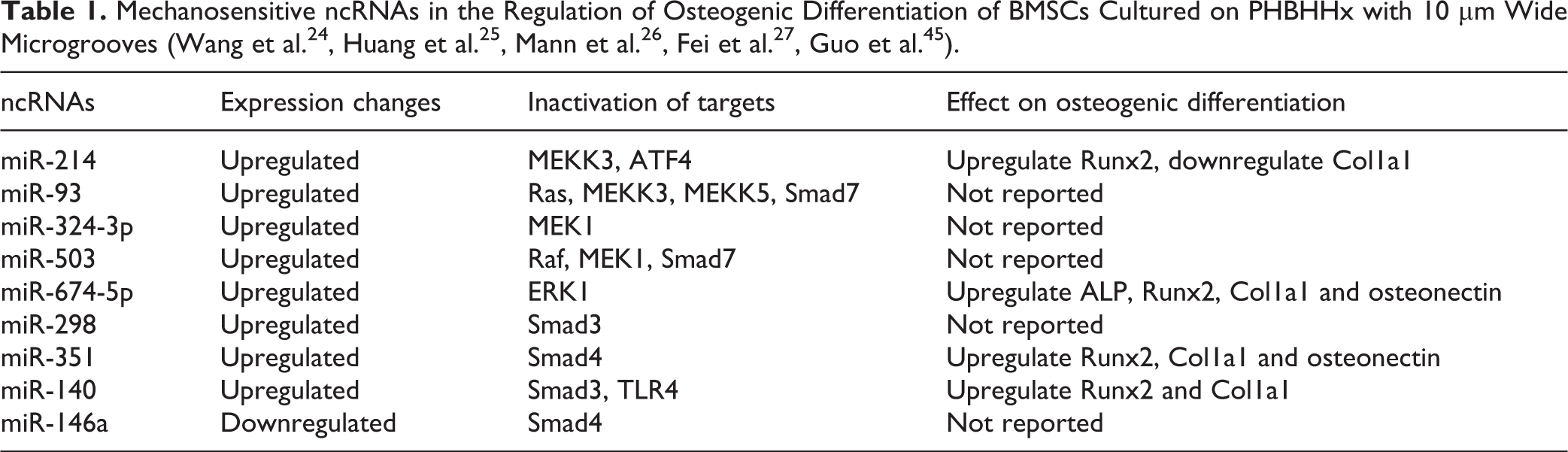
MAPK could mediate the phosphorylation of RSK2 and then enhance the activity of activate transcription factor 4 (ATF4), which is a pivotal mediator of procollagen gene transcription at the later stages of osteoblast differentiation 23 . ATF4 is demonstrated to be the direct target of miR-214 which is an inhibitor of osteogenesis. It is suggested that miR-214 plays vital roles in the modulation of osteogenic differentiation of MSCs cultured on micro-grooved surface pattern 24 . It is showed that the suppression of miR-214 could stimulate the expression of lncRNA MALAT1 and ATF4 significantly, while overexpression of lncRNA MALAT1 remarkably decreases the expression level of miR-214 but upregulates the expression of ATF4. Dual luciferase reporter assay further confirms the binding between MALAT1 and miR-214, and miR-214 and ATF4 25 . Therefore, lncRNA MALAT1 might sponge miR-214 to regulate ATF4 and enhance osteogenesis in steroid-induced avascular necrosis of the femoral head. Whether lncRNA-MALAT1/miR-214/ATF4 axis functions as a regulator in mechano-induced osteogenesis, however, remains to be elucidated.
It is reported that miR-210, miR-296, miR-324-5p, miR-320, miR-324-3p, miR-503, miR-423, miR-500, miR-674-5p, and miR-93 are upregulated in the rat BMSCs cultured on poly(3-hydroxybutyrate-co-3-hydroxyhexanoate) (PHBHHx) with 10 µm wide microgrooves, while miR-196a, let-7a, and let-7i are downregulated 24 . MiR-93 is predicted to target Ras, MEKK3, and MEKK5. MiR-503 is predicted to target Raf and MEK1, while MEK1 is the target of miR-324-3p 24,26,27 . Therefore, these mechano-sensitive miRNAs could influence different steps of Ras-Raf-MEK-ERK-cascade to regulate osteogenesis of BMSCs. Additionally, ERK1 is predicted to be the target of miR-674-5p 24 . ERK1/2 is phosphorylated significantly at the onset of strain 28 , and thus ERK1/2 is suggested as one kind of mechano-sensing protein. Additionally, it is reported that ERK1/2 is a downstream effector of focal adhesion kinase (FAK) signaling 29 , whose activity is related to the expression level of osteogenic markers in human adipose-derived stem cells (hASCs) 30 . Therefore, miR-674-5p might target ERK1 to affect ERK1/2, which orchestrated MAPK signal and FAK signal to regulate the osteogenic differentiation of MSCs.
BMPs Signaling Pathway
PLLA nanofiber alignment is reported to be efficiently committed to osteogenic differentiation 31,32 . A significantly higher expression level of lncRNA H19 is observed on aligned PLLA than random PLLA after osteogenic differentiation of 14 days and 21 days respectively. MicroRNAs is also showed to express differentially between random and aligned nanofibers. The expression levels of miR-30c, miR-24b, and miR-15b are significantly increased on aligned PLLA nanofibers compared with random ones during osteogenic differentiation of MSCs. MiR-15b could target Smad7 and SMAD specific E3 ubiquitin protein ligase 1 (Smurf1) which are known as inhibitors of BMPs signaling 32,33 .
Smad3, Smad4, Smad8 and bone morphogenetic protein receptor type 2 (BMPR2) are positive regulators of BMPs signaling. The smooth PHBHHx substrates are showed to initiate chondrogenesis of MSCs without any induction medium, while the PHBHHx films with 10 µm wide grooves/ridges and 10 µm depths could enhance osteogenic differentiation of MSCs 24,34 . It is reported that miR140, miR-298, and miR-351 are upregulated in the rat BMSCs cultured on PHBHHx with 10 µm wide microgrooves, while miR-146a is downregulated 24 . It is showed that miR-140 and miR-298 might target Smad3 while miR-351 and miR-146a might target Smad4 potentially to regulate osteogenesis 24 . Therefore, these miRNAs might interact with diverse target genes to contribute to the behaviors of MSCs, and the complex and precise regulatory network is an interesting issue to be studied. MiR-153 is identified as a mechanosensitive miRNA, which regulates osteoblast differentiation of hMSCs by directly targeting BMPR2 35 . MiR-125b and miR-20a are significantly downregulated on aligned PLLA nanofibers compared with random ones during osteogenic differentiation of MSCs 32 . It is supposed that miR-20a could target Smad8 and BMPR2 to suppress osteogenic differentiation, while miR-125b inhibits osteogenic differentiation of MSCs through downregulation of BMPR2, Smad4, and osterix. LncRNA MEG3 is predicted to have four binding sites of miR-125b. Meanwhile, lncRNA MEG3 is upregulated while miR-125b is downregulated during osteogenesis, suggesting lncRNA MEG3 might be regulated by miR-125b and microRNA-triggered lncRNA decay mechanism (Fig. 1) 32 . However, more experiments are needed, such as loss- and gain-of-function experiments, to confirm this conclusion.
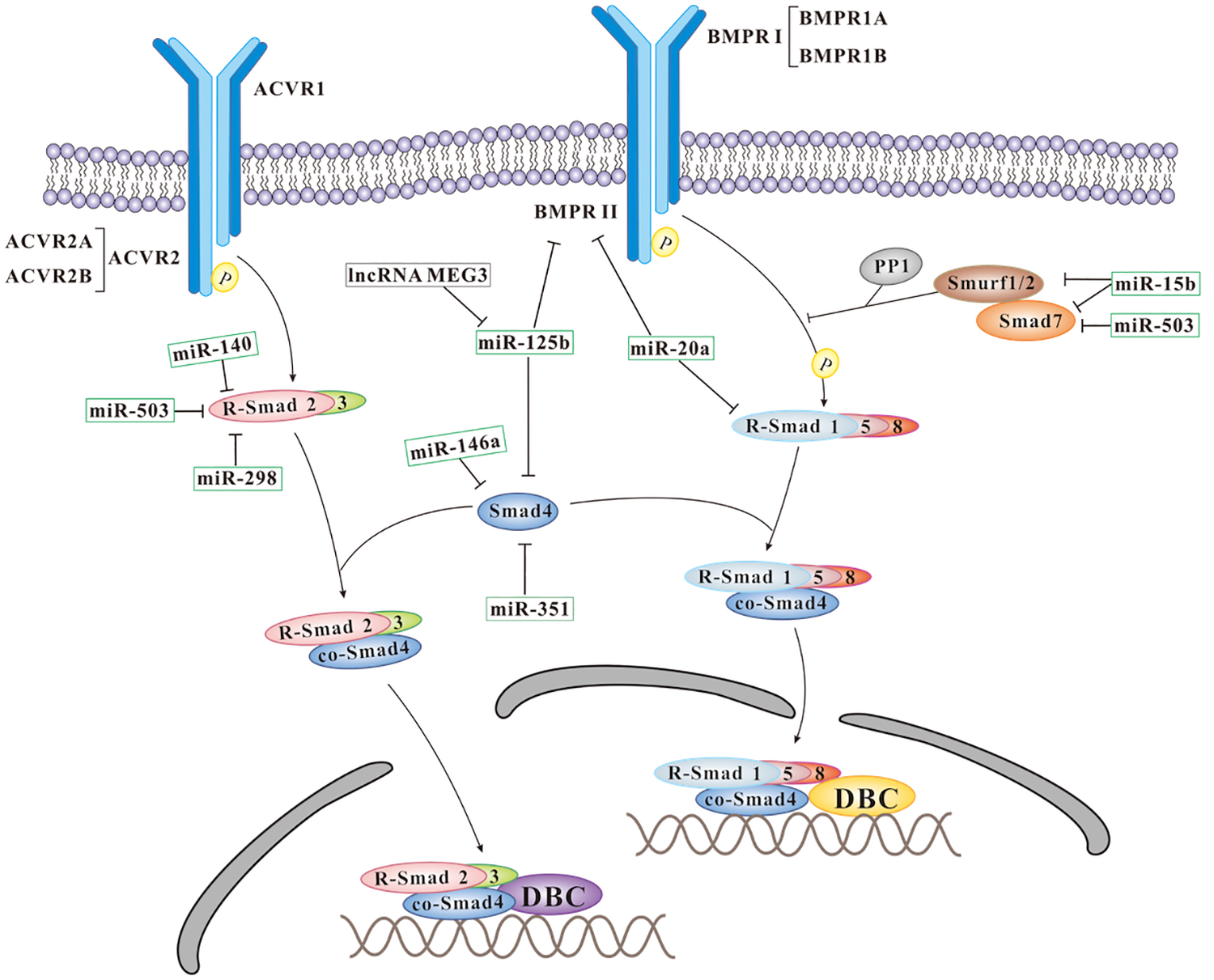
Schematic representation of the mechanosensitive ncRNAs influencing BMPs signals during the osteogenesis of MSCs. ACVR: Activin receptor; DBC: DNA binding co-factors; PP1: protein phosphatase 1; Smurf: Smad ubiquitination regulating factor; R-Samd: receptor activated Smads; Co-Smad: co-mediator Smad.
Mammalian Target of Rapamycin (mTOR) Signaling Pathway
It is reported that mTOR complex 1 (mTORC1) and mTOR complex 2 (mTORC2) are two structurally and functionally distinct protein complexes of mTOR. MTORC1 could activate p70 ribosomal protein S6 kinase (P70S6 K) to positively regulate the expression of the adipogenic gene PPARγ and negatively regulate the expression of the osteogenic marker gene Runx2, and thus is pivotal for initiation and maintenance of adipogenesis 36 . MTORC2 is reported to regulate Ras homolog family member A (RhoA) and cytoskeletal remodeling 37 . RhoA is known to promote actin polymerization to function as an important mechanotransducer of mechanical stimuli, and RhoA-mediated cytoskeletal contractility played essential roles in the proliferation and differentiation of MSCs 38 .
Substrate stiffness is a critical cue in directing MSCs proliferation and differentiation. It has proven that soft substrates enhance adipogenesis while stiff substrates favor differentiation into osteoblasts 39 . It is reported that miR-100-5p and miR-143-3p are upregulated in hMSCs cultured on stiff substrates, while both of them are downregulated in hMSCs cultured on soft substrates 40 , which suggest both of them are mechanically-sensitive miRNAs. Interestingly, genetic modified hMSCs encapsulated in photo-crosslinkable gelatin-PEG hydrogels, which highly expressed miR-100-5p and miR-143-3p, could increase the amounts of hydroxyapatite accumulation greatly 40 . This demonstrates the regulation of these two mechanosensitive miRNAs could generate the mature osteoblast population in injectable hydrogels. This finding is really important because it shows modulation of mechanosensitive miRNAs could drive MSCs fate in 3D gelatin hydrogels, which is necessary to apply biomaterial systems successfully in the clinic to regenerate functional bone mass.
MiR-100 has been demonstrated to suppress the expression of mTOR through targeting its 3’UTR directly in a post-transcriptional manner to modulate proliferation and migration of human esophageal squamous cell carcinoma cell lines 41 . It is reported that miR-100-5p also affects mTORC1 and mTORC2 to regulate mTOR signaling pathway and drive MSCs differentiation to osteogenesis in 3D soft hydrogels significantly in vitro 40 . MiR-143 is also linked to the regulation of mTOR signaling pathway, as it could inhibit the phosphorylation of mTOR and Akt 42 . It is revealed that miR-143-3p might influence the expression of Larp1 and Rictor to exert effects on mTORC1 and mTORC2 40 . Therefore, miR-100-5p and 143-3p could impact the activation of mTOR to regulate mechanotransductive signaling and enhance osteogenic differentiation of hMSCs (Fig. 2). The relative contributions of these two mechanosensitive miRNAs to mTORC1 and mTORC2, however, remains a significant topic for future investigation.
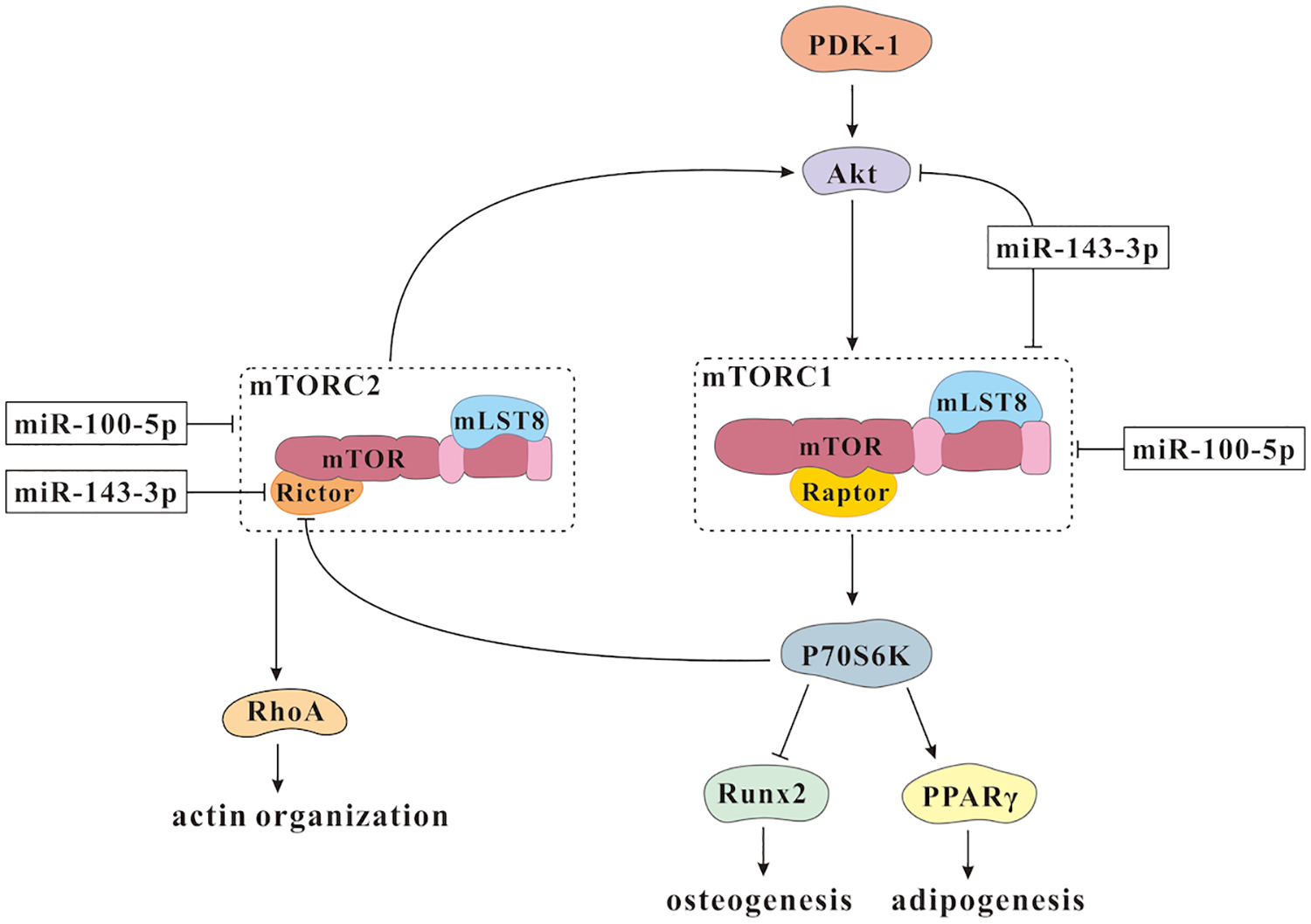
Schematic representation of the mechanosensitive miRNAs affecting mTOR signaling pathway during the osteogenesis of MSCs.
Toll-Like Receptors (TLRs) Signalling Pathway
The microgrooves on PHBHHx with 10 µm wide grooves/ridges and 10 µm depths could trigger the changes of MSCs interfacial behaviors 24 . Osteogenesis-related markers, including cbfa1, col1a1, and bmp2, are upregulated in the MSCs cultured on the microgrooved PHBHHx, while adipogenesis-related genes and myogenesis-related genes are downregulated 24 . MiR-140 is revealed to express differently between the MSCs cultured on microgrooved PHBHHx and those cultured on smooth PHBHHx 24 . Interestingly, lncRNA MEG3 is reported to target miR-140-5p and inhibit adipogenesis and promote osteogenesis of human adipose-derived stem cells 43 . Moreover, TLR4, which could inhibit osteogenesis and enhance adipogenesis of hPDLSCs, is demonstrated to be the direct target gene of miR-140-5p 44 . When lentiviruses-mediated knockdown of miR-140-5p, TLR4 is upregulated significantly, and these genetically modified ASCs combine with scaffold could significantly promote bone formation and fracture-healing in the atrophic nonunion rat model 45 . However, it needs to be further studied whether lncRNA MEG3 could sponge miR-140-5p to regulate the expression of TLR4 to direct MSCs interfacial behaviors on the microgrooved PHBHHx. It might provide important clues for the roles of ceRNAs in the regulation of the topography triggered osteogenesis and the development of biomedical materials.
YAP/TAZ Signaling Pathway
Yes-associated protein (YAP) is necessary for cancer-associated fibroblasts to enhance matrix stiffening, angiogenesis, and cancer cell invasion. Responsively, matrix stiffness further promotes YAP activation, and thus it establishes a feed-forward self-reinforcing loop to maintain the phenotype of cancer-associated fibroblasts 46 . Connective tissue growth factor (CTGF) is a target gene of YAP, which could be induced by mechanical stimulation and play roles in osteogenic differentiation of mechanical stretched-associated ASCs 47 . Transcriptional coactivator with PDZ-binding motif (TAZ) is the transcriptional coactivator of YAP in regulating the MSCs differentiation balance between adipogenesis and osteogenesis.
YAP/TAZ is now widely recognized as mechanosensors and mechanotransducers. Meanwhile, YAP/TAZ serves as the intracellular mechanical rheostat, which could store complex information from past physical environments and influence cells fate 48 . It is reported that YAP/TAZ could directly bind with the TEA domain transcription factor 2 (TEAD2) family that locate in the nucleus, and function as the co-activator to modulate cell proliferation, neovascularization, bone differentiation, and cell fate decisions. In addition, YAP/TAZ could regulate Runx2 and signal transducer and activator of transcription factor 3 (STAT3), which are crucial for bone homeostasis. ECM stiffness could localize YAP and TAZ transcription factors to the nucleus and induce the positive modulation of targets to regulate cell responses, while nanotopography has no effect on the YAP/TAZ complex 15 .
The miR-33a family, including miR-33a-3p and miR-33a-5p, are reported to be possible regulators of YAP/TAZ during hMSCs osteoblast differentiation 49 . The presence of epidermal growth factor receptor (EGFR) inhibitor is able to upregulate miR-33a-3p, and miR-33a-3p inhibition is able to induce a dramatically increase of YAP and TAZ in terms of both mRNAs and proteins. MSCs modified with miR-33a-3p inhibitor and treated with EGFR inhibitor, however, display a block of YAP and TAZ modulation 49 . These results indicate that miR-33a-3p might crosstalk with EGFR signaling to regulate the expression of YAP/TAZ during hMSCs osteogenesis. As for the roles of miR-33a-5p in the osteogenesis, it has demonstrated that overexpression of miR-33a-5p could stimulate the osteoblasts activity and induce a relative increase of osteoid formation in the osteopenia mouse model 50 . The gain and loss of function studies show that either miR-33a-5p overexpression or inhibition has no influence on the expression of YAP/TAZ in hMSCs, while the expression level of TAZ mRNA is increased in pre-osteoblast cells transfected with miR-33a-5p mimics, suggesting miR-33a-5p could upregulate TAZ probably by blocking a certain TAZ inhibitor factor when MSCs acquire a pre-osteoblast phenotype 48 . The specific mechanism, which orchestrates the responses of different cell lineages to mechanical stimuli, deserves more well-designed research.
Conclusions
In recent years, a vast body of work has addressed and unveiled the importance of mechanical force in the skeletal development, function, and repair. Mechanosensitive ncRNAs could response to physical forces and convert them into biochemical signals, which govern osteogenesis of MSCs through various signaling pathways (Table 1 to Table 3). Still there are some areas waiting for further exploration. For example, many kinds of ncRNA, such as circRNAs and lncRNAs, are known as critical modulators in osteogenesis, whereas most studies focus on the roles of mechanosensitive miRNAs. Additionally, most studies only give an insight on the influence of the ncRNAs in vitro, and in vivo studies are needed to provide better support to show the clinical relevance of these modulators of cell behavior. It is also very interesting to study the effect of mechanosensitive ncRNAs in the bone disease mimicking models, which might decipher the code of molecular pathomechanisms and result in the translation of basic biological findings into clinical applications.


In conclusion, it is with undoubted significance to dig out specific mechanosensitive ncRNA signaling networks that regulate the functions of MSCs in their unique microenvironment, thus boosting the processes of bone tissue repair and providing a novel avenue to treat skeletal disorders.
Footnotes
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Ethical Approval
This study was approved by our institutional review board.
Statement of Human and Animal Rights
This article does not contain any studies with human or animal subjects.
Statement of Informed Consent
There are no subject in this article and informed consent is not applicable.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported by National Natural Science Foundation of China (81771048;81900981), China Postdoctoral Science Foundation (2019M663530), Sichuan Science and Technology Program (2020YFS0170;2021YJ0149), Research and Development Foundation of West China Hospital of Stomatology (RD-02-201904), and Research Funding from West China School/Hospital of Stomatology Sichuan University (RCDWJS2020-18).