
Abstract
Asherman syndrome (AS) has an adverse effect on reproductive health and fertility by affecting endometrial regeneration. Stem cell-based therapies hold promise for future use in activating non-functional endometrium and reconstructing the endometrium in vivo. It has been postulated that various endometrial stem cells (EnSCs) are responsible for endometrial regeneration. Numerous studies have focused on bone marrow-derived stem cells (BMDSCs), which may provide new ideas for repairing endometrial lesions and reconstructing the endometrium. Other sources of stem cells, such as menstrual blood, umbilical cord, and amniotic membrane, have also attracted much attention as candidates for transplantation in AS. This review discusses the features and specific biomarkers among four types of resident endometrial stem cells, applications of four different sources of exogenous stem cells in AS, and development of stem cell therapy using biomaterials and exosomes.
Keywords
Introduction of Asherman Syndrome
Asherman syndrome (AS), a severe intrauterine adhesion (IUA), is the complete obliteration of the endometrium with fibrous tissue 1 that often occurs after various gynecological surgeries or pregnancy complications, as a result of damage to the endometrium 2 .
The incidence of AS and IUA among women ranges between 2% and 22%, and its rate is influenced by the number of abortions performed, the incidence of genital tuberculosis, and the criteria used to detect IUA 3–5 .
Patients with mild IUA may present with menstrual abnormalities, including irregular bleeding, hypomenorrhea, or amenorrhea. Severe adhesion can lead to infertility, miscarriage, and poor implantation5,6 . The remaining endometrium becomes atrophic because of poor uterine perfusion 7 . The gold standard for diagnosis and treatment is hysteroscopy 5 .
Certain adjunctive therapies after surgery, such as intrauterine contraceptive devices, anti-adhesive barriers, low-dose aspirin, and periodic hormone treatments have also been applied for preventing the reformation of adhesion and for restoring the normal endometrium 5 . Besides, an integrated approach combining various mentioned measures was proposed to restore the endometrium and achieve better fertility outcomes 8 . However, none of these therapies have proven clinical effectiveness and satisfactory pregnancy outcomes 9 . In order to prevent recurrence and prompt tissue regeneration, it is necessary to uncover the underlying pathology and molecular mechanisms of AS and investigate more effective and precise treatments. IUA therapy is thought to inhibit adhesion formation, promote endometrial regeneration, and improve pregnancy outcomes. Studies have revealed that severe injury to the basal layer of the endometrium leads to a decrease in the number of resident endometrial stem/progenitor cells 10 –12 . Therefore, most studies have focused on stem cell-based therapy to treat damaged endometrium and restore fertility.
Stem Cell-Related Pathogenesis of AS
The underlying pathology of AS remains largely unknown. From a histological point of view, biopsies of AS patients show decreased densities of epithelia and glands and replacement of stroma by fibrous tissue in both endometrial and myometrial layers. The epithelium becomes non-responsive to hormonal stimulation, leading to the absence of functional endometrium 4,13 .
Continuous menstrual cycles through the female reproductive life suggest remarkable regenerative capacity of the human endometrium, which may be associated with stem/progenitor cells residing in the endometrium. Current evidence suggests that resident endometrial stem/progenitor cells may be damaged because of deep trauma involving the basalis and myometrium. Insufficient resident stem cells with limited capacity finally lead to the failure of functionalis regeneration 3,14,15 . On one hand, potential vascular occlusion after various forms of damage may prevent the replenishment of hematopoietic stem cells and tissue repair-related molecules from the bone marrow (BM) via the circulatory system and further result in dysfunctional stem cell niches, which are located perivascularly in the stroma, as the communication between the niches and stem cells is regulated by paracrine factors and cellular signaling pathways 16 . On the other hand, the downregulation of vascular endothelial growth factor (VEGF) in patients with AS impairs angiogenesis, which also requires the participation of endothelial progenitor cells released from the bone marrow 11,17,18 .
Reconstructing Endometrium by Activating Endometrial Stem/Progenitor Cells
It has been widely demonstrated that human endometrium has both endogenous and exogenous sources of stem cells, which are indispensable to its remarkable regeneration capability 19 . The endogenous stem/progenitor cells are derived from the tissue-resident reserve and are crucial in restoring the uterine cavity and recovering endometrial function. Chan et al. isolated endometrial stem cells from endometrial tissue in 2004 20 . Subsequently, numerous studies have been carried out on the identification and application of EnSCs 11,21,22 . The EnSCs include several types of cells, such as the endometrial mesenchymal stem cells (eMSCs), epithelial progenitor cells (EPCs), endometrial side populations (ESPs), and endometrial regenerative cells (ERCs), that are responsible for endometrial regeneration (Table 1). These cells can be identified by their functional attributes, such as clonogenicity, long-term culturing capability, multi-lineage differentiation potential, expression of stem cell markers, and most accurately, by verifying their capacity for in vivo endometrium reconstruction in xenograft animal models 23 –27 . Great efforts have been made to investigate specific surface markers of these EnSCs for the purpose of isolation and targeting certain cells as cell-based therapies.
Resources and Features of Stem Cells which Contribute to Endometrium Regeneration.

eMSC, endometrial mesenchymal stem cell; EPC, epithelial progenitor cell; ESP, endometrial side populations; ERC, endometrial regenerative cells; BMDSC, Bone marrow-derived stem cell; UC-MSC, Umbilical cord-derived mesenchymal stem cell; menSC, menstrual blood-derived stem cell; HAMSC, human amnion-derived mesenchymal stem cell.
Endometrial Mesenchymal Stem Cells (eMSCs)
The eMSCs in the endometrium were detected by the co-expression of CD146 and platelet-derived growth factor receptor β (PDGF-Rβ), called CD140b in 2007 28 . Located on the outer layer of small vessels in both the functionalis and basalis, the eMSCs act as perivascular cells or pericytes that promote endometrial regeneration, maturation, and hemostasis 36,37 . Human endometrial CD146+CD140+ cells have gene profiles similar to those of menstrual blood-derived MSC-like cells 38 and other types of MSCs 37 in immunomodulatory potential and low immunogenicity.
Another significant cell surface marker, sushi domain containing-2 (SUSD2), also known as W5C5, can aid in detecting clonogenic MSCs from the endometrium 23,39 . In vivo endometrium reconstruction shows that human endometrial stromal cells are primarily composed of stromal fibroblasts and blood vessels, both of which are derived from W5C5+ cells 24,37,40 . The W5C5+ cells may also act as a stem cell niche for endometrial progenitor cells 23 . In accordance with clonogenicity, higher colony forming units (CFUs) could be observed in magnetic bead-selected W5C5+ cells, rather than in flow cytometry-sorted W5C5+ cells 23 .
Recently, Trapero et al. identified NTPDase2 as a marker of the endometrial stromal basal layer, which is expressed by the SUSD2+ endometrial MSC, rendering it useful in improving the isolation of MSCs needed for regenerative medicine therapies 41 .
Epithelial Progenitor Cells (EPCs)
EPCs are observed in the basalis in the human endometrium and are not normally shed during menstruation 20,42 . They are characterized as large, single-cell-derived epithelial CFUs with high proliferative potential, and can be differentiated into large gland-like structures in a three-dimensional (3D) cell culture 43 . The 3D cultures contain abundant EPC niche factors in the stromal feeder layer, which can promote differentiation and morphogenesis.
Previous studies suggest that stage-specific embryonic antigen 1+ (SSEA-1 or CD15) and nuclear SOX9+ epithelial cells are capable of generating gland-like structures in vitro 44,45 . SSEA-1 expression is the strongest in the glandular epithelium of endometrial tissue in menstruating and postmenopausal women. In comparison to SSEA-1- epithelial cells, SSEA-1+ cells show greater telomerase activity with longer telomeres and lower proliferation rates, which are the properties of progenitor cells 44 . It is hypothesized that the human endometrium may contain multiple epithelial progenitor cell pools. The basalis is SSEA-1++SOX9++LGR5+, which provides a cellular basis for massive regeneration of the functional layer after menstruation or childbirth. Another cell with a different phenotype known as LGR5++SSEA-1+SOX9+, may facilitate embryo implantation and replenish the luminal epithelial cells that are likely to be shed every day 46 .
Recently, it has been shown that N-cadherin serves as a specific EPC marker. The EPCs associated with N-cadherin can differentiate into large gland-like structures with cytokeratin+ epithelial lumen in vitro 29 . N-cadherin+ EPCs are located on the endometrial gland bases at the endometrial/myometrial interface 29 . After analyzing the spatial relationship of N-cadherin+ EPC and SSEA-1+ EPC, obvious epithelial differentiation can be observed among the epithelia in the endometrial glands 29 (Fig. 1).
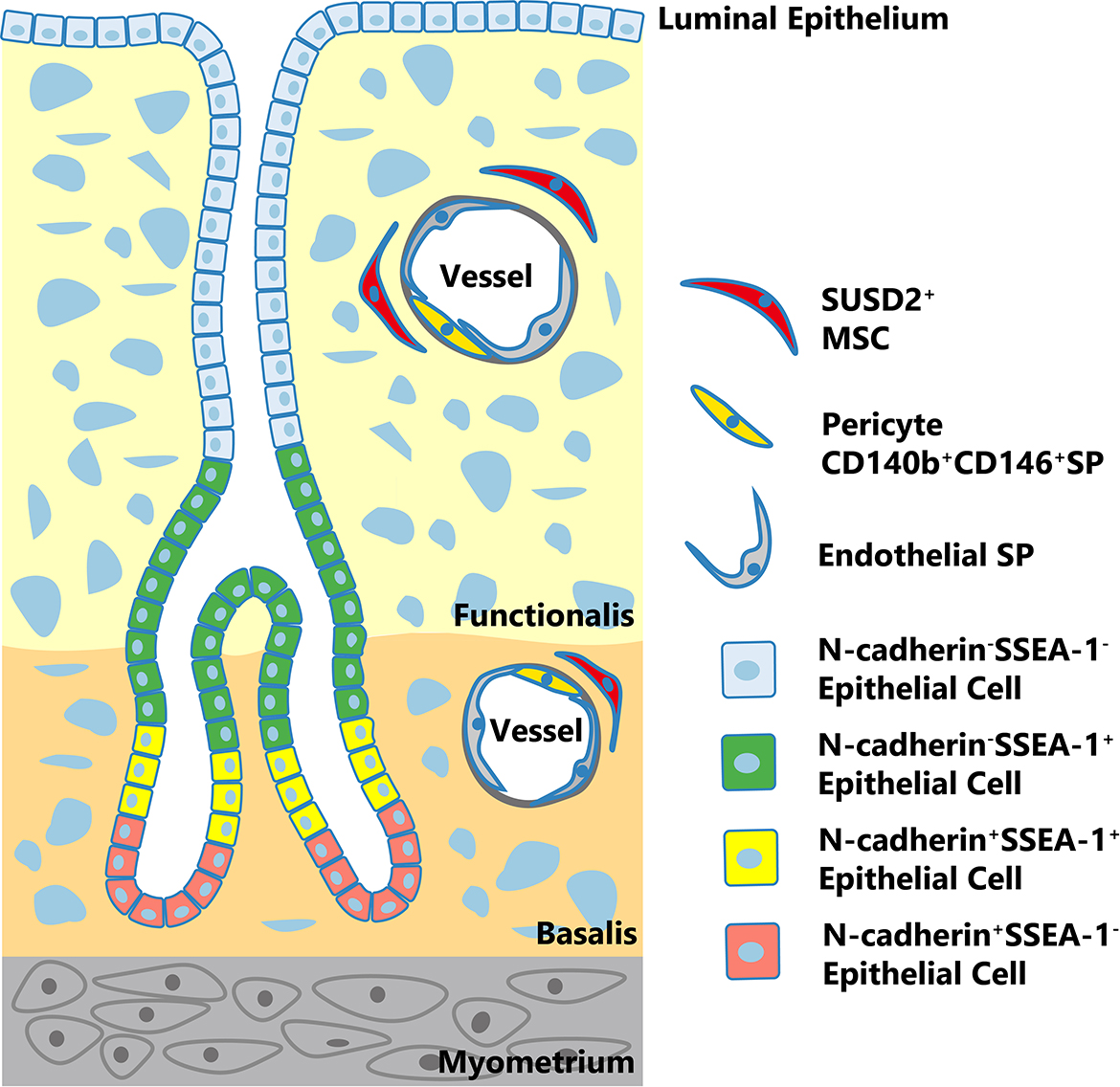
Localization of human resident endometrial stem/progenitor cells. SUSD2+ MSCs are located perivascularly and CD140b+CD146+ cells are mostly pericytes in vessels in both basalis and functionalis. ESP cells are also components of blood vessels. N-cadherin+ epithelial progenitor cells localize to endometrial gland bases in the basalis adjacent to the myometrium. There may be an epithelial cell hierarchy in the human endometrium. N-cadherin+SSEA-1-cells (red) may be the most primitive, which may give rise to double-positive cells (yellow) located closer to the functionalis, which in turn generates N-cadherin-SSEA-1+ cells (green) lining the glands at the boundary of the basalis and functionalis. Double-negative cells (blue) comprise the luminal epithelium.
Endometrial Side Population (ESP)
The ESP has been studied thoroughly in vitro and in vivo, and has the most references among all endometrial stem/progenitor cell candidates 30 . It was identified using dual-color flow cytometry with the efflux DNA-binding dye, Hoechst 33342 47 . The ESP accounts for 1.68% to 2.7% of the epithelial part and 0.4% to 3.1% of the stromal part 24,47 and higher percentages are observed in proliferative and menstrual stages 48,49 . Most of the freshly isolated human ESP cells are in the G0 phase of the cell cycle and show little clonogenic growth and intermediate levels of telomerase; on the contrary, cultured ESPs are in G1 and G1/M/S phases and show raised clonogenicity 47,48 . In a study by Masuda, immunostaining with the ESP marker ABCG2 revealed their co-localization with CD31 in the capillaries and small vessel walls in both the functionalis and basalis, further confirming their endothelial phenotypes 24 . However, human ESP is a mixed population, consisting mainly of endothelial cells(CD31+, 51%), as well as epithelial (CD326+, 27%) and stromal SP cells(CD10+ or CD140b+, 10-14%) 26 .
The ESP cells have a high clonogenic activity with a phenotype similar to that of the MSCs. Interestingly, there are four different types of tissue that may differentiate in vivo with different dominant structures or cell components, including glandular, vascular endothelial, migrating endothelial, and stromal type 24,25,30 . However, the efficiency of endometrium reconstruction remains fairly low due to the lack of a niche, which may be provided by the endometrium main population (EMP, unable to excrete Hoechst, significantly fewer CD31+ endothelial cells than ESP). Even though EMP alone is incapable of differentiation into the endometrium tissue, the coculture of EMP and ESP shows a high rate of success with a mimicked uterine microenvironment 24,26,30 . Another experiment using a xenograft mouse model also suggested that human SP is a heterogeneous population with the optimum ability to regenerate endometrial-like tissue. In contrast, in the absence of niche-like cells, SUSD2+ and ICAM1+ cells alone have the lowest functional regeneration capacity 50 .
Endometrial Regenerative Cells (ERCs)
The ERCs refer to multipotent MSCs and stromal fibroblasts isolated from menstrual blood, with high proliferative potential 21 . The ERCs are localized in the functional layer and basalis layer around the vasculature and are shed every menstrual cycle, providing an alternative source in a non-invasive manner for research in regenerative medicine 31 . Cultured ERCs express telomerase reverse transcriptase and the markers of embryonic stem cells (Oct4++, SSEA+, Nanog+, CXCR4+) but not specific BM MSC marker, STRO-1 51 . The ERCs have a wide differentiation potential for producing all mesodermal lineages, including skeletal, cardiomyocytes, and neural lineages 52,53 . When transplanted into immunocompetent mice, ERCs have low immunogenicity and may have immunomodulatory properties 38 . With such a high proliferation rate, stable genetic characteristics, and remarkable pluripotency, these new type of stem cells may exhibit therapeutic properties beyond expectation 34 .
Activated EnSCs in AS Cases
The presence of stem cells in the human endometrium implies that these cells may be activated in the presence of dysfunctional or atrophic endometrium. Indeed, cytokines essential for endometrial proliferation and angiogenesis, including VEGF and basic fibroblast growth factor (bFGF), were proven to be effective when injected or delivered by collagen scaffold or other biomaterials in AS patients and prompted tissue regeneration before the application of stem cell-based therapy 54,55 . Concurrent curette may also activate resting adult stem cells and initiate cellular renewal, fulfilling their role in recovering tissue homeostasis by releasing tissue repair-related cytokines, thereby generating a thicker endometrium that gives rise to prompted rates of pregnancy and live birth in patients undergoing IVF 56,57 .
The human endometrium is the only source of tissue-derived MSCs that can be separated with minimal invasiveness without the need for anesthetics. EnSC-specific markers facilitate the isolation and purification of MSCs using magnetic bead or flow cytometry sorting, and provide insights for further identification and application in cell-based therapies. It is expected that future studies will clarify the relationship between these stem cells in the endometrial stem/progenitor cell hierarchy in detail.
Reconstructing Endometrium by Replenishing Exogenous Cells and Organs
Until now, the most promising treatment for AS has been stem cell transplantation. There have been many reports about the therapeutic effects of MSC transplantation to AS in animals and humans. The MSCs extracted from the adipose tissue, bone marrow, placenta, and umbilical cords are characterized by their non-immunogenic, angiogenic, antifibrotic, antiapoptotic, and anti-inflammatory properties (Table 1). Various paracrine signaling molecules, including cytokines, chemokines, and growth factors secreted from MSCs, are required for tissue repair 58,59 .
Bone Marrow-Derived Stem Cells (BMDSCs)
Adult BM is a reservoir for several types of stem cells, including hematopoietic stem cells, MSCs, and progenitor cells. Numerous studies on humans and mice have shown that a small number of BMDSCs are integrated into the endometrium and transdifferentiate into epithelial, stromal, and endothelial cells, remodeling the endometrium.
The MSCs are defined by their plastic adherence, clonogenicity, and multilineage differentiation into mesodermal lineages in vitro. They can be distinguished from hematopoietic stem cells by their surface phenotypes (CD73+, CD90+, CD105+, CD34-, CD45-, and HLA-DR-) 59 .
Donor-derived endometrial epithelial cells and stromal cells were detected in endometrial sample of a patient with leukemia who had received bone marrow from a single-HLA antigen mismatched donor for the first time in 2004 60 , suggesting that BMDSCs are responsible for endometrial injury repair and tissue regeneration 10 . Subsequently, this phenomenon was observed in a mouse allogeneic transplantation model, showing that BMDSCs can be implanted in the uterus and positioned in the endometrium when transplanted through the tail vein or intrauterine injection 32 .
Numerous animal studies have focused on the underlying therapeutic effects of BMDSCs in AS. It has been demonstrated that BMDSCs can be recruited to the endometrium in response to damage, and pregnancy rates of AS mice increase substantially after BMDSC transplant, compared to the control group 10 . Another study suggested that BMDSCs might play a crucial role in the reconstruction of damaged endometrium and have great potential for improving endometrial receptivity, which may be due to the expression of endometrial receptivity marker, leukemia inhibitory factor (LIF). The implantation and conception rates of AS rats treated with BMDSCs were equivalent to those of rats with normal uteruses 61 .
CD133 is considered a marker of immature hematopoietic and progenitor cells with high proliferative potential and circulating cells with endothelial regenerative capacity. After being injected into an immunodeficient AS mouse model, human CD133+ BMDSCs were found primarily implanted around stem cell niche, located at the blood vessels of wounded endometrium. Both direct injection into the uterus and tail vein injection are proven to be effective when comparing Ki-67 expression with the non-treated group, suggesting that CD133+ BMDSCs induce the proliferation of peripheral cells via paracrine molecules, such as thrombospondin 1 and insulin-like growth factor 1, which activate mitosis in surrounding cells, thereby initiating regeneration of the endometrium 62 . The VEGF gene transfected BMDSC transplantation is supposed to promote the regeneration of endometrium and improve the endometrial receptivity. The combination revealed a better therapeutic effect than BMDSC treatment alone 63 . BMDSCs show greater ability to migrate to the uterus by systemic injections than EnSCs, resulting in a higher proportion of in situ donor cells 64 .
Clinical studies showed a significant increase in endometrial thickness (ET) and enhanced pregnancy rates after BMDSC treatment in infertile AS patients. Autologous stem cells were selected as an alternative treatment in a woman with severe AS in 2011, after failure of an intrauterine contraceptive device (IUCD) and cyclical hormonal therapy. CD9/90/144+ BMDSCs were isolated and inserted into the endometrial cavity after curettage with ultrasound guidance. When the ET was 8 mm and blood vessels grew well, in vitro fertilization and embryo transplantation were performed. This resulted in a positive biochemical pregnancy, and the subsequent ultrasound examination confirmed the existence of a fetal sac, a yolk sac, and embryonic heart activity on ultrasound examination, indicating that BMDSCs were successfully applied for the first time in an AS patient 65 . In 2014, six cases of refractory AS were involved in mononuclear stem cell (MNC)-based treatment. The MNCs were implanted in the sub-endometrial area and treated with exogenous oral estrogen, resulting in endometrial regeneration reflected by resumption of menstruation in five out of six cases 66 . Later, Santamaria et al. 67 transplanted autologous CD133+ BMDSCs into 11 AS and 5 endometrial atrophies (ET < 5 mm) via spiral arterioles by catheterization, and endometrial function recovered two months after cell therapy. All the patients showed prompted uterine cavities, and three of them became pregnant spontaneously, while another seven became pregnant after using assisted reproductive technology. Another clinical trial showed that five patients with severe AS and secondary infertility received autologous BM MNCs supported by collagen scaffolds transplanted to the endometrium, and satisfactory pregnancy outcomes were achieved in all these patients. In this study, ΔNp63 was also demonstrated as a biomarker of AS, giving rise to endometrial quiescence. At the same time, ΔNp63+ cells showed a distinct AS stemness alteration. It was concluded that BMDSCs reverse quiescence and stemness alterations induced by ΔNp63 in the endometria of AS patients, even though further studies are needed to clarify how different stemness properties change cell fate 13 .
The implantation rate of BMDSCs into the endometrium is 0.2% to 48% for endometrial epithelial cells and 0.3% to 52% for stromal cells, depending on the method of detection and time after BM transplantation 20,60 .
In ischemia-reperfusion injury or inflammation of the uterus, the recruitment of BMDSCs into the endometrium is dramatically promoted, independent of the menstrual cycle, gonadal hormone, or granulocyte-colony stimulating factor (G-CSF) treatment 68 . Moreover, cigarette smoking is detrimental to the recruitment and differentiation of BMDSCs to the endometrium 69 . It has been proven that treatment with CXCL12 leads to increased migration of BMDSCs to the uterus in AS, which in turn leads to increased fertility 70 . In other words, BMDSCs participate in both physiological regeneration and pathological repair as well as endometrium reconstruction after injury.
The mechanism by which these cells are mobilized and directed to the endometrium may involve the CXCL12/CXCR4 signaling pathway. To be more specific, human endometrial stromal cells secrete the chemokine C-X-C motif chemokine 12(CXCL12). Its receptor is C-X-C chemokine receptor type 4 (CXCR4), which is expressed by BM cells, and their combination drives BMDSCs to endometrium. In response to this injury, endogenous CXCL12 expression is increased, thereby enhancing the recruitment of BMDSCs 71 . In the late proliferative phase, elevated E2 causes increased expression of CXCL12/CXCR4 in the endometrium and BM, which subsequently prompts the migration of stem cells to endometrium 72 .
The effect of exogenous CXCL12 has been investigated using a mouse model 70 . Mice were treated with recombinant CXCL12, AMD3100 (CXCR4 antagonist), or both after BM transplantation and endometrial scratch. Administration of CXCL12 increased the degree of BMDSC implantation in the injured uterus, which was significantly blocked by AMD3100. It was speculated that expansion of CXCL12 directed the stem cell migration to uterus, inhibited fibrosis formation, enhanced uterine function, increased litter size, resulted in shorter conception time, and possibly improved fertility in AS mice, even without complementary BM transplantation. This evidence indicates that based on the biological effects of CXCL12 on stem cell mobilization, CXCL12 has therapeutic value in stem cell-related therapies, which suggests that it can be used as a substitute or replacement for stem cell transplantation 70 .
To date, the regenerative characteristics of transplanted BMDSCs may be attributable to the following mechanisms: direct engraftment, transdifferentiation into endometrial stem/progenitor cells, or cell-to-cell contact and secretion of related molecules. Although the transdifferentiation of stem cells into organ-specific cells is believed to be the main principle of stem cell therapy, the number of BMDSCs implanted into the endometrium is low in all injury models, and they do not replace the entire endometrium by clonal expansion 10,64 . In a new 5-fluorouracil (FU)-based non-gonadotoxic sub-myeloablation mouse BMT model, it was demonstrated that BMDSCs could migrate to the endometrium, but only to stromal compartment. It is worth noting that only a portion of stromal cells originate from the BM instead of endothelial or epithelial cells 73 . However, in an irradiated mouse BMT model, no bone marrow-derived stroma, epithelium, or endothelium was observed in the endometrium 74 . Although BMDSCs have been considered to be the origin of endometrial stem cells, current data indicate that the possibility of BMDSCs being ancestors is very low. In addition, only a small number of transdifferentiated cells (<0.1% of all tested cell types) can be detected in the damaged organs after transplantation 75 , suggesting that attractions of secreted molecules are the most likely mechanism of recruitment 70 (Fig. 2).
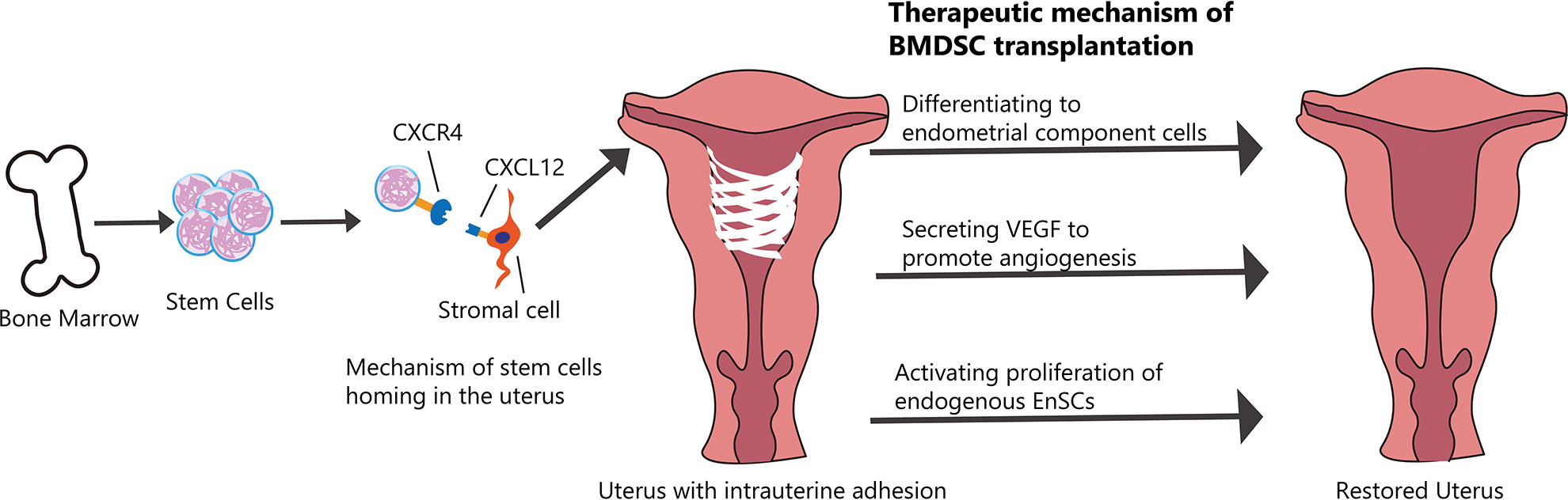
Therapeutic effects of BMDSC transplantation for AS patients. Stem cells are isolated from bone marrow and then injected through vein or into injured uterus directly. CXCL12/CXCR4 signaling pathway is involved in the process of homing of BMDSCs. After transplantation, stem cells can differentiate to stromal cells, epithelial and endothelial cells. Stem cells also stimulate the angiogenesis and proliferation of endogenous cells through the secretion of trophic factors.
Further study of these molecular mechanisms is needed to investigate the therapeutic effects of stem cell therapy 76,77 . In fact, BMDSCs secrete many nutritional factors, including chemokines, cytokines, and growth factors, which stimulate angiogenesis, immune regulation, and stem/progenitor cells, thereby enhancing the endogenous repair system 78 . The therapeutic effects of MSCs may depend on their capacity to secrete soluble factors that promote several key biological activities 76,78 . Extracellular vesicles and membrane nanotubes play an important role in cell-to-cell communication. As with other mechanisms in stem cell research, such research can enrich the further development of new strategies and treatments in regenerative medicine 76 .
Umbilical Cord-Derived Mesenchymal Stem Cells (UC-MSCs)
Human umbilical cord mesenchymal stem cells, also known as WJMSCs, are located in the umbilical cord Wharton’s jelly and areas around the blood vessels and are widely applied in regenerative medicine. Compared with BMSCs, UC-MSCs are highly proliferative with a higher cartilage differentiation rate 79 . Moreover, owing to their advantages, including easy availability, painless collection method, and low immunogenicity, UC-MSCs are more suitable candidates for endometrial damage repair 80 . Finally, quality management during the standardized acquisition of UC-MSCs is easy to control. UC-MSCs are capable of moderately damaging human endometrial stromal cells (ESCs) in vitro or in vivo 81,82 , indicating their potential application value in AS for stem cell-based therapy.
In AS chronic phase, Zhang et al. 83 injected UC-MSCs through the tail vein once, twice, or thrice into AS rats and found that UC-MSC transplantation might repair endometrial damage and restore fertility by inhibiting excessive fibrosis and inflammation, and enhancing endometrial cell proliferation and vascular remodeling. Some circRNAs, such as hsa_circRNA_ 0111659, may be involved in the biological pathways associated with endometrial injury and repair through various regulatory mechanisms. Additionally, the association of circRNA-miRNA-mRNA may also participate in regulating the repair of endometrial injury 84 .
Cao’s group 85 conducted a phase clinical trial by transplanting UC-MSCs and degradable collagen scaffold (CS) as a carrier into the uterine cavity of patients undergoing adhesiolysis. By collecting stem cells, maintaining their viability, and increasing the contact time with the damaged part of the endometrium, the proliferation and differentiation ability of the endometrium was significantly improved. As a result, 10 out of 26 patients successfully became pregnant, and 8 out of 10 patients gave birth without placental complications. This process involves the downregulation of ΔNp63, which may be a contributing element for endometrial regeneration.
Recently, another CS loaded with UC-MSCs was manufactured and applied for endometrial regeneration. The CS/UC-MSC complex accelerated the proliferation of endometrial stromal cells and suppressed cell apoptosis in vitro through paracrine effects. The transplantation of CS/UC-MSCs can maintain normal endometrium structure, facilitate endometrial regeneration and collagen remodeling, and upregulate the expression of estrogen receptor alpha and progesterone receptor. It has been confirmed that the regenerated endometrium has an improved capacity to receive embryos 86 .
Menstrual Blood-Derived Stem Cells (MenSC)
MenSC-based stem cell therapy has received a lot of attention because of the outstanding features of stem cells, such as a non-invasive isolation protocol, abundant source material, their ability to be regularly donated, excellent proliferative capabilities, and potential for use in autologous transplantation 34 .
One group transplanted human endometrial stem cells from menstrual blood organized in spheroids into AS rats. MenSCs retain all the properties of MenSCs in monolayers but with increased synthesis of angiogenic and anti-inflammatory factors. The application of MenSCs significantly improved rat fertility, compared to autologous BM cells 87 . It was shown that MenSCs could differentiate into endometrial cells in vitro and reconstruct endometrial tissue in NODSCID mice in vivo, with impaired cloning efficiency and OCT4 expression of MenSCs from patients with IUA 88 . MenSCs may promote the repair of endometrial lesions in mice by increasing the expression of vimentin, VEGF, and keratin 89 . MenSCs are able to recover the proliferation and migration abilities of impaired endometrial stromal cells and inhibit their apoptosis, which may involve the AKT and p38 signaling pathways 90 . The Hippo signaling pathway is also involved in the induction of endometrial damage repair. When combined with platelet-rich plasma (PRP), this cell therapy was more effective 91 .
In a non-controlled prospective clinical study, autologous MenSC transplantation significantly increased the ET in seven women with AS; in five of them it was increased to 7 mm, which is the optimum thickness for embryo implantation. Four of them underwent frozen embryo transfer (FET). Surprisingly, only one patient became pregnant after the second MenSC transplantation, indicating that autologous MenSC transplantation is an alternative for AS treatment 92 .
Human Amnion-Derived Mesenchymal Stem Cells (HAMSCs)
Human amnion-derived mesenchymal stem cells (HAMSCs) are a novel source of human mesenchymal stem cells isolated from discarded human tissue. They are easy to obtain and are not ethically problematic. In addition, HAMSCs express stem cell markers, including CD90, CD105, CD59, CD49d, and CD44, and possess differentiation potentials 35,93 . Thus, HAMSCs may play an important role in both xenograft and allograft transplantation along with other well-established stem cells, such as BMDSCs. HAMSCs have already been used in many preclinical treatments, and their role in IUA has been clarified.
Yang et al. observed the migration of HAMSCs by labeling them with PKH26 in the endometrium of AS rats 94 . The PKH26 labeling technique proved to be a reliable method for tracing HAMSCs in the treatment of IUA and AS, according to their results. In addition, PKH26 staining had no obvious effect on cell activity.
Gan’s group investigated the therapeutic effect of HAMSCs in AS rats 95 . In the HAMSC transplant group, the endometrium was found to be thicker with alleviated fibrosis. In addition, HAMSC transplantation can promote the regeneration of the endometrium while reducing the expression of pro-inflammatory cytokines (interleukin-1β, tumor necrosis factor-α, and interleukin-8), and upregulating anti-inflammatory cytokines (interleukin 6 and interleukin 10), VEGF, bFGF, and hepatocyte growth factor.
Another type of stem cells that also have multiple differential potential are human amniotic epithelial cells (hAECs). It has been demonstrated that hAECs have the capability to restore fertility in IUA mice by activating the autophagy pathway and accelerating damage repair 96 .
However, current research has focused only on animal models, and further studies are required to explore the feasibility and efficiency of HAMSC engraftment in AS patients.
Challenges and Concerns of Stem Cell Therapy
In spite of the promising prospect of stem cell therapy, we have to face its limitations and difficulties when it comes to clinical application, including the lack of public understanding, the endless ethical arguments and ambiguities in national legislation and regulations. More importantly, there are unsolved barriers in the development of stem cell technology. To be more specific, current obstacles include determining the optimal source of stem cells and the effective mode of cell delivery, establishing perfect criteria for clinical evaluation and assessment of therapeutic effects, and reducing risk of transplantation rejection. The source of another inevitable problem resides in its specialty of high proliferative capacity, which will contribute to endometriosis or the formation of endometrial carcinomas when ectopic endometria grow 32 .
A number of strategies are currently being pursued to overcome these limitations. Selecting the proper mode of delivery is key for efficacy. Various targeted delivery methods including intra-arterial catheterization, trans-myometrial injection and intra-uterine infusion show different positive effects depending on the cell types 65 –67 . Additionally, the timing of transplantation is another controversial factor since the immune cells may act differently during menstrual cycle 97 . Therefore, deep investigation is essential to clearly define the mechanism of each stem cell population in order to carry on individual therapy with satisfied safety and efficacy 98 .
Future Perspectives on Stem Cell-Based Therapy
Cell Therapy Combined with Biomaterials
For stem cell engraftment to the uterine cavity, proper stem cell delivery tools or systems need to be considered. There are several new approaches in the field of cell engineering that fit this criterion, such as cell sheets and membranes 99 . Oral mucosal epithelial cell sheets (OMECS) have been proven to be effective in preventing IUA in rat models, as epithelial cells are observed after transplantation 100 . Recently, Chen et al. investigated the role of decellularized and lyophilized amniotic membrane (DL-AM) seeded with OMECs in inhibiting the progression of IUA in rats 101 . There are many advantages of amniotic membrane (AM), including non-immunogenicity, lower cost, anti-inflammatory, and few ethical issues; thus, AM is thought to be an ideal scaffold for cell delivery 102 . The removal of AM epithelial cells promotes tighter cell adhesion, proliferation, and differentiation, while lyophilization facilitates the recovery of AM. When DL-AM is used alone, there is no regeneration within the epithelium and glands. However, the process can be enhanced with the OMEC combination after 14 and 28 days, for stromal cells may transdifferentiate into epithelial cells in this condition.
Administration of Extracellular Vesicles and Exosomes
The factors released by MSCs, including extracellular vesicles (EVs) and exosomes, make a huge difference in the therapeutic effect of stem cell transplantation 103 . Recently, cell-derived EVs were shown to be a special mechanism for communication between cells 104 . EVs are defined as vesicles originating from the plasma membrane, and are released into the microenvironment by stem/progenitor cells. Additionally, EVs may enhance the angiogenic process in endothelial cells or regulate the phenotypes of cells damaged by a horizontal transfer with miRNA or mRNA 105 . The EVs derived from UC-MSCs also show an impressive ability to decrease inflammation and fibrosis in IUA rats, especially when combined with estrogen. The synergistic effect of UCMSCs-EVs and estrogen results in a highly regenerative agent for AS treatment 106 .
The exosomes derived from MSCs are highly active paracrine components with the potential to repair damaged tissue and promote angiogenesis, and act as mediators of cell-cell communication by transferring mRNAs, miRNAs, and proteins 107,108 . The exosomes have an edge over other cells, as they can be sterilized by filtration with a longer shelf life. They can easily reach the damaged sites through circulation 109 . The exosomes are also less immunogenic, and there is no risk of tumor formation without replication ability 110 . Xiao et al. 111 found that effective antifibrotic function of BMDSCs allows the exosomes to transfer microRNA-340 (miR-340) to ESCs in rats with damaged endometria, which leads to the conclusion that prompting the transfer of BMDSC-derived miR-340 is another way to prevent intrauterine adhesion. Yao et al. 112 concluded that like BMDSCs, the effect of BMDSC-derived exosomes is the reversal of endothelial-mesenchymal transition (EMT) induced by TGF-β1 in rabbits. Saribas et al. 18 transplanted uterine-derived MSCs or exosomes derived from MSCs into AS rats uteri and found increased proliferation and vascularization in uterine tissue and decreased fibrosis in both groups. Furthermore, exosomal administration can improve the damage from AS in a shorter period of time, compared to the MSCs. These findings suggest that MSC-derived exosomes may be a promising alternative therapy for AS.
Conclusion
As a disease that plagues millions of women in childbearing age, AS urgently needs effective treatment methods other than adhesiolysis and assistant management. Stem cell-based therapy has gained considerable attraction in the recent years for treating refractory diseases. It has been shown that there are four types of stem cells in the endometrium that are capable of regenerating endometrial tissue and reconstructing the endometrium in vivo. In the future, researchers should focus on the discovery of specific markers for human EnSC in order to increase the feasibility of using endometrial stem/progenitor cells in AS and IUA.
Other sources of adult stem cells, including the bone marrow, menstrual blood, umbilical cord, and amniotic membrane, that have the potential to differentiate into human endometrial stromal cells, epithelial cells, and endothelial cells, provide alternative approaches for rebuilding the endometrium in cases of AS and IUA. Obstacles that need to be overcome include the risk of tumor formation after transplantation and ethical controversy. Fortunately, the availability of both new technology as well as materials makes the research and application of stem cell base therapy more attractive and practical.
Footnotes
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: Funding for this work was provided by the Provincial Natural Science Foundation of China (20180101140JC and 20190201148JC), the Health Technology Innovation Project of Jilin Province(3D517DS93429), and the Science and Technology Development Project of Jilin Province(20200404169YY).