
Abstract
Pneumonia is a serious respiratory tract infection disease in children, which threatens to the health or life of children patients. Ginsenoside Rb1 (Rb1) is a principle active ingredient extracted from the root of Panax notoginseng (Burk.) F.H. Chen with anti-inflammatory effect. Our study aimed to determine the effects and molecular mechanisms of Rb1 on lipopolysaccharide (LPS)-induced inflammatory injury of lung fibroblasts WI-38 cells. Cell viability and apoptosis were evaluated by CCK-8 and flow cytometry, respectively. The production of inflammatory cytokines were measured by ELISA and RT-qPCR. miR-222 expression was examined by RT-qPCR. The expression levels of the nuclear factor-kappa B (NF-κB) p65 and phosphorylated p65 were detected by western blot. We found that LPS stimulation induced WI-38 cell inflammatory injury by inhibiting cell viability, and inducing apoptosis and inflammatory cytokine production, while treatment with Rb1 significantly attenuated LPS-induced inflammatory injury in WI-38 cells. Additionally, Rb1 decreased LPS-induced upregulation of miR-222 and activation of the NF-κB pathway in WI-38 cells. Overexpression of miR-222 abolished the inhibitory effects of Rb1 on LPS-induced viability reduction, apoptosis, inflammatory cytokine production and activation of the NF-κB pathway. In conclusion, Rb1 alleviated LPS-induced inflammatory injury in WI-38 cells via downregulating miR-222 and inactivation of the NF-kB pathway.
Introduction
Pneumonia is lung inflammation which is caused by infection with viral, bacterial, or fungal pathogens, and remains a common and serious respiratory tract infection disease worldwide occurred in children aged <5 years 1 . The typical clinical characteristics of pneumonia are mainly fever, cough, severe dyspnea, and even breathing disorder, heart failure, and death 2 . Pneumonia remains the frequent cause of global mortality and morbidity in children due to their under-developed immunity and respiratory anatomies 3 , and threatens to the health or life of children patients 4 . During pneumonia, the release of inflammatory cytokines, such as tumor necrosis factor-α (TNF-α), interleukin (IL)-lβ, and IL-6, could cause apoptosis and lung tissue destruction 5 . Despite significant advances in prevention and treatment in recent years, pediatric pneumonia is still confronted with a high risk of recurrence and hospitalization 6 . Therefore, more experimental and clinical studies are indispensable to explore novel effective therapeutic agents for pneumonia.
Panax notoginseng (Burk.) F.H.Chen (P. notoginseng) is a highly valued medicinal herb that is extensively used in Chinese medicine for over 5000 years in China and other Asian countries 7 . Panax notoginseng saponins are thought to be the major pharmacologically active ingredients extracted from the root of P. notoginseng. Ginsenoside Rb1 (Rb1) (its chemical structure was shown in Fig. 1), a principal active component of Panax notoginseng saponins, has been shown to exert multifaceted pharmacological activities, including anti-tumor, anti-aging, anti-apoptotic, anti-angiogenic, anti-oxidative, anti-inflammatory and neuroprotective properties 8 –11 . Pharmacological studies have shown that Rb1 may have beneficial therapeutic effects on multiple human diseases, such as neurodegenerative diseases 12 and cardiovascular disorders 13 . It was reported that Rb1 attenuated pulmonary inflammatory cytokine release and lung histological injury following intestinal ischemia reperfusion (IIR) injury 14 . However, the effects of Rb1 on pneumonia development as well as the underlying mechanism remains to be further explored.
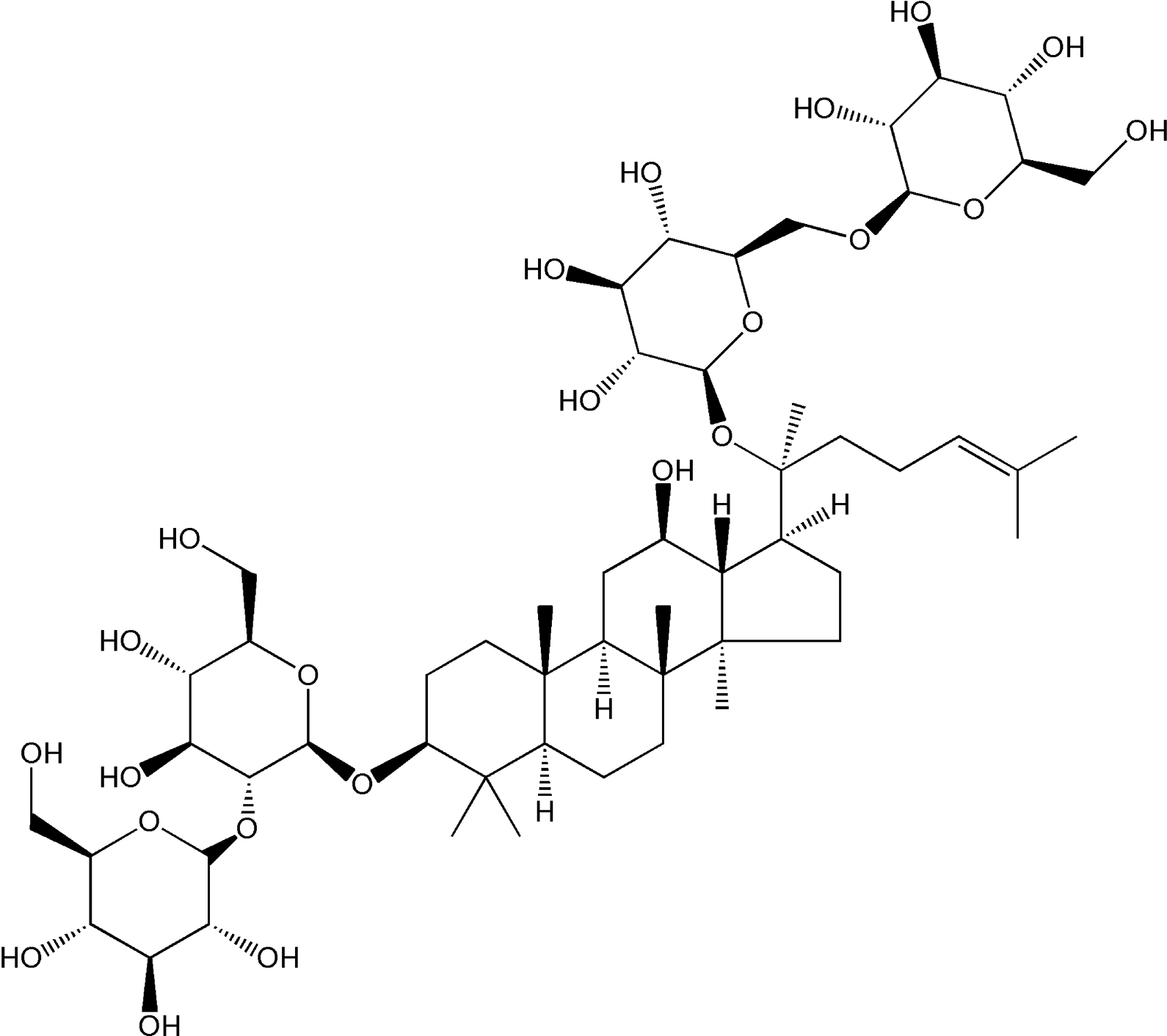
The chemical structure of Rb1.
A previous study reported that Panax notoginseng saponins treatment significantly decreased miR-222 expression in lung cancer 15 . miR-222 has been reported to be highly expressed in children with Mycoplasma pneumoniae pneumonia 16 . Accordingly, we hypothesized that miR-222 was involved in the anti-inflammatory effect of Rb1 on pneumonia. In the present study, human lung fibroblasts WI-38 cells were administrated with lipopolysaccharides (LPS) to establish a cell model of pneumonia. Our study demonstrated that Rb1 inhibited LPS-induced the viability reduction, apoptosis and release of inflammatory cytokines in WI-38 cells by decreasing miR-222 expression and inactivating the nuclear factor-kappa B (NF-κB) signaling pathway.
Materials and Methods
Cell Culture and Treatment
Human fetal lung fibroblasts WI-38 cells were purchased from American Type Culture Collection (Manassas, VA, USA) and cultured in Dulbecco’s modified Eagles medium (Thermo Fisher Scientific, Inc., Waltham, MA, USA) supplemented with 10% fetal bovine serum (Gibco, Grand Island, NY, USA), 100 U/ml of penicillin and 100 μg/ml of streptomycin (Beyotime, Shanghai, China) at 37°C in a humidified atmosphere containing 5% CO2. To induce inflammatory injury in this research, WI-38 cells were treated with appointed doses of LPS (Sigma-Aldrich, St. Louis, MO, USA) for 24 h. Rb1 (purity > 98%) was purchased from Shanghai Tauto Biotech Co (Shanghai, China) and dissolved into DMSO (final DMSO concentration was <0.05%). 0.05% DMSO was used as a blank control, which was nontoxic to WI-38 cells.
Cell Transfection
miR-222 antagomir (miR-222) and antagomir scrambled control (miR-con) were designed and synthesized by GenePharma (Shanghai, China). When growing to 70%–80% confluence, WI-38 cells were transfected with miR-222 or miR-con using lipofectamine 2000 reagent (Invitrogen, Carlsbad, CA, USA).
Cell Viability Assay
Cell counting kit-8 (CCK-8) assay was performed to evaluate the viability of WI-38 cells. Briefly, WI-38 cells were seeded into 96-well plates at 2 × 104 cells/well and incubated overnight. After different treatments, 10 µl of CCK-8 reagent (Dojindo, Tokyo, Japan) was added into the culture medium of each well, followed by incubation for 1 h at 37°C. Subsequently, cell viability was determined by measuring the absorbance at a wavelength of 450 nm using a microplate Reader (Bio-Rad Benchmark, Hercules, CA, USA).
Flow Cytometry Analysis
The apoptosis of WI-38 cells was assessed using an Annexin V-fluorescein isothiocyanate (FITC) Apoptosis Detection Kit (KeyGen Biotech, Nanjing, China). Following different treatments, WI-38 cells were collected and washed with cold PBS. After being resuspended in 100 µL 1 × binding buffer, WI-38 cells were double-stained with 5 µL of Annexin V-FITC and 5 µL propidium iodide (PI) for 15 min in the dark. Finally, the percentage of apoptotic cells was detected by a FACScan Flow Cytometer (BD Biosciences, San Diego, CA, USA).
Caspase-3/7 Activity Assay
The activity of caspase-3/7 in the treated WI-38 cells was measured using a commercially available Caspase-Glo 3/7 assay kit (Promega, Madison, WI, USA) according to the manufacturer’s instructions.
Enzyme-Linked Immunosorbent Assay (ELISA)
Following different treatments, the culture supernatant of WI-38 cells was collected. The levels of inflammatory cytokines including TNF-α, IL-lβ, and IL-6 in the supernatant were measured using corresponding commercially available ELISA kits (R&D Systems, Inc., Minneapolis, MN, USA).
Real Time-Quantitative PCR (RT-qPCR)
Total RNA was extracted from treated WI-38 cells using TRIzol reagent (Invitrogen) and RNA concentration was measured using a Nanodrop 2000 spectrophotometer (NanoDrop Technologies, Wilmington, DE, USA). cDNA was synthesized from 2 µg of total RNA by reverse transcription using NCode mRNA First-Stand cDNA synthesis kit (Invitrogen). To detect miR-222 and the mRNA expression of TNF-α, IL-lβ, and IL-6, qPCR were performed with SYBR® premix Ex Taq™ kit (Takara, Dalian, China) using a Lightcycler™ 480 Real-Time PCR system (Roche Applied Science, Rotkreuz, Switzerland). The primer sequences were as follows: miR-222 forward, 5′-CGCA GCTA CATC TGGC TACT G-3′, and reverse, 5′-GTGC AGGG TCCG AGGT-3′; U6 forward, 5′-GCGC GTCG TGAA GCGT TC-3′ and reverse, 5′-GTGC AGGG TCCG AGGT-3′; TNF-α forward, 5′-CCGA GTGA CAAG CCTG TAGC-3′ and reverse, 5′-AGGA GGTT GACC TTGG TCTG-3′; IL-lβ forward, 5′-ACAG ATGA AGTG CTCC TTCC A-3′ and reverse, 5′-GTCG GAGA TTCG TAGC TGGA T-3’; IL-6 forward, 5′-TACA TCCT CGAC GGCA TCTC-3′ and reverse, 5′-AGCT CTGG CTTG TTCC TCAC-3′; GAPDH forward, 5′-GAGT CAAC GGAT TTGG TCGT-3′ and reverse, 5′-TTGA TTTT GGAG GGAT CTCG-3′. The relative mRNA expression levels were normalized to U6 or GAPDH, and data analysis was performed using the 2-ΔΔCt method.
Western Blot Analysis
Whole cellular protein and nucleoprotein from treated WI-38 cells were extracted using RIPA lysis buffer (Invitrogen) with protease inhibitors (Sigma) and NE-PER Nuclear Extraction Reagents (Thermo Fisher Scientific Inc.) according to the instructions, respectively. Equivalent amounts of protein samples (20 μg/lane) were subjected to 10% sodium dodecyl sulphate-polyacrylamide gel electrophoresis and then electrotransferred to a polyvinylidene difluoride membrane (Millipore, Billerica, MA, USA). The membrane was then blocked with 5% skimmed milk in Tris Buffered Saline with Tween (TBST) at room temperature for 2 h and incubated overnight with the following antibodies pohsphrylated p65 (p-p65) (Abcam, Cambridge, MA, USA), p65 (Abcam) and β-actin (Abcam) at 4°C, followed by incubation with a corresponding horseradish peroxidase-conjugated secondary antibody (Abcam) at room temperature for 1 h. The protein signals were visualized using an enhanced chemiluminescence kit (Pierce, Rockford, IL, USA). The intensity of bands was quantified using ImageJ software (NIH, Bethesda, MD, USA).
Statistical Analysis
All experiments were repeated three times and data are shown as the means ± standard deviation (SD). All statistical analyses were carried out using Graphpad 6.0 statistical software (Graphpad, San Diego, CA, USA) with one-way analysis of variance analysis (ANOVA) or Student’s t test. P < 0.05 was considered to indicate a statistically significance.
Results
Rb1 Suppressed LPS-Induced Viability Reduction of WI-38 Cells
Firstly, we evaluated the viability of WI-38 cells after stimulation with 0, 1.25, 2.5, 5, and 10 mg/ml LPS for 24 h. As demonstrated by CCK-8 assay, cell viability was dose-dependently inhibited by LPS stimulation at 2.5, 5, and 10 mg/ml (Fig. 2A). Next, the effect of Rb1 on the viability of WI-38 cells was assessed by CCK-8 assay and the result manifested that Rb1 showed no significant effect on cell viability in WI-38 cells (Fig. 2B). Furthermore, the effects of Rb1 on LPS-induced viability reduction of WI-38 cells were explored. CCK-8 assay demonstrated that Rb1 treatment alleviated LPS-induced reduction of the viability of WI-38 cells in a concentration-dependent manner (Fig. 2C). These results demonstrated that Rb1 suppressed LPS-induced viability reduction of WI-38 cells.
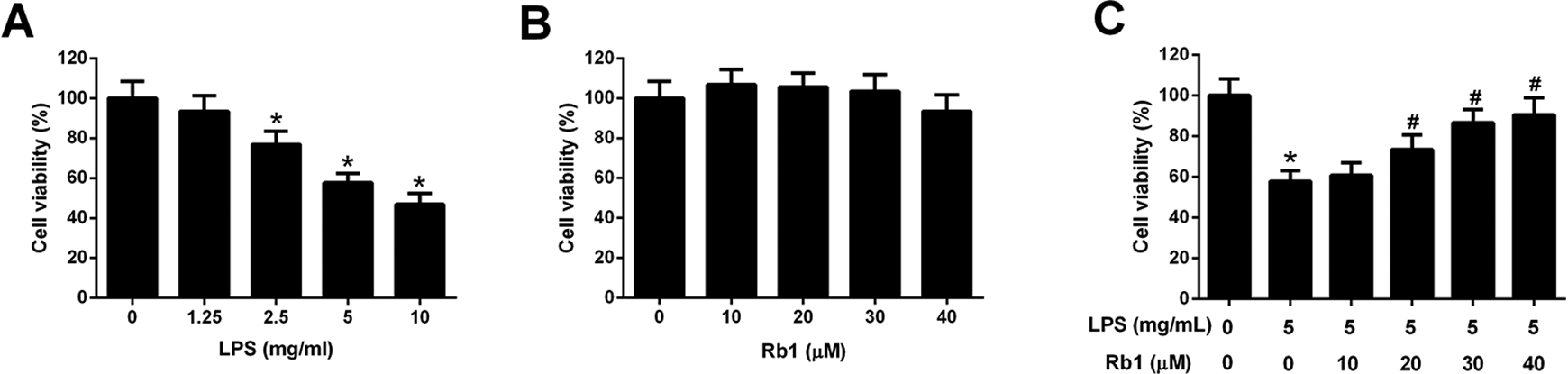
Effect of Rb1 on LPS-induced viability reduction of WI-38 cells. (A) CCK-8 assay was performed to detect cell viability in WI-38 cells after stimulation with 0, 1.25, 2.5, 5, and 10 mg/ml LPS for 24 h. (B) CCK-8 assay was conducted to assess the viability of WI-38 cells following by treatment with 0, 10, 20, 30, and 40 μM Rb1 for 24 h. (C) CCK-8 assay was applied to evaluate cell viability after WI-38 cells were treated with Rb1 (0, 10, 20, 30, and 40 μM) in the presence or absence of 5 mg/ml LPS for 24 h. *P < 0.05.
Rb1 Inhibited LPS-Induced Apoptosis of WI-38 Cells
Flow cytometry analysis and caspase-3/7 activity assay showed that 30 μM Rb1 alone caused no noticeable change of the apoptotic rate and caspase-3/7 activity in WI-38 cells, and LPS exposure alone distinctly increased the percentage of apoptotic cells and caspase-3/7 activity in WI-38 cells (Fig. 3A, B). However, LPS-induced elevation of apoptotic rate and caspase-3/7 activity in WI-38 cells were antagonized following treatment with 20 and 30 μM Rb1 (Fig. 3A, B). Collectively, these data suggested that Rb1 inhibited LPS-induced apoptosis of WI-38 cells.
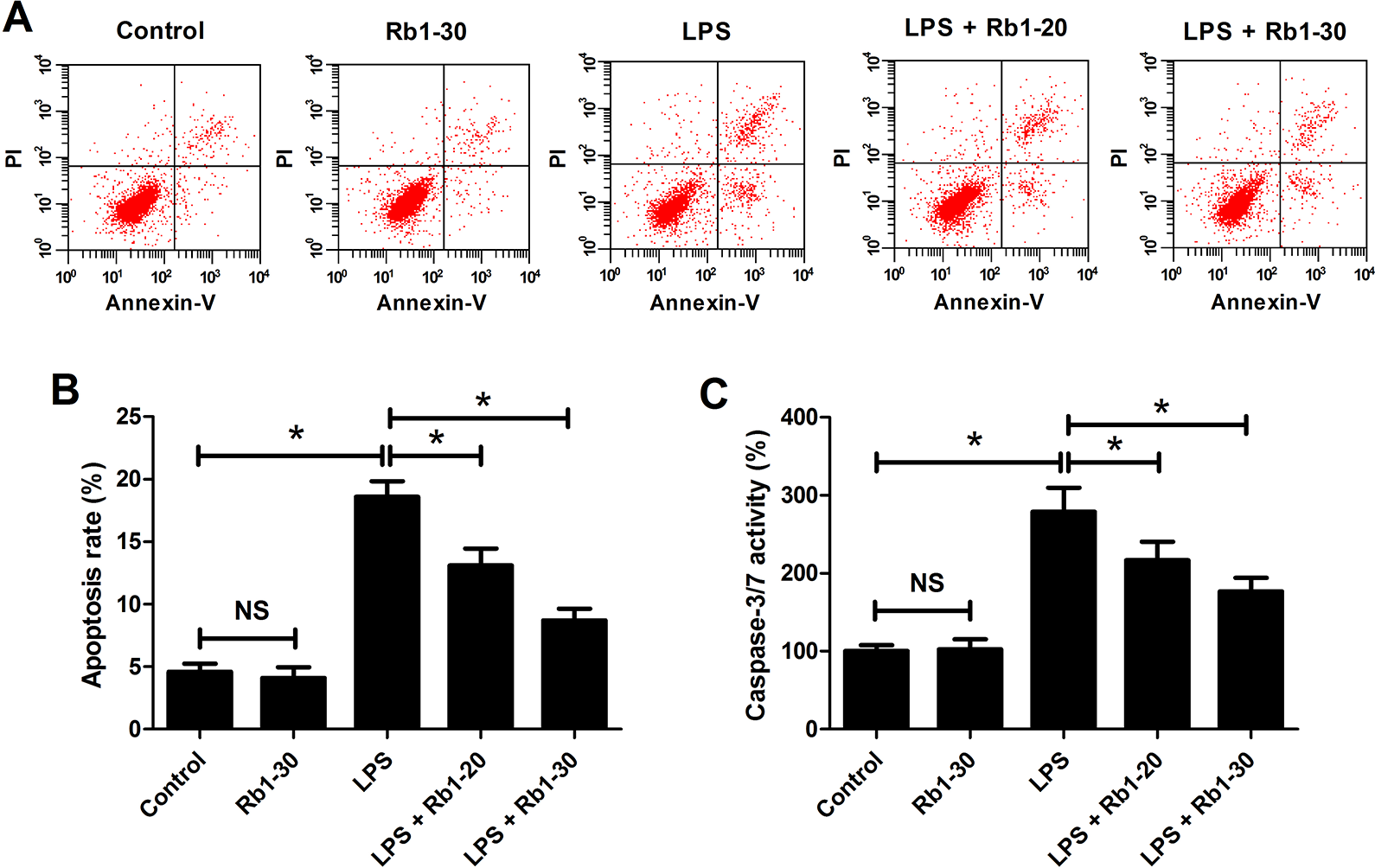
Effect of Rb1 on LPS-induced apoptosis of WI-38 cells. WI-38 cells were treated with 5 mg/ml LPS or 30 μM Rb1 for 24 h, or administrated with 20 or 30 μM Rb1 in the presence of 5 mg/ml LPS for 24 h. (A) The percentage of apoptotic WI-38 cells was measured by flow cytometry analysis. (B) Caspase-3/7 activity in the treated WI-38 cells was measured using caspase-3/7 activity assay. *P < 0.05. NS, not significant.
Rb1 Attenuated LPS-Induced Inflammatory Cytokines Production in WI-38 Cells
As shown in Fig. 4A–F, the mRNA expression and concentrations of IL-1β, IL-6, and TNF-α in WI-38 cells were not changed in response to 30 μM Rb1 compared to control group but were prominently enhanced following LPS stimulation. Relative to LPS group, the mRNA expression and concentrations of IL-1β, IL-6, and TNF-α in WI-38 cells were declined in LPS + 20 or 30 μM Rb1 co-treatment group, suggesting that Rb1 mitigated LPS-induced inflammatory cytokines production in WI-38 cells.
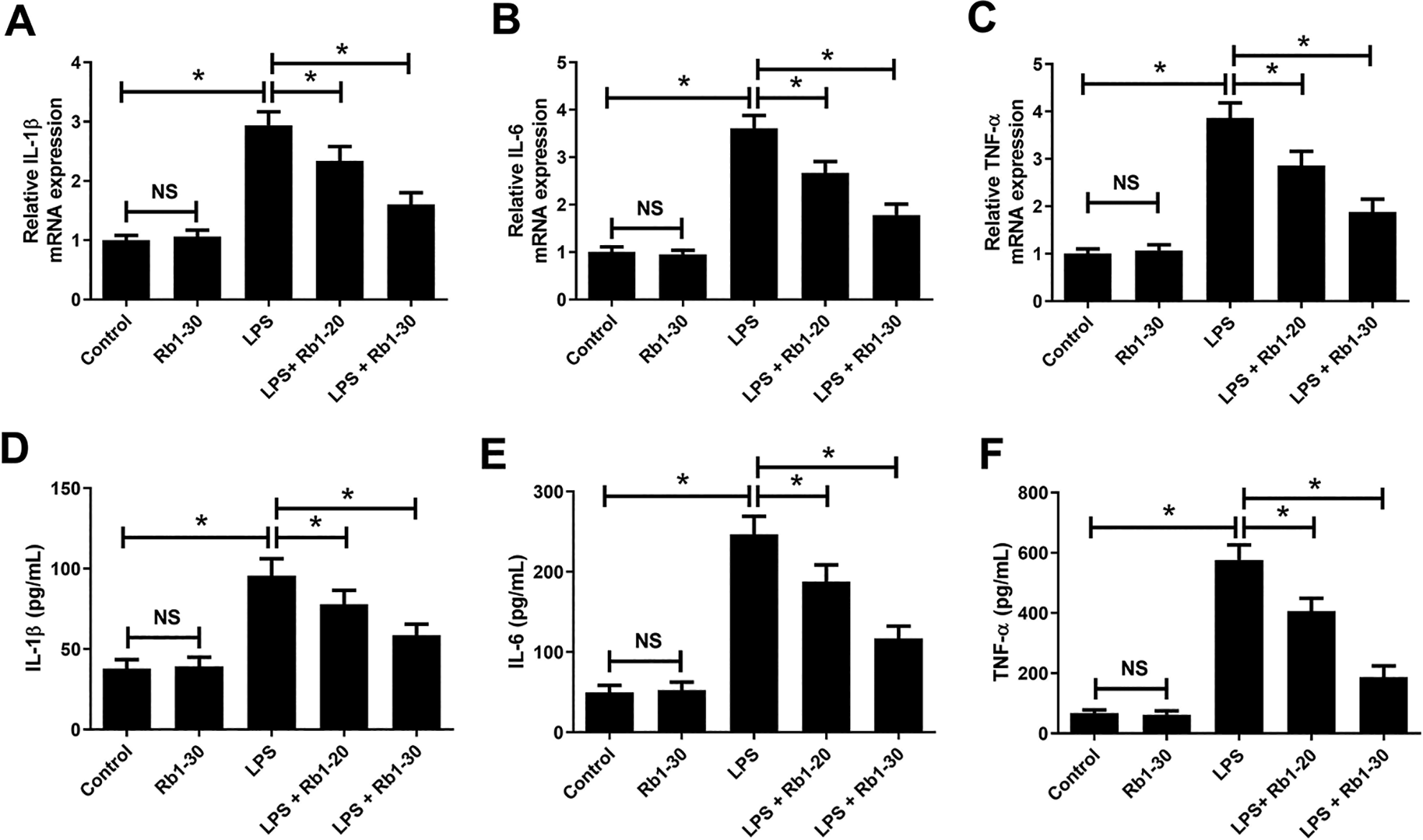
Effect of Rb1 on LPS-induced inflammatory cytokines production in WI-38 cells. WI-38 cells were treated with 5 mg/ml LPS or 30 μM Rb1 for 24 h, or administrated with 20 or 30 μM Rb1 in the presence of 5 mg/ml LPS for 24 h. Then, the mRNA expression (A–C) and concentrations (D–F) of IL-1β, IL-6 and TNF-α in WI-38 cells were detected by RT-qPCR and ELISA, respectively. *P < 0.05. NS, not significant.
Rb1 Decreased LPS-Induced Upregulation of miR-222 in WI-38 Cells
The expression of miR-222 in WI-38 cells after LPS and/or Rb1 treatment was examined by RT-qPCR. As displayed in Fig. 5, 30 μM Rb1 treatment slightly suppressed miR-222 expression in WI-38 cells. miR-222 expression was considerably augmented in response to LPS stimulation, while treatment with 20 or 30 μM Rb1 weakened LPS-induced increase of miR-222 expression in WI-38 cells (Fig. 5).
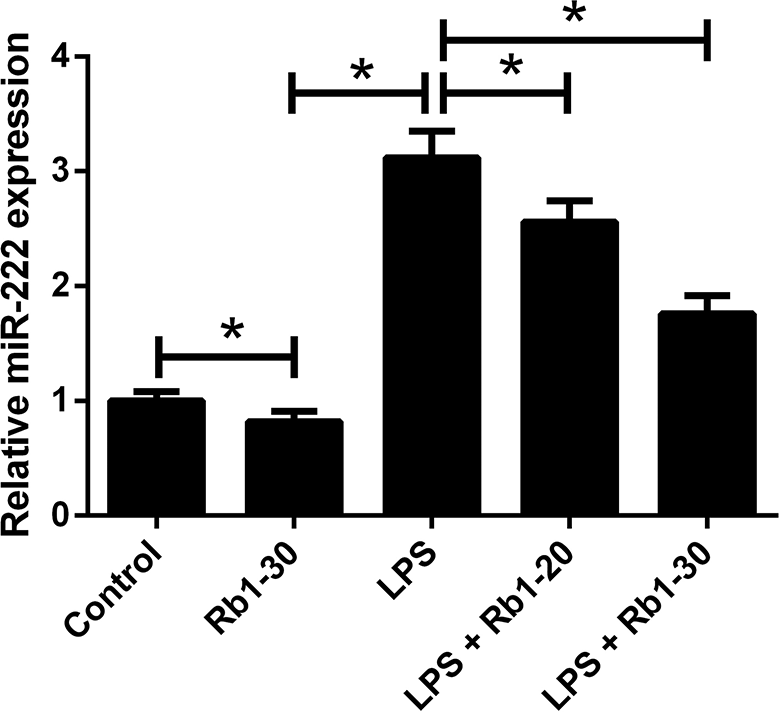
Effect of Rb1 on LPS-induced upregulation of miR-222 in WI-38 cells. WI-38 cells were treated with 5 mg/ml LPS or 30 μM Rb1 for 24 h, or administrated with 20 or 30 μM Rb1 in the presence of 5 mg/ml LPS for 24 h, followed by RT-qPCR analysis of miR-222 expression.
Rb1 Inhibited LPS-Induced Activation of NF-κB Pathway in WI-38 Cells
To investigate the molecular mechanism underlying the anti-inflammatory effect of Rb1 on LPS-stimulated WI-38 cells, we focused on the NF-κB pathway, which is well-known to be closely associated with inflammation 17 . As shown in Fig. 6A, B, no significant changes of p-p65, p65, and nuclear p65 expression were observed in Rb1-treated WI-38 cells. Moreover, LPS exposure led to a significant increase of p-p65/p65 and nuclear p65 expression in WI-38 cells, suggesting that LPS induced activation of the NF-κB pathway in WI-38 cells. However, this effect was inhibited by treatment with 20 or 30 μM Rb1. These results demonstrated that Rb1 inhibited LPS-induced activation of NF-κB pathway in WI-38 cells.
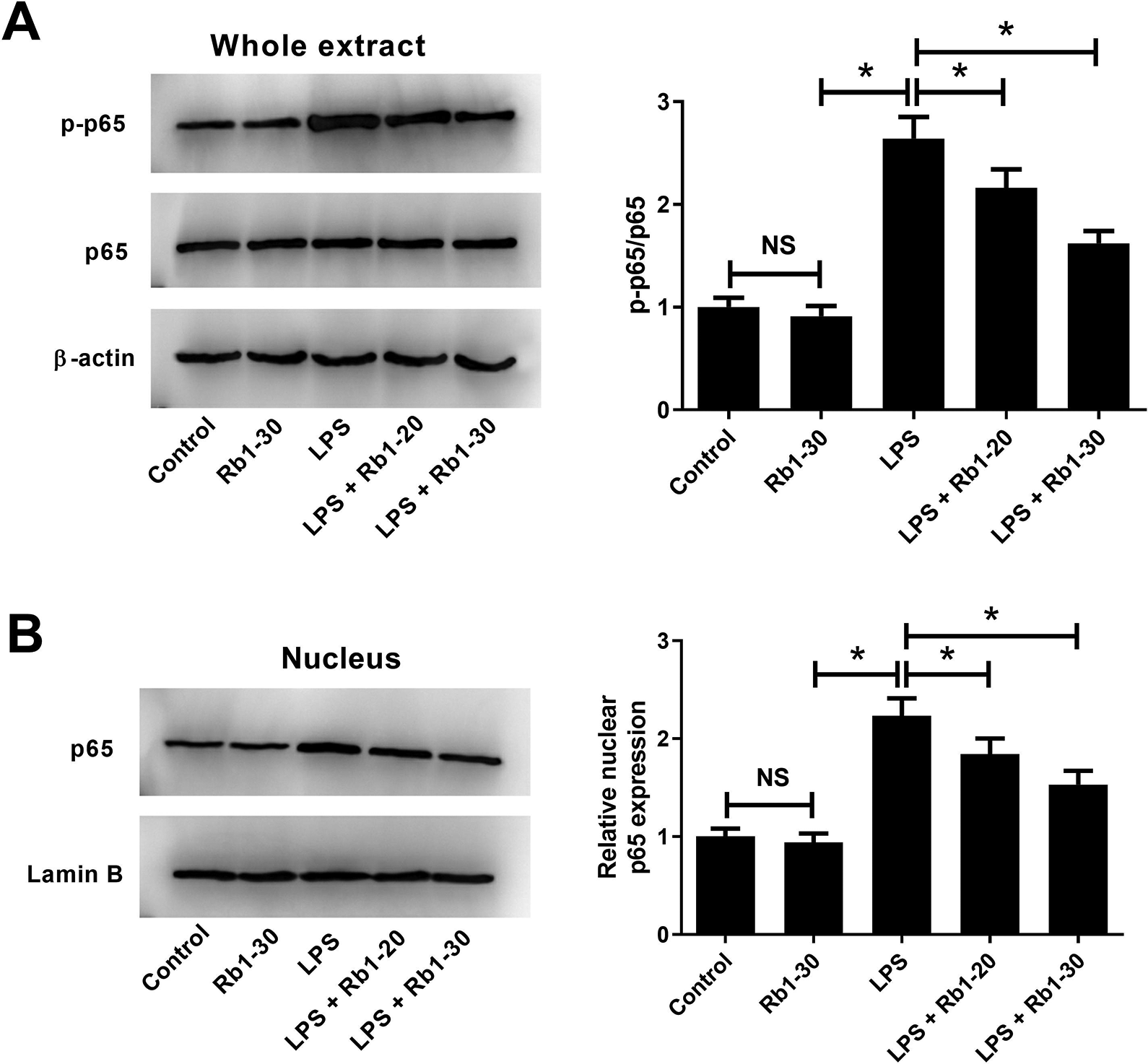
Effect of Rb1 on LPS-induced activation of NF-κB pathway in WI-38 cells. (A, B) WI-38 cells were treated with 5 mg/ml LPS or 30 μM Rb1 for 24 h, or administrated with 20 or 30 μM Rb1 in the presence of 5 mg/ml LPS for 24 h. Western blot analysis was conducted to detect the expression of p-p65, p65, and nuclear p65 in WI-38 cells. *P < 0.05. NS, not significant.
Overexpression of miR-222 Restrained the Effects of Rb1 on LPS-Induced Viability Reduction and Apoptosis in WI-38 Cells
To further clarify the roles of miR-222 on the anti-inflammatory effect of Rb1, WI-38 cells were transfected with miR-222 or miR-con and RT-qPCR analysis revealed that miR-222 expression was augmented by delivery of miR-222 compared with miR-con group (Fig. 7A). CCK-8 assay disclosed that Rb1 strikingly alleviated LPS-induced viability reduction of WI-38 cells, which was abolished by promotion of miR-222 (Fig. 7B). Flow cytometry analysis and caspase-3/7 activity assay manifested that the inhibitory effects of Rb1 on LPS-induced enhancement of apoptotic rate and caspase-3/7 activity in WI-38 cells were prominently ameliorated by miR-222 overexpression (Fig. 7C, D). Taken together, we concluded that Rb1 suppressed LPS-induced viability reduction and apoptosis in WI-38 cells by downregulating miR-222.
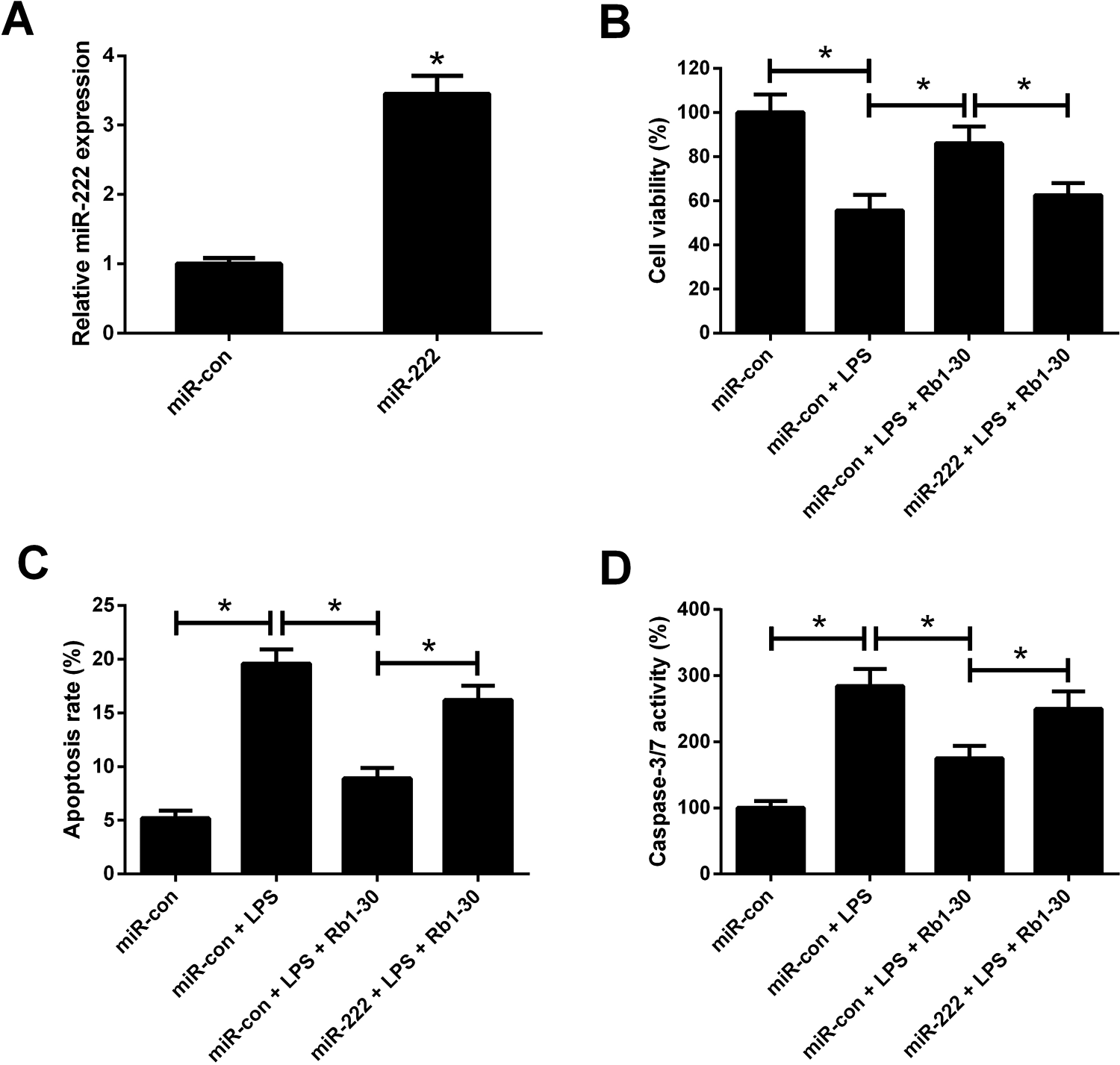
Effects of Rb1 or combined with miR-222 on LPS-induced viability reduction and apoptosis in WI-38 cells. (A) RT-qPCR analysis of miR-222 expression in WI-38 cells after transfection with miR-222 or miR-con for 48 h. WI-38 cells were transfected with miR-222 or miR-con for 24 h, and then treated with 5 mg/ml LPS, or treated with 5 mg/ml LPS plus 30 μM Rb1 for another 24 h, followed by assessment of cell viability (B), apoptosis (C), and caspase-3/7 activity (D) by CCK-8, flow cytometry analysis and caspase-3/7 activity assay, respectively. *P < 0.05.
Overexpression of miR-222 Resisted the Effect of Rb1 on LPS-Induced Inflammatory Cytokines Production and Activation of NF-κB Pathway in WI-38 Cells
ELISA showed that ectopic expression of miR-222 reversed the suppressive effect on LPS-induced increase of concentrations of IL-1β (Fig. 8A), IL-6 (Fig. 8B), and TNF-α (Fig. 8C) in WI-38 cells mediated by Rb1. In addition, western blot analysis implied that the inhibitory effect of Rb1 on LPS-induced elevation of p-p65/p65 (Fig. 8D) and nuclear p65 (Fig. 8E) in WI-38 cells was effectively relieved following by enforced expression of miR-222. Collectively, these results uncovered that Rb1 impeded LPS-induced inflammatory cytokines production and activation of NF-κB pathway in WI-38 cells by downregulating miR-222.
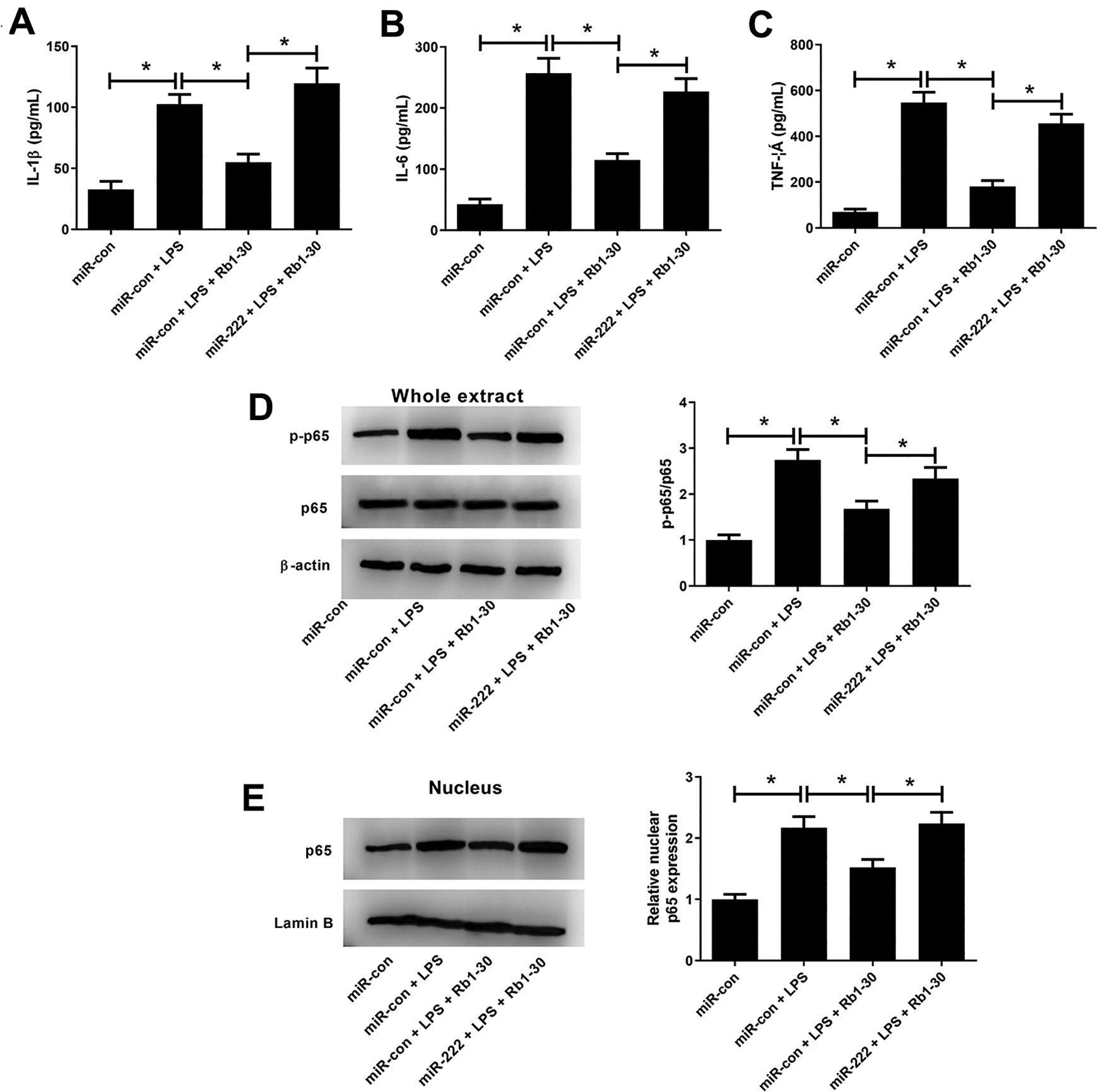
Effects of Rb1 or along with miR-222 on LPS-induced inflammatory cytokines production and activation of NF-κB pathway in WI-38 cells. WI-38 cells were transfected with miR-222 or miR-con for 24 h, and then treated with 5 mg/ml LPS, or treated with 5 mg/ml LPS plus 30 μM Rb1 for another 24 h. The levels of inflammatory cytokines IL-1β (A), IL-6 (B), and TNF-α (C) in treated WI-38 cells were measured by ELISA. Western blot analysis was conducted to detect the expression of p-p65, p65 (D), and nuclear p65 (E) in treated WI-38 cells. *P < 0.05.
Discussion
Pneumonia is a major contributor to childhood morbidity, leading to about one million mortalities annually 18 . Obviously, identifying safe and effective medicines to treat pneumonia has become a research focus worldwide. In the present study, we investigated the effects of Rb1 on LPS-induced inflammatory injury in WI-38 cells. We demonstrated that Rb1 remarkably attenuated LPS-induced inflammatory injury by suppressing LPS-induced viability reduction, apoptosis and the release of inflammatory cytokines in WI-38 cells. More notably, miR-222 overexpression reversed the anti-inflammatory effect of Rb1 on WI-38 cells. Furthermore, we discovered that Rb1 inhibited LPS-induced activation of NF-κB pathway in WI-38 cells via downregulating miR-222.
In recent years, plant-derived natural drugs in the treatment of human diseases have received increasing attention worldwide, owing to their safety, high efficiency and low side effects 19 . Rb1, a principle active saponin extracted from P. notoginseng, has been extensively studied due to its anti-inflammatory activity. For instance, Rb1 was documented to possess potential anti-inflammatory and anti-apoptotic properties in human articular chondrocytes 20 . A previous study demonstrated that Rb1 attenuated IIR-induced inflammation and oxidative stress by activating the phosphoinositide 3-kinase (PI3 K)/protein kinase B/nuclear factor erythroid 2-related factor 2 (Nrf2) pathway 21 . More importantly, it has been proved that Rb1 ameliorated IIR- or LPS-induced lung injury via inhibiting NF-kB-dependent inflammatory response 22,23 . However, whether Rb1 could exert beneficial effects on pneumonia remains unclear. LPS, a key cellular wall component of Gram-negative bacteria, could repress cell proliferation and induce inflammatory response in lung fibroblasts 24 . It is well-established that LPS-induced cell model of pneumonia is extensively applied to investigate the progression of pneumonia and identify effective therapeutic agents for pneumonia 25 . In our study, we demonstrated that LPS stimulation induced WI-38 cell inflammatory injury by inhibiting cell viability, and inducing apoptosis and inflammatory cytokine production. In addition, we found that Rb1 alone showed no significant effect on the viability, apoptosis, and inflammatory cytokine release in WI-38 cells. However, to the best of our knowledge, it is the first time to demonstrate that Rb1 attenuated LPS-induced inflammatory injury in WI-38 cells by suppressing LPS-induced viability reduction, apoptosis and inflammatory cytokine production, suggesting that Rb1 may be a potential anti-inflammatory drug for the treatment of pneumonia.
miRNAs are a group of small endogenous non-coding RNAs of approximately 19–21 nucleotides in length, which exert important functions on the posttranscriptional regulation in the expression of target mRNAs 26 . Compelling evidence has shown that miRNAs participate in the modulation of cell proliferation, apoptosis, immune system development, and release of inflammatory cytokines 27 . It has been proposed that some miRNAs are differentially expressed in pneumonia and involved in the pathogenesis and development of inflammatory lung diseases including pneumonia 28 . miR-222, which is clustered in tandem with miR-221 on human chromosome Xp11.3, is highly conserved in vertebrates and identified as an inflammation-related miRNA 29 . It was documented that miR-221/miR-222 knockdown in mice abolished the inhibitory effects of resveratrol on TNF-α-induced inflammation and intercellular adhesion molecule (ICAM)-1 expression in endothelial cells 30 . In our study, we found that miR-222 expression was dramatically increased in response to LPS stimulation in WI-38 cells. Similarly, it was previously demonstrated that LPS also caused upregulation of miR-221 in WI-38 cells 31 . We further manifested that treatment with Rb1 decreased LPS-induced upregulation of miR-222 in WI-38 cells. Moreover, we found that miR-222 overexpression attenuated the inhibitory effects of Rb1 on LPS-induced viability reduction, apoptosis, and inflammatory cytokine production in WI-38 cells. These results suggested that Rb1 exerted protective effects on LPS-induced inflammatory injury in WI-38 cells via downregulating miR-222.
The NF-κB signaling pathway is a crucial signaling pathway that plays a vital role in regulating the expression of numerous genes that are important for apoptosis, cell survival, release of inflammatory cytokines, and autoimmune diseases 32 . Improper activation of NF-κB pathway can promote cellular inflammatory response via enhancing the expression of various inflammatory cytokines 33 . It has been demonstrated that LPS could induce activation of NF-κB pathway in pneumonia, thus inducing inflammatory response 34,35 . Previously, it was reported that inhibition of the NF-κB pathway improved median survival times and recipient oxygenation in a mouse model of idiopathic pneumonia syndrome 36 . In the present study, we demonstrated that LPS induced the activation of NF-kB pathway in WI-38 cells, while incubation with Rb1 suppressed LPS-induced activation of NF-kB pathway in WI-38 cells, consistently with the previous studies 22,23 . Furthermore, promotion of miR-222 abolished the inhibitory effect of Rb1 on LPS-induced activation of NF-kB pathway in WI-38 cells. Therefore, we concluded that Rb1 alleviated LPS-induced inflammatory injury in WI-38 cells by downregulating miR-222 and inactivation of the NF-kB pathway. It has been reported that miR-222 directly targeted PDZ and LIM domain protein 2 (PDLIM2), leading to activation of NF-κB pathway 37 . miR-222 was also reported to target adiponectin receptor 1 (ADIPOR1), a negative regulator of NF-κB pathway 38 . miR-222 posttranscriptionally regulated peroxisome proliferator-activated receptor γ coactivator 1α (PGC-1α) 39 , and PGC-1α repressed the NF-κB pathway, affecting inflammatory processes 40 . Thus, we deduced that Rb1 inhibited the NF-κB pathway by regulating multiple target genes of miR-222, thus exerting its effects in LPS-induced inflammatory injury in WI-38 cells.
In summary, our study clarified the protective role of Rb1 in LPS-induced inflammatory injury in WI-38 cells and provided the first evidence that Rb1 alleviated LPS-induced inflammatory injury in WI-38 cells via downregulating miR-222 and inactivation of the NF-kB pathway, contributing to our understanding of the effects and molecular mechanism of Rb1 in pneumonia. Our findings suggested that Rb1 might be a promising therapeutic agent for pneumonia. A major limitation of this study is the lack of evidence showing the protective effects of Rab1 against pneumonia in vivo. Further studies are required to explore whether Rab1 exerts anti-pneumonia activity, regulates miR-222, and inactivates the NF-kB pathway in an animal model of pneumonia.
Supplemental Material
Supplemental Material, sj-docx-1-cll-10.1177_09636897211002787 - Ginsenoside Rb1 Alleviates Lipopolysaccharide-Induced Inflammatory Injury by Downregulating miR-222 in WI-38 Cells
Supplemental Material, sj-docx-1-cll-10.1177_09636897211002787 for Ginsenoside Rb1 Alleviates Lipopolysaccharide-Induced Inflammatory Injury by Downregulating miR-222 in WI-38 Cells by Erhu Wei, Xiao Fang, Peisheng Jia, Mingxia Li, Peina Jin, Fengyan Li, Huaili Wang and Dan Gao in Cell Transplantation
Footnotes
Authors’ Contribution
Erhu Wei, Xiao Fang and Peisheng Jia performed experiments, analyzed data, and wrote the manuscript. Mingxia Li, Peina Jin and Fengyan Li designed research, performed experiments, and analyzed data. Huaili Wang and Dan Gao conceived and designed research.
Ethical Approval
This study was approved by our institutional review board.
Statement of Human and Animal Rights
This article does not contain any studies with human or animal subjects.
Statement of Informed Consent
There are no human subjects in this article and informed consent is not applicable.
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
This work was supported by the Medical Education Research Project of Henan Province (No. Wjlx2019021), the Bethune Foundation for Medical Science Research (No. SCE183AN), and the Joint Construction Project of Henan Medical Science and Technology Research Plan (No. LHGJ20200338).
Supplemental Material
Supplemental material for this article is available online.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.