
Abstract
Mammalian fat comprises white and brown adipose tissue (WAT and BAT, respectively). WAT stores energy, whereas BAT is used for thermogenesis. In recent years, the incidence of obesity and its associated disorders have increased tremendously. Considering the thermogenic capacity and decreased levels of BAT with increasing age, BAT can be used as a suitable therapeutic target for the treatment of obesity and diabetes. In several studies, using positron emission tomography and computed tomography images, adult humans have been shown to have functional BAT in interscapular fat. Results of these basic research studies on BAT have shed light on the new components of transcriptional regulation and the role of hormones in stimulating BAT growth and differentiation. In this review article, we have summarized the thermogenic regulators identified in the past decades by focusing on peroxisome proliferator-activated receptor gamma/uncoupling protein 1 activators, branched-chain amino acids, fatty acids (lipokine), and adenosine monophosphate-activated protein kinase mediators. We have also presented the progress of a few ongoing clinical trials aimed at the treatment of obesity and its associated metabolic disorders. The main purpose of this review was to provide a comprehensive introduction to the latest knowledge of the representative thermogenic regulators for the treatment of obesity. The fat combustion capacity of BAT may have great potential and can be considered as a suitable target for the therapeutic application of drugs from bench-to-bed treatment of obesity and the associated diseases.
Introduction
According to the World Health Organization, the global obese population based on body mass index has increased from 105 million in 1975 to 650 million in 2015, and >1.9 billion adults (>18 years of age) in the world are overweight. In the past 40 years, the incidence of obesity has increased by approximately five times. Approximately 39% of the adults aged ≥18 years were reported to be overweight in 2016, whereas 13% of the adults aged >18 years were reported to be obese. More than 340 million 5–18-year-old children and adolescents have been reported to be overweight or obese, and approximately 41 million young children aged <5 years have been reported to be overweight or obese, indicating that obesity has developed into a modern epidemic disease.
Because of westernization, factors such as sedentary life, aging, drug abuse, smoking, staying-up late, and pregnancy may be the risk for obesity 1 ; the global prevalence of obesity continues to increase greatly. These factors cause excessive adipose tissue to accumulate in the body, thereby causing harm to the health of the body. Excessive adipose tissue accumulation affects energy expenditure and homeostasis. Except in cases of children with congenital heart diseases, obesity is closely related to the daily lifestyle and work schedules. Almost one billion people have been reported to have chronic metabolic diseases that are affected by obesity, including diabetes, hyperinsulinemia, cardiovascular disease, osteoarthritis, cancer, stroke, nonalcoholic fatty liver disease, lung disease, and sleep-disordered breathing 2 . Because the high incidence of obesity and metabolic syndrome continues to increase the systemic burden and risk of chronic diseases, finding potential and better treatments that are safe and effective for obesity and the related metabolic diseases is as important as other researches and developments.
Therapeutic Strategy Based on Fat Combustion and Clinical Development
The annual estimates of obesity in the past several years show that the prevalence of obesity and the associated diseases is rising. Since the brown adipose tissue (BAT) shows substantial capacity to expedite energy, targeting brown adipogenic thermogenesis may be considered as a suitable therapeutic approach for the treatment or prevention of obesity and its associated disorders. Three review articles have discussed the potential and the application of thermogenesis in obesity and diabetes 3 –5 .
The therapeutic strategy for obesity can be classified into two types: first, the pharmacological in vivo approach, represented by sympathetic activation. Although the human BAT expresses the β-adrenergic receptor, the early human trials for β-adrenergic receptor agonists were not successful 6 . The reasons for the failed trails were speculated to be that human BAT has several binding characteristics. Because BAT therapies based on β-adrenergic agonists are clinically not feasible, alternative strategies were investigated. The second approach is the ex vivo cell-based therapy, which involves cell regeneration and induced differentiation. Research studies on adipogenesis have used precursor cells for regeneration, which are then implanted in patients with metabolic disorders. Embryonic stem cells undergo unlimited multiplication, but they are not generally used for cell therapy owing to the limited access to the cell source, ethical controversies, higher risk for teratoma formation, and autoimmune response. Mesenchymal stem cells (MSCs) can be used as precursor cells because their sources are accessible, they are available in unlimited supply, and they cause lower risk of autoimmune responses and oncogenic development. However, the effects of MSC transplantation are incomplete and temporary; long-term administration and adjunct treatments are required. Recent studies have examined the effects of induced pluripotent stem cells (iPSCs) in several diseases. IPSCs are differentiated from somatic cells; therefore, these cells have no controversial issues and they are easily obtained from the patients. However, IPSCs have the potential to undergo mutations through reprogramming methods and there are barriers to long-term transplant viability and functionality 7 . The findings of all these studies show that pharmacological approaches have progressed considerably; therefore, the main focus of interest in this review is targeting obesity using small molecules.
To date, several persuasive evidences have shown that thermogenic regulation has a therapeutic effect on obesity. With growing demands for treatments for obesity and the associated disorders, several clinical studies have been conducted safely and effectively in recent years. The clinical trials registration site of United States shows that 16 clinical trials involving small molecules targeting obesity are underway (Table 1). Two trials are active but not recruiting (NCT02964442 and NCT02133040), two trials are recruiting now (NCT03379181 and NCT04086381), and the other 12 projects have been completed. These drugs have been tested for the treatment of obesity, insulin resistance, and hyperlipidemia. To date, there are no effective and proved drugs that can increase the activation and the amounts of BAT in the clinical settings for the treatment of metabolic diseases. Therefore, reviewing recent research advances in thermogenic regulators is significant for determining their potential as antiobesity drugs.
Thermogenic Regulators in Clinical Trials.

NCT: national clinical trials; MRI: magnetic resonance imaging.
Small-Molecule-Mediated Peroxisome Proliferator-Activated Receptor Gamma/Uncoupling Protein 1 Gene Induction in Beige Cells
Earlier, many studies focused on the regulatory molecules or environmental factors related to fat browning. In fat browning, white adipose tissue (WAT) can change its phenotype to that of a brown-like adipose tissue, which is known as beige adipose tissue. This conversion is characterized by an increase in thermogenic capacity mediated by uncoupling protein 1 (UCP1). This browning process is a potential new target for treating obesity 8 . For example, intermittent exercise or low-temperature environmental (cold acclimatization) stimulation in animal experiments promotes BAT activation from WAT, and sympathetic activation plays a key role 9 . The activation of the β-adrenergic receptor initiates a cascade of signals, leading to the hydrolysis of triglycerides, which are oxidized as a source of heat energy in a large number of mitochondria in the brown fat cells. A large number of UCP1 protein channels carry the electron transport chain and the oxidative action continues to occur without phosphorylation, i.e., no adenosine triphosphate is produced, which is also called as the granulocyte-decoupling respiration 10,11 . This reaction also promotes the consumption of nutrients and oxygen, but the free energy released during the process is lost in the form of heat. However, it is still unclear whether the regulation of sympathetic activation balances with the proliferation or differentiation of the precursor cells.
Recent studies have shown that UCP1 plays an important role in fatty acid and glucose utilization. β-Adrenergic receptor promoters (CL316, 243); peroxisome proliferator-activated receptor gamma (PPAR-γ) promoters; miRNAs; and genes such as PRDM16, BMP7, and PGC1α are currently known to induce fat browning 12 –17 . Stimulating energy consumption can increase the metabolic rate by increasing the brown fat accumulation in the body.
In addition, recently published articles have shown that tofacitinib (Janus kinase, JAK inhibitor), adenosine (acts on A1 and A2A receptors), and proanthocyanidin (polyphenols found in various plants) can activate fat browning. White fat cells treated with tofacitinib show increased UCP1 expression and increased glandular activity. Further, a study showed that two inhibitors of JAK activity conferred brown-like metabolic activity to white adipocytes. These metabolically converted adipocytes showed elevated UCP1 expression and increased mitochondrial activity. The repression of interferon signaling and activation of hedgehog signaling in JAK-inactivated adipocytes contributed to the metabolic conversion in these cells 18 . Pfeifer et al. found that adenosine can activate brown adipocytes—the key to signal transmission is the A2A receptor. Thus, pharmacological stimulation of A2A receptors or injection of lentiviral vectors expressing the A2A receptor into white fat induces the formation of brown-like cells, called the beige adipocytes. Adenosine-A2A signaling plays an important role in sympathetic BAT activation and offers protection from diet-induced obesity 19 .
Proanthocyanidin has potent antioxidant and anti-inflammatory effects, but little is known about its effects on granulocyte function and energy metabolism. One study showed that at 5 h after feeding rats with proanthocyanidin mixed with lard through oral gavage, the gastrocnemius muscle and brown fat mitochondria showed higher oxygen consumption rate but lower levels of blood triglyceride, free fatty acid, glycerol, and urea. Proanthocyanidin significantly enhanced the expression of the BAT energy metabolism gene, PGC1-α, and regulated its action. The citrate cycle of the enzyme-active transport chain was also upregulated, indicating that proanthocyanidin had a significant effect by increasing the oxidative capacity of the cells rapidly 20 .
Several small molecules extracted from Chinese herbs have been demonstrated to play a role in fat browning, such as flavanol, butylphthalide, berberine, curcumin, resveratrol, celastrol, bitter orange, isoquercitrin, ginger extract, β-lapachone, betulinic acid, cordycepin, platycodin D, liraglutide, and apigenin 21 –35 . All of these compounds can induce anti-inflammation, antioxidation, and antiobesity effects in high-fat diet-induced obese or streptozotocin (STZ) diabetic animal models.
Branched-Chain Amino Acids Control Energy Homeostasis in BAT
Branched-chain amino acids (BCAAs) are amino acids that have an aliphatic side chain with a branch, such as valine (Val), leucine (Leu), and isoleucine (Ile), and these BCAAs are essential amino acids. Therefore, BCAAs can only be obtained from the daily diet. BCAAs promote protein synthesis in the skeletal muscle, thereby increasing glucose uptake, and reduce fatigue and muscle soreness during exercise 36 . BCAAs are metabolized into the molecules for the tricarboxylic acid (TCA) cycle in the adipose tissue; therefore, the accumulation of circulating BCAAs is strongly associated with insulin resistance.
Studies have shown that animals fed with high-fat diets supplemented with BCAAs showed the same insulin resistance as animals fed with only high-fat diets. Pair-feeding with standard chow with/without BCAAs did not cause insulin resistance. The mechanism of action of BCAAs was mediated via increased levels of phosphorylated mTOR, JNK, and IRS1Ser307 and by the accumulation of acylcarnitine in the muscle 37 . These findings indicated that BCAAs contributed to the development of obesity-related insulin resistance. Herman et al 2 published that BCAA metabolism modulated the circulating BCAA levels in the adipose tissue. In obese and insulin-resistant humans, the overexpression of GLUT4 specifically in the adipose tissue resulted in the downregulation of the BCAA metabolizing enzymes. Higher levels of circulating BCAAs in the plasma are caused by decreasing BCAA oxidation rates in the adipose tissue.
Obese individuals are classified into three subgroups: lean healthy (LH), metabolically healthy obese (MHO), and metabolically unhealthy obese (MUO). In subcutaneous adipose tissue gene expression profile analysis, the expression of genes related to BCAA catabolism and the TCA cycle were mildly decreased in MHO individuals compared with those in the MUO individuals. MHO individuals have an intermediate amino acid homeostasis and they are more positively associated with homeostasis model assessment-insulin (HOME-IR) compared with LH and MUO individuals 38 .
The mechanism of action underlying the development of diabetes due to BCAA catabolic flux can be attributed to 3-hydroxyisobutyrate, a catabolic intermediate of valine, which is secreted from the muscle cells and activates endothelial fatty acid transport. Subsequently, fatty acid uptake is stimulated in vivo and lipid accumulation is promoted in the muscle, leading to insulin resistance in mice 39 . However, the role of systemic protein and amino acid metabolism in BAT has not been elucidated. Kajimura et al. reported that increasing utilization of BCAAs resulted in BAT thermogenesis and clearance of the circulating BCAAs. However, the specific deficiency of mitochondrial BCAA enzymes such as the branched-chain alpha-keto acid dehydrogenase complex induces obesity and glucose intolerance (Fig. 1). BCAA catabolism is mediated though a mitochondrial transporter, SLC25A44, as a starter for BCAA oxidation 40 . This mechanism of action shows the functional role of BCAA in thermogenesis and its applicability in the clinical treatment of obesity.

The accumulation of BCAAs in the blood vessels results in insulin resistance. BCAA catabolism in BAT enhances thermogenic activity because the upregulation of branched chain aminoacid amino-transferase and branched-chain alpha-keto acid dehydrogenase facilitate the oxidation. In the skeletal muscle, the utilization of BCAAs increases the glucose uptake and reduces the fatigue and muscle soreness. Increasing the clearance of BCAAs is a potential strategy for improving diabetes. BAT: brown adipose tissue; BCAA: branched-chain amino acid.
Lipid as a Regulator of Thermogenic Fat Activation
Fatty acid uptake is a type of fuel in BAT to promote thermogenesis; therefore, lipids must be the key mediators of communication among different organs. In 2017, a study showed that the activation of the β-adrenergic receptor by cold exposure induced the secretion of 12,13-dihydroxy-9Z-octadecenoic acid (12,13-diHOME). This study also showed that 12,13-diHOME was formed by hydrolysis. This reaction was catalyzed by soluble epoxide hydrolases (sEHs), especially Ephx1 and Ephx2 of four sEH-encoding genes, which are the major isoforms expressed in the adipose tissue 41 . 12,13-diHOME can be an autocrine or a paracrine stimulator that promotes fatty acid translocation by fatty acid transporter proteins, including CD36 and fatty acid transport protein 1, both of which are hormone-sensitive fatty acid transporters 42 . The acylcarnitine produced in the liver is transcribed from hepatocyte nuclear factor 4 alpha. Plasma acylcarnitine is uptaken by brown adipocytes in response to cold for adaptive thermogenesis 43 .
Dietary recommendations have considered that insufficient uptake of omega-3 polyunsaturated fatty acids (PUFAs) and excessive intake of omega-6 are associated with overweight/obesity. PUFA supplementation has a beneficial effect on the thermogenic function of adipocytes, and the degree of thermogenicity is affected by the levels of the individual oxylipins, which are controlled by the amount of lipids present 44 .
Sustarsic et al. 45 showed that the synthesis of the mitochondrial phospholipid cardiolipin is the major effector of the thermogenesis in brown adipocyte and beige cells (brown-like adipocytes) exposed to cold environments. Cardiolipin as a key effector triggers the thermogenic process and regulates the mitochondria-to-nucleus cross talk through the endoplasmic reticulum stress factor. Conversely, the blockage of cardiolipin in brown and beige fat mitochondria causes insulin resistance. Alkylglycerol in breast milk maintains the beige adipose tissue in infants and children below 5 years of age and impedes the transformation of beige adipose tissue into WAT. Alkylglycerol induces the beige adipocyte phenotype in the adipose tissues in a macrophage-dependent manner, and its function in beige adipose tissue development is mediated by the platelet-activating factor via IL-6/STAT3 signaling activation 46 .
A recent study reported that peroxisomes perform essential functions in lipid metabolism, and the peroxisomal biogenesis factor Pex16 in the adipose tissue is required for mitochondrial fission, increased mitochondrial copy number, and function recovery. Plasmalogens are synthesized in the peroxisomes; the knockdown of the plasmalogen synthesis enzyme glyceronephosphate-O-acyltransferase causes Pex16 inactivation, thereby disturbing mitochondrial morphology and function 47 (Fig. 2). Several studies are focusing on the role of lipids in thermogenesis, thereby revealing the potential use of lipids in the treatment of obesity in metabolic diseases.

HNF4-α expression regulates the release of acylcarnitine from the liver. Acylcarnitine acts as a fuel source for thermogenesis. PRDM16 expression regulates peroxisomes to induce the secretion of plasmalogens to communicate with the mitochondria. 12,13-diHOME promotes CD36/FATP1 translocation to the cell membrane to increase fatty acid uptake. Breast milk alkylglycerols regulate beige fat development via IL-6 and STAT3 signals. Lipokines are a critical fuel source for thermogenesis. 12,13-diHOME: 12,13-dihydroxy-9Z-octadecenoic acid; FATP1: fatty acid transport protein 1; HNF4-α: hepatocyte nuclear factor 4 alpha.
AMPK Activator-Mediated Phosphorylated ACC is a Potential Signal Extension Regulated by the PGC-1α/PPAR-γ/UCP1 Pathway
AMPK plays a pivotal role in the energy homeostasis of the central nervous system and peripheral organs, serving as a cell energy sensor and regulator. Hypothalamic AMPK activation increases food intake by inducing appetite and decreases adaptive heat generation in adipose tissue to reduce energy expenditure, thereby maintaining energy balance 48 . Several metabolic hormones, such as leptin, adiponectin, ghrelin, and insulin, integrate with neural circuits and exert energy-regulating effects through hypothalamic AMPK. Although AMPK activation in peripheral tissues can promote fatty acid oxidation and insulin sensitivity, its chronic activation in the hypothalamus causes obesity by reducing hyperphagia in humans and rodents 49 .
In the last 2 years, about eight published articles showed that some small molecules could be considered as novel candidates for the treatment of obesity through AMPK activation (Table 2). Cordycepin increases the energy expenditure and adaptive thermogenesis and improves the insulin sensitivity. Cordycepin is regulated by promoting the UCP1/AMPK-dependent pathway to enhance WAT browning in high-fat diet-fed mice 30 .
List of AMPK of Regulators for Thermogenetic Function.
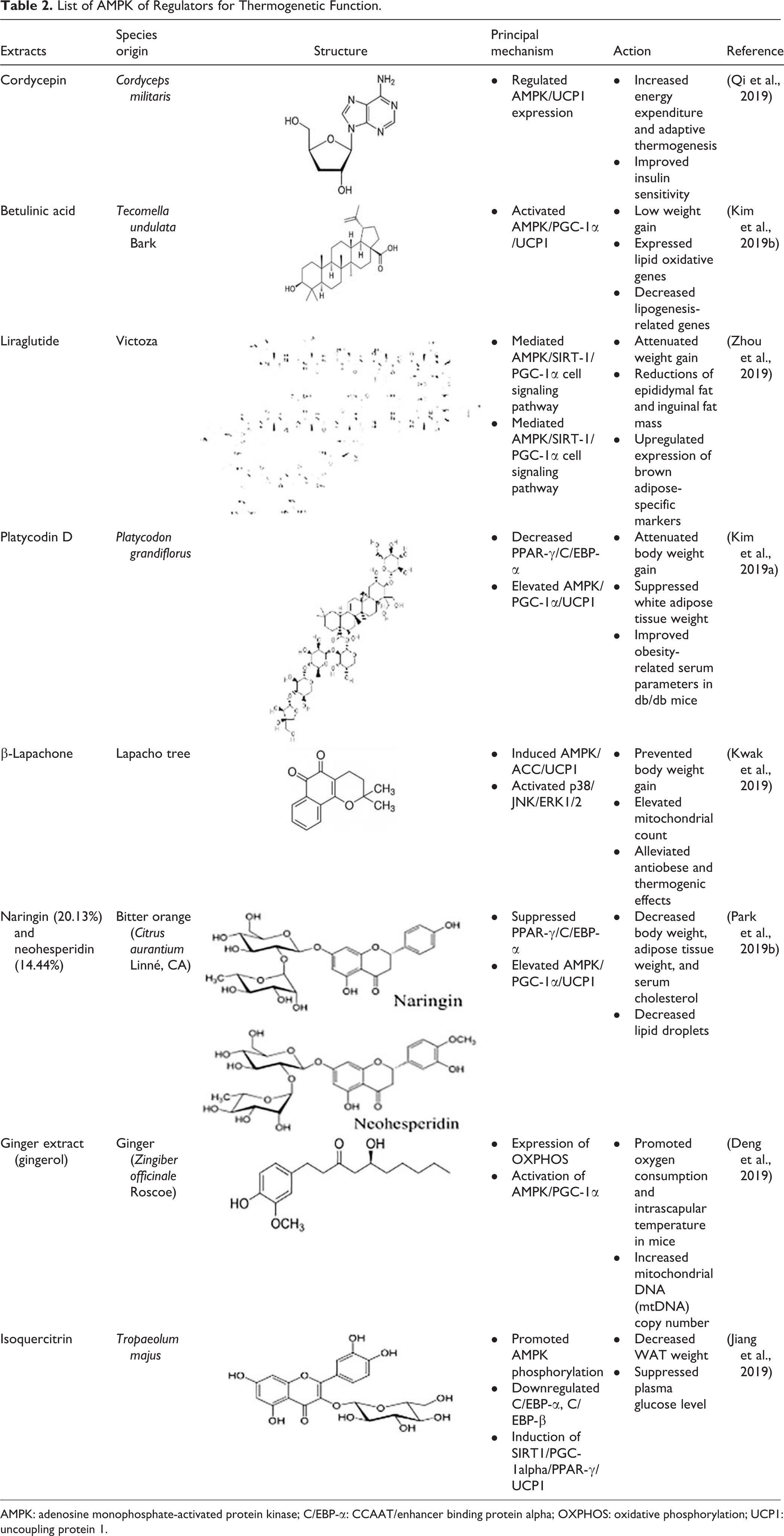
AMPK: adenosine monophosphate-activated protein kinase; C/EBP-α: CCAAT/enhancer binding protein alpha; OXPHOS: oxidative phosphorylation; UCP1: uncoupling protein 1.
Betulinic acid modulates energy expenditure, lipid oxidation, and thermogenesis in obese mice, and this is mediated by increased phosphorylated ACC/phosphorylated AMPK signals 25 .
Liraglutide is an agonist of the glucagon-like peptide 1 (GLP-1) receptor, and it upregulates brown adipose-specific markers in perigonadal fat in association with the upregulation of AMPK/SIRT-1/PGC-1α cell signaling; thus, liraglutide can be a potential medication for the prevention of obesity 35 .
Platycodin D treatment attenuated the body weight gain, suppressed the WAT mass, and improved the obesity-related serum parameters in db/db mice. The major mechanism of action was found to be decreased PPAR-γ and CCAAT/enhancer binding protein alpha (C/EBP-α) and activated AMPK both in in vivo and in vitro studies 24 .
β-Lapachone prevents body weight gain in Lepr−/− mice and increases the mitochondrial count in BAT and adipose-derived MSCs. β-Lapachone induces AMPK phosphorylation, but AMPK and its related signals ACC and UCP1 can be diminished by the AMPK inhibitor. Additionally, mitogen-activated protein kinase pathways are regulated by β-lapachone via p-38/JNK/ERK1/2. Thus, AMPK plays an important role in thermogenic activity 24 .
Bitter orange extracts, containing 20.13% naringin and 14.44% neohesperidin, administered by oral gavage in obese mice resulted in a significant decrease of body weight and serum cholesterol, while bitter orange-treated 3T3-L1 adipocytes showed decreased numbers of lipid droplets. Bitter orange extracts suppressed PPAR-γ/C/EBP-α but increased AMPK/PGC-1α/UCP1 activation, thereby suggesting its antiadipogenic and pro-thermogenic activity 29 .
Another study showed the effects of ginger extract on thermogenesis in BAT and mitochondrial biogenesis in the liver. They found that the major components of ginger, namely, 6-gingerol and 6-shogaol, promoted oxygen consumption and increased the interscapular temperature in mice; these components also increased the mitochondrial DNA copy number in the muscle and the liver. Ginger extracts activated the mitochondrial respiratory chain complex I and IV and promoted the expression of oxidative phosphorylation (OXPHOS), AMPK/PGC-1α, and related proteins. Thus, ginger consumption can improve mitochondrial function and can be useful for the treatment of metabolic syndrome 22 .
Isoquercitrin has been reported to decrease WAT weight, promote thermogenic function, and suppress plasma glucose levels. Isoquercitrin mediates the downregulation of C/EBP-α and C/EBP-β and induces SIRT1/PGC-1α/PPAR-γ/UCP1 via AMPK activation 23 (Fig. 3).
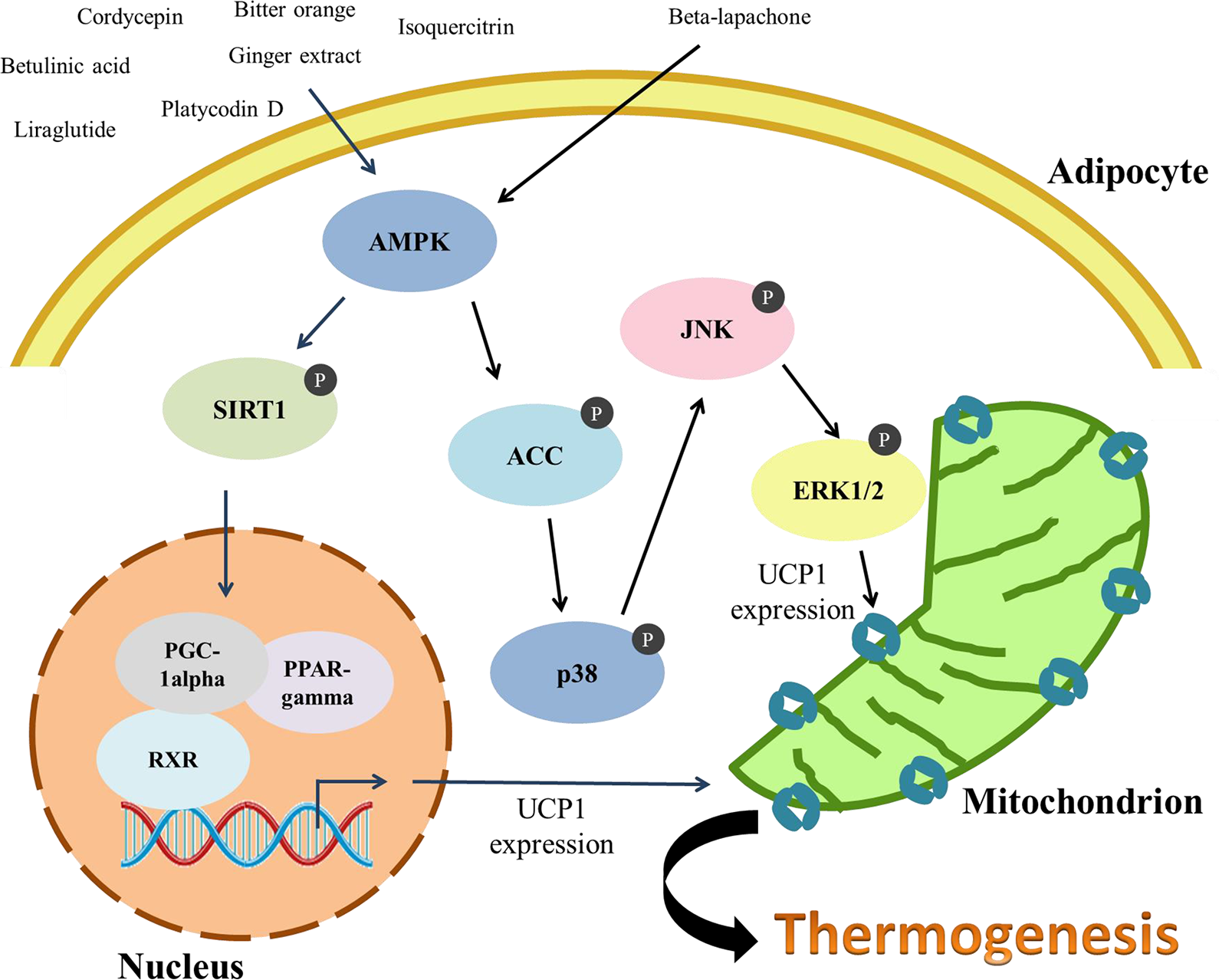
Several small molecules induce SIRT1/PGC-1α/UCP1 to mediate thermogenesis. β-Lapachone phosphorylates AMPK/ACC/p38/JNK/ERK1/2, increases the mitochondrial count, and increases the antiobese and thermogenic effects. All the small molecules listed in Table 2 can improve insulin sensitivity and increase energy expenditure. ACC: acetyl-CoA carboxylase; AMPK: adenosine monophosphate-activated protein kinase.
In summary, several small molecules can inhibit adipogenesis and promote adipogenic thermogenesis. Increasing number of evidences show that AMPK is involved in the signal pathway. All the studies show that AMPK could be an effective target for treating obesity and the associated disorders.
Use of AMPK Activators in Clinical Trials for the Treatment of Metabolic Syndrome
Currently, five drugs targeting AMPK are being tested in clinical trials for the treatment of obesity and metabolic syndrome (Table 3).
AMPK as a Target for Antiobesity and Metabolic Syndrome in Clinical Trials.

AMPK: adenosine monophosphate-activated protein kinase; GLP-1: glucagon-like peptide 1.
Clinical Trial 1: The consumption of genistein from soy for 2 months has been found to improve insulin resistance and modify the gut microbiota taxonomy in rodents and humans with metabolic diseases. Increasing AMPK phosphorylation and increased gene expression of the fatty acids undergoing oxidation were considered as the mechanisms of action of the diet consumed by the patients (NCT04105023).
Clinical Trial 2: GLP-1 injected subcutaneously in isolated adipocytes could induce both lipogenic and lipolytic mechanisms in WAT. GLP-1 agonists also stimulated BAT thermogenesis and browning through hypothalamic AMPK activation in animals (NCT02170324).
Clinical Trial 3: The participants were administered a dietary supplement containing Hibiscus sabdariffa L. and Lippia citriodora L. extracts as capsules. This formulation could significantly activate the AMPK enzyme and reduce triglyceride accumulation in the hypertrophied adipocyte model (NCT03568877).
Clinical Trial 4: Many studies have shown that C1q/TNF-related protein 9 (CTRP9) exerts beneficial effects on metabolic and cardiovascular homeostasis. CTRP9 levels were detected in obese pre-eclamptic pregnant women and nonobese pre-eclamptic pregnant women, but results have not been published yet. CTRP9 levels were found to be reduced in patients with diabetes and obesity (NCT03650790).
Clinical Trial 5: NCT03342742 is an active clinical trial that is recruiting participants currently. The investigators have novel preliminary data that overweight and obesity are independently associated with substantially faster kidney growth in patients with autosomal dominant polycystic kidney disease. In this trial, the participants will have a restricted caloric diet to reduce the energy intake, and the body weight and AMPK signaling intervention will be analyzed.
In summary, we have discussed the role of AMPK in adipose tissues in five clinical trials, with AMPK related to regulating the carbohydrate and lipid metabolism, adipogenesis, and proinflammatory effect in physiological and pathophysiological conditions. Our review discusses a large number of studies wherein AMPK activity is specially manipulated for further characterization of its role in the constituent cells of the adipose tissues and the potential use of this activity for future therapeutic strategies.
Conclusion
Undoubtedly, in recent years, pandemic obesity and diabetes have been some of the emerging and challenging diseases, but there is no powerful and effective drug for their treatment to date. Our review article has discussed studies that have focused on the activation of BAT in the body, thereby increasing the metabolic function, as a suitable therapeutic approach for the treatment of obesity and its associated metabolic disorders.
Small molecules such as proteins, amino acids, and lipids can also be considered as regulators of BAT thermogenesis. Chinese herbal extracts that have anti-inflammatory effects have been found to activate thermogenesis. Moreover, recent studies have pointed out that hormones secreted by brown adipocytes, which can promote the release of different cytokines, lipokines, or activate protein translocation, increase fatty acid oxidation and glucose utilization. While BCAAs are positively correlated with insulin resistance, the clearance of BCAAs improves the metabolism and promotes energy expenditure.
AMPK regulates the energy homeostasis of several cells. It activates glucose and fatty acid uptake and oxidation when energy is not in conformity with the requirements in the cells. AMPK has been established to be directly involved in the regulation of energy homeostasis in BAT. Several studies have showed the efficacy of activating downstream molecular signals via AMPK to achieve thermogenic effects. In current clinical trials, oral administration, peptide injection, and caloric restriction activate AMPK, thereby delaying body weight gain, lowering plasma parameters, and increasing metabolic activity, and activating thermogenesis. In the future, the development of specific drugs targeting obesity and the associated metabolic disorders is of prime importance because of the increase in the incidence of mortality due to obesity.
Footnotes
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) received financial support for the research, authorship, and/or publication of this article: This project was supported by the Buddhist Tzu Chi Bioinnovation Center, Buddhist Tzu Chi Medical Foundation, Hualien, Taiwan (project no. MF00A130SS02), and the Graduate Institute of Biomedical Science, China Medical University, Taichung, Taiwan, and Ministry of Science and Technology, Taiwan (MOST 106-2320-B-303-001-MY3 and MOST 106-2320-B-303-002-MY3). The authors also thank the Department of Medical Research, Hualien Tzu Chi Hospital, for funding this study.