
Abstract
The term episomal induced pluripotent stem cells (EiPSCs) refers to somatic cells that are reprogrammed into induced pluripotent stem cells (iPSCs) using non-integrative episomal vector methods. This reprogramming process has a better safety profile compared with integrative methods using viruses. There is a current trend toward using episomal plasmid reprogramming to generate iPSCs because of the improved safety profile. Clinical reports of potential human cell sources that have been successfully reprogrammed into EiPSCs are increasing, but no review or summary has been published. The functional applications of EiPSCs and their potential uses in various conditions have been described, and these may be applicable to clinical scenarios. This review summarizes the current direction of EiPSC research and the properties of these cells with the aim of explaining their potential role in clinical applications and functional restoration.
Introduction
Application of the reprogramming techniques first developed by Yamanaka et al. 1 has made possible the conversion of somatic cells to pluripotent stem cells that resemble embryonic stem cells (ESCs). ESCs have been touted as the “Holy Grail” for unrestricted regeneration because of their potential to differentiate into any cell lineage in the body and to replace damaged tissue. Opponents of this technique commonly cite the potential for unethical use or donation of ESCs, which are potential sources of life and embryo formation. With the development of a method to create induced pluripotent stem cells (iPSCs), it is possible to harness the regenerative properties of iPSCs, which resemble ESCs, yet without the ethical controversies associated with the sources of ESCs.
For research purposes, iPSCs can be readily cultured in the laboratory from various somatic cells. Somatic cells of various lineages have been shown to be capable of changing both their morphology and pluripotent potential through the overexpression of four main pluripotent factors: Oct3/4, Sox2, Klf4, and c-Myc (OSKM). The replacement of c-Myc and Klf4 by Nanog and Lin28 has also been shown to be possible when used in conjunction with Oct3/4 and Sox2 during the reprogramming of cells into iPSCs 2 . Overexpression of Oct4, Sox2, and Nanog can also reprogram human fetal gut mesentery-derived cells into iPSCs 3 . Although Nanog is a dispensable reprogramming factor 4 , it has been reported to be essential for the ability for self-renewal 5 and generation of stable iPSCs 6 . In some cases, such as adult mouse neural stem cells, expression of only one factor (Oct4) is sufficient for the generation of iPSCs 7 , even using episomal reprogramming 8 . A possible reason is the endogenous expression of Sox2, c-Myc, and Klf4 in neural stem cells. In addition, downregulation of p53 using knockdown 9 and knockout 10 methods can markedly improve the efficiency of iPSC generation 11 , and only Oct4 and Sox2 are sufficient for iPSC generation under conditions of p53 loss 12 . One possible mechanism to explain the suppression of iPSC generation by p53 is through the inhibition of the expression of Nanog and Oct4 13 . Certain somatic and adult stem cells, such as keratinocytes 14 and dental pulp stem cells 15 , have a greater propensity to be reprogrammed into iPSCs compared with fibroblasts.
The introduction of these pluripotent reprogramming techniques can be divided into integrative and non-integrative methods. When first discovered, viral transduction was used for the insertion of the OSKM genes into the somatic cell genome. However, this may cause disruption to the host’s genome and, with the use of the proto-oncogene c-Myc, gene reactivation could increase the risk of transgene-derived tumor formation 16,17 . Other methods have been used to reprogram somatic cells into iPSCs. In particular, non-integrative methods such as episomal plasmid delivery of pluripotent OSK genes without c-Myc have been touted as a safer alternative for iPSC generation 18,19 . Episomal plasmid vectors are also combined with p53 knockdown to generate iPSCs 10,20,21 . This review focuses on studies of the use of iPSCs generated using episomal plasmids, which are termed “episomal iPSCs” (EiPSCs) to reflect their origin.
Integrative Methods of iPSC Production
Potential issues with integrative methods of iPSC production include the inadvertent introduction of potentially harmful viral components that express certain oncogenes that may lead to tumor formation. Insertion of genes also carries the risk of disrupting the expression of host cell tumor suppressor genes, especially if the insertions occur in an open reading frame or alter the expression of oncogenes in close proximity 17 . Several viral systems have been introduced to circumvent this issue such as Cre-deletable 22 or inducible lentiviruses, which reduce the risk of integration, although concerns have been raised about the use of viral vectors for therapeutic applications. Cre-deletable lentiviruses refer to a Cre recombinase-mediated deletion of a viral genome once integration is complete. This allows for the time-controlled integration, which is self-limiting and hence minimizes the chance of permanent viral genomic integration. Inducible lentivirus commonly refers to a tetracycline- or doxycycline-inducible expression system in lentiviruses, which allows the activation or deactivation of viral genomic DNA in the presence of tetracycline. This also provides some control over the duration of the viral genomic presence and, therefore, the risk of integration with the host genome.
Non-Integrative Methods of iPSC Production
The use of plasmids for the introduction of OSK pluripotent transcription factors has been described, and this method has been successful for reprogramming somatic and adult stem cells. The use of integration-defective viral delivery systems (adenoviruses 23 , Sendai viruses 24 ), piggyBac systems 25 , minicircle vectors 26 , episomal delivery 19 , mRNA delivery 27 , protein delivery 28 , and chemical induction 29 in the production of iPSCs has been described. The piggyBac transposition system allows the direct cutting and pasting of sections of DNA to allow insertion and removal of certain sections. Specific deletion and insertion of reprogramming genes can be performed. Minicircle vectors are small circular plasmid derivatives that are freed from all prokaryotic vector parts or bacterial plasmid DNA. However, these are not in widespread use because of the intensive production process. The delivery of mRNA and protein can be much more costly for experimental purposes, and the mRNA or protein is not always inducible downstream. Chemical induction uses specific sets of chemicals and mutagens to allow the reprogramming of cells. By subjecting cells to a specific set of chemicals, researchers have shown that cells can be reprogrammed to their pluripotent state. However, this method is extremely laborious and has unknown safety parameters at present. Episomal plasmid delivery is cost effective, and the plasmid does not integrate when used with common transfection methods. This method provides an effective method for the delivery of plasmids into cells for reprogramming 30 . The plasmids are readily available on multiple gene platforms and are removed from the host cell through cell division and serial dilution. The use of non-integrative methods of iPSC production has the advantage of producing iPSCs that are free of transgene integration. This has been confirmed by polymerase chain reaction (PCR) analysis, which shows no residual transgenes of viral origin 31 . For example, the Yamanaka group reported that no residual episomal plasmid DNA could be detected in clones after 11–20 passages 20 .
Two components of the oriP and Epstein–Barr nuclear antigen-1 (EBNA-1) are also used widely in episomal plasmids 32 –38 . Yu et al. reported that human iPSCs that are completely free of plasmids and transgene sequences could be derived from fibroblasts by a single transfection with oriP/EBNA-1-based episomal plasmids 39 –42 . The footprint-free iPSCs make them safer for clinical application because of the loss of plasmids and transgenes. OriP/EBNA-1 plasmids have a wide host cell range for episomal reprogramming, and a single transfection of episomal plasmids is sufficient for iPSC generation. Compared with the original episomal plasmids without EBNA-1, oriP/EBNA-1 can improve the efficiency of iPSC generation 43 through the oriP/EBNA-1-mediated nuclear import and retention of vector DNA 44 . It can replicate only once per cell cycle. Episomal DNA is lost from cells at a rate of 5% per cell generation because of defects in plasmid synthesis and partitioning 45 . Subsequently, episome-free iPSCs can be easily harvested. Although plasmids appear in the first few cell passages immediately after transfection, many studies have used PCR analysis to show complete loss of plasmids and transgenes during extended cell culture 20,39 –42,46 –58 . The time point of successful loss of episomes varies between different somatic cell types. The oriP/EBNA-1-based episomal plasmids have been proven to generate iPSCs very efficiently without the risk of transgenic sequences inserted into the somatic cell genome. Certain comments exist saying iPSCs generated using episomal plasmids with EBNA-1 expressed have residual episomal DNA. This does not appear to be true after reviewing the data presented by Yu et al. 39 , where the group demonstrated that in fact that there were no residual plasmids and transgenes in iPSCs generated by EBNA-based plasmids using PCR and Southern blot analysis.
In an interesting experiment, the efficiency of reprogramming was compared between various methods for generating iPSCs. Reprogramming success rates were similarly high, at around 80%, with Sendai-viral, episomal, and lentivirus methods. mRNA methods alone were reported to have a lower success rate because of massive cell death and detachment. Episomal methods seem to be a good method for producing iPSCs, and the materials can be manufactured using current good manufacturing practice compatible processes (cGMP). As such, episomal methods remain very useful in the clinical setting 59 .
Human EiPSC Sources
Multiple cell lineages from humans have been reprogrammed into iPSCs using episomal plasmids. Episomal plasmid reprogramming differs markedly from other forms of reprogramming techniques, such as retroviral transduction, and varied success rates have been reported. Recent trends include the increasing use of episomal plasmid methods for reprogramming various types of human cells for research use and potential clinical applications. The human cells reported to have been successfully reprogrammed into EiPSCs include fibroblasts, epithelial cells, keratinocytes, mononuclear cells from adult peripheral blood, cord blood cells, amniotic fluid stem cells, mesenchymal stromal cells, lymphoblasts, lamina propria progenitor cells from oral mucosa, and urothelial cells obtained from urine. A summary of these reported human EiPSC sources is shown in Table 1 8,20,21,32 –43,47 –52,55,56,58,60 –207 . The variety of cell lineages that can be reprogrammed by episomal techniques demonstrates the versatility of this technique, which has been used in many laboratories around the world.
Summary of Reported Human EiPSC sources available in the Literature.

The ease of obtaining several of the human cell types, such as those in peripheral blood or urine, for reprogramming make this method less invasive for donors, and there is an equivalent success rate for producing EiPSCs of similar nature. Obtaining EiPSCs from donors with known specific genetic manipulations also allows for the establishment of cell lines for further research, especially with regards to future developments of gene therapy for these specific conditions. Episomal plasmids have been used to generate EiPSCs from various somatic cells of differing origins. Human mesenchymal stromal cells have been developed into EiPSCs through the episomal plasmid-based expression of Oct4, Sox2, Nanog, Lin28, SV40LT, Klf4, and c-Myc 60 . Other sources reported include human fetal foreskin fibroblasts, and CD34+ cells from cord and peripheral human blood, which shows that peripheral blood mononuclear cells may be a good source of cells for iPSC reprogramming especially because of the low invasiveness when obtaining samples.
Gene Delivery Methods
Delivery of plasmids into cells for reprogramming has been described. These methods include electroporation techniques (using Nucleofector kits), liposomal magnetofection, and Lipofectamine transfection reagents. Liposomal transfection using magnetofection 208 or Lipofectamine refers to the delivery of plasmid DNA via liposomes and allows the merging of cationic liposomes carrying the DNA into the cells. Electroporation is the use of an electric current across cell membranes, which forcibly opens their channels to allow entry of reagents. This method has been described widely and has a higher rate of delivery efficiency but is known to cause cell damage 209 . The use of non-liposomal transfection reagents for episomal plasmid delivery, such as FuGENE HD, has also been reported to have good transfection efficiencies 210 . The company produces a proprietary formula that is touted as non-liposomal but can still deliver DNA and plasmids into cells and therefore works as a transfection kit. Recent work has also shown promise in producing EiPSCs using small molecules instead of feeder cells. A cocktail of molecules has been described for the reprogramming of human somatic cells to iPSCs. These include the MEK inhibitor PD0325901, GSK3β inhibitor CHIR99021, TGF-β/activin/nodal receptor inhibitor A-83-01, ROCK inhibitor HA-100, and human leukemia inhibitory factor 40 .
In Vivo Animal Model Applications
Cardiogenic Regeneration
Much attention has focused on the use of iPSCs in cardiac regeneration especially after cardiac infarction, which causes loss of cardiomyocytes that cannot regenerate and are replaced with scar tissue. Both EiPSCs and iPSCs have been differentiated into cardiomyocytes, which shows their potential use in both autologous and allogeneic therapies. A recent study demonstrated that allogeneic EiPSCs cultured from cynomolgus monkeys, when differentiated into cardiomyocytes and injected intramuscularly infarcted cardiac muscle, induced remuscularization of infarcted muscle tissue. Fibroblasts obtained from the monkeys were reprogrammed using episomal plasmids into EiPSCs, and the EiPSCs-derived cardiomyocytes were then injected into the infarcted cardiac muscle. After a clinical regimen of immunosuppression using methylprednisolone and tacrolimus, the hearts showed improvement in cardiac contractile function without any signs of rejection on postoperative week 12 211 . The results are promising in showing that direct application of EiPSCs-derived cardiomyocytes is possible. The local environment and conditions under which the EiPSCs were directly injected allowed for their direct use and differentiation according to clinical need. A diagram of the potential application for an EiPSCs-engineered cardiac cell sheet is shown in Fig. 1.
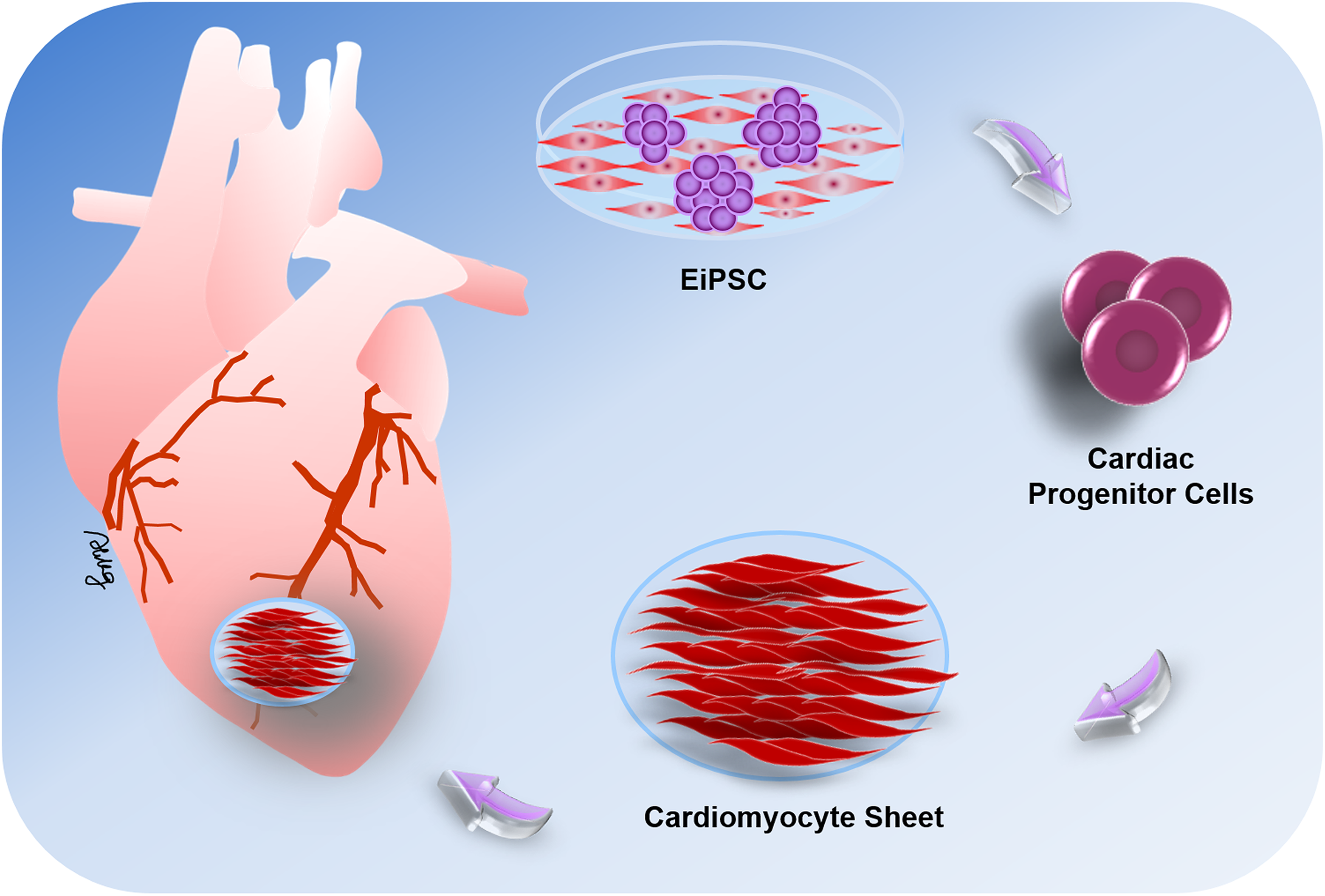
The potential application for cardiac cell sheet strategies using EiPSC-derived cardiomyocytes. EiPSCs can be differentiated into cardiac progenitor cells, which are then induced to form cardiomyocytes in vitro. These cardiomyocytes can then be organized into a cell sheet and applied to damaged areas of cardiac muscle in vivo via intracoronary or intracardiac injections or epicardially by tissue-engineered cardiac patches. The cell sheets exhibit regenerative capabilities and induce the restoration of cardiac function after muscle damage.
One problem with bioengineered tissue is that it cannot be used to create a large structure, which requires thorough oxygenation, because of the lack of vascularization in the bioengineered construct. EiPSCs were reported to regenerate vascular tissue if some were first converted to patient-specific cardiovascular progenitor cells, which then differentiated into vascular smooth muscle cells to make up the vascular scaffold present in blood vessels. This new development heralds the potential for integration and creation of larger bioengineered constructs that can become vascularized. This suggests the potential ability to design whole organs with vascularized networks made from the patient’s cells, which are then attached using conventional surgical methods. This may allow the organ to be manufactured in the laboratory and vascularized 61 .
Peripheral Nerve Regeneration
EiPSCs have shown promise in promoting the regeneration of peripheral nerves in a mouse sciatic transection model 212 . Transection or neurotmesis of peripheral nerves is notoriously difficult to recover and usually leads to wasting of motor end plates, muscle atrophy, and functional loss, which markedly impairs the patient’s quality of life. In this mouse model, undifferentiated EiPSCs were applied to the transected ends of the sciatic nerves after coaptation of both ends by suturing. Compared with the negative control without cell administration, sciatic nerves treated with EiPSCs displayed significantly faster axonal regeneration and a ration of the degree of myelination to axonal diameter. These positive changes were similar to those observed in the ESC group, which acted as a positive control. The results of this study demonstrate the neuroregenerative potential of EiPSCs. One possible mechanism includes the increased expression of neutrotrophin-3, a neuronal growth factor, which can accelerate axonal regeneration and myelination. Direct application of EiPSCs to the site of injury and nerve transection presumably allowed the EiPSCs to act through a paracrine mechanism due to its direct effect and fast nature; they probably differentiate but rather, when applied to the environment, promoted sciatic nerve recovery through the upregulation of neutrotrophin-3 and subsequent secretion of neuronal growth factor by the EiPSCs themselves. The diagram in Fig. 2 shows a depiction of the actions of EiPSCs on mouse transected peripheral nerve regeneration.

Topical application of EiPSCs to transected peripheral nerves. After surgical repair of transected peripheral nerves in a mouse sciatic nerve model, axonal regeneration was accelerated by topical application of EiPSCs to the site of injury. The increased production of neurotrophic factor-3 as a growth factor was one of the causes of acceleration of axonal growth and maintenance of muscle function and gait. Compared with negative controls without cell administrations, the regenerated axons exhibited a higher quality of myelination and more cells were obtained.
Ischemic Stroke Therapy
Mouse embryonic fibroblasts reprogrammed into EiPSCs using episomal plasmid transfection were delivered and used to treat mice in an ischemic stroke model 213 . To avoid oncogenic and virus integration, while generating EiPSCs, two expression plasmids, Oct4 and Sox2, were repeatedly transfected into fibroblasts under hypoxic condition. The EiPSCs were first differentiated into neural precursor cells before being injected into the brain of mice after the induced ischemic stroke. The authors observed evidence of the differentiation of precursor cells into neurons and astrocytes. They concluded that these observed changes resulted in better behavioral recovery including locomotor activity, beam walking, and rotarod movement compared with control mice.
Smooth Muscle Regeneration
The use of EiPSCs in the regeneration of smooth muscle cells for the treatment of stress-induced urinary incontinence has also been described 214 . Inadequate muscle sphincter function is a possible cause for urinary incontinence, and regeneration of smooth muscles at this sphincter may increase sphincter tone and hence urinary continence and control. The group here conducted experiments in rat urethral sphincters which were surgically weakened, resulting in urinary incontinence. Smooth muscle precursor cells were then differentiated from human EiPSCs and injected periurethrally to enhance muscle tone. Leak pressure and sphincter muscle electromyography were measured as markers of recovery. The group with the EiPSCs-derived smooth muscle injection showed recovery of the sphincter compared with the control without cell administration, which suggested that this method of treatment may be possible for restoration of urethral sphincter function.
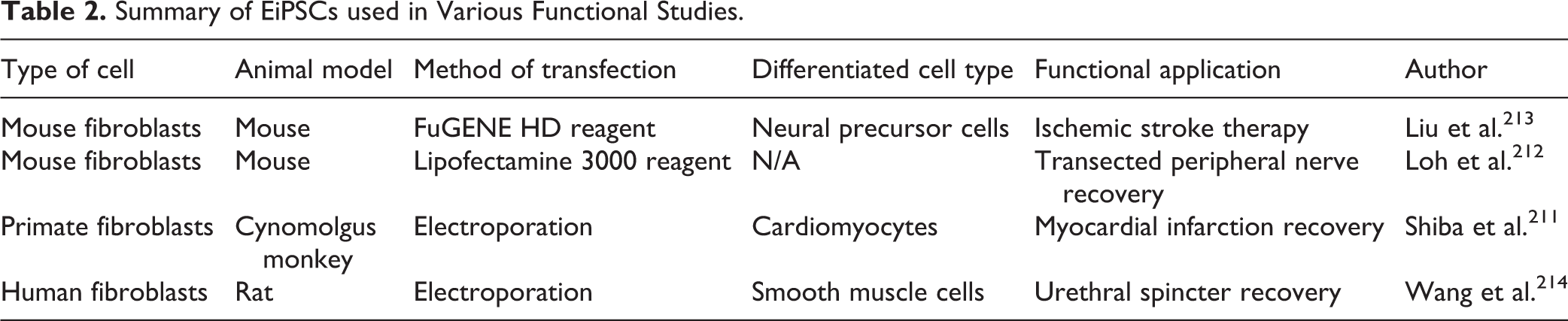
EiPSCs currently have the potential for cardiac regeneration to replace damaged myocardium. The topical application of cells to regenerate a limited area appears promising, but reconstructing an entire cardiac structure requires further bioengineering advances to deliver blood supply to the entire organ. The use of EiPSCs in peripheral nerve regeneration can improve transected nerve recovery. Direct applications after surgical repair can enhance recovery through nerve growth factor secretion. In ischemic stroke therapy, EiPSCs may have potential for improving the recovery of damaged neural cells in the brain by differentiating into neurons and astrocytes, which should result in better motor recovery. EiPSCs can also play a role in regeneration of smooth muscle cells, leading to restoration of muscle sphincter function in urinary incontinence. A summary of the EiPSCs used in various functional studies is shown in Table 2 211 –214 .
Summary of EiPSCs used in Various Functional Studies.
Clinical Trials
Current clinical trials known at the time of writing this review all involve iPSCs derived from retroviral transduction. A trial of the replacement of retinal pigment epithelium cells (RPEs) in age-related macular degeneration (ARMD) was recently continued and is in progress. The human iPSCs used in this case were derived from retroviral reprogramming 215 . Human iPSC-derived RPE cell sheets were generated without any artificial scaffolds, express typical RPE cell markers, form tight junctions that exhibit polarized secretion of growth factors, and show phagocytotic ability and gene-expression patterns similar to those of native RPE cells. The monolayer cell sheets have potential use as a graft for tissue replacement therapy for ARMD.
The trial was temporarily halted because spontaneous genetic mutations were found in the generation of iPSCs 216 . The spontaneous mutations comprised six mutations, in which three genes had been deleted and another three nucleotides changed. One of the mutations was an “oncogene,” which has a low-risk link to cancer. None of these mutations were present in the patient’s original DNA makeup. The appearance of mutations was deemed to be either the result of the iPSC induction procedure or the presence at undetectable levels in the patient’s somatic skin cells initially. As the risk of carcinogenesis was low, the trial was continued as planned 217 .
Currently registered trials focus on platelet generation for treating various anemias. However, the research group faced the problem of the mass production of platelets required for effective clinical use. Other trials involve the differentiation of dopaminergic neurons piloted for use in Parkinson’s disease. Retinal ganglion cells are also being used for treatment of glaucoma and optic neuropathies. Except for the current trial in Japan of the use of RPE cells for wet ARMD mentioned above, all of these trials are in the preclinical or animal model stages 218 . Guidelines for further clinical trials for stem cell research involving patients and iPSCs were issued by the International Society for Stem Cell Research in 2016. These guidelines suggest that the donor cell procurement should be checked for iPSCs intended for use in human, and that these cells should be excluded from specialized reviews because they are now acknowledged to have different implications in the treatment of disease compared with human ESCs 219 .
Safety
The evidence for the safety of the use of EiPSCs in animal models is only now emerging, and there is a paucity of evidence in this area. Mice given EiPSCs at the site of the transected sciatic nerve displayed no formation of tumors locally at the site of application 212 . Distant sites and major organs were also examined histologically for the presence of tumors after 1 year of follow-up. Normal behavior and health of the mice were recorded, and no ill effects of EiPSCs were reported in this group. Another study found no aberrant growths from differentiated EiPSCs that were added to primate hearts regeneration after a myocardial infarction 211 . Neither macroscopic nor microscopic analysis revealed any evidence of tumor formation at 12 weeks after transplantation of the EiPSCs-derived cardiomyocytes. One possible reason is that the EiPSCs used were completely differentiated, with minimal residual EiPSCs present remaining after grafting, which resulted in no tumor formation. Because viral or integrative reprogramming techniques may alter gene expression, the use of episomal reprogramming techniques in the production of EiPSCs may reduce the risk of tumor transformation. Further evidence is required to understand more about the safety of the use of EiPSCs and to substantiate the hypothesized mechanisms of action. One possible future direction is to compare the formation of tumors in two groups—undifferentiated EiPSCs and iPSCs generated from other methods such as retroviruses grafted onto recipients. The possible lack of tumor formation seen in EiPSC-transplanted groups from such a study design would provide evidence of its safety.
Yamanaka Cell Bank: A Future in Autologous and Allo-iPSC Therapy
With the recent establishment of the Yamanaka stem cell bank at the RIKEN BioResource Center in Japan, several human iPSC cell lines have been produced in preparation for future research or clinical application. A switch to episomal methods of iPSC production is evident from the most recent human iPSC cell lines formed at the Yamanaka stem cell bank 220 . As mentioned previously, to enable use of the cells clinically, particular attention should be paid to the methods of iPSC production that favor episomal methods. The Yamanaka stem cell bank has successfully produced multiple human cell lines of EiPSCs using transformation of human cells from various types. EiPSCs have been produced from human skin fibroblasts (cell numbers HPS0076, HPS0077) 221,222 , human cord blood (cell numbers HPS0328, HPS0331), and human peripheral blood (cell number HPS0360) 43 . Other than the two initial cell lines produced from human, which were obtained using retroviral transduction without c-Myc, the five more recently developed human EiPSC cell lines were produced purely via episomal vectors. A diagram showing the potential for an EiPSC bank is depicted in Fig. 3.

The potential for an EiPSC bank. The pluripotent potential of EiPSCs allows them to differentiate into various cell lineages for repair and regeneration. Fibroblasts from healthy donors can be harvested and reprogrammed into EiPSCs and stored in a cell bank. These EiPSCs can then be differentiated into various cell lineages for repair and regeneration according to the needs of individual patients. So far, these cells include cardiomyocytes, hematopoietic progenitor cells, neural cells, adipocytes, and pancreatic islet cells. Each of these cells can be used to replace damaged cells in patients and provide a novel therapeutic potential for each clinical scenario. For any allogeneic transfer of EiPSCs, MHC mismatch typing can first be performed to minimize any chance of MHC mismatch incompatibility before selecting the least antigenic EiPSC bank sample to be transferred to the patient.
The immunogenicity of iPSCs generated remains an unknown area of research. A recent study showed a possible immune response toward smooth muscle cells derived from iPSCs but not the RPE from iPSCs 223 . Possible strategies for bypassing a possible immune response include the development of humanized iPSCs-derived RPE cells for transplantation. In a humanized mouse model, Zhao et al. found that the smooth muscle cells induced a strong antigenic response by the host’s immune system which was not evident when RPE cells were used. The potential use of immunosuppression must be considered for the clinical use and clinical trials of iPSCs. If there is evidence of rejection of tissue as for a foreign host, immunosuppressants should be administered when the cells are delivered to the recipient. The use of iPSCs for treating Parkinson’s disease has been reported in Japan. However, certain clinical considerations, such as the safety and therapeutic use of iPSCs in patients with Parkinson’s disease, are needed. Clinical trials are underway for examining this aspect of iPSC use and treatment. Current rapid integration into its clinical use should continue to surface within the next few years 224 .
Conclusion
The future of EiPSCs lies in the increasing trend for the use of cell therapy in the treatment of various diseases because of their regenerative properties. Differentiating these cells before use or their direct application requires further investigation. The immunogenicity of allogeneic EiPSCs will also need to be determined for the tissue both derived and differentiated from these cells.
Footnotes
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported by grants from Chang Gung Medical Foundation, Chang Gung Memorial Hospital, Taiwan (CMRPG1F0082, CMRPG3F1431, CMRPG1H0081 and CMRPG1H0082) and Ministry of Science and Technology, Taiwan (MOST 108-2314-B-182A-009-).