
Abstract
Adoptive transfer of autologous polyclonal regulatory T cells (Tregs) is a promising option for reducing graft rejection in allogeneic transplantation. To gain therapeutic levels of Tregs there is a need to expand obtained cells ex vivo, usually in the presence of the mTOR inhibitor Rapamycin due to its ability to suppress proliferation of non-Treg T cells, thus promoting a purer Treg yield. Azithromycin is a bacteriostatic macrolide with mTOR inhibitory activity that has been shown to exert immunomodulatory effects on several types of immune cells. In this study we investigated the effects of Azithromycin, compared with Rapamycin, on Treg phenotype, growth, and function when expanding bulk, naïve, and memory Tregs. Furthermore, the intracellular concentration of Rapamycin in CD4+ T cells as well as in the culture medium was measured for up to 48 h after supplemented. Treg phenotype was assessed by flow cytometry and Treg function was measured as inhibition of responder T-cell expansion in a suppression assay. The concentration of Rapamycin was quantified with liquid chromatography coupled to tandem mass spectrometry (LC-MS/MS). Azithromycin and Rapamycin both promoted a FoxP3-positive Treg phenotype in bulk Tregs, while Rapamycin also increased FoxP3 and FoxP3+Helios positivity in naïve and memory Tregs. Furthermore, Rapamycin inhibited the expansion of naïve Tregs, but also increased their suppressive effect. Rapamycin was quickly degraded in 37°C medium, yet was retained intracellularly. While both compounds may benefit expansion of FoxP3+ Tregs in vitro, further studies elucidating the effects of Azithromycin treatment on Tregs are needed to determine its potential use.
Introduction
Adoptive cell therapy with immunosuppressive regulatory T cells (Tregs) raises hope for clinical applications in transplantation settings and for prevention of immunological disorders 1 –4 . Previous studies have shown promising results for prevention of graft-versus-host disease in allogeneic stem cell transplantation and in reducing beta cell destruction in newly onset type-1 diabetes 1 –3 .
Currently, the effects of adoptive Treg therapy in liver and kidney transplantation are under study at several clinics worldwide 4 . The exact numbers of Tregs needed to reach therapeutic levels have not been defined; however, previous trials suggest dose ranges between 0.1 × 106 and 20 × 106 polyclonal natural Tregs (nTregs) per kilo of bodyweight 1 –3,5 . In order to amass such cell numbers, in vitro expansion of nTregs is required prior to adoptive cell transfer 6 . However, Treg cultures risk contamination of T effector (Teff) cells and of Tregs converting to pro-inflammatory Th17 cells during expansion 7 –11 . The ideal in vitro culture condition of Tregs would therefore promote fast expansion of stable functional Tregs while reducing non-Treg contamination.
Rapamycin (RAP) is a macrolide with immunosuppressive properties used in transplantation settings for the prevention of graft rejection 12 . It is also utilized for improving in vitro cultures of Tregs due to its ability to suppress proliferation of non-Treg T cells, thus promoting a purer Treg yield 7,8,13,14 . RAP works by binding the 12-Kda FK506- and-Rapamycin-binding protein (FKBP12), forming a complex that exerts an inhibitory effect on the mammalian target of Rapamycin (mTOR) kinase, which is involved in the regulation of dendritic, B-cell, and T-cell activation 15,16 . While both Treg and Teff cells are affected by the suppressive properties of RAP, the impact is less profound on Tregs 15 . Still, the growth inhibition of Tregs treated with RAP remains an issue, and therefore alternatives for improving Treg in vitro expansions are sought 17 –19 . Apart from RAP, other macrolides such as Clarithromycin and especially Azithromycin (AZM) have also been recognized for their immunomodulatory effects 20 –22 . AZM is widely used for treatment of soft tissue infections and respiratory tract diseases, and has favorable safety and tolerability qualities 22 –29 . Interestingly, the drug shows immunomodulatory properties even in therapeutic dosages 22 –24,27 . AZM is also known for high intracellular accumulation in peripheral blood mononuclear cells (PBMCs), polymorphonuclear cells, and fibroblasts, with pleotropic effects on their function 21,22,30 –33 . Notably, the intracellular concentration of RAP in lymphocytes over time during expansion in vitro has not been analyzed before. A previous study by Ratzinger et al. suggests that AZM may, in similarity to RAP, suppress T-cell activation by modulation of the mTOR pathway, thus hampering CD4+ T-cell proliferation and cytokine secretion 20,34 . However, while the effects of AZM on bulk CD4+ T cells have been studied, the impact on Tregs has not been explored. The aim of the present study was to investigate if AZM is a possible alternative to RAP in enhancing the quality of in vitro expanded Tregs. We measured the effects of treating Treg cultures with AZM compared with RAP, in terms of expansive capacity, phenotype, and suppressive activity. In addition, we examined if the effects of AZM and RAP differ between expanded bulk, naïve CD45RA+, and memory CD45RA- Treg cultures. Furthermore, the degradation of RAP was examined by measuring intracellular and extracellular concentrations over time in vitro.
Materials and Methods
Blood Samples
Buffy coats were collected from healthy blood donors at the Uppsala University Hospital Blood Bank (Uppsala, Sweden) with approval from the Regional Ethics Committee (Dnr 2010/69). Female and male donors aged 18–65 were eligible for donation and a total of 27 donors were included in the study.
Cell Purification
PBMCs were isolated from buffy coats by separation over a Ficoll-Paque gradient (GE Healthcare, Chicago, Il, USA). Adherent cells were removed during 2 h incubation at 37°C in T175 flasks using standard RPMI-1640 medium modified with L-glutamine (Gibco, Thermo Fisher Scientific, Waltham, MA, USA), 1% 4-(2-hydroxyethyl)-1-piperazineethanesulfonic acid (HEPES), 1% penicillin-streptomycin, 0,04% β-mercaptoethanol and 2% pooled human serum. T cells were purified from the non-adherent PBMCs by magnetic activated cell sorting (MACS) using either the CD4+CD25+ Regulatory T Cell Isolation Kit or the CD4+ T Cell Isolation Kit (Miltenyi Biotec GmbH, Bergisch Gladbach, Germany) according to the manufacturer’s instructions. Isolated Tregs were used for bulk Treg cell culture, while isolated CD4+ T cells were used for further Treg purification by flow cytometric sorting or cryopreserved for later use with CryoStor® cell cryopreservation media (Merck, Darmstadt, Germany), frozen with CoolCell (Corning, NY, USA) and stored at –70°C for up to 3 weeks.
Flow Cytometric Sorting
MACS-sorted CD4+ T cells were thawed in 37°C water baths and washed with RPMI-1640 medium containing 10% fetal bovine serum. Cells were stained for flow cytometric sorting using the following antibodies: CD4-Brilliant Violet 421, CD25-Phycoerythrin, CD127-Fluorescein Isothiocyanate, and CD45RA-Phycoerythrin Cyanin 7, all from BD Biosciences, San Jose, CA, USA. Sorting was performed on FACS Aria III and FACS Melody from BD Biosciences. Different populations were selected for sorting: Effector T cells CD4+CD25lowCD127+, bulk Tregs CD4+CD25highCD127low or naïve Tregs CD4+CD25highCD127lowCD45RA+ and memory Tregs CD4+CD25highCD127lowCD45RA–. The purified Tregs were used for cell culture and the Teffs were cryopreserved for later use as described above.
Flow Cytometric Analyses
The expression of Treg-associated markers Forkhead box P3 (FoxP3), Helios, L-selectin (CD62L), and Interleukin 1 receptor type I (IL-1RI) was analyzed and compared between the expanded Treg cultures. In addition to the previously mentioned antibodies for flow cytometric sorting, the following antibodies were used for analysis: IL-1RI-FITC (R&D Systems, Minneapolis, MN, USA), CD45-Allophycocyanin-Hilite 7 (BioLegend, San Diego, CA, USA), CD62L-Brilliant Violet 421 (BioLegend), CD25-Brilliant Violet 510 (BioLegend), FoxP3-Alexa Fluor 647 (BD Biosciences), CD4-Peridinin-Chlorophyll-Protein Cyanin 5.5 (BD Biosciences) and Helios-Phycoerythrin (BD Biosciences). Cells were prepared for intracellular staining using FoxP3 Staining Buffer Set (eBioscience, Thermo Fisher Scientific) according to the manufacturer’s instructions. All flow cytometric analyses were performed on a FACSVerse™ (BD Biosciences).
Cell Cultures
During preparatory studies we found a large reduction of CD4+ T-cell numbers at AZM concentrations of 50 ug/ml or more, which is in concordance with what has been previously reported 20 . An intermediate AZM concentration of 25 ug/ml was therefore chosen to avoid cell toxicity and still retain a possible inhibition of the mTOR pathway. RAP was used at concentration of 100 ng/ml, which is commonly used in the literature and known to prevent Teff expansion 14 . Tregs purified by MACS or flow cytometric sorting were cultured in RPMI-1640 medium with the previously mentioned supplements and the addition of 10% human serum and IL-2 500 IU/ml. Starting populations consisted of 80,000–100,000 cells per well using 96-well plates. Human T-Activator CD3/CD28 beads (Gibco, Thermo Fisher Scientific) were added during culture start at a bead:cell ratio of 4:1 for Tregs and 1:1 for Teffs. Rapamycin 100 ng/ml (LC-laboratories, Woburn, MA, USA) or the AZM trade name Zithromax 25 ug/ml (Pfizer, New York, NY, USA) were added at culture start and continuously every second day. Beads were removed after a week and replaced at a 1:1 bead:cell ratio. Cultures were split to further wells and larger culture plates as the expansions grew, scaling up to 12-well and 6-well plates. Cultures were harvested after 2 weeks of growth and cells were either used for phenotype analysis by flow cytometry or rested for 48 h with IL-2 100 IU/ml without AZM or RAP before use in suppression assays.
Suppression Assays
FACS-sorted CD4+CD25lowCD127+ T cell responders (Tresps) were thawed in 37°C water baths and washed with RPMI-1640 medium containing 10% fetal bovine serum. Cells were stained with CellTrace™ Violet (Molecular Probes, Thermo Fisher Scientific) at a 5 μM concentration and incubated at 37°C for 20 min. Stained cells were washed using RPMI-1640 medium with 10% human serum. Tresps were cultured in 96-well plates at starting populations of 50,000 cells together with expanded resting Tregs at Treg:Tresp ratios of 1:1, 1:2, and 1:5. Human T-Activator CD3/CD28 beads were added during culture start at a bead:Tresp ratio of 0.25:1. Tresps co-cultured with expanded Teffs, or Tresps cultured alone, were used as positive controls. Tresps cultured without the addition of beads were used as negative controls. The suppression assays were harvested after 72 h, stained with LIVE/DEAD® Fixable Near-IR Dead Cell Stain (Molecular Probes, Thermo Fisher Scientific) and analyzed by flow cytometry. The frequencies of live non-proliferating Tresps were calculated using the proliferation tool in FlowJo version 10.6.0 (FlowJo LLC, Ashland, OR, USA).
Quantification of Rapamycin
MACS-sorted CD4+ T cells were thawed and washed as previously described. RPMI-1640 medium was used with addition of the previously mentioned supplements plus 10% human serum and IL-2 at a concentration of 100 IU/ml. The medium was stored in 12-well plates at 4°C or incubated at 37°C with or without the addition of 1 × 106 CD4+ T cells per well. RAP was added to all wells at the start of the experiment at a concentration of 100 ng/ml. Medium, cells, and supernatants were collected at multiple times during 48 h. Supernatants and cell pellets were separated by centrifugation and the cells were washed with 4°C phosphate buffer saline. Both supernatants and cells pellets were stored at –70°C until analysis. RAP in cells, medium, and supernatants was measured with liquid chromatography coupled to tandem mass spectrometry (LC-MS/MS) using a Transcend II LX-2 TSQ Quantiva system (Thermo Fisher Scientific). HPLC-grade methanol was used as extraction reagent for the cell analysis and methanol:zinc sulfate 0.1 mol/l (2:1) for analysis of the cell medium and supernatant. The extraction solutions contained the internal standard 13C, D3-rapamycin. Following mixing on a multishaker and centrifugation, the extracts were transferred to the instrument autosampler and injected on an Accucore C8 column (50 × 2.1 mm, 2.6 µm; Thermo Fisher Scientific). The mobile phases consisted of UHPLC-MS grade (A) water and (B) methanol added 0.10% formic acid and 2.0 mmol/l ammonium acetate. The chromatographic starting condition was 50% B, and elution of the analyte and internal standard was performed with 90% B (flow rate 0.600 ml/min, temperature 75°C). Electrospray ionization combined with selective reaction monitoring was applied for the mass spectrometric detection of positively charged ammonium adducts (RAP m/z 931.6 to 864.5 and the internal standard m/z 935.6 to 864.5). The peak area ratio between analyte and internal standard was used as instrument response, and the quantification was based on calibrators with declared RAP concentrations (Recipe, Munich, Germany).
Statistical Analysis
The Wilcoxon signed-rank test was used for comparing data between two populations. Groups of three populations were compared using the Friedman test with Dunn’s multiple comparisons post hoc test. A p-value <0.05 was considered statistically significant. FlowJo version 10.6.0 was used for analyzing flow cytometric data. Graphpad Prism version 6 (Graphpad Software Inc., San Diego, CA, USA) was used for statistical computations and visualizations.
Results
Phenotype of Expanded Tregs
Bulk, naïve, and memory Tregs were analyzed for CD4+FoxP3 along with the common Treg-associated markers Helios and CD62L after 2 weeks of culture. RAP treatment presented higher frequencies of CD4+FoxP3+Helios+ cells among memory Tregs compared with AZM treatment (p = 0.0373) (Fig. 1f). In addition, RAP showed higher frequencies of CD4+FoxP3+CD62L+ cells in naïve Tregs compared with AZM (p = 0.0231) (Fig. 1h). In comparison to no treatment, RAP treatment presented higher frequencies of CD4+FoxP3+ cells in all three populations: bulk Tregs (p = 0.0226), naïve Tregs (p = 0.0009), and memory Tregs (p = 0.0015) (Fig. 1a–c). RAP also showed higher frequencies of CD4+FoxP3+Helios+ cells compared with no supplement for both naïve Tregs (p = 0.0003) and memory Tregs (p = 0.0179) (Fig. 1e,f). AZM presented higher frequencies of CD4+FoxP3+ cells compared with no supplement only in bulk Tregs (p = 0.0099) (Fig. 1a). The CD4+FoxP3+ frequencies were also analyzed prior to expansion: bulk Tregs 91.6% (±6.31 SD), naïve Tregs 90.9% (±5.15 SD), and memory Tregs 86.3% (±3.82 SD).

Azithromycin and Rapamycin affect Treg phenotype. Graph showing the effect of Azithromycin and Rapamycin on the frequency of (
Expansion of Tregs
The expansion of Tregs was investigated after 2 weeks of culture. No significant differences in fold expansion were found between RAP and AZM treatment in either Treg sub-population. However, naïve Tregs treated with RAP presented lower fold expansion compared with no treatment (p = 0.0226) (Fig. 2b).

The influence of Azithromycin and Rapamycin on Treg expansion. The graphs summarize fold expansion of (
The Treg populations treated with AZM showed a trend of larger mean numbers of cells compared with the Treg populations treated with RAP: bulk Tregs 28.9%, naïve Tregs 29.4%, and memory Tregs 58.5% more cells than with RAP treatment (Fig. 2d). However, these differences in the mean number of cells did not reach statistical significance. The bulk Tregs treated with RAP showed a mean number of 2.55 (±2.61 SD) × 106 cells, compared with bulk Tregs treated with AZM which presented 3.27 (±1.91 SD) × 106 cells. A similar difference in mean cell numbers was found between naïve Tregs treated with RAP and AZM, which showed 8.19 (±4.27 SD) and 10.61 (±7.50 SD) × 106 cells, respectively. A larger difference was found between memory Tregs treated with RAP and AZM, which presented 4.07 (±3.99 SD) × 106 cells and 6.46 (±3.64 SD) × 106 cells, respectively.
FoxP3 Expression in Relation to Expansion
To investigate the Treg preserving effects of AZM and RAP during expansion, both fold expansion and the frequencies of CD4+FoxP3+ cells were compared after 2 weeks of culture. Tregs cultured with AZM presented larger variations than RAP in the frequencies of CD4+FoxP3+ cells, for which we found no significant correlation with fold expansion. RAP-treated Tregs presented higher frequencies of CD4+FoxP3+ cells than no treatment for all three populations, though no correlation was found between FoxP3 positivity and fold expansion (Fig. 3a–c). Neither did Tregs cultured with no supplement present any correlation between CD4+FoxP3+ cell frequencies and fold expansion (Fig. 3a–c). Though no significant differences were found between AZM and RAP, the Treg populations treated with AZM showed a trend of larger mean numbers of CD4+FoxP3+ cells compared with the Treg populations treated with RAP: bulk Tregs 16.2%, naïve Tregs 25.0%, and memory Tregs 17.6% more CD4+FoxP3+ cells than with RAP treatment (Fig. 2e). The bulk Tregs treated with RAP showed a mean number of 2.12 (±2.14 SD) × 106 CD4+FoxP3+ cells, compared with bulk Tregs treated AZM which presented 2.46 (±1.96 SD) × 106 CD4+FoxP3+ cells. Naïve Tregs treated with RAP and AZM presented slightly larger differences in mean numbers, showing 7.52 (±3.89 SD) and 9.40 (±6.81 SD) × 106 CD4+FoxP3+ cells, respectively. Memory Tregs treated with RAP and AZM showed differences in mean numbers comparable to that of bulk Tregs, presenting 3.3 (±3.06 SD) and 3.88 (±2.50 SD) × 106 CD4+FoxP3+ cells, respectively.

Treg fold expansion in relation to FoxP3 frequency. Graph showing the frequencies of CD4+FoxP3 positive cells in relation to fold expansion of (
Suppression Assay Using Cultured Tregs
Tregs expanded with AZM, RAP, or no supplements were tested for their suppressive activity in an antigen-presenting cell independent suppression assay. CellTrace™ Violet-stained CD4+ T cells (Tresps) were co-cultured with Tregs for 72 h at different Treg:Tresp ratios. The data are presented as the frequencies of live non-proliferating Tresps. The Treg cultures suppressed Tresps in a dose-dependent manner. RAP-treated naïve Tregs showed higher rates of suppression than AZM-treated Tregs for 1:2 Treg:Tresp ratios (p = 0.0424) (Fig. 4a). Also, RAP-treated naïve Tregs presented higher rates of suppression than Tregs with no treatment for both 1:1 (p = 0.0117) and 1:2 (p = 0.0183) Treg:Tresp ratios (Fig. 4a). No significant differences in the rates of suppression were found among memory Tregs (Fig. 4b).

Suppressive capacity of Tregs expanded with Azithromycin or Rapamycin. The graph summarizes the suppressive capacity of Tregs expanded with no treatment or in the presence of Azithromycin or Rapamycin. The rate of suppression is presented as the frequency of live non-proliferating autologous CD4+ T cell responders (Tresps) after co-culture with expanded (
CD4+FoxP3+IL-1RI Positivity in Expanded Tregs
As IL-1RI positivity in Tregs has been associated with potential conversion to a pro-inflammatory phenotype, the frequency of Tregs positive for CD4+FoxP3+IL-1RI was analyzed after 2 weeks of culture. No differences were found between treatment with RAP and AZM. Memory Tregs treated with RAP showed higher frequency of cells positive for CD4+FoxP3+IL-1RI compared with no treatment (p = 0.0081) (Fig. 5b), while no differences were found for naïve Tregs (Fig. 5a).
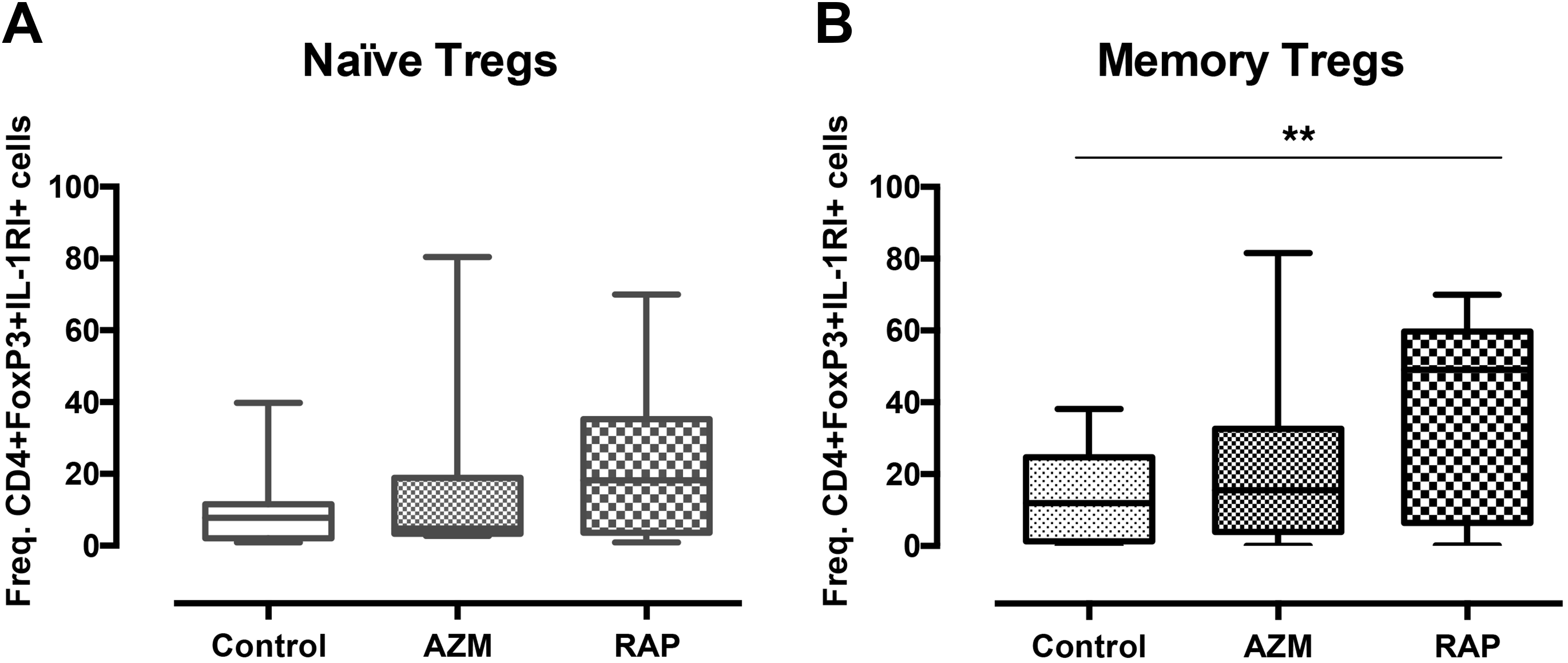
Rapamycin induces IL-1R1 expression. The graph present the frequencies of IL-1RI positive CD4+FoxP3+ cells among expanded (
Quantification of Rapamycin
To investigate the degradation of RAP in CD4+ T cells and in culture medium, both the intracellular and extracellular concentrations were measured over time. The medium, cells, and supernatants were harvested at multiple times during 24- or 48-h culture and the concentration of RAP was analyzed by LC-MS/MS. Measurement of RAP concentration in 37°C medium showed a pattern of quick degradation during the first 24 h regardless of whether it was cultured with cells or not, while the degradation in 4°C medium progressed more slowly (Fig. 6a). The CD4+ T cells presented a mean intracellular concentration of 383.2 (± 89 SD) ρg/106 cells 3 h after the addition of RAP, which dropped to 328 (± 63.6 SD) ρg/pellet in 24 h (Fig. 6b).

Rapamycin is retained in CD4+ T cells. Graph depicting the concentration of Rapamycin in RPMI medium, supernatant and cell pellet at multiple times during 48 h. Medium was stored at 4°C or incubated at 37°C with or without the addition of MACS-sorted CD4+ T cells. The concentration of Rapamycin was measured by LC-MS/MS. (
Flow Cytometric Scatter Analysis
To measure the potential effect of AZM and RAP on cell size and internal cellular complexity, forward scatter (FSC) and side scatter (SSC) was compared between cell cultures after 2 weeks of expansion. Significant differences in FSC and SSC were found between Tregs cultured with RAP and AZM for both naïve Tregs (p = 0.0004, p = 0.0004) and memory Tregs (p = 0.004, p = 0.004) (Fig. 7b).

Azithromycin increases Treg FSC and SSC compared with Rapamycin. Flow cytometric data showing forward-scatter (FSC) and side-scatter (SSC) properties of expanded Tregs. Cells enriched by FACS were stimulated with CD3/CD28 beads and cultured with no treatment or in the presence of Azithromycin or Rapamycin for 14 days. (
Discussion
The main objective of this study was to investigate the possible effects of AZM compared with RAP on in vitro expanded Tregs. Compounds that improve the quality of Treg in vitro expansions are sought as Treg cultures risk contamination of Teffs and conversion of Tregs to pro-inflammatory Th17 cells 7 –11 . Various agents such as RAP, transforming growth factor beta (TGF-beta), butyrate, and all-trans retinoic acid (ATRA) have presented Treg-enhancing capabilities in previous studies, though no consensus guidelines exist regarding optimal regimen for improving Treg in vitro expansions 7,8,14,35 –38 . While the mTOR inhibitor RAP is commonly used for improving the quality of Treg cultures, the growth inhibition of RAP-treated Tregs is an issue 17 –19 . AZM has known immunomodulatory properties and a previous study by Ratzinger et al. of AZM-treated CD4+ T cells suggests that AZM may, in similarity with RAP, suppress T-cell activation by modulation of the mTOR pathway 20 . Today AZM is widely used in the treatment of soft tissue infections and respiratory tract diseases, and has favorable safety and tolerability qualities that make the drug suitable for use in clinical applications 22 –29 .
In this study we analyzed the impact of AZM compared with RAP on Treg phenotype, expansion, and function. The effects were studied in bulk, naïve, and memory Tregs due to the known heterogeneity of Tregs subpopulations 39,40 . We found that RAP treatment induced higher frequencies of FoxP3+Helios+ cells among memory Tregs than AZM treatment. Also, RAP-treated naïve Tregs were more suppressive compared with AZM or no treatment. Previous studies have shown that FoxP3+Helios+ Tregs maintain a stable phenotype and feature more immune-suppressive characteristics compared with FoxP3+Helios– Tregs 41 –44 . This stability may be vital, as unstable Tregs can differentiate to pro-inflammatory Th17 cells during expansion, which could possibly be detrimental in clinical applications 9 –11 . For bulk Tregs, both RAP and AZM treatment induced higher frequencies of FoxP3+ cells than cultures with no treatment. This trend was also evident among naïve and memory Tregs, but was only statistically significant for RAP treatment. Thus, further studies are required to elucidate how and to which degree AZM treatment affects FoxP3 expression. Also, analysis of the Treg-specific demethylated region, which is considered a signature of stable Tregs when demethylated, would be interesting for future studies in terms of determining whether AZM treatment induces a stable or transient Treg phenotype 45 .
Regarding FoxP3+IL-1RI expression, we found higher frequencies of positive cells among the memory Treg cultures than the naïve Treg cultures, which is in concordance with previous studies on memory Tregs and IL1-RI expression 46,47 . In this study, RAP treatment, but not AZM treatment, further increased memory Treg IL-1RI frequency compared with no treatment. Interestingly, IL-1RI expression in Helios– Tregs has been associated with secretion of the suppressive cytokine IL-10, but also with secretion of the inflammatory cytokine IL-17, which is known to increases when Tregs are stimulated in the presence of IL-1 46,47 . The stability of the Treg lineage may be essential in clinical applications, and this notion suggests that memory Tregs and RAP treatment should be used with caution as IL-17 has been associated with multiple forms of inflammatory autoimmune pathologies 48 . According to our findings, culturing naïve Tregs may be preferred to minimize the frequencies of potentially inflammatory IL1-RI+ cells in Treg expansions. In addition, we found that naïve Treg cultures presented high frequencies of classic FoxP3+Helios+CD62L+ Tregs even with no treatment. Naïve Tregs may therefore be suitable for stable expansion of Tregs, which has been previously suggested in the literature 40,49 . However, naïve Tregs constitute only a minority of the Treg population in adults, and have been found to decrease with age while CD45RO memory Tregs increase 50 . Thus, the relatively small proportion of naïve Tregs may hinder amassment of large cell numbers for clinical applications, especially when dealing with elderly patients with lower numbers of naïve cells. We found that RAP-treated naïve Tregs expanded less than Tregs with no treatment, which is line with the literature 17 –19 . Interestingly, our study did not show any differences in fold expansion for AZM-treated Tregs, compared with studies on CD4+ T cells by Ratzinger et al. and Lin et al., which present a dose-dependent negative effect of AZM treatment on expansion, viability and cytokine secretion 20,34 . While we did not analyze for viability or cytokine secretion in this study, our findings showed that the mean number of cells after expansion with AZM was 28–58% more compared with expansion with RAP. Although this trend did not reach statistical significance in our study, it may be of importance as Treg therapies are dose-dependent and require ex vivo expansion of Tregs to accumulate therapeutic levels 6 . Notably, even though RAP treatment may retain the purity of Treg expansions better than AZM treatment, the mean number of FoxP3+ cells acquired in our study was still 16–25% higher in Treg expansions treated with AZM compared with RAP. While the mechanisms of AZM that inhibits CD4+ T cells has not been elucidated, the study by Ratzinger et al. suggested that AZM may in similarity with RAP interfere with the mTOR pathway, though in an FKBP12-independent manner. As Tregs are more resistant to the suppressive effects of RAP compared with conventional CD4+ T cells, this resistance could potentially apply for AZM treatment of Tregs as well. Further research exploring how AZM affects Treg viability, expansion, phenotype, and cytokine secretion would be of interest, preferably in comparison with conventional CD4 T cells. Also, studies of AZM dosing and its potential effects on Tregs are needed. Notably, we found that memory Treg expansions treated with RAP or AZM from two different blood donors did not expand while their respective naïve Treg cultures thrived, which may indicate a lower resistance among memory Tregs for these compounds. Our analysis of RAP concentration in CD4+ T cells and cell medium showed stable intracellular levels of RAP and rapid degradation in 37°C cell medium within 24 h. Similarly, a previous study of AZM presented an intracellular accumulation of AZM in leukocytes that slowly degraded over days, while the concentration in plasma quickly dropped within hours 32 . However, considerable individual variations in AZM concentration were found in the study. Another pharmacological study of AZM concentration in PBMCs and polymorphonuclear leukocytes also showed high uptake of AZM with large individual variations, as well as inconsistent fluctuations of AZM concentration over time 33 . Notably, we found larger individual variations in the frequencies of FoxP3+ cells between donors treated with AZM compared with RAP treatment. Further research is required to elucidate the mechanisms behind intracellular accumulation of AZM and whether individual variations in uptake may affect Tregs differently.
In the flow cytometric analysis, Treg cultures treated with AZM showed significantly increased FSC and SSC in comparison with the cultures treated with RAP for both naïve and memory Tregs. These findings indicate that Tregs treated with AZM are both larger and show increased internal cellular complexity in comparison with Tregs treated with RAP. The internal complexity of AZM Tregs can possibly be explained by the known intracellular accumulation of AZM in lysosomes, which are highly efficient structures for scattering light 33,51,52 .
In conclusion, we found that RAP treatment induced a FoxP3+Helios+ phenotype and increased suppressive function, but may also inhibit Treg expansion. In comparison, AZM treatment promoted a FoxP3+ phenotype, but to a lesser extent than RAP and the AZM-treated Tregs are possibly less suppressive. While AZM treatment showed a trend of marginally more Treg expansion than RAP treatment, no significant differences were found in this regard. These findings imply that further elucidation of the biological effects and functional impact of AZM is required to determine if the compound may benefit Treg in vitro cultures for clinical applications.
Supplemental Material
Supplemental_material - Comparing the Effects of the mTOR Inhibitors Azithromycin and Rapamycin on In Vitro Expanded Regulatory T Cells
Supplemental_material for Comparing the Effects of the mTOR Inhibitors Azithromycin and Rapamycin on In Vitro Expanded Regulatory T Cells by Marcus Bergström, Malin Müller, Marie Karlsson, Hanne Scholz, Nils Tore Vethe and Olle Korsgren in Cell Transplantation
Footnotes
Ethical Approval
This study was approved by the Regional Ethics Committee (Dnr 2010/69), Uppsala, Sweden.
Statement of Human and Animal Rights
This article does not contain any studies with human or animal subjects.
Statement of Informed Consent
There are no human subjects in this article and informed consent is not applicable.
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) received no financial support for the research, authorship, and/or publication of this article.
Supplemental Material
Supplemental material for this article is available online.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.