
Abstract
There is growing clinical interest in the utilization of mesenchymal stem cells (MSCs) in the management of acute graft-versus-host disease (aGvHD), yet the effect of major histocompatibility complexes (MHCs) on B lymphocytes in this process has been less well documented. Working in an MHC fully mismatched murine aGvHD model, we found that MSC co-transfer significantly prolonged the survival time of the recipients. More interestingly, analysis on immunophenotypic profiles of posttransplant splenocytes showed that surface expression of CD69 (an early activation marker) and CD86 (a costimulatory molecule) was suppressed predominantly on donor derived B lymphocytes by MSC infusion. Additionally, mRNA level of interleukin-4, a potent B lymphocyte stimulator, was strikingly reduced from MSC-treated mice, while interleukin-10, the regulatory B lymphocytes inductor, was increased; these may underlie the lesser activation of B lymphocytes. In consistence, depletion of B lymphocytes in the transfusion inoculum further prolonged the survival time of aGvHD mice regardless of MSC administration. Therefore, B lymphocytes played an important role in the development of aGvHD, and they are targets in MSC-regulated immune response cascade in vivo. This study may provide a mechanistic clue for the treatment of human clinical aGvHD.
Introduction
Acute graft-versus-host disease (aGvHD) continues to be a leading cause of morbidity and mortality after allogeneic hematopoietic stem cell transplantation (HSCT), which significantly limits the successful outcome of HSCT in the treatment for a number of hematologic malignancies. aGvHD usually manifests within 100 days following HSCT. Clinically significant aGvHD (grade II or higher) developed in 20–65% of patients. Mortality due to this complication accounts for approximately 50% 1,2 .
T lymphocytes are the main actor in aGvHD development. aGvHD is mediated by the cytotoxic T lymphocytes attacking 3 –5 . The pathogenesis of aGvHD can be divided into three sequential steps 6,7 . The myelosuppressive conditioning regimen causes tissue damage that leads to a pro-inflammatory environment and antigen-presenting cell activation. Donor T cells activation and proliferation are induced by the pro-inflammatory milieu. The activated alloreactive T cells induce apoptosis of target cells and secrete pro-inflammatory cytokines, enhancing aGvHD.
Mesenchymal stem cells (MSCs) are a progenitor cell population with multilineage potency. MSCs were initially discovered in bone marrow and were subsequently found in almost every type of tissue, such as adipose tissue, dental pulp, amnion, and umbilical cord 8,9 . A large number of studies have demonstrated that MSCs are low allogeneic stimulators and may even suppress an ongoing immune process, in which T lymphocytes 10 –13 and dendritic cells 14,15 are involved. In light of their immunoregulatory properties and ease of in vitro expansion, MSCs have been utilized clinically to treat patients with aGvHD, especially in the treatment of steroid-refractory aGvHD 16 . Clinical data suggest the complete response to MSC treatment of patients to be about two-thirds 17 –19 . The incidence of cytomegalovirus, Epstein–Barr virus infections, and tumor relapse was not different between the non-MSCs group and the MSCs group during aGvHD treatment and follow-up 20 .
B lymphocytes have been considered as effector cells in chronic GvHD 21,22 . MSCs have been reported to regulate B lymphocyte activation in vitro and in vivo 23,24 . Previous studies about the immunoregulatory effects of MSCs on B lymphocytes mainly related to the B lymphocyte maturation and immunoglobin section functions 25 . Rosado et al. reported that inhibition of B-cell proliferation and antibody production by MSCs was mediated by T cells 26 .
B lymphocytes can also be competent antigen-presenting cells for delivery of protein antigens to CD4+ T cells in vivo 27 . Up to now, the basic question of how MSCs act on B lymphocytes in aGvHD remains rarely documented. Our previous study revealed that the functional and phenotypic alteration of T lymphocytes was affected by MSCs in the aGvHD model 28 . In the present study, we examined the immunoregulatory effect of MSCs on B lymphocytes. The results revealed that donor B lymphocytes played an important role in the development of aGvHD, MSCs potently inhibited the expression of CD69 and CD86 on B lymphocytes, and B lymphocyte deletion further prolonged the mean survival time (MST) of aGvHD mice.
Materials and Methods
Cell Culture
Primary MSCs were isolated from C57BL/6 (B6) murine compact bone fragments, and culture-expanded as described in our previous report 29 . Cells were grown in minimal essential medium (GIBCO, Grand Island, NY, USA) with 4 mM L-glutamine, 100 U/ml penicillin, 100 U/ml streptomycin, and 10% fetal bovine serum in a humidified atmosphere of 5% CO2 at 37°C.
Murine aGvHD Model
aGvHD was induced as previously described with some modifications 30 . Briefly, inoculum of 7×107 unfractionated B6 (H2b) nucleated spleen cells, with or without MSCs (1×106), was intravenously injected into sublethally irradiated (60Co, 5 Gy) female recipient BALB/c (H2d) aged 6–8 weeks, which were designated as aGvHD+MSCs or aGvHD group respectively. Syngeneic controls received BALB/c splenocytes (7×107). In some experiments, splenocytes (3.85×107) immuno-magnetically depleted of B220+ cells (B lymphocytes at all stages) were used with or without co-transfer of MSCs (B220–/aGvHD+MSCs or B220–/aGvHD group).
Alloreactive Cytotoxic Assay
Aliquots (2×106) of nucleated splenocytes of aGvHD and aGvHD+MSCs harvested on day 5 were cultured in triplicate in 96-well plates with 3H-TdR-labeled P815 cells (H2d) for 5 h, at an effector/target ratio of 40:1. The percent specific cytolysis was determined as follows: specific lysis (%) = (cpm maximum – cpm experimental)/cpm maximum.
Flow Cytometry Analysis
At different posttransplant time points, nucleated splenocytes from two or three mice were pooled and surface stained with fluorescein isothiocyanate or phycoerythrin-conjugated monoclonal antibodies. Antibodies against mouse CD3 (145-2C11), H2b (28-8-6) were purchased from BD-Pharmingen (BD Biosciences, San Jose, CA, USA). Antibodies against mouse CD11b (M1/70), mIgM (RMM-1), CD69 (H1.2F3), CD86 (GL-1), CD25 (3C7) and CD4 (GK1.5) were from BioLegend (San Diego, CA, USA). Cells were examined on a FACSCalibur with CellQuest software (BD Biosciences). Data were analyzed using FlowJo 7 software (Joseph Trotter, La Jolla, CA, USA). Acquired events were gated for the designated population.
Real-time Polymerase Chain Reaction
At different posttransplant time points, B220 positive splenocytes from aGvHD were collected by magnetic bead sorting (R&D Systems, Minneapolis, MN, USA) and total RNA was extracted using Trizol (Invitrogen, Carlsbad, CA, USA). Quantitative real-time polymerase chain reaction (qRT-PCR) was performed as described by the manufacture’s instructions (TOYOBO, Osaka, Japan). The primers are: interleukin (IL)-4: upper, 5′-CCATATCCACGGATGCGACA-3′, lower, 5′-CTGTGGTGTTCTTCGTTGCTG-3′, IL-10: upper, 5′-GGCCCAGAAATCAAGGAGCA-3′, lower, 5′-ACAGGGGAGAAATCGATGACAG-3′, HPRT: upper, 5′-AGCCTAAGATGAGCGCAAGT-3′, lower, 5′-GGCCACAGGACTAGAACACC-3′. The levels of gene expression were calculated by relative quantification using HPRT as the endogenous reference genes. The gene expression level at the time point of 12 h post transplantation was designated as 1.
Statistical Analysis
Statistical analysis was performed using Prism 6 (GraphPad Software, San Diego, CA, USA). The Kaplan–Meier product-limit method was used to calculate survival curve. Differences between groups in survival studies were determined using log-rank statistics. Statistical analysis was performed with the log-rank test or the Student t test. p value less than 0.05 was considered statistically significant.
Results
MSCs Infusion Prolonged the MST of aGvHD Mice
In agreement with the clinical trials 18 –20,31 , alleviation of aGvHD severity by co-transfer of expanded MSCs was also evident in this study, in that the MST of MSC co-infusion in mice was significantly prolonged (7.89±0.73 days for aGvHD+MSCs group vs. 5.31±0.45 days for aGvHD group, n = 15, p < 0.05). Correspondingly, by day 5, the anti-H2d (P815 cells) cytotoxicity of posttransfusion splenocytes was significantly reduced (Fig. 1A), which suggests that MSC treatment suppressed anti-host (H2d) reactivity of donor derived T lymphocytes (H2b).
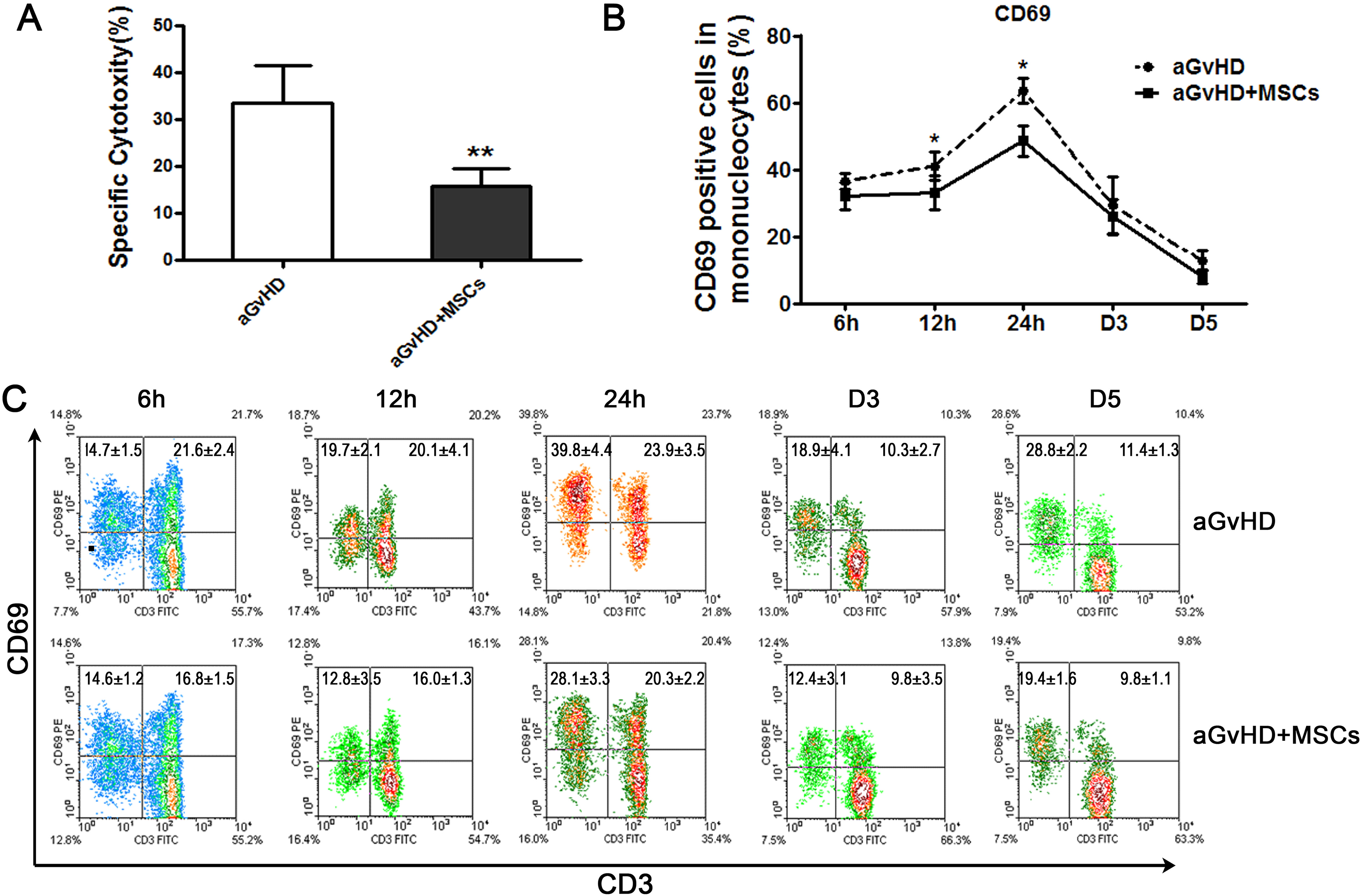
Mesenchymal stem cell (MSC) infusion decreased the CD69 expression on the splenic mononucleocytes of acute graft-versus-host disease (aGvHD) mice. A. Co-infusion of MSCs significantly suppressed specific killing capacity of donor splenocytes (H2b) against P815 cells (H2d). Splenocytes from at least three mice five days after transplantation were used in each group and the results are pooled from three separate experiments; n = 15, **p < 0.01. B. The splenocytes were collected and examined using flow cytometry (FCM) technique at different time points post transplantation. The mononucleocytes were gated for analysis. The linear graph showed that CD69 expression on the splenic mononucleocytes was suppressed by MSC infusion; n = 12, *p < 0.05. C. The FCM data exhibited the CD69 expression status on the CD3 positive T lymphocytes and CD3 negative cells; n = 12.
MSC Infusion Significantly Down-regulated the Expression of CD69, an Early Activation Marker, on B Lymphocytes
CD69 is known as a very early activation marker; it is upregulated on T cells during the first kinetics phase of brief contacts between T cells and antigen presenting cells 32 . We, then, attempted to define cellular events involved in MSC functionality by characterizing the phenotypes of splenocytes harvested 12 h, 24 h, 48 h, day 3, and day 5 post-transplantation, focusing on surface activation marker expression. Flow cytometry analysis showed that CD69 expression exhibited an inverse “V” curve, peaking at 24 h, and MSC co-transfer down-regulated CD69 expression obviously at 12 h and 24 h. The percentage dropped from 41.0±4.2% to 33±5.0%, and from 63.5±3.8% to 48.5±4.7% for aGvHD and aGvHD+MSCs groups respectively (Fig. 1B, C).
Nevertheless, percent CD69 reduction in CD3+ cells was not obvious (1.2–5.0% of all spleen mononucleated cells), in apparent contrast to that in the CD3- population (Figs 1C, 2A). Then, its expression on mature B lymphocytes (membrane IgM, mIgM+), monocytes (CD11b+), and natural killer (NK) cells (NK1.1+) between the two groups was compared separately. Interestingly, decrease of percent CD69 in mature B lymphocytes was predominantly evident, as the proportion was 72.0% (63.4–85.3%) of total CD69 decrease in CD3- cells at 12 h (Fig. 2B) and 58.8% (49.6–72.3%) at 24 h. A decline of percent CD69 was also observed in CD11b+ cells, but it contributed less to the total decrease (Fig. 2C). The proportion of NK cells was extremely low in the aGvHD recipient mice. Therefore, they had few contributions to the CD69 expression (Fig. 3).

The inhibited CD69 expression was mainly on the B lymphocytes in spleens in the acute graft-versus-host disease (aGvHD) model. A. CD69 expression status on the CD3 negative cells using flow cytometry (FCM) technique; n = 12, *p < 0.05. B, C. Splenocytes were collected post-infusion, and CD69 expression on different immune subsets was analyzed. The representative FCM density diagram showed that activation (CD69 expression) of mature B lymphocytes (membrane IgM+) was chiefly suppressed (B) when compared with monocyte-macrophages (CD11b+) (C); n = 12.
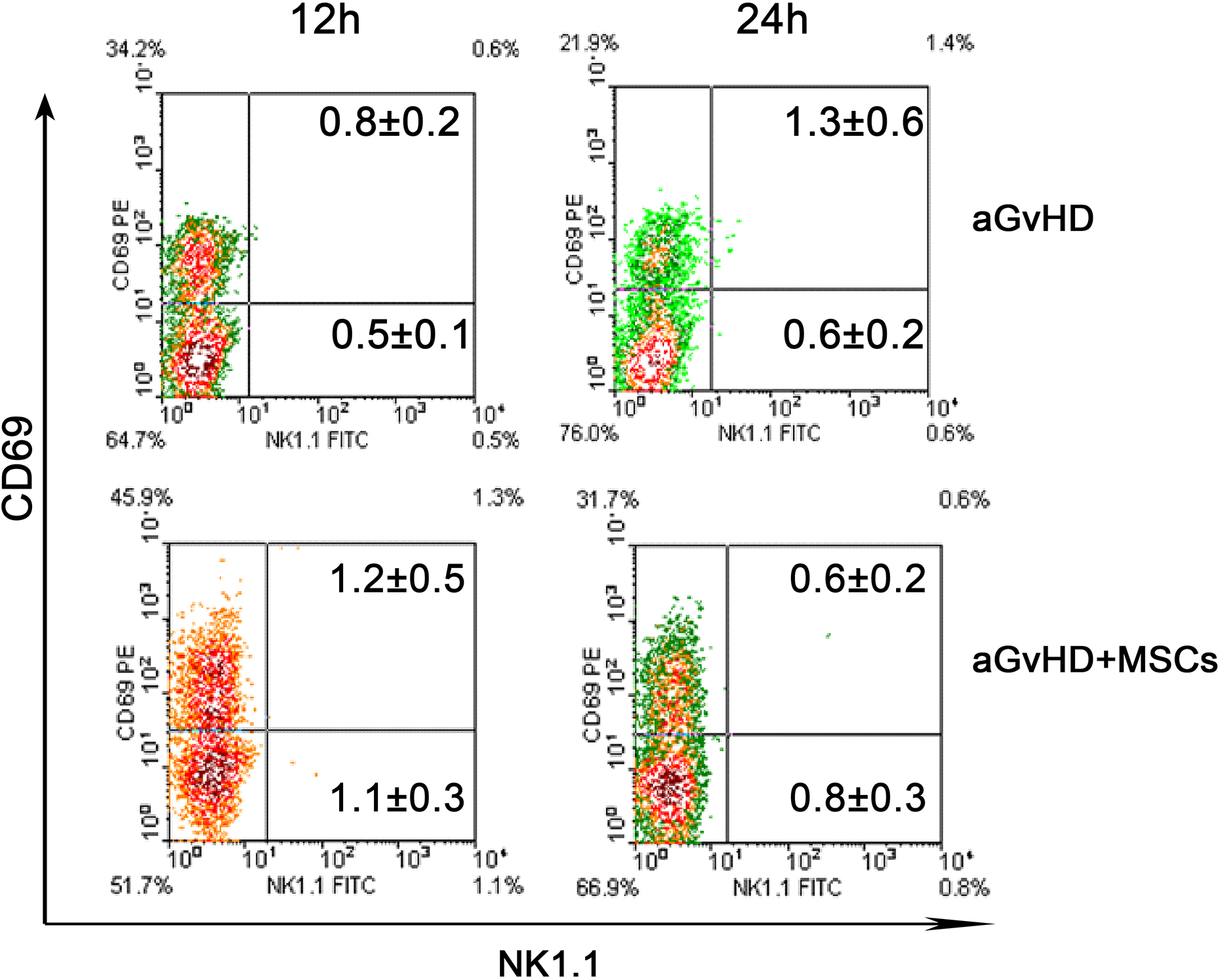
Natural killer (NK) cells were rarely affected in the acute graft-versus-host disease (aGvHD) model. The representative flow cytometry density diagram showed that the proportion of NK cells was relatively very low in the splenocytes and there was no significant difference between aGvHD and aGvHD+MSCs groups of mice at 12 h and 24 h post transplantation; n = 12.
The Co-stimulatory Molecule CD86 Decreased by MSC Infusion
B lymphocytes are a kind of antigen-presenting cell, but the contribution is controversial as they can either prime naïve T cells or induce T cell anergy, depending partially on surface co-stimulatory molecule expression 33,34 . In this study, we found that over 75% of IgM+ cells were positive for CD86 molecule at 12 h and 24 h (Fig. 4A, C). Similar to CD69 expression, MSC co-transfer also down-regulated CD86 expression on B lymphocytes at all observing time points after transplantation (6 h,12 h, 24 h, day 3, day 5), implying that MSCs exert immunoregulatory effects by down-regulating alloantigen presenting function of B to T lymphocytes.

Mesenchymal stem cell (MSC) infusion decreased the CD86 expression on B lymphocytes, which were predominantly donor derived. A. CD86 expression on B lymphocytes indicated by the ratio of CD86 and IgM double positive cells to IgM positive cells at different time point post infusion detected by flow cytometry (FCM) technique; n = 12, *p < 0.05, **p < 0.01. B. The proportion of donor derived B lymphocytes, which was displayed by the ratio of co-expressed H2b+ mIgM+ to mIgM+ B lymphocytes detected by FCM technique; n = 12. C, D. The representative flow cytometric density diagram of CD86 co-expression (C) and co-expressed H2b+ (D) on IgM+ B lymphocytes; n = 12. aGvHD: acute graft-versus-host disease

B lymphocyte deletion further prolonged the mean survival time (MST) of acute graft-versus-host disease (aGvHD) mice. A. The survival curve of the four groups of mice. The MST was prolonged significantly when compared between aGvHD (n = 14) and B220–/aGvHD (n = 14), and aGvHD+MSCs (n = 6) and B220–/aGvHD+MSCs (n = 6), *p < 0.01. B. Real-time polymerase chain reaction analysis showed that mesenchymal stem cells (MSCs) affected cytokine profiles in B220+ cells. C. The flow cytometry (FCM) result showed that the B lymphocytes were mostly deleted by B220 antibody, and the CD69 expression was restrained simultaneously; n = 3. D. The FCM result of syno-infusion group of mice at 24 h post infusion; n = 3.
It was also noteworthy that most IgM+ cells co-expressed H2b at 12 h and afterwards post transplantation (Fig. 4D), indicating the donor origin of the activated B lymphocytes. Therefore, our results linked B lymphocytes to MSC-regulated immune response cascade in vivo.
B Lymphocyte Deletion Further Prolongs the MST of Mice of aGvHD and aGvHD+MSCs Groups
To confirm the functional role of B lymphocytes in this experimental model, we depleted B220+ cells (B lymphocytes at all stages) from the transfusion inoculum. The results showed that the MST was prolonged significantly when compared with the aGvHD group with B220–/aGvHD group (5.31±0.45 days vs.10.00±2.10 days, p = 0.0167), the aGvHD+MSCs with B220–/aGvHD+MSCs (7.89±0.73 days vs.12.53±2.71 days, p = 0.0256) (Fig. 5A), indicating that B lymphocyte activation was a critical step in the development of aGvHD. Nevertheless, there were no differences between the B220–/aGvHD and B220–/aGvHD+MSCs group of mice, which indicates that B lymphocytes are the target of MSC regulation in vivo.
We further analyzed the cytokine profiles of splenic B220+ cells of aGvHD and aGvHD+MSCs by qRT-PCR. As indicated in Fig. 5B, the mRNA level of IL-4, a well-characterized potent stimulator for B lymphocytes, was remarkably down-regulated at every time point, implying that suppression of B lymphocyte activation by MSCs might be at least partially attributed to decreased IL-4 autocrine. Also, MSCs up-regulated IL-10 expression, but these changes occurred at late stages, when B lymphocyte activation declined. This change might be related to the formation of B regulatory cells 35 .
In the B220–/aGvHD and B220–/aGvHD+MSCs groups of mice the membrane IgM positive cells were almost deleted by the B220 antibody at the 24 h time point after infusion, whatever host (H2b–) or donor (H2b+) derived (Fig. 5C), therefore, the CD69 and CD86 expression was mainly concentrated in the IgM negative cells. CD69 expression on the CD3 positive T lymphocytes was significantly lower than those of the aGvHD and aGvHD+MSCs mice (Fig. 5C), which may explain the prolonged MST by B220 deletion. In this aGvHD model, the syno-splenic cell infusion mice acted as the control group, the mice were alive long-term (Fig. 5A), and very few T and B lymphocyte were activated (Fig. 5D).
Discussion
MSCs play a key role in the regulation of the immune responses against allo- and auto-antigens, and have been employed in clinical trials to treat steroid-resistant aGvHD. Our study revealed that B lymphocytes are targets of MSCs’ immunoregulatory effect and play an important role in the development of aGvHD.
aGvHD is a life-threatening complication of allogeneic HSCT. Previous efforts emphasized the crucial role of T cells in the development of aGvHD 28 . CD69 is a membrane-bound, type II C-lectin receptor. It is known that CD69 is a classical early marker of T lymphocyte activation during the first kinetics phase of brief contacts between T cells and antigen presenting cells, as it rapidly appears on the surface of the plasma membrane after stimulation 36 . CD69 knockout mice enhanced susceptibility to different inflammatory diseases 37 . Cross-linking of CD69 by monoclonal antibody induced a prolonged elevation of intracellular [Ca2+], which was induced by the influx of extracellular Ca2+. This signal resulted in ERK1/2 kinases activation, induction of synthesis of IL-2 and interferon-γ, and cell proliferation. We examined the CD69 expression status at 6 h, 12 h, 24 h, day 3, and day 5 post transplantation. Our study demonstrated that MSC co-infusion surely down-regulated CD69 expression on the mononucleated splenic cells of aGvHD recipient mice. However, CD69 up-regulation was not so obvious on the CD3+ T lymphocytes (Fig. 1C). CD69 is also promptly up-regulated on cells of most hematopoietic lineages, including B lymphocytes, macrophages, NK cells, etc 36,38,39 . Thus, the expression kinetics of CD69 on antigen presenting cell types is one of costimulatory molecules 40 . Then we also examined CD69 expression status on the other cell subtypes. Our results indicated that the proportions of CD11b positive cells (monocytes, macrophages) and NK1.1 positive NK cells were very low. CD69 was significantly down-regulated on the B lymphocytes (Fig. 2B), therefore, this present study focused on what role of B lymphocytes play in the aGvHD and their relationship with infused MSCs.
Previous investigations revealed that MSCs can regulate B lymphocyte function. But the regulatory direction is mainly dependent on the treatment microenvironment. It was reported that treatment with human MSCs resulted in an increase of proliferation, differentiation of B lymphocytes into plasma cells, and production of antibodies in vitro. But it is also reported that mouse MSCs significantly enhanced T cell dependent and independent antibodies production in vivo in mice 41 . Human palatine tonsil-derived MSCs ameliorate B-cell-mediated immune responses and increase IL-10-expressing regulatory B lymphocytes. On the contrary, Luk et al. found that in the inflammatory conditions treated MSCs potently reduced B lymphocyte proliferation and IgG production but did not induce regulatory B cells or IL-10 production 42 . Conditioned medium from cultivation of MSCs alone has no effect on B-cell expansion. MSCs need to be activated to exert their suppressive properties. Human MSCs and B lymphocytes were cocultured with different B-cell tropic stimuli. B-cell proliferation, maturation and production of IgM, IgG, and IgA was significantly impaired 43,44 . Transwell experiments indicated soluble factors were the major mechanism of B-cell suppression. The Corcione group reported that human MSCs also significantly down-regulated B-cell chemotaxis property to CXCL12, the CXCR4 ligand, CXCL13, and the CXCR5 ligand 45 .
An important function of B lymphocytes is antigen presenting. Corcione et al. reported that hMSCs did not affect the expression of B-cell costimulatory molecule, MHC-II, CD40, CD86, and CD80, and cytokine production, which indicated that the antigen presenting properties of B lymphocytes are not affected 45 . However, the immunoregulatory function exerting is dependent on the inflammatory circumstance exposure. Our present data demonstrated that MSCs potently inhibited the costimulatory molecule CD86 expression on B lymphocytes. These processes were more obvious at the early stage of aGvHD. To further investigate the B cells’ function in the development of aGvHD, B lymphocytes were deleted by B220 antibody. Data in Fig. 5 show that B lymphocyte deletion prolonged the MST of aGvHD mice. The MST was 5.31±0.45 days for aGvHD group, 10.00±2.10 days for B220–/aGvHD group, 12.53±2.71 days for B220–/aGvHD+MSCs group. There was no difference between the B220–/aGvHD and B220–/aGvHD+MSCs group of mice. The underlying reason may be due to the critical role of B lymphocyte in aGvHD development, and B lymphocytes are a target of MSC regulation in vivo.
Conclusion
To the best of our knowledge, this study is the first to represent the evidence that MSCs target B lymphocyte function to inhibit the development of aGvHD. The cell activation and antigen presenting function of B lymphocytes was restrained by MSC infusion. This present study might shed light on therapeutic targets for the treatment of human clinical aGvHD.
Footnotes
Acknowledgment
The authors thank ZiKuan Guo for the kind help in manuscript writing.
Ethical Approval
This study was carried out in strict accordance with the recommendations in the national guidelines for the use of animals in scientific research “Regulations for the Administration of Affairs Concerning Experimental Animals.” Additional approval was granted by the Animal Care and Use Committee of Chinese PLA General Hospital, with the approval number PGH-16021103.
Statement of Human and Animal Rights
All experimental procedures were approved by the animal care committee of the Chinese PLA General Hospital.
Statement of Informed Consent
There are no human subjects in this article and informed consent is not applicable.
Declaration of Conflicting Interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: this work was supported by National Natural Science Foundation of China (No. 81571619), the National Key Research and Development Program of China (Nos. 2016YFE0204400).