
Abstract
Airway epithelial cell injury is a key triggering event to activate allergic airway inflammation, such as asthma. We previously reported that administration of mesenchymal stem cells (MSCs) significantly alleviated allergic inflammation in a mouse model of asthma, and the mmu-miR-21/ACVR2A axis may be involved. However, whether MSCs protect against bronchial epithelial cell injury induced by hypoxia, and the underlying mechanism, remain unknown. In our study, the human bronchial epithelial cell line BEAS-2B was induced to undergo apoptosis with a hypoxia mimic of cobalt chloride (CoCl2) damage. Treatment of MSCs derived from induced pluripotent stem cells (iPSCs) significantly decreased apoptosis of BEAS-2B cells. There was high miR-21 expression in injured BEAS-2B cells after MSC treatment. Transfection of the miR-21 mimic significantly decreased apoptosis of BEAS-2B, and transfection of a miR-21 inhibitor significantly increased apoptosis. More importantly, the protective effects of MSCs on injured BEAS-2B were reversed by transfection of the miR-21 inhibitor. Binding sites of human miR-21 were identified in the 3’UTR of human ACVR2A. We further determined that CoCl2 stimulation increased ACVR2A expression at both the mRNA and protein levels. Moreover, transfection of the miR-21 mimic further up-regulated ACVR2A expression induced by CoCl2, whereas transfection of the miR-21 inhibitor down-regulated ACVR2A expression. In addition, MSCs increased ACVR2A expression in BEAS-2B cells; however, this effect was reversed after transfection of the miR-21 inhibitor. Our data suggested that MSCs protect bronchial epithelial cells from hypoxic injury via miR-21, which may represent an important target. These findings suggest the potentially wide application of MSCs for epithelial cell injury during hypoxia.
Introduction
The airway epithelium is considered an essential controller of inflammatory, immune and regenerative responses to allergens, viruses and environmental pollutants, which contribute to asthma pathogenesis 1,2 . Epithelial cells express pattern recognition receptors that detect environmental stimuli and secrete endogenous danger signals, which thereby activate dendritic cells and bridge innate and adaptive immunity. Epithelial inflammation dominates the pathophysiology in several lung diseases, such as asthma and chronic obstructive pulmonary disease (COPD) 3 . Asthma is a chronic inflammatory disease characterized by inflammation of the airways and frequently accompanies hypoxic conditions 1 . In asthma, the respiratory epithelium is damaged and produces more mucus, structurally becomes more permeable and more sensitive to oxidants and exhibits a deficiency in the innate immune response to respiratory virus infections 4 . Accordingly, research efforts are focused on understanding the cell-specific signaling responsible for these inflammatory disease states and developing more targeted therapies to circumvent them.
Mesenchymal stem cells (MSCs) are multipotent cells that are capable of differentiation into numerous mesenchymal lineages, such as osteoblasts, chondrocytes, myocytes, adipocytes 5,6 and endothelial cells 7 . Increasing data in animal studies and preliminary clinical trials have indicated that MSCs are considered a promising resource for cell therapy, including a series of immunological diseases, such as graft-versus-host disease (GvHD) 8 , arthritis 9,10 , Crohn’s disease 11 and systemic lupus eryematosus 12,13 . Herrera’s group reported that transplanted MSCs enhanced tubular proliferation, differentiated into tubular epithelial cells, and regenerated tubular epithelial cells 14 . We have previously reported the suppressive effects of MSCs, including bone marrow-derived MSCs (BM-MSCs) or induced pluripotent stem cell-derived MSCs (iPSC-MSCs), to allergic rhinitis in humans 15 and asthma in mice 16 . The treatment of MSCs in asthmatic mice mitigated Th2 inflammation, as well as pulmonary injury, and the mmu-miR-21/ACVR2A axis played an important role 17 . However, whether MSCs protect airway epithelial cells and the relative mechanisms are unclear.
MicroRNAs (miRNAs), which are 19–22-nucleotides long, are non-coding RNAs that are in all eukaryotic cells and mediate the post-transcriptional silencing of target genes 18 . In animals, miRNAs tend to regulate target gene expression via translational inhibition or mRNA degradation through binding to complementary sites in the 3’ untranslated region (UTR) of target genes 19 . miRNAs are involved in many pathological processes, including inflammation, the stress response, solid tumors, and cardiac abnormities. Numerous reports have shown that miRNAs are involved in the function of airway epithelial cells. A total of 28 miRNAs were differentially expressed in the bronchial airway epithelium of smokers compared with healthy controls 20 . MiR-126 was down-regulated in cystic fibrosis bronchial epithelial cells and was correlated with a significant up-regulation of TOM1 mRNA 21 . Substantial changes in airway epithelial cell miRNA levels are a common feature of asthma 22 . MiR-21 is an abundantly expressed member of the small non-coding miRNA family in multiple mammalian cell types 23 . MiR-21 was highly expressed in asthmatic children 24,25 and mice 17 , and is considered one biomarker of asthma. We have previously reported that mmu-miR-21a-3p was significantly up-regulated in the lung in asthmatic mice and recovered to normal levels after MSC treatment 17 . However, whether miR-21 mediates the protective effects of MSCs for bronchial epithelial cells and how miR-21 functions remain unclear.
In our study, we investigated the effects of MSCs on injured respiratory epithelial cells induced by cobalt chloride (CoCl2) and the potential mechanisms of miR-21 involvement.
Materials and Methods
Cell Culture
Human bronchial epithelial cells, BEAS-2B (ATCC, Manassas, VA, USA), were grown in Dulbecco’s modified Eagle’s F12 medium (DMEM/F12, Life Technologies, Carlsbad, CA, USA) supplemented with 10% fetal bovine serum (FBS; Hyclone, Logan, UT, USA). Human induced pluripotent stem cell-derived MSCs (iPSC-MSCs) were prepared as described in our previous study 26 . Briefly, urine cell-derived iPSCs donated by the Guangzhou Institute of Biomedicine and Health, Chinese Academy of Science (Guangzhou, Guangdong, China) were reprogrammed from human urine-derived cells via electroporation with the plasmid pEP4EO2SET2 K (which contained octamer-binding transcription factor 4 [OCT4], sex-determing region y-box 2 [SOX2], SV40 large T antigen [SV40LT], Kruppel-like factor 4 [KLF4]) 27 . U-iPSCs were maintained on Matrigel (BD Biosciences, San Jose, CA, USA) and cultured in iPSC medium that contained mTeSR1 (Stem Cell Technologies, Vancouver, Canada). MSCs were induced from iPSCs via MSC-inducing culture media and were passaged with MSC maintenance medium as described in our previous study 26 . Briefly, iPSCs were plated onto Matrigel-coated six-well culture plates. The passaged iPSCs were cultured in mTeSR1 medium for 2 days and subsequently cultured with MSC-inducing-culture media with minimum essential medium Eagle-α-modified (α-MEM; Gibco, Grand Island, NY, USA), 10% FBS (Gibco, Invitrogen Corporation, Carlsbad, CA, USA), penicillin/streptomycin, sodium pyruvate, 10 mM l-ascorbate-2-phosphate (Sigma-Aldrich, Inc., St. Louis, MO, USA), l-glutamine and non-essential amino acids for 2 weeks. The heterogeneous cells were subsequently passaged as a single cell suspension using Accutase® (Stem Cell Technologies) with a 1:2 split ratio. The cells were plated onto 0.1% gelatin-coated flasks (passage 1, P1). The iPSC-MSCs were cultured in DMEM-high glucose (DMEM-HG) supplemented with 10% FBS (Gibco), 5 ng/ml epidermal growth factor (EGF) (PeproTech, Rocky Hill, NJ, USA), 10 ng/ml basic-fibroblast growth factor (bFGF) (PeproTech), 0.1 mM non-essential amino acid (Millipore, Darmstadt, Germany), 1 mM L-glutamine and 0.1% β-mercaptoethanol. These cells were maintained in a humidified atmosphere of 5% CO2 and 95% air at 37°C and were routinely sub-cultured every 2–3 days. The iPSC-MSCs were morphologically highly similar to the classical MSCs, had the capacity to differentiate into osteoblasts, adipocytes, and chondroblasts, and exhibited a similar expression profile as surface antigens 26 .
Induction of Injured BEAS-2B and Co-culture with MSCs
CoCl2 is a hypoxia mimetic, which may stabilize hypoxia inducible factors (HIFs), induce the transcription of HIF target genes and cause a hypoxic injury and dysfunction to epithelial cells 28 . BEAS-2B cells, an airway epithelial cell line (1×105/well), were seeded in 6-well plates, treated with 0, 400, 600 or 800 μM CoCl2 for 12 h after their adherence to the plates and evaluated for apoptosis using flow cytometry. In different experiments of our study, BEAS-2B cells were treated with 400 μM CoCl2 for 24 h or 48 h or 800 μM CoCl2 for 12 h.
For the co-culture system, BEAS-2B cells (1×105/well) treated with 800 μM CoCl2 for 12 h were subsequently co-cultured with 2×104, 3×104, or 4×104/well MSCs for 24 h to determine BEAS-2B cell apoptosis. We determined that with the 4:10 ratio of MSCs: BEAS-2B cells, MSCs decreased apoptosis of injured BEAS-2B cells to nearly normal levels. Thus, we used this ratio of MSCs: BEAS-2B cells in the subsequent experiments. The cells were harvested, and apoptosis, miR-21 expression or ACVR2a expression were assessed in BEAS-2B cells. To investigate the effects of cell–cell contact of MSCs on BEAS-2B cells, BEAS-2B cells were seeded in a 12-well plate and treated with 800 μM CoCl2 for 12 h after adherence, followed by co-culture with MSCs for 24 h, which were seeded in the upper chamber of transwells (VWR, Fontenay-sous-Bois, France). BEAS-2B cells were subsequently examined for apoptosis using flow cytometry.
For the cell apoptosis assay, BEAS-2B cells in the co-culture systems with MSCs were sorted using flow cytometry. BEAS-2B cells were labeled with cell trace violet (1 μM, Molecular Probes, Invitrogen, Eugene, OR, USA) for 20 min and then seeded in the plates. For the RNA extraction of BEAS-2B cells in the co-culture systems, MSCs were labeled with green fluorescent protein (GFP), and BEAS-2B cells were sorted using flow cytometry.
MicroRNA Transfection and Treatments
BEAS-2B cells were seeded in a six-well plate (1×105/well). After the cells reached 60% confluence, miRNA-21 mimic (50 pmol) or miRNA-21 inhibitor (250 pmol) were transfected into BEAS-2B cells using lipo2000 reagent (Invitrogen). Transfection without the RNA sequence was used as a control. The cells were further treated with 400 μM CoCl2 8 h after transfection. BEAS-2B cells were harvested for RNA isolation or cell apoptosis examination at different time points after CoCl2 treatments. Apoptosis of BEAS-2B cells was analyzed using flow cytometry.
Flow Cytometry Assay and Cell Sorting
BEAS-2B cell apoptosis was detected using an Annexin V-FITC/propidium iodide (PI) kit (Miltenyi Biotec, Bergisch Gald-bach, Germany). In brief, BEAS-2B cells or cells of the co-culture system were dissociated with 0.25% trypsin solution without ethylenediaminetetraacetic acid (EDTA; Hyclone) and stained with 10 μl of Annexin V-FITC for 15 min in the dark at room temperature. After washing, the cells were stained with 5 μl of PI solution immediately prior to analysis with a flow cytometer. For the cells in the co-culture system, BEAS-2B cells were gated using the cell trace violet.
The ACVR2A expression in BEAS-2B was detected using flow cytometry. The cells were stained with APC-ACVR2A antibody (R&D Systems, Minneapolis, MN, USA) per the manufacturer’s protocol. All flow cytometry assays were performed using a Beckman flow cytometer (Beckman-Coulter, Miami, FL, USA), and the data were analyzed using Kaluza software (Beckman-Coulter).
To examine the mRNA expression in BEAS-2B cells co-cultured with GFP-MSCs, the cells were dissociated with 0.25% trypsin/EDTA solution (Gibco, Invitrogen Corporation) from the co-culture system, and BEAS-2B cells were collected by cell sorting with BD FACS AriaTM II (BD Biosciences, Heidelberg, Germany) for RNA extraction.
RNA Extraction and Quantification
BEAS-2B cells were collected as previously described. Total RNA was isolated using RNAisoTM Plus (TAKARA, Otsu, Japan). For miR-21 quantification, cDNA was synthesized with the M-MLV Reverse Transcriptase Kit (Promega, Madison, WI, USA). Mature miRNAs were quantified via stem-loop real-time polymerase chain reaction (PCR) with SYBR Premix ExTaq (TAKARA). The primers for the miRNAs used in the reverse transcription and quantitative PCR assays are as follows: Forward-miR-21: AGTGCGTGTCGTGGAGTC, reverse-miR-21: CACCAGTCGATGGGCTG. TR-hsa-miR-21-3p: GTCGTATCCAGTGCGTGTCGTGGAGTCGGCAATTGCACTGGATACGACacagccc, Forward-U6: CGCTTCACGAATTTGCGTGTCAT. Reverse&RT-U6: GCTTCGGCAGCACATATACTAAAAT.
Moreover, the activin A receptor type IIA (ACVR2A) mRNA expression in BEAS-2B was confirmed via quantitative real-time (qRT)-PCR. For mRNA quantification, total RNA was reverse transcribed with the Revert Aid First Strand cDNA Synthesis Kit (Thermo Scientific, Inc., Rockford, IL, USA), and the synthesized cDNA was quantified by SYBR (Roche, Basel, Switzerland) with a 7500 Fast system (ABI, Foster City, CA, USA). The Ct values were normalized to β-actin, and the relative expression was calculated using the 2-ΔΔCt method. The primer sequences were as follows: Forward-β-actin: TGAGACCTTCAACACCCCAGCCATG, Reverse-β-actin: CGTAGATGGGCACAGTGTGGGTG. Forward-hAcvr2a: GATGGAAGTCACACAGCCCA, Reverse-hAcvr2a: GGTCCTGGGTCTTGAGTTGG.
Western Blot Analysis
BEAS-2B cells were cultured under 400 μM CoCl2 with or without MSCs for 36 h. BEAS-2B cells were subsequently dissociated, sorted and lysed using radioimmunoprecipitation assay (RIPA) buffer with protease inhibitor (Pierce, Rockford, IL, USA). The lysed proteins were separated via sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE) and then transferred to polyvinylidene difluoride membranes (Roche Diagnostics, Indianapolis, IN, USA), which were probed with antibodies to p53 (1:1000) and actin (1:1000). Enhanced Chemiluminescence Plus (Millipore Corporation, Billerica, MA, USA) was used to visualize the membrane after incubation with horseradish peroxidase (HRP)–conjugated secondary antibodies. All primary antibodies were purchased from Abcam (Cambridge, MA, USA). Densitometric analyses of the immunoblots were performed using GenoSens Software.
Statistical Analysis
The data were expressed as the mean ± standard deviation (SD). All statistical analyses were performed with SPSS software, version 19.0 (IBM, Armonk, NY, USA). One-way analysis of variance ANOVA followed by Tukey's post hoc test (for equal homogeneity) or Dunnett T3 (for unequal homogeneity) were used for multiple comparisons between the different groups with Gaussian distribution data. A Kruskal–Wallis rank sum test followed by a Mann–Whitney U test were performed for the comparisons with non-normal distribution data. A P-value less than 0.05 was considered statistically significant.
Results
Co-culture with MSCs Attenuated Hypoxia-Induced Apoptosis of BEAS-2B Cells
To investigate the protection of MSCs to pulmonary epithelial cells induced by hypoxia, BEAS-2B cells were exposed to CoCl2. The CoCl2 treatments induced both early and late apoptosis of BEAS-2B cells in a dose-dependent manner, and there was clear apoptosis for 400 μM, 600 μM and 800 μM (Figure 1(a)). Next, BEAS-2B cells were treated with 400 μM CoCl2 for 24 h or 48 h or 800 μM CoCl2 for 12 h. The injured BEAS-2B cells were co-cultured with MSCs at different ratios. In the positive control group, CoCl2 treatment substantially induced both early and late apoptosis in BEAS-2B cells. The early and late apoptosis rates of BEAS-2B cells increased to 18% and 26%, respectively (Figure 1(a) and (b)). However, co-culture with MSCs (2×104, 3×104 or 4×104/well) significantly decreased the apoptotic ratio of BEAS-2B cells (1×105/well) in a dose-dependent manner (p < 0.01 or p < 0.001). The apoptotic ratio of BEAS-2B cells reached normal levels with 4% and 6% for early and late apoptosis, respectively, when the ratio of MSCs (4×104/well): BEAS-2B cells (1×105/well) was 4:10 (Figure 1 [b]). Moreover, the protection of MSCs on BEAS-2B cells was further evaluated by detecting p53 expression using Western blotting, which was responsible for cell apoptosis 29 . We determined that the CoCl2 treatment increased the p53 expression in BEAS-2B cells, which was blocked after co-culturing with MSCs (Figure 1(c)). We subsequently investigated the role of cell–cell contact in MSC inhibition effects on apoptosis of BEAS-2B cells using transwells. We determined that transwells significantly reversed the inhibition of MSCs on apoptosis of BEAS-2B cells. Moreover, BEAS-2B cells separated with MSCs by transwells exhibited less early apoptosis and late apoptosis (Figure 1(d)), which suggests that both cell–cell contact and soluble factors were involved in the protection of MSCs on epithelial apoptosis.
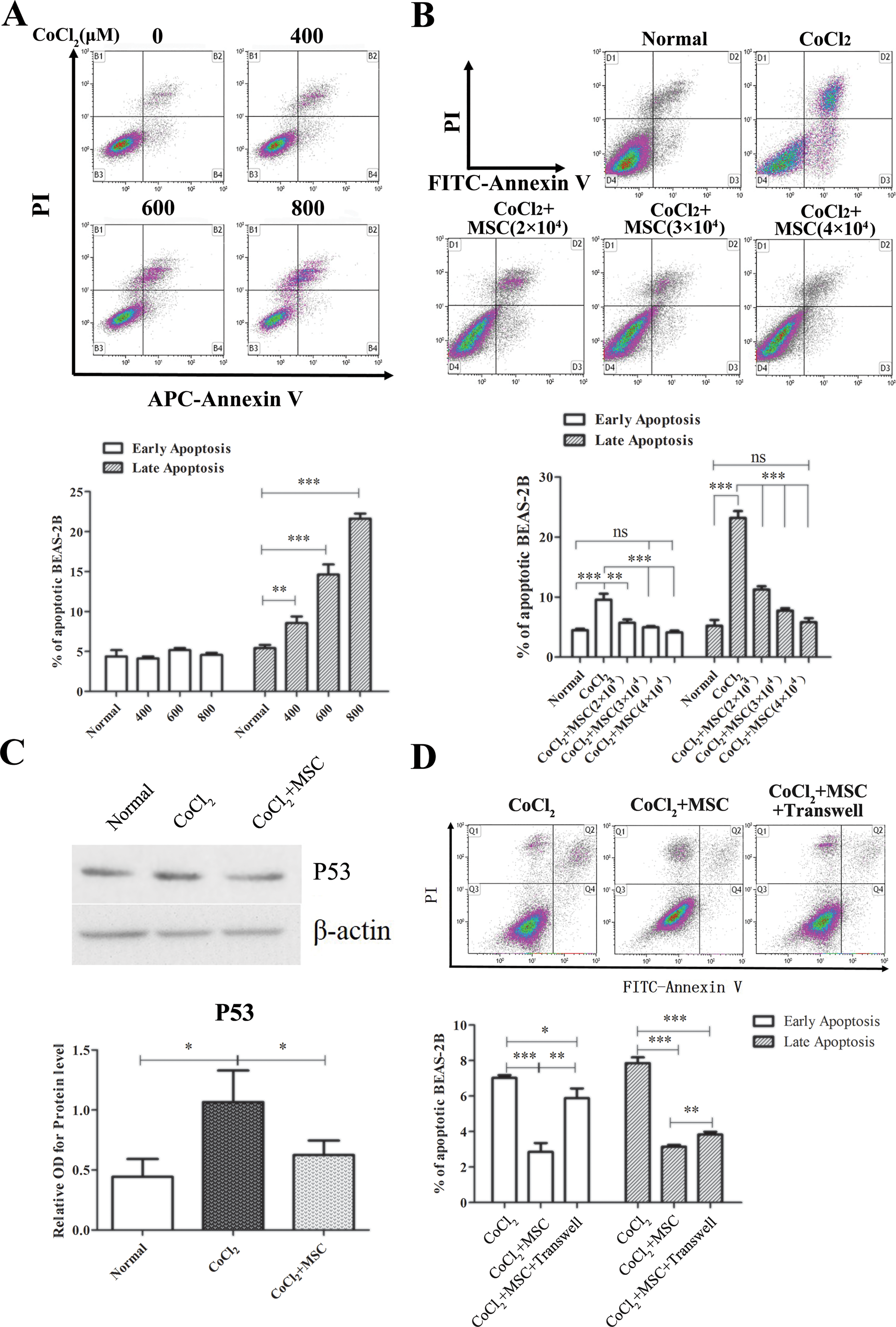
Co-culture with MSCs attenuated hypoxia-induced apoptosis. (a) BEAS-2B cells were cultured with different concentrations of CoCl2 (0, 400, 600, and 800 μM, respectively) for 12 h and subsequently cultured in complete medium for 24 h. Apoptosis was examined using flow cytometry. (b–c) BEAS-2B cells (1×105/well) were labeled with cell trace violet, seeded in a six-well plate and treated with 800 μM CoCl2 for 12 h after adherence. BEAS-2B cells were further co-cultured with MSCs with different concentrations for 24 h. BEAS-2B cells were analyzed via an apoptosis assay (b) or sorted for Western blot (c). The normal group was not cultured with CoCl2 or MSCs. (d) BEAS-2B were cultured with 800 μM CoCl2 and co-cultured with MSCs for 24 h using transwell. Data are representative of three separate experiments. *p < 0.05; **p < 0.01; ***p < 0.001.
Co-culture with MSCs Up-regulated miR-21 Expression in Human Bronchial Epithelial Cells of BEAS-2B
We have previously reported that the infusion of MSCs alleviated Th2 inflammation and pulmonary injury, and it may be involved in the mmu-miR-21/ACVR2A axis in an ovalbumin (OVA) induced asthma mouse model 17 . However, whether MSCs protect the injured bronchial epithelial cells induced by hypoxia via miR-21 remains unknown. We subsequently examined the miR-21 expression in BEAS-2B cells after CoCl2 stimulation. No difference in miR-21 expression was identified in BEAS-2B cells at different time points after CoCl2 treatment (Figure 2(a)). Interestingly, after co-culture with MSCs, the miR-21 expression increased in BEAS-2B cells under CoCl2 stimulation (Figure 2(b)). These data indicated that miR-21 may be involved in the protection of MSCs to injured human bronchial epithelial cells.

Co-culture with MSCs up-regulated miR-21 expression in BEAS-2B cells. (a) BEAS-2B cells were treated with 400 μM CoCl2 for 0, 12 h, 24 h and 36 h. The relative expression of miR-21 was examined via qRT-PCR. (b) BEAS-2B cells were treated with 800 μM CoCl2 for 12 h and subsequently cultured with GFP-labeled MSCs for 24 h. BEAS-2B cells were sorted, and the miR-21 mRNA expression was examined. Data are shown as the mean ± SD. ***p < 0.001.
MiR-21 Mediated the Anti-apoptotic Effects of MSCs to Bronchial Epithelial Cells
We subsequently examined the role of miR-21 in the early and late apoptosis of BEAS-2B cells. We transfected BEAS-2B cells with miR-21a mimic (Figure 3(a)) or miR-21a inhibitor (Figure 3(b)) or their controls prior to exposure to CoCl2. For the miR-21 control, the total ratio of early and late apoptosis for BEAS-2B cells without CoCl2 stimulation was approximately 12%. The transfection of miR-21 mimics significantly reduced the early (p < 0.01) and late (p < 0.05) apoptosis rates (compared with those of the miR-21 control). Under the CoCl2 stimulation, the late apoptosis rate of BEAS-2B cells in the miR-21 control group increased to 12%. However, pre-transfection of miR-21 mimics significantly decreased the late apoptosis of BEAS-2B cells (p < 0.001) (Figure 3(a)). These data suggested that miR-21 exerts anti-apoptosis effects for BEAS-2B cells. We subsequently investigated the effects of a miR-21 inhibitor on cell apoptosis. The transfection of miR-21 inhibitor significantly increased both early (p < 0.01) and late (p < 0.001) apoptosis ratios in BEAS-2B cells without CoCl2 treatments. Moreover, compared with the CoCl2 group, the pre-transfection of miR-21 inhibitor combined with CoCl2 treatment induced a significant increase of early apoptosis in BEAS-2B cells (p < 0.05) (Figure 3(b)). Increased late apoptosis was also identified without statistical significance. More importantly, we determined that the protective effects of MSCs on injured BEAS-2B cells with the CoCl2 stimulation were significantly reversed with the transfection of miR-21 inhibitor (Figure 3(c)). There was a higher apoptosis ratio particularly for late apoptosis for the MSC-treated BEAS-2B if they were transfected with miR-21 inhibitor (p < 0.05) (Figure 3(c)). These data suggested that miR-21 is an important factor to mediate the protective effects of MSCs to hypoxia-induced bronchial epithelial cell injury.

MiR-21-mediated the anti-apoptotic effects of MSCs to hypoxia-induced BEAS-2B cell injury. Beas-2B cells were transfected with miR-21 mimic (a) or miR-21 inhibitor (b), exposed to CoCl2 for 12 h and cultured in complete medium for 24 h. Cells were collected for apoptosis assay. A random sequence was used as a control for the miR-21 mimic group. The administration of lip2000 was used as a control for the miR-21 inhibitor group. (c) BEAS-2B cells were labeled with cell trace violet for 20 min, seeded in plates, transfected with miR-21 inhibitor and exposed to CoCl2 for 12 h, followed by co-culture with or without MSCs for an additional 24 h. Apoptosis of cell trace positive BEAS-2B cells was analyzed using flow cytometry. Data are representative of three separate experiments. Data are shown as the mean ± SD. *p < 0.05; **p < 0.01; ***p < 0.001.
ACVR2A was Involved in CoCl2-Induced Cell Injury
We have previously reported that mouse ACVR2A was targeted with mouse miR-2117. We searched the online Computational Medicine Center and determined that there were binding sites of miR-21 in the 3’UTR of human ACVR2A (Figure 4(a)). We subsequently investigated the expression of ACVR2A in the injured BEAS-2B and the treatment of MSCs. The treatment of CoCl2 up-regulated the ACVR2A expression in a time-dependent manner (Figure 4(b)). The ACVR2A mRNA expression was significantly up-regulated at 24 h (up to four-fold) and 36 h (up to 10-fold) after the stimulation of CoCl2 compared with the normal cells (both p < 0.001), and there were higher levels of ACVR2A at 36 h compared with 24 h (p < 0.001) (Figure 4(b)). Next, BEAS-2B cells were stimulated with CoCl2 for 36 h, and the ACVR2A protein levels were evaluated using a flow cytometer. In the normal group, there were approximately 9% ACVR2A positive BEAS-2B cells with 6% moderate and 3% high, respectively. CoCl2 treatment significantly increased the percentage of ACVR2A positive cells with 9% ACVR2Amoderate and 8% ACVR2Ahigh (p < 0.001) (Figure 4(c)). Furthermore, BEAS-2B cells exhibited higher ACVR2A expression following co-culture with MSCs (p < 0.001) (Figure 4(d)). These data suggested that hypoxia on epithelial cells increases ACVR2A expression at both the mRNA and protein levels, and the treatment of MSCs also increased the ACVR2A expression.
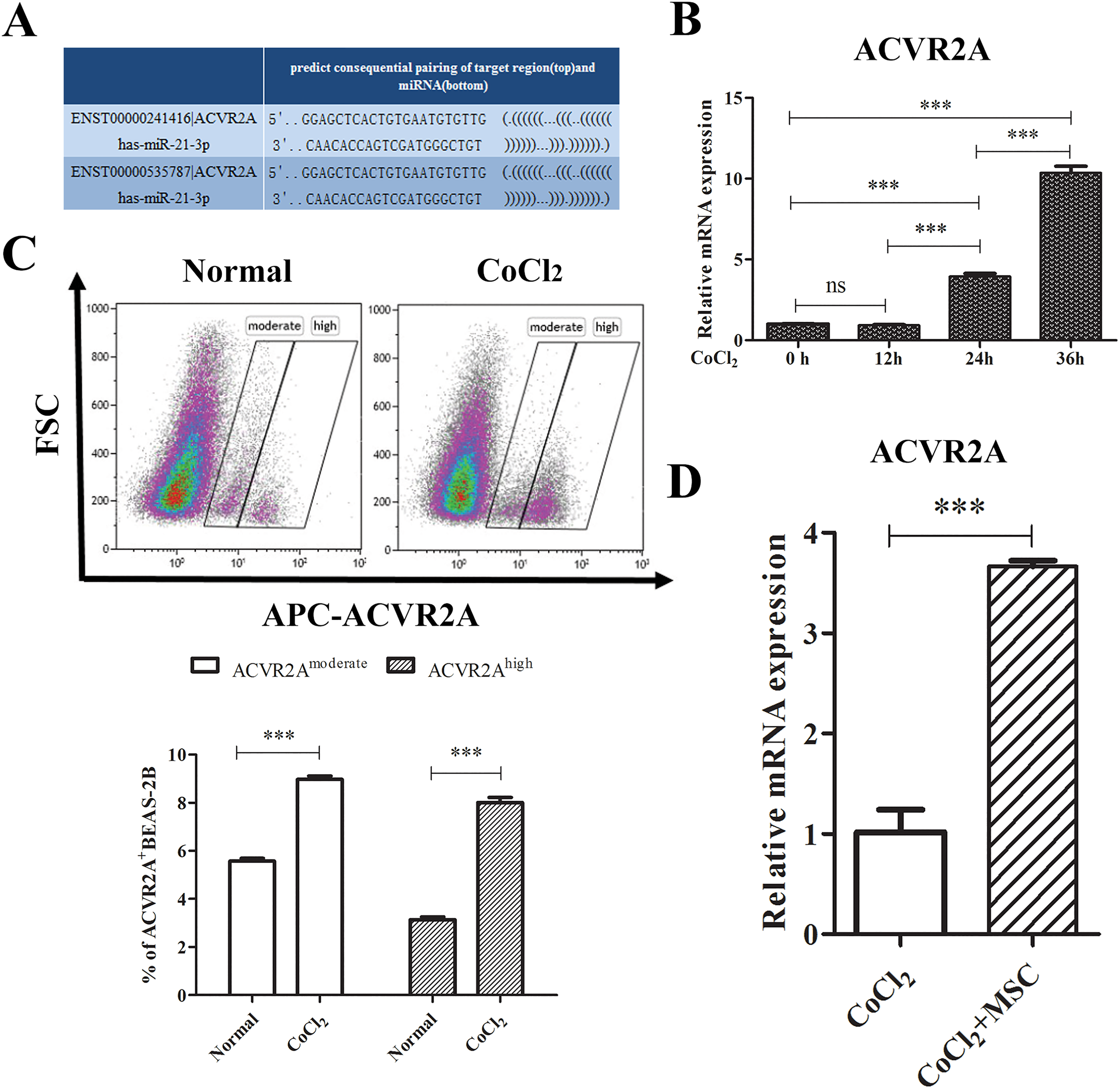
ACVR2A expression increased in CoCl2 induced BEAS-2B cells. (a) Schematic indicates the interaction sites for human miR-21, which is harbored in the 3’UTR of human ACVR2A. (b) BEAS-2B cells were treated with 400 μM CoCl2 for 0, 12 h, 24 h and 36 h. The relative expression of ACVR2A mRNA was examined via qRT-PCR. (c) BEAS-2B cells were treated with 400 μM CoCl2 for 36 h, and the ACVR2A protein level was examined using flow cytometry. (d) BEAS-2B cells were treated with 400 μM CoCl2 for 12 h and subsequently co-cultured with or without MSCs for 36 h. BEAS-2B cells were harvested and isolated by cell sorting for PCR assay. Data are shown as the mean ± SD from three independent experiments. ***p < 0.001.
Transfection of MiR-21 Mimic Up-regulated ACVR2A Expression
We subsequently investigated the effects of a miR-21 inhibitor or miR-21 mimic on the ACVR2A expression 36 h after CoCl2 treatment. Pre-transfection of the miR-21a mimic had no effects on the ACVR2A mRNA expression in normal cells; however, it led to a significant increase of ACVR2A under the treatment of CoCl2 (p < 0.05) (Figure 5(a)). Moreover, we determined that pre-transfection of the miR-21 inhibitor significantly decreased the ACVR2A mRNA expression in the injured BEAS-2B cells. More importantly, pre-transfection of the miR-21 inhibitor significantly reversed the effects of MSCs on increasing the ACVR2A expression (p < 0.001) (Figure 5(b)), which suggests that MSCs increased the ACVR2A expression, at least in part, via miR-21. We obtained similar results for the ACVR2A protein levels under the transfection of the miR-21 inhibitor or miR-21 mimic. Pre-transfection of the miR-21 mimic or miR-21 inhibitor resulted in opposite effects on the ACVR2A protein levels. Transfection of the miR-21 mimic induced a substantial increase of ACVR2A expression (38% for ACVR2Amoderate and 35% for ACVR2Ahigh) compared with the miR control under the treatment of CoCl2 (p < 0.001) (Figure 5(c)). Moreover, transfection of the miR-21 inhibitor significantly decreased ACVR2A protein to nearly normal levels compared with the controls (3.3% are ACVR2Amoderate and 4.1% are ACVR2Ahigh) (p < 0.001) (Figure 5(d)). Taken together, our data suggest a positive effect of miR-21 on ACVR2A expression in injured BEAS-2B under hypoxia.

MiR-21 regulated CoCl2-induced ACVR2A expression. BEAS-2B cells were transfected with miR-21 mimic (a and c) or inhibitor (b and d) and cultured in the presence or absence of CoCl2 for 36 h. (a, b) ACVR2A mRNA expression was detected via qRT-PCR. (c–d): Percentage of ACVR2A-positive BEAS-2B cells was detected using flow cytometry. Data are shown as the mean ± SD from three independent experiments. (b) BEAS-2B cells were treated with MSCs.
Discussion
Respiratory epithelial cells play important roles in the maintenance of proper homeostasis 3 . Asthma is a chronic inflammatory disease that frequently accompanies hypoxic conditions characterized by inflammation of the airways, during which the airways become inflamed and constricted. Evidence has indicated that respiratory epithelial cells were structurally and functionally abnormal in asthma 4 . MSC therapy is a promising source for tissue repair and immunotherapy, and these cells have been widely used in clinical and scientific research. MSCs have shown a trophic effect for injured kidneys, and they have been considered to exert a potential contribution to tubular epithelial cell regeneration and recovery from acute renal failure (ARF) 14 . We have also shown that the infusion of MSCs attenuated lung inflammation in an OVA-induced asthmatic mouse model. However, the effect of MSCs on airway epithelial cell injury remain unclear. Here, we determined that treatment of CoCl2 lead to apoptosis in human bronchial epithelial cells. However, co-culture with MSCs significantly protected the injured epithelial cells during hypoxia, which was dependent on both cell-to-cell contact and soluble factors. Furthermore, we determined that MSCs significantly decreased the high levels of apoptosis-related protein p53 under the stimulation of CoCl2. This finding is consistent with the protective effects of MSCs on renal epithelial cells 14 . Many cells may be injured during hypoxia. Our data have shown that MSCs protected epithelial cells during hypoxia and suggested the potential wide application of MSCs on injured cells during hypoxia.
microRNAs have been shown to be involved in the development of asthma. MiR-21 was up-regulated and appeared to be a biomarker for asthma diagnosis 24,25 . We have previously reported that miR-21 was up-regulated in an OVA-induced asthmatic mouse model, and the treatment of MSCs down-regulated miR-21 expression and alleviated lung inflammation 17 . These data led us to investigate whether MSCs protect respiratory epithelial cells via miR-21. MiR-21 has been reported to be an anti-apoptotic factor in human glioblastoma cells 30 and prostate cancer cells 31 . By suppressing the tumor suppressor phosphatase and tensin homologue (PTEN) 32 and Fas-ligand 33 , miR-21 also appeared to serve as a biomarker for non-small cell lung cancer 34 , hepatocellular carcinoma 35 and esophageal squamous cell carcinoma 36 . Here, we determined that pre-transfection of miR-21 mimic inhibited the pro-apoptotic effect of CoCl2 on epithelial cells. In contrast, pre-transfection of a miR-21 inhibitor increased apoptosis of epithelial cells of BEAS-2B. These data suggested that miR-21 exerts the protection on epithelial cells particularly during hypoxia. More importantly, the protective effects of MSCs on epithelial cells were significantly reversed by pre-transfection of a miR21 inhibitor, which suggests that miR-21 played an important role in the effects of MSCs on epithelial cells. In addition, an up-regulation of miR-21 in injured BEAS-2B cells was identified when co-cultured with MSCs but not in injured BEAS-2B cells cultured alone. These data provided further evidence that miR-21 mediates, at least in part, the protective effects of MSCs to injured BEAS-2B cells.
We have previously reported that the miR-21-ACVR2A axis was important in a mouse model of asthma with the treatment of MSCs. We searched the online Computational Medicine Center database and determined that there were binding sites of miR-21 in the 3’UTR of human ACVR2A. We identified high ACVR2A levels of mRNA and protein during the hypoxia of CoCl2. Furthermore, transfection of a miR-21 mimic significantly up-regulated the ACVR2A expression, whereas transfection of a miR-21 inhibitor down-regulated the ACVR2A expression. These data suggest that ACVR2A is downstream of miR-21. We have previously reported that mouse ACVR2A was targeted by mouse miR-21 and down-regulated in an asthmatic mouse model; moreover, it recovered to a normal level after MSC treatment 17 . Furthermore, we determined that MSC treatment increased the ACVR2A levels at the same time as protecting injured BEAS-2B cells, which suggests ACVR2A plays a role of anti-apoptosis in BEAS-2B cells.
ACVR2A is the receptor type II A of Activin, a member of the transforming growth factor-beta (TGF-β) superfamily. Activin binds with ACVR2A and initiates the intracellular transmission of signals. Activin has exhibited complex functions with a pro-inflammatory role and an anti-inflammatory role 37 . Moreover, expression of ACVR2A also exhibited different and, in some cases, completely opposite activities in different and the same disease or condition. For example, there was increased ACVR2A expression in asthma after an allergen challenge 38 . However, other studies reported a decreased frequency of ACVR2A expression in epithelial cells in asthma 39,40 . Here, we hypothesize that ACVR2A was an anti-apoptotic factor and was up-regulated in response to CoCl2 treatment to protect cells from apoptosis. We determined that pre-transfection of a miR-21 inhibitor down-regulated the ACVR2A expression, whereas pre-transfection of a miR-21 mimic up-regulated a further increase of ACVR2A after CoCl2 treatment. Nevertheless, it was inconsistent with a previous report that pre-transfection of a miR-21 inhibitor increased the ACVR2A expression after lipopolysaccharide (LPS) treatment 17 . This difference may be because LPS and CoCl2 induced different cellular injuries via different pathways. LPS exerts pro-inflammatory effects via the toll-like receptor 4 (TLR4) signaling pathway 41 and has been reported to induce a rapid increase of ACVR2A in a short time of 1.5 h 42 . CoCl2 induces cell injury by providing a condition similar to hypoxia, in which the increase of ACVR2A expression requires a longer time. We acknowledge several limitations in our study. The relationship between miR-21 and ACVR2A requires further clarification via luciferase assay. Blocking ACVR2A function should be performed to further examine the role of ACVR2A in the effects of MSCs on injured epithelial cells. As previously discussed, both ACVR2A and activin are complex factors and exhibit unexpected activities in different conditions. It is all possible that ACVR2A plays positive, negative or no functions in the effects of MSCs on epithelial cells. It should be carefully and clearly investigated in the future.
Conclusion
To summarize this article, we have provided a schematic figure to indicate the effects of MSCs on BEAS-2B cells injured by CoCl2 and the role of miR-21 (Figure 6). The treatment of CoCl2 induces apoptosis in the epithelial cells of BEAS-2B. Co-culture with MSCs attenuated this apoptosis via miR-21 and up-regulated miR-21 expression in BEAS-2B cells. In response to CoCl2, ACVR2A is up-regulated and MiR-21 enhances ACVR2A expression during hypoxia. The roles of ACVR2A in cell apoptosis and the protective effects of MSCs should be further investigated.

The effects of MSCs on BEAS-2B cells injured by hypoxia via miR-21 in this article. CoCl2, a hypoxia mimic, induces apoptosis of epithelial cells of BEAS-2B cells. MSCs attenuate apoptosis of epithelial cells via miR-21. Both the stimulation of CoCl2 and miR-21 increased the ACVR2A expression. The roles of ACVR2A in cell apoptosis and the protective effects of MSCs should be further investigated.
In general, MSCs are isolated from adult tissues of bone marrow (BM) or adipose tissue. However, there are several potential limitations of using adult MSCs, including their limited capacity for proliferation, the significant variability in cell quality derived from different donors and the rapid loss of multipotentiality 43,44 . We have previously determined that iPSC-MSCs exhibited a higher proliferation rate with less cell senescence than BM-MSCs 26 . Furthermore, we reported that human iPSC-MSCs are insensitive to pro-inflammatory-induced human leukocyte antigen (HLA)-II expression and have a stronger immune privilege after transplantation 45 . Moreover, iPSC-MSCs exerted good stability and did not induce tumor formation 46 ; however, the safety concerns associated with reprogramming using viruses and the genomic instability of iPSCs require careful evaluation.
In conclusion, we determined that MSCs protected epithelial cells during hypoxia with the treatment of CoCl2 via miR-21. These findings provide a novel mechanism for the repair function of MSCs and a basis for the application of MSCs in clinical therapy.
Footnotes
Acknowledgements
Cheng-Lin Li, Zhi-Bin Xu These authors contributed equally to this manuscript.
Ethical Approval
Ethical Approval is not applicable for the article.
Statement of Human and Animal Rights
Statement of Human and Animal Rights is not applicable for the article.
Statement of Informed Consent
Statement of Informed Consent is not applicable for the article.
Declaration of Conflicting Interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The authors disclosed receipt of the following financial support for the research and/or authorship of this article: This study was supported by grants from the NSFC for Excellent Young Scholars (81322012 to Prof. QL Fu), NCET-13-0608 to Prof. QL Fu, NSFC (81373174, 81272062, 81471832 and 81770984) and the key grant from the Science and Technology Foundation of Guangdong Province of China (2015B020225001), and the Natural Science Foundation of Guangdong Province (2014A030313051 and 2016A030308017).