
Abstract
Although emerging evidence links mesenchymal stem cells (MSCs) with cancer metastasis, the underlying mechanisms are poorly understood. In the present study, we found that human umbilical cord-derived MSCs (UC-MSCs) promoted MCF-7 cell migration in vitro and metastasis in vivo. To explore the mechanisms, the characteristics of MCF-7 cells cocultured with UC-MSCs were assessed. The expression and secretion of interleukin-8 (IL-8) and IL-6 were induced in MCF-7 cells cocultured with UC-MSCs. However, neutralization of IL-8 or IL-6 secreted by UC-MSCs could attenuate the enhanced expression of IL-8 and IL-6 in MCF-7 cells cocultured with UC-MSCs, which subsequently alleviated the enhanced migration. Similar to UC-MSCs, exogenous human recombinant IL-8 or IL-6 also promoted IL-8 and IL-6 expression and MCF-7 cell migration. In addition to enhanced IL-8 and IL-6 expression, MCF-7 cells cocultured with UC-MSCs displayed enhanced mammosphere-forming ability and increased percentage of CD44+/CD24- cells. However, epithelial-to-mesenchymal transition (EMT) was not observed in MCF-7 cells cocultured with UC-MSCs. Taken together, these results suggested that IL-8 and IL-6 secreted by UC-MSCs activated the autocrine IL-8 and IL-6 signaling in MCF-7 cells and induced CD44+/CD24- cells, which subsequently promoted MCF-7 cell migration in vitro and metastasis in vivo.
Keywords
Introduction
Mesenchymal stem cells (MSCs) are considered as promising tools for immunomodulation and tissue regeneration (18). The fact that MSCs can home to cancer has prompted researchers to use MSCs as vectors for the targeted therapy of cancer (13,15,41). Several reports have confirmed the therapeutic efficacy of MSCs in cancer (24,36). However, evidence suggests that interactions between MSCs and cancer cells may promote their metastatic potential (10,17,31). Karnoub et al. reported that MSCs enhanced breast cancer metastasis when they were coinjected with breast cancer cells. The interaction of chemokine (C-C motif) ligand 5 (CCL5) secreted by MSCs and C-C chemokine receptor type 5 (CCR5) expressed in cancer cells played an important role in MSC-mediated cancer metastasis (17). Subsequently, it was shown that MSCs promoted migration of breast cancer cells by secreting stromal cell-derived factor-1 (SDF-1), interleukin-6 (IL-6), and vascular endothelial growth factor (VEGF) (7,10,31). Although the cross talk between MSCs and cancer cells is gradually being revealed, the exact mechanisms remain unknown. Recent studies focus on the effect of epithelial-to-mesenchymal transition (EMT) and cancer stem-like cells (CSCs) on cancer metastasis (3,39). EMT is a process essential to organogenesis during embryonic development (29). It has recently been documented to promote the detachment of cancer cells from the primary cancer and facilitate their subsequent migration to other organs (3,39). It was shown that breast cancer cells with a CD44+/CD24- cell surface profile were capable of driving tumor development and were proposed as CSCs (1). Furthermore, CD44+/CD24- breast cancer cells were associated with enhanced invasion (33). Since both EMT and CD44+/CD24- cells were implicated in metastasis, a connection between EMT and CD44+/CD24- cells was proposed. Mani et al. demonstrated that induction of EMT caused an increase in CD44+/CD24- cells and cancer initiation frequency (26). Moreover, it was shown that CD44+/CD24- cells expressed EMT markers (16). On the contrary, another study showed that the signaling pathways governing CSCs may not be identical to those pathways fostering EMT (11).
Multiple cytokines, secreted either by cancer cells or cells in the cancer microenvironment, lead to cancer survival, growth, and progression (19,27). For example, elevated serum IL-8 level has been reported in patients with pancreas, breast, or prostate cancer. Furthermore, a positive correlation was established between serum IL-8 level and diminished survival of breast cancer patients (2,20). Recently, it was shown that IL-8 and IL-6 could induce EMT (12,37). There are evidences that IL-8 and its receptor, chemokine (C-X-C motif) receptor 1 (CXCR1), play an important role in inducing CSCs. Moreover, blocking CXCR1 decreases the number of CSCs and inhibits metastasis (14,35).
Compared with bone marrow-derived MSCs (BM-MSCs), UC-MSCs have several advantages, such as no ethical concern, painless collection procedure, better expandable ability, and low immunogenicity (25,42). Therefore, UC-MSCs possess great potential in clinical application. Our research group showed that UC-MSCs exhibited great therapeutic potential in several disease models, such as liver fibrosis, stroke, acute lung injury, and immune-mediated disorders (22,30,38,45). Recently, we found that UC-MSCs inhibited the proliferation and induced the differentiation of leukemia cells (9,40). However, it is unknown if UC-MSCs accelerate solid cancer growth and metastasis. In the present study, we will investigate if UC-MSCs promote breast cancer growth and metastasis. Based on the fact that UC-MSCs secrete a variety of cytokines, we will explore cytokines, which may contribute to breast cancer metastasis. At the same time, we will determine if CD44+/CD24- cells and EMT are involved in UC-MSC-induced breast cancer metastasis.
Materials and Methods
Reagents
Phycoerythrin (PE)-conjugated anti-chemokine (C-X-C motif) receptor 4 (CXCR4), CCR5 and CCR7 antibodies, human IL-8 and IL-6 neutralizing antibodies, and their isotype antibodies were from R&D Systems (Minneapolis, MN, USA). PE-conjugated anti-E-cadherin and vimentin antibodies, fluorescein isothiocyanate (FITC)-conjugated anti-CD24 antibody, allophycocyanin (APC)-conjugated anti-CD44 antibody, and their isotype controls were from BD Pharmingen (San Jose, CA, USA). Recombinant IL-8, IL-6, epidermal growth factor (EGF), and basic fibroblast growth factor (bFGF) were from Peprotech (London, UK).
Isolation and Culture of UC-MSCs
UC-MSCs were isolated from three umbilical cords obtained from a local maternity hospital with donors’ informed consent. Human tissue collection for research was approved by the Institutional Review Board of the Chinese Academy of Medical Science. The details of isolation, expansion, and identification of UC-MSCs were essential as described previously (22,25,30). Briefly, cords were cut into pieces and digested with 0.075% collagenase II (Sigma-Aldrich, St. Louis, MO, USA) for 30 min and then 0.125% trypsin (Gibco, Grand Island, NY, USA) for another 30 min at 37°C. The digested mixture was passed through a 100-μ filter (Beyotime Biotech, Haimen, China) to obtain single cell suspensions. Cells were plated onto plastic flasks (Corning, Tewksbury, MA, USA) in DMEM/F12 medium (Gibco) supplemented with 10% fetal bovine serum (FBS) (Hyclone, Logan, UT, USA), 100 U/ml penicillin–streptomycin (Sigma-Aldrich), 2 mmol/L glutamine (Sigma-Aldrich), and 10 ng/ml EGF. Cultures were maintained at 37°C in a humidified atmosphere containing 5% CO2 and passaged at 80% confluence. UC-MSCs at passages 4-6 were employed.
Culture of Human Breast Cancer Cell Line
MCF-7 cells were purchased from Cell Center of Basic Medical Institute, Chinese Academy of Medical Sciences. Cells were grown in DMEM/F12 medium supplemented with 10% FBS (Hyclone), 100 U/ml penicillin-streptomycin (Sigma-Aldrich), 2 mmol/L glutamine (Sigma-Aldrich), and 1x nonessential amino acids (Invitrogen, Grand Island, NY, USA). Cultures were maintained at 37°C in a humidified atmosphere containing 5% CO2.
Transwell Coculture Experiment
When UC-MSCs in six-well plates (Corning) became 80% confluent, cells were irradiated (cesium-137, 30 Gy; low dose rate research irradiator, Gammacell 40 Exactor; MDS Nordion, Sweden) to inhibit proliferation. After irradiation, hanging cell culture inserts with 0.4-μm pore size permeable membrane (Millipore, Darmstadt, Germany) were put on six-well plates to separate MCF-7 cells from UC-MSCs. MCF-7 cells were seeded onto inserts at a density of 2 × 105 cells/insert and cultured for 48 h.
In Vitro Migration and Invasion Assay
For the migration assay, when UC-MSCs in 24-well plates (Corning) became confluent, hanging cell inserts with 8-μm pore size permeable membrane (BD Biosciences, San Jose, CA, USA) were put on 24-well plates, and 2.0 × 104 MCF-7 cells after serum starvation for 4 h were plated onto inserts and cultured for 18 h. MCF-7 cells remaining on the top side of the inserts were removed, and migratory cells to the bottom side were fixed with methanol (Sigma-Aldrich) and stained with hematoxylin and eosin (Beyotime Biotech). Six random fields per insert were photographed, and migratory cells were counted.
The protocol of the invasion assay was similar to that of the migration assay, and 5.0 × 104 MCF-7 cells were plated onto Matrigel (BD Biosciences)-coated inserts with 8-μm pore membrane. After cells were incubated for 24 h, noninvasive cells were removed. The invasive cells were fixed, stained, photographed, and counted.
Proliferation Assay
MCF-7 cells were seeded at a density of 2,000/well in 96-well plates and were incubated for 48 h in DMEM/F12 medium without or with conditioned medium (CM) of UC-MSCs (DMEM/F12 medium/UC-MSCs CM = 1:1). At the end of incubation, cell counting kit-8 (CCK-8; Beyotime Biotech) reagent was added into the wells, and cells were incubated for another 1-4 h (defined according to the density of cells) at 37°C. The absorbance at 450 nm was measured with a microplate reader (Biotek, Winooski, VT, USA).
Quantitative Real-Time PCR Analysis
After being cocultured or cultured alone for 48 h, UC-MSCs, and MCF-7 cells were collected. Total RNA was extracted with TRIzol (Invitrogen), following the manufacturer’ s protocol. After RNA extraction, cDNA synthesis was performed using MLV RT (Promega, Madison, WI, USA) in the presence of oligo-dT primer. Based on the nucleic acid sequence, specific primers were designed (Table 1). The level of mRNA expression was measured using SYBR Green Master Mix on an Applied Biosystems 7500 Real-time PCR system. The relative quantification of genes was calculated with GAPDH as a reference.
Sequence of Primers Specific for Cytokines, Chemokine Receptors, and EMT Markers

Quantification of Cytokines by ELISA
Cell-free media were collected and kept frozen at −80°C. IL-8, IL-6, and VEGF ELISA kits were purchased from Xinbosheng Biotech (Shenzhen, China). Prostaglandin E2 (PGE2) ELISA kit was purchased from Cayman Chemicals (Ann Arbor, MI, USA). All assays were carried out according to the protocols provided by the suppliers.
Flow Cytometry
MCF-7 cells cultured alone or cocultured with UC-MSCs were harvested and washed twice with PBS (Beyotime Biotech). Then cells were labeled within PE-conjugated antibody of CXCR4, E-cadherin, or vimentin, FITC-conjugated antibody of CD24, and APC-conjugated antibody of CD44 and their isotype controls for 45 min at 4°C. The final concentration of antibody was 10 μg/ml. After incubation, cells were washed with PBS and resuspended in 2% paraformaldehyde (Sigma-Aldrich) followed by flow cytometric analysis (LSR II; BD Biosciences).
Cell Sorting
MCF-7 cells cocultured with UC-MSCs for 48 h were trypsinized and suspended in PBS plus 1% FBS at a density of 1 × 107 cells/ml. FITC-conjugated antibody of CD24 and APC-conjugated antibody of CD44 were added and incubated on ice for 40 min. The final concentration of antibody was 10 μg/ml. After being washed with PBS containing 1% FBS, labeled cells were sorted by CD44 and CD24 markers on a BD FACS AriaIII cell sorter. CD44+CD24- and CD44+CD24+ cells were harvested.
Mammosphere Culture
MCF-7 cells cultured alone or cocultured with UC-MSCs were seeded at 2.5 × 104 cells/ml in six-well ultralow adherence plates (Corning) and cultured for 7 days in serum-free DMEM/F12 medium with 20 ng/ml EGF, 20 ng/ml bFGF, 1x B27 supplement (Invitrogen). At the end of mammosphere culture, six random fields per well were photographed. The number and size of mammospheres were compared between different groups.
Mouse Xenograft Model
Five-week-old female Balb/c athymic nude mice were purchased from Vital River Laboratory Animal Technology Co. Ltd. (Beijing, China). Mice were housed in specific pathogen-free conditions, and 1 × 107 MCF-7 cells in 100 μl serum-free RPMI-1640 medium (Gibco) were injected subcutaneously into the dorsum of the right forelimb. When tumors reached the mean size of 50 mm3, intravenous injection of 4 × 104, 2 × 105, 1 × 106 UC-MSCs or PBS was carried out at days 0 and 14. Eight mice were used for each group. The volume of tumors was measured with a caliper every 3 days for 42 days. Tumors and organs such as lung, liver, heart, spleen, and kidney were kept in 1% formalin (Sigma-Aldrich) solution for histologic analysis.
Histologic Analysis
Tissues were fixed in formalin (Sigma-Aldrich) and then embedded in paraffin (Sigma-Aldrich). Sections (4 μm thick) were deparaffinized, rehydrated, and stained by hematoxylin and eosin (Beyotime Biotech).
Statistical Analysis
Data were presented as mean ± SD. Differences between group means were determined by Student's t-test or Newman–Keuls (SNK) test after one-way ANOVA analysis. Values of < 0.05 were considered statistical significance (p < 0.05, p < 0.01).
Results
UC-MSCs Promoted the Migration and Invasion of MCF-7 Cells In Vitro and Metastasis In Vivo
The migration and invasion of cancer cells are two requisite steps of metastasis (4). In order to know if UC-MSCs influence the migration and invasion of cancer cells in vitro, the migration and invasion of MCF-7 cells without or with UC-MSCs were tested. Without UC-MSCs in the lower chambers of the Transwell plate, the number of migratory and invasive MCF-7 cells in each field was 60 ± 11 and 54 ± 11, respectively. However, in the presence of UC-MSCs, the number of migratory and invasive MCF-7 cells reached 219 ± 22 and 109 ± 13, respectively (Fig. 1A, B). These results showed that UC-MSCs promoted the migration and invasion of MCF-7 cells in vitro. Since UC-MSCs and MCF-7 cells were separated by a membrane permeable for soluble factors rather than cells, the effect of UC-MSCs was attributed to soluble factors.
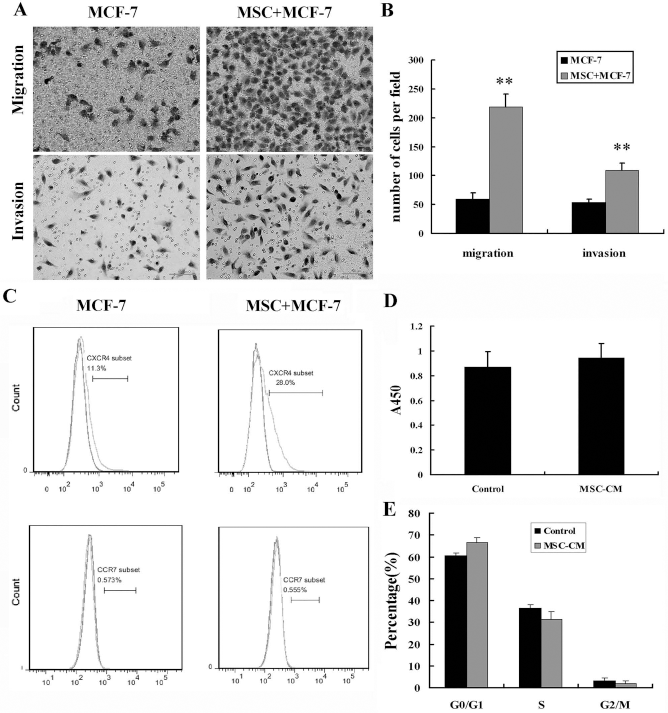
UC-MSCs induced MCF-7 cell migration and invasion in vitro. (A) MCF-7 cells were plated onto inserts with a permeable membrane coated with or without Matrigel and cultured with or without UC-MSCs in the lower chamber. Representative pictures of migratory and invasive cells on the bottom of inserts after hematoxylin and eosin staining. (B) Cell migration and invasion were evaluated by counting six random fields per insert. **p < 0.01 versus MCF-7 group. (C) Representative flow cytometric pictures of chemokine receptors CXCR4 and CCR7 protein expression in MCF-7 cells with or without coculture. (D, E) MCF-7 cells were treated with or without UC-MSCs CM for 48 h; then cell proliferation and cell cycle of MCF-7 cells were assayed.
Previous studies have demonstrated that the migration and invasion of tumor cells depend on chemokine receptors (28); therefore, we examined the expression of chemokine receptors by flow cytometry. The positive proportion of CXCR4 increased from 11.6 ± 0.9% in MCF-7 cells alone to 21.8 ± 1.9% in those cocultured with UC-MSCs (Fig. 1C). The basal expression of CCR7 and CCR5 in MCF-7 cells was less than 1% and was not increased by coculture (Fig. 1C and data not shown). Therefore, CXCR4 might play an important role in the enhancement of migration and invasion. To exclude the possibility that the effect of UC-MSCs on MCF-7 cell proliferation might be responsible for the increased migration and invasion, the proliferation and cell cycle of MCF-7 cells were determined. The results indicated that UC-MSCs had no significant effect on the proliferation and cell cycle of MCF-7 cells (Fig. 1D, E).
We further evaluated if UC-MSCs affected breast cancer metastasis in vivo. MCF-7 cells were injected subcutaneously into mice. When the implanted tumors reached a mean size of 50 mm3, various doses of UC-MSCs or PBS were injected into tumor-bearing mice. The growth and metastasis of tumors were assessed 6 weeks later. The volume and weight of tumor in mice treated with or without UC-MSCs were similar at different time points (Fig. 2A, B), which indicated that UC-MSCs had no significant effect on tumor growth. Sporadic tumor cells were observed in lungs of four tumor-bearing mice treated with 2 × 105 UC-MSCs and five tumor-bearing mice treated with 1 × 106 UC-MSCs (Fig. 2C, D). Moreover, one to two metastatic tumor knobs were observed in lungs of three tumor-bearing mice treated with 1 × 106 UC-MSCs (Fig. 2C, D). However, metastasis was not observed in all tumor-bearing mice treated with PBS or with 4 × 104 UC-MSCs. Tumor cells were not found in other tissues of mice with metastatic tumor.

UC-MSCs induced MCF-7 cell metastasis in vivo. (A, B) MCF-7 cells were injected subcutaneously into mice. When tumors reached the mean size of 50 mm3, different doses of UC-MSCs and PBS were intravenously injected into tumor-bearing mice. The tumor volume and weight of tumor-bearing mice treated without and with UC-MSCs are similar. (C) Representative hematoxylin and eosin-stained sections of lungs of tumor-bearing mice treated with UC-MSCs. Metastatic tumor cells are delineated by an arrow. Scale bar: 100 μm. (D) The frequency of metastatic tumor cells and knobs in each group was calculated.
The Expression and Secretion of IL-8 and IL-6 in MCF-7 Cells Were Induced by Coculture with UC-MSCs
In order to figure out certain cytokines that contributed to increased MCF-7 cell migration, the secretion of IL-8, IL-6, PGE2, VEGF, and SDF-1 by UC-MSCs and MCF-7 cells with and without coculture was determined since these cytokines were reported to be related to cancer growth and metastasis (19). ELISA assays showed that UC-MSCs secreted much higher level of IL-8, IL-6, and PGE2 and lower levels of VEGF when compared with MCF-7 cells. The secretion of these cytokines was not increased in the coculture system (Fig. 3A-D). SDF-1 was not detected in the CM of UC-MSCs, MCF-7 cells, and the coculture system. The mRNA level of IL-8, IL-6, VEGF, and cyclooxygenase-2 (COX-2, prostaglandin synthase) in UC-MSCs was not increased by coculture (Fig. 3E and data not shown). However, compared with MCF-7 cells cultured alone, the mRNA level of IL-8 and IL-6 rather than VEGF and COX-2 in cocultured MCF-7 cells (MCF-7-Co) was much higher (Fig. 3F). After coculture for 48 h, UC-MSCs, and MCF-7 cells were separated and continued to be cultured alone for an additional 48 h. Consistent with the mRNA levels, the secretion of IL-8 and IL-6 was increased in MCF-7-Co CM rather than MSC-Co CM (Fig. 3G, H).

The secretion and expression of cytokines in UC-MSCs and MCF-7 cells with or without coculture. (A-D) The concentration of IL-6, IL-8, PGE2, and VEGF in CM of UC-MSCs alone, MCF-7 cells alone, and coculture system. **p < 0.01 versus MCF-7 group, ##p < 0.01 versus MSC group. (E) The expression of IL-6 and IL-8 in UC-MSCs without and with coculture. (F) The expression of IL-6 and IL-8 in MCF-7 cells without and with coculture. **p < 0.01 versus MCF-7 group. (G) UC-MSCs and MCF-7 cells were cocultured for 48 h, and then they are separated and cultured alone for 48 h. The secretion of IL-6 and IL-8 in CM of UC-MSCs cultured for 48 h and UC-MSCs cultured for 48 h after coculture. (H) The secretion of IL-6 and IL-8 in CM of MCF-7 cells cultured for 48 h and MCF-7 cells cultured for 48 h after coculture. **p < 0.01 versus MCF-7 group.
In summary, UC-MSCs constitutively secreted high levels of IL-8 and IL-6, and their expression and secretion were not altered by coculture. MCF-7 cells constitutively secreted low levels of IL-8 and IL-6. However, the expression and secretion of IL-8 and IL-6 were significantly increased in MCF-7 cells cocultured with UC-MSCs. These results implied that UC-MSCs induced IL-6 and IL-8 production in MCF-7 cells.
IL-8 and IL-6 Secreted by UC-MSCs Induced Autocrine IL-8 and IL-6 in MCF-7 Cells
Recently, it was reported that paracrine transforming growth factor-βl (TGF-β1) by adipose-derived MSCs could induce autocrine of TGF-β in MCF-7 cells (43). We supposed that IL-8 and IL-6 constitutively secreted by UC-MSCs induced the autocrine of IL-8 and IL-6 in MCF-7 cells. To verify our hypothesis, the neutralizing antibodies (NA) of IL-8 and IL-6 were respectively added to the coculture system, and isotype antibodies were used as controls. Then the expression of IL-8 and IL-6 in UC-MSCs and MCF-7 cells was respectively examined. It was shown that both IL-8 NA and IL-6 NA had no effect on IL-8 and IL-6 expression in MSC-Co (Fig. 4A). Nevertheless, either IL-8 NA or IL-6 NA inhibited IL-8 and IL-6 expression in MCF-7-Co (Fig. 4B). On the other hand, IL-8 NA or IL-6 NA had no effect on IL-8 and IL-6 expression in MCF-7 cells cultured alone (data not shown). To further validate our hypothesis, 50 ng/ml exogenous human recombinant IL-8 and 10 ng/ml recombinant IL-6, which mimicked the level of IL-8 and IL-6 secreted by UC-MSCs, were respectively added into the media of MCF-7 cells. More interestingly, the expression of IL-8 and IL-6 in MCF-7 cells was simultaneously upregulated by either IL-8 or IL-6 (Fig. 4C).
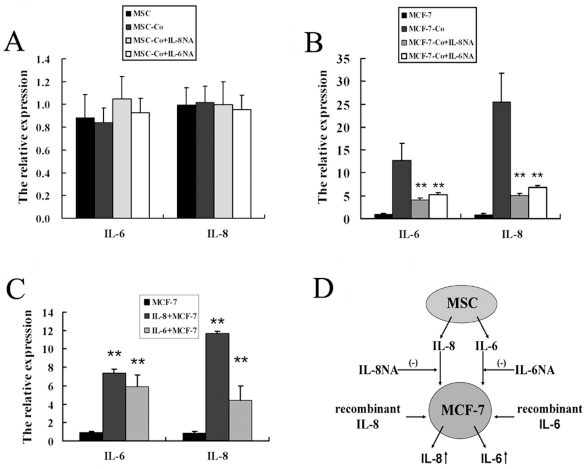
The neutralizing antibody of IL-8 and IL-6 has different effects on IL-8 and IL-6 expression in UC-MSCs and MCF-7 cells. (A) IL-6 and IL-8 expression in UC-MSCs did not change when IL-8 NA and IL-6 NA were added into coculture system. (B) Enhanced expression of IL-6 and IL-8 in MCF-7 cells with coculture was blocked when IL-8 NA and IL-6 NA were added into the coculture system. **p < 0.01 versus MCF-7-Co group. (C) To determine if IL-6 and IL-8 can mimic the effect of UC-MSCs on induction of IL-6 and IL-8, MCF-7 cells were treated with exogenous IL-6 and IL-8. IL-6 and IL-8 expression in MCF-7 cells was also upregulated by exogenous recombinant IL-6 or IL-8. **p < 0.01 versus MCF-7 group. (D) A proposed model of regulation of IL-6 and IL-8 expression in MCF-7 cells by UC-MSCs. It was proposed that paracrine IL-6 and IL-8 by UC-MSCs played a pivotal role in triggering IL-6 and IL-8 expression in MCF-7 cells.
These results suggested that IL-8 and IL-6 secreted by UC-MSCs in coculture system activated the autocrine IL-8 and IL-6 signaling in MCF-7 cells. IL-8 NA or IL-6 NA blocked the activity of IL-8 and IL-6 secreted by UC-MSCs and thereby inhibited the autocrine IL-8 and IL-6 signaling in MCF-7 cells. Exogenous IL-8 and IL-6 mimicked the effect of UC-MSCs on the autocrine IL-8 and IL-6 signaling in MCF-7 cells (Fig. 4D).
Autocrine IL-8 and IL-6 Played an Important Role in MCF-7 Cell Migration
We supposed that the enhanced autocrine IL-8 and IL-6 induced the migration of MCF-7 cells. IL-8 NA and IL-6 NA were also added to the migration assay system, and isotype antibodies were used as controls. IL-8 NA and IL-6 NA, especially IL-8 NA, significantly inhibited the migration of MCF-7 cells induced by UC-MSCs (Fig. 5A). In the presence of UC-MSCs, the number of migratory MCF-7 cells in each field was 219 ± 22. However, the number of migratory MCF-7 cells, respectively, dropped to 75 ± 7 and 126 ± 14 when IL-8 NA and IL-6 NA were added. The protein expression of CXCR4 decreased from 21.8 ± 1.9% to 13.9 ± 2.2% and 16.0 ± 1.4% in the presence of IL-8 NA and IL-6 NA, respectively (Fig. 5B, C). However, in the absence of UC-MSCs, IL-8 NA and IL-6 NA exhibited no effect on migration and CXCR4 expression of MCF-7 cells (data not shown). According to the fact that IL-8 is also a chemokine, we also examined the expression of its receptors CXCR1 and CXCR2. The data showed that the expression of CXCR1 and CXCR2 was increased in MCF-7 cells cocultured with UC-MSCs. However, enhanced CXCR1 and CXCR2 expression was partly reversed in the presence of IL-8 NA or IL-6 NA (Fig. 5C). Like UC-MSCs, IL-8 and IL-6 promoted the migration of MCF-7 cells and the expression of CXCR4 (Fig. 5D-F). Furthermore, IL-8 significantly upregulated the expression of CXCR1 rather than CXCR2 (Fig. 5F), while IL-6 did not upregulate the expression of CXCR1 or CXCR2.

The role of IL-8 and IL-6 in enhanced MCF-7 cell migration induced by UC-MSCs. (A, B) When IL-8 NA and IL-6 NA were added into the migration system, they inhibited enhanced migration and CXCR4 expression of MCF-7 cells cocultured with UC-MSC. (C) The effects of IL-8 NA and IL-6 NA on chemokine receptor expression of MCF-7 cells cocultured with UC-MSC. *p < 0.05 versus MSC + MCF-7 group. (D, E) To determine if IL-6 and IL-8 can mimic the effect of UC-MSCs on induction of IL-6 and IL-8 expression, MCF-7 cells were treated with exogenous IL-6 and IL-8. Recombinant IL-6 and IL-8 induced migration and increased CXCR4 protein expression of MCF-7 cells. (F) The effects of recombinant IL-6 and IL-8 on chemokine receptor expression of MCF-7 cells. **p<0.01 versus MCF-7 group, *p<0.05 versus MCF-7 group.
These results suggested that IL-8 and IL-6 secreted by UC-MSCs activated the autocrine IL-8 and IL-6 signaling in MCF-7 cells, which subsequently promoted MCF-7 cell migration and the expression of chemokine receptors.
The Role of EMT in MCF-7 Migration Induced by UC-MSCs
Since EMT was involved in the migration of cancer cells, we also studied if UC-MSCs induced EMT. MCF-7 cells were shown to express E-cadherin at high levels (>90%) and vimentin at low levels (<2%). It was found that the protein expression of E-cadherin and vimentin did not vary in MCF-7 cells either cocultured with UC-MSCs or stimulated with IL-8 and IL-6 for 48 h (Fig. 6A). Consistently, the mRNA expression of EMT markers, E-cadherin, N-cadherin, vimentin, Snail, and Twist did not alter either (Fig. 6B). To further investigate if EMT was induced when the time of stimulation was prolonged, MCF-7 cells were stimulated by UC-MSCs CM or IL-8 and IL-6 for 7 days. Morphologic alteration was observed in MCF-7 cells stimulated by UC-MSCs CM rather than IL-8 and IL-6 (data not shown). MCF-7 cells stimulated by UC-MSCs CM for 7 days became elongated spindle shape from cobblestone-like epithelial morphology. However, the protein and mRNA expression of EMT markers remain unchanged. Therefore, it was unlikely that UC-MSCs promoted MCF-7 cell migration by induction of EMT.
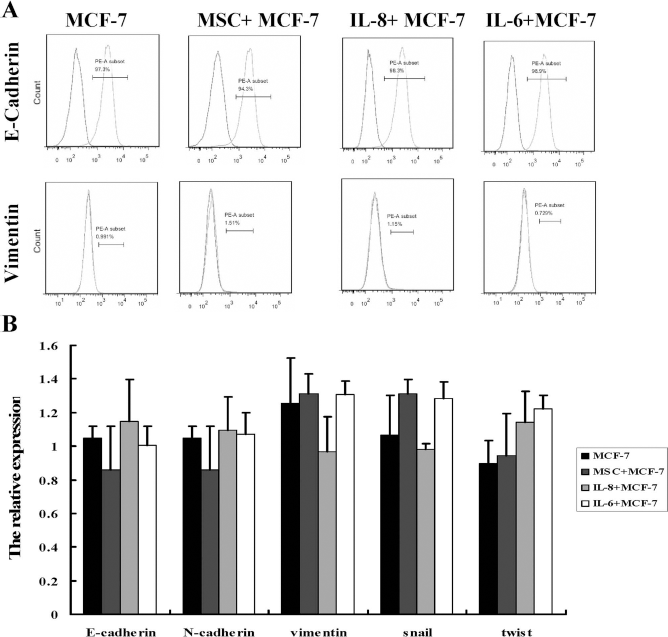
The effects of UC-MSCs, IL-6, and IL-8 on EMT of MCF-7 cells. (A) E-cadherin is a marker of epithelial cell, and vimentin is a marker of mesenchymal cells. When EMT is induced, the expression of E-cadherin is decreased. However, the expression of vimentin is increased. UC-MSCs, IL-6, and IL-8 had no significant effect on E-cadherin and vimentin protein expression in MCF-7 cells. (B) UC-MSCs, IL-6, and IL-8 had no significant effect on mRNA expression of EMT markers in MCF-7 cells.
The Role of CD44+/CD24- Cells in MCF-7 Cell Migration Induced by UC-MSCs
It was reported that CSCs were rich in CD44+/CD24- breast cancer cells and associated with enhanced invasion (1,33). Hence, we investigated the proportion of CD44+CD24- cells in MCF-7 cells. The baseline proportion of CD44+/CD24- cells in MCF-7 control cells was 0.87 ± 0.52% (Fig. 7A), which significantly increased upon induction by UC-MSCs, IL-8, and IL-6 (Fig. 7A). The proportion of CD44+/CD24- cells rose to 13.28 ± 2.47% in MCF-7 cells cocultured with UC-MSCs. The mammosphere culture system has now been used to identify and enrich putative stem cells in breast cancer cells (6). When MCF-7 control cells were cultured in mammosphere culture system, the majority of cells suspended in the medium, and few cells formed small spheres. However, most of MCF-7 cells formed spheres after being cocultured with UC-MSCs or stimulated with IL-8 and IL-6. More importantly, many more mammospheres were formed, and their sizes were significantly larger (Fig. 7B).

The effects of UC-MSCs, IL-6, and IL-8 on the percentage of CD44+CD24- cells and mammosphere-forming ability of MCF-7 cells. (A) Cancer stem cells are rich in CD44+CD24- breast cancer cells. The percentage of CD44+CD24- cells of MCF-7 cells was increased by coculture with UC-MSCs and stimulated with IL-6 and IL-8. (B) The mammosphere-forming ability of MCF-7 cells was promoted by coculture with UC-MSCs and stimulated with IL-6 and IL-8. (C, D) MCF-7 cells cocultured with UC-MSCs were sorted into two subpopulations: CD44+CD24- and CD44+/CD24+ cells. The comparison of mammosphere-forming and migratory abilities of CD44+CD24- cells and CD44+CD24+ cells. (E, F) The comparison of IL-6, IL-8, chemokine receptor expression of CD44+CD24- cells and CD44+CD24+ cells. **p<0.01 versus CD44+CD24+ cells, *p<0.05 versus CD44+CD24+ cells.
In order to know the biological characteristics of these CD44+/CD24- cells, MCF-7 cells cocultured with UC-MSCs were sorted, and the migratory and mammosphere-forming abilities of CD44+/CD24- and CD44+/CD24+ cells were compared. Despite that both CD44+/CD24- and CD44+/CD24+ cells formed mammospheres, CD44+/CD24- cells formed more spheres with larger sizes, compared with CD44+/CD24+ cells (Fig. 7C). More importantly, it was shown that CD44+/CD24- cells had higher migratory ability than CD44+/CD24+ cells (Fig. 7D). The expression of IL-6, IL-8, CXCR1, and CXCR4 in CD44+/CD24- cells was also higher than that of CD44+/CD24+ cells (Fig. 7E, F). These results suggested that CD44+/CD24- cells were involved in MCF-7 cell migration. Previous reports showed that CSCs expressed the markers of EMT and embryonic stem cells. However, we found that the expression of markers of EMT and embryonic stem cells was similar between CD44+/CD24+ cells and CD44+/CD24- cells (data not shown).
Discussion
UC-MSCs are attractive tools for the targeted therapy of cancer (44). However, we demonstrated that UC-MSCs promoted breast cancer metastasis in the present study. We further documented that the effect of UC-MSCs on breast cancer was mediated by secreting IL-8 and IL-6. It was reported that LoVo cells induced MSCs to secrete PGE2 by secreting IL-1 (21). Discrepant with this result, we found that the secretion of PGE2, IL-8, IL-6, and VEGF did not increase in UC-MSCs cocultured with MCF-7 cells. The reason for this inconsistency might be attributed to different types of cancer cells. Unlike LoVo cells, MCF-7 cells were reported not to express IL-1 (34).
It was shown that cytokines, such as IL-8 and IL-6, could be involved in the aggressiveness of breast cancer (8). Therefore, the increased expression of IL-8 and IL-6 in cocultured MCF-7 cells might be responsible for the enhanced migration and invasion. More interestingly, exogenous recombinant IL-8 and IL-6 promoted the migration of MCF-7 cells and simultaneously upregulated the expression of IL-8 and IL-6. A previous study reported that IL-6 upregulated its own expression in MCF-7 cells (32). We further revealed that the positive feedback loop of IL-8 expression existed in MCF-7 cells and played an important role in MCF-7 cell migration. We also showed that the expression of CXCR1 increased in MCF-7 cells cocultured with UC-MSCs. This result further confirmed the role of IL-8 signaling in the migration of MCF-7 cells. Although SDF-1 was not detected in the coculture system, CXCR4 was expressed by MCF-7 cells, and its expression was increased by coculture. It was reported that CXCR4 was a key receptor in the cross talk between cancer cells and their microenvironment (5,31). Our results indicated that CXCR4 might also be an important receptor for MCF-7 cell migration.
Previous studies demonstrated that induction of EMT led to induction of CD44+/CD24- cells, which expressed EMT markers at high levels (26,33). However, EMT was not shown in MCF-7 cells cocultured with UC-MSCs in the present study. We also found that the CD44+/CD24- MCF-7 cells did not express EMT markers. Similar to our results, El-Haibi et al. showed that lysyl oxidase (LOX) was essential for MSC-induced EMT of breast cancer cells. But LOX did not mediate MSC-induced expansion of cancer stem cells (11). These data argued that the signaling pathways governing the propagation of CSCs might not be identical to those pathways fostering EMT. Although we did not find evidences of EMT in MCF-7 cells cocultured with UC-MSCs, EMT was undoubtedly an important mechanism for breast cancer metastasis.
In the present study, we showed that the paracrine IL-8 and IL-6 by UC-MSCs induced the autocrine IL-8 and IL-6 and CD44+/CD24- cells in MCF-7 cells, finally contributing to enhanced cell migration and metastasis. Recently, it was also reported that BM-MSCs regulated breast CSCs through cytokine loops involving IL-6 and CXC chemokine ligand 7 (CXCL7) (23). Our data suggested that interfering with cytokine secretion such as IL-8 and IL-6 might represent a novel strategy to target cancer metastasis. Estrogen receptor (ER)-positive breast cancer cells were used in this study, which possessed low metastatic capacity and constitutively expressed low levels of IL-8 and IL-6. The effect and mechanism of UC-MSCs on ER-negative breast cancer cells might be different because these cells display higher metastatic capacity and constitutively express high levels of IL-8 and IL-6. Therefore, more studies should be carried out to provide further insights into the effects and mechanism of UC-MSCs on various kinds of cancer cells.
Footnotes
Acknowledgments
This work was supported by the National Basic Research Program of China (2011CB964802, 2013CB966900), National Natural Science Foundation of China (81330015, 81200116), Tianjin Research Program of Application Foundation and Advanced Technology (12JCZDJC25000). The authors would like to thank Mengrou Lu (Department of Pharmaceutical Sciences, University of California, Irvine) for reviewing the manuscript and correcting grammatical errors. The authors declare no conflicts of interest.