
Abstract
In this review, we introduce current developments in induced pluripotent stem cells (iPSCs), site-specific nuclease (SSN)-mediated genome editing tools, and the combined application of these two novel technologies in biomedical research and therapeutic trials. The sustainable pluripotent property of iPSCs in vitro not only provides unlimited cell sources for basic research but also benefits precision medicines for human diseases. In addition, rapidly evolving SSN tools efficiently tailor genetic manipulations for exploring gene functions and can be utilized to correct genetic defects of congenital diseases in the near future. Combining iPSC and SSN technologies will create new reliable human disease models with isogenic backgrounds in vitro and provide new solutions for cell replacement and precise therapies.
Keywords
Induced Pluripotent Stem Cell (iPSC) Technology
In 2006 and 2007, Dr. Takahashi and Dr. Yamanaka overexpressed four pluripotency-related transcriptional factors (octamer-binding transcription factor 4 (Oct4), Kruppel-like factor 4 (Klf4), sex-determining region y box 2 (Sox2), and c-myc) and successfully reversed mouse and human somatic cells back to a pluripotent status. These embryonic stem cell (ESC)-like cells are called induced pluripotent stem cells (iPSCs) 1,2 . iPSCs share similar properties with ESCs, including self-renewal, a normal karyotype, a 3-germlayer cell formation and germline transmission ability 1,2 . These unique advantages of ESC-like properties and personalized fabrication from somatic cells rapidly garnered world-wide attention to this technology. Accumulative research has steered the fundamental improvement of the efficacy of iPSC establishment, including culture conditions, optimal cell sources 2,3 , vector designs 4 –8 , and reprogramming assistance by proteins and small molecules 9 –11 . Notably, Dr. Hou reported the success of iPSC production by chemical induction without the introduction of Yamanaka factors 12 . Currently, iPSCs are widely applied in basic research and have become a reliable in vitro platform for developmental studies, disease modeling and drug screening (Fig. 1).
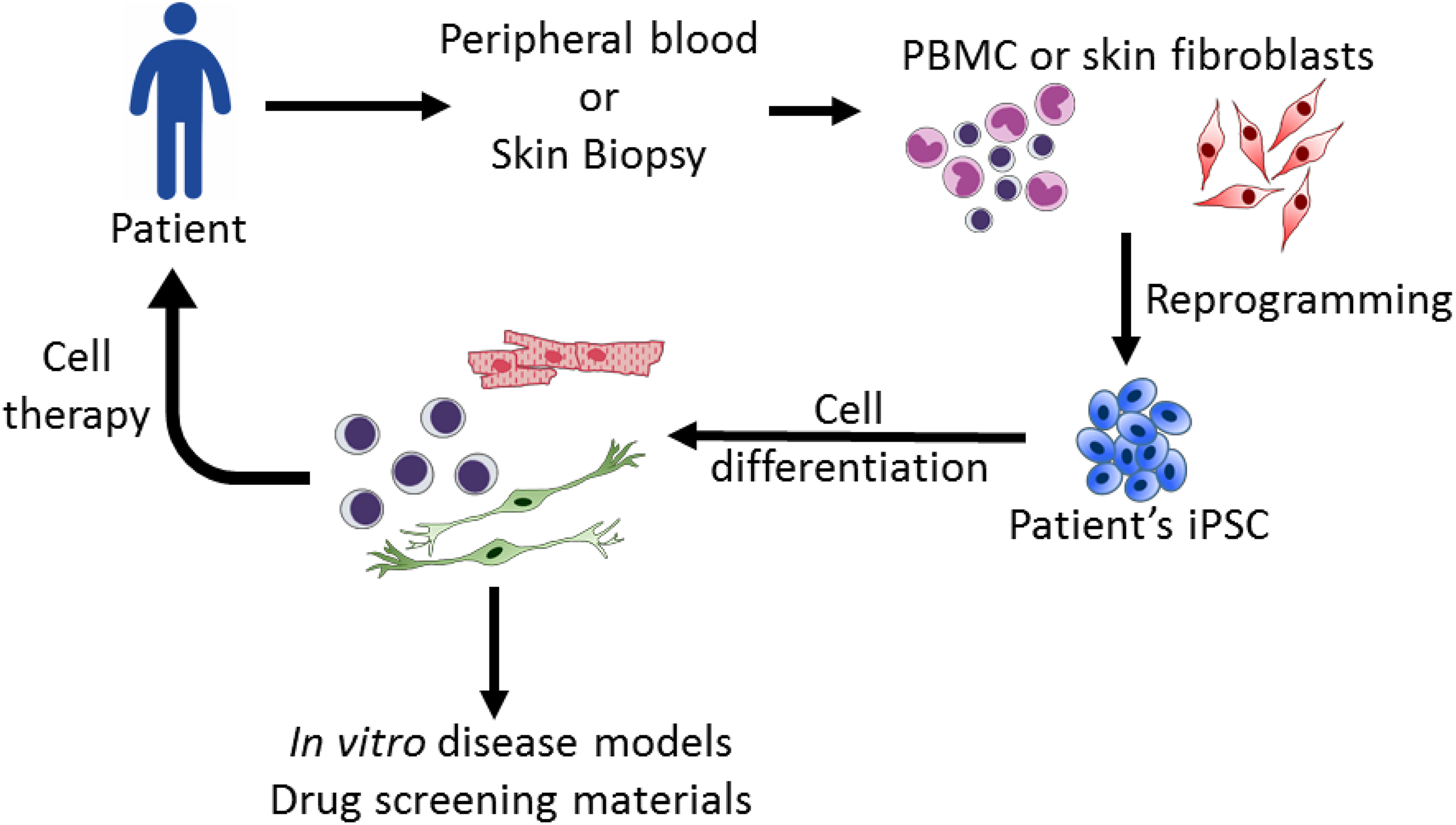
Applications of induced pluripotent stem cell (iPSC) technology. iPSCs derived from patients can be differentiated into specific cell lineages to recapitulate cytopathies for disease studies and potential drug screening. For therapies, iPSC-derived cells can provide materials for transplantation.
Genome modifications in pluripotent stem cells (PSCs) will fundamentally improve the feasibility for researchers to delineate the cell fate, patterning of gene expression, and niche environment regulation at different developmental stages or in 3D organoid architecture. The following text will briefly introduce the genetic editing tools through both random insertion and site-specific modification.
Development of Genome Editing Tools: Genome Modifications Before Site-Specific Nucleases (SSNs)
For genetic modification, there are two major strategies, random insertion and site-specific targeting. For random insertion, lentiviruses 13 and retroviruses 14 are the most commonly used vectors. Other well-known random insertion tools are transposons, including Sleeping Beauty 15 , piggyBac 16 , and others. Through the help of the transposase protein, DNA fragments surrounded with a terminal repeat sequence can be randomly inserted into a host genome. Different from lentiviruses or retroviruses, the transposon can be excised from the host genome via re-expression of transposase and reverse back to transgene-free cell clones 15,16 .
Foreign DNA fragments can be inserted into the host cell genome for different purposes, like gene-specific reporters and gene overexpression. Despite the convenience of the genetic tools, this approach has several shortcomings. First, the random inserted segments may induce mutagenesis in host cells. In addition, the expression level of random inserted genes may be different from the natural expression level of host cells. In some cases, the inserted genes may be silenced, depending on the insertion sites of chromosomes.
Compared with random insertion strategies, site-specific DNA targeting provides higher stability and accuracy for genetic research. For instance, transcription regulatory elements of most genes are still not clear and restrict the application of transgenic systems to genetic function research. Site-specific DNA targeting can overcome these defects of the transgenic approach and become powerful tools for genetic research and therapies. To implant a foreign DNA segment into a specific position of a chromosome, homologous recombination (HR)-based targeting is the traditional approach. Two homologous arms on the 5’ and 3’ ends of foreign DNA are essential for spontaneous HR 17 . Site-specific HR is widely used in mouse ESCs (mESCs) for generating knock-in/knockout mice 18 . Several genetically modified human PSC (hPSC) lines have also been established for disease models. These strategies have also been used to establish gene-specific reporter hPSCs, such as Oct4 (a pluripotent specific marker) and Oligo2 (a neuroglia specific marker), for cell differentiation research or specific cell lineage purification 19,20 .
Although the HR approach is widely applied to mESCs, this genetic targeting approach is limited in hPSCs. The major challenge is the dissociation-induced cell death of hPSCs. Most hPSCs undergo anoikis and die after cell dissociation due to loss of the cell–cell surface cadherin junction 21,22 . This property not only reduces the DNA transfection or electroporation efficiency, but also influences the efficacy to obtain targeted hPSC lines from a single cell. Recent evidence indicates that hPSC dissociation-induced cell death can be inhibited by adding Y27632, which blocks the activation of Rho/ROCK signaling pathway and sustains the cytoskeleton architecture 21 –23 . In addition, this cell death can also be dramatically attenuated when the hPSCs status is converted to naïve state. Naïve hPSCs share similar features with mESCs, including cell morphologies, germline markers expression and high survival rates in single cell status 24,25 . These properties of naïve hPSCs may benefit the improvement of genome editing efficiency in hPSCs. Moreover, newly-developed feeder-free and defined hPSC culture medium fundamentally accelerate the generation of genetic modified hPSCs by avoiding the disturbance of feeder cells during gene transfer and increasing the homogeneity of expanded cultured cells 26 –28 .
The Development of Genome Editing Tools: Site-Specific Nuclease-Mediated Genome Editing
The efficacy of genetic editing by HR can be enhanced by creating DNA break by SSNs. SSNs can initiate the DNA repair system, including both homology-directed repair (HDR) and non-homologous end joining (NHEJ). The most widely used SSNs for genome editing are zinc-finger nucleases (ZFNs), transcription activator-like effector nucleases (TALENs), and clustered regularly interspaced short palindromic repeat (CRISPR)/CRISPR-associated system (Cas9) 29 (Fig. 2).

Site-specific nuclease (SSN)-based genome editing tools. (A) Zinc-finger nucleases (ZFNs). Two zinc-finger nucleases (ZFNs) cooperate for site-specific recognition with a 3-bp pairing/zinc finger and dimerized FokI to create double-strand breaks (DSBs) on DNA. (B) Transcription activator-like effector-based nucleases (TALENs). Two TALENs cooperate for site-specific recognition with 1-bp pairing/repeat variable di-residues (RVDs) and dimerized FokI to create DSBs on DNA. (C–E) Cas9-based SSN system. sgRNA recognizes specific sites via Watson–Crick pairing and cleavage target DNA (C) with wild-type Cas9 (which makes DSB), (D) Cas9 nickase (double-strand nicking with PAM-out and PAM-in orientations), and (E) FokI-dCas9 (which creates DSBs with dimerized FokI).
The zinc finger domain of transcription factor, a DNA-binding motif that recognize and bind to specific DNA sequences, was artificially conjugated with DNA nuclease as zinc-finger nuclease (ZFN) for human genome targeting in 2005 30 . ZFNs are composed of tandem repeating DNA-binding domain which recognize three bases of nuclear acids. For the balance of DNA specificity and targeting efficiency, 3 to 6 zinc fingers are the most suitable length for genome editing. The most common nuclease of ZFN is the FokI restriction enzyme 31,32 (Fig. 2A). ZFNs are widely used for human genome editing 33 . A clinical trial using ZFNs to modify the chemokine receptor CCR5 for human immunodeficiency virus (HIV) therapy is now in progress 29,34 –37 .
Although ZFN is a powerful tool for genome editing, there are still some limitations for this SSN. First, the optimal couple between the zinc finger domain and the nucleotides is a challenge. It is a tremendous work to tailor proper amino acid sequence of the DNA-binding domain for properly fitting the variables of 3-mer DNA sequence. Second, the pairing efficiency of nucleic acids and zinc fingers may be influenced by up- and downstream nucleic acid sequences and zinc finger compositions. Finally, the low binding specificity may also cause off-target mutations and targeting failure.
Transcription activator-like effector (TALE) nucleases (TALENs) is another common SSN for genetic targeting and contain both DNA-binding domain and nuclease domain. The DNA-binding domain of TALE proteins, derived from the plant bacterium, Xanthomonas, are composed of a series of repeating residues with 33∼35 amino acids. The 12th and 13th amino acids within a TALE residue are variable and play major roles in DNA recognition, called repeat variable di-residues (RVDs). Each RVD can specifically recognize a nucleic acid (Asn-Gly for thymine, His-Asp for cytosine, Asn-Ile for adenine, and Asn-Asn for guanine) 38 –40 . Combinations of continuous designed TALE theoretically can recognize all DNA sequences. After DNA-binding, the FokI nuclease can break DNAs with DSB nicks and induce HDR or NHEJ DNA repair reactions 41 (Fig. 2B).
Compared with ZFNs, TALENs have only four types of RVDs to cover the four nucleotides. This advantage makes it much easier to generate TALEN-targeting clones for gene targeting. To overcome difficulties of assembling continuously repeating residues, many strategies have been developed for TALEN assembly, including the Golden Gate method, Platinum Gate method, and ligation-independent cloning (LIC) 42 –46 .
The Development of Genome Editing Tools: CRISPR/Cas9
CRISPR/Cas9 system, the most common SSN from Streptococcus pyogenes, is first discovered from a microbial adaptive immune system against phage infection 47 (Table 2). There are three components of CRISPR/Cas9-mediated DNA recognition and cleavage: Cas nuclease, CRISPR (cr)RNA and trans-activating crRNA (tracrRNA) 48 –50 .
Cas9 contains two nuclease domains, HNH and RuvC, to make a DSB 48 –50 . For CRISPR/Cas targeting, tracrRNA promotes crRNA maturation and conjugates with processed crRNA to form small-guide (sg)RNA for targeting site recognition. The specificity of the gene targeting depends on the pairing of targeting sequencing (also named as protospacer) with sgRNA and protospace-adjacent motif (PAM), which interact with Cas9 protein 48 –50 (Fig. 2C).
Accumulative evidence indicates that off-targeting effect of CRISPR/Cas9 mainly causes by the high tolerance in the sgRNA sequence (up to five mismatches) and the DSB-induced NHEJ repair. To decrease the off-targeting rate and increase the accuracy of genetic modification, Cas9 nickase is generated by inactivating the RuvC nuclease region of Cas9 (D10A mutant in RuvC) to trigger a DNA single-strand break and predominant repair by the high-fidelity base excision repair pathway. Paired Cas9 nickases with reverse orientation DNA-binding are able to make double-strand nicks (Fig. 2D). This modified Cas9 nickase approach can improve targeting specificity by more than 50–1500-fold compared with wild-type Cas9, and also lowers the off-targeting rate 51 –53 . Truncated sgRNAs may also reduce off-targeting effects of CRISPR/Cas9 54 .Moreover, replacing the Cas9 with FokI, called fCas9, increased targeting specificity more than 140-fold compared with the wild-type Cas9 in human cells (Fig. 2E) 55,56 .
Application of SSNs in Clinical Trials
For gene therapy, the development of efficient genome targeting tools provides possibilities for clinical applications. Corrected cells may be helpful for transplantation therapies. The first application of SSNs in clinical trials was to treat acquired immune deficiency syndrome (AIDS). For preclinical AIDS therapy, researchers applied genome editing tools to generate mutations in HIV co-receptor CCR5 that blocks HIV infection and proliferation on CD4 T-cells 34 –36,57 . The modified T-cells are resistant to HIV infection. Based on the positive results, a phase I study which used ZFNs to modify CCR5 of patient CD4 T-cells for autologous transplantation was initiated in 2009. A total of 12 patients with chronic aviremic HIV infection were enrolled, and 6 of them were treated with CCR5-edited CD4 T cell autologous transplantation. Results showed that HIV RNA levels in one patient (this patient was heterozygous for CCR5 delta 32) became undetectable and decreased in most patients. This trial demonstrated the safety and curative efficacy of this strategy. Notably, replenished CCR5 edited T-cells are required for long-term therapy to overcome the short-term survival of edited T-cells or alternatively using hematopoietic stem cells as the next genetic engineered target for providing long-term T-cells.
Application of hPSCs in Clinical Trials
Clinical trials of PSCs have shown the safety and efficacy of the transplantation of PSC-derived differentiating cells, including the treating for age-related macular degeneration (AMD), type I diabetes, Parkinson’s disease (PD), and myocardial infarction (Table 1) 58 .
List of current human pluripotent stem cell (hPSC)-based clinical trials.

ESC: embryonic stem cell; hPSC: human pluripotent stem cell; iPSC: induced pluripotent stem cells; PSC: pluripotent stem cells.
Comparison of site-specific nuclease (SSN) genome editing tools of zinc-finger nucleases (ZFNs), transcription activator-like effector nucleases (TALENs), and clustered regularly interspaced short palindromic repeat (CRISPR)/CRISPR-associated system 9 (Cas9).

The first clinical trial using human ESCs (hESCs) targeted spinal cord injury, supported by Geron company. This phase I clinical trial was initiated in October 2010. hESC-derived oligodendrocyte progenitor cells (GRNOPC1) were used to treat spinal cord injury patients in the subacute stage 59 . Unexpectedly, Geron halted this program in 2013. However, Asterias re-initiated this clinical trial using the same cell type in 2016 due to the success of using hESCs in AMD.
For type I diabetes mellitus (DM), a phase I/II clinical trial using hESC-derived pancreatic precursor cells 60 was initiated in October 2014 by the company Viacyte. Type I DM is caused by the loss of β-islet cells, and patients need to inject insulin for the rest of their lives. In this trial, researchers used a biocompatible capsule to protect hESC-derived pancreatic precursor cells from immune rejection 61 . Animal studies revealed that these encapsulated precursor cells can sense the blood glucose level and secrete insulin over the long-term without the formation of a teratoma. However, detailed results from human clinical trials are still under investigation.
hESC-derived cardiomyocytes have potential for myocardial infarct therapy. In non-human primate preclinical studies, transplantation of ESC-derived cardiomyocytes improved heart function after a myocardial infarct with no ventricular arrhythmia- related problems 62 . For the phase I clinical trial, hESC-derived cardiac progenitor cells were merged with fibrin to form a patch, which was then engrafted onto the damaged area of the heart. After 3 months, the heart function had improved without arrhythmias being observed 63,64 .
The functional improvements of PSC-derived cells were first validated in the AMD studies. AMD, the most common type of blindness in the elderly, is caused by the progressive degeneration of the retinal pigment epithelium (RPE). Stargardt macular dystrophy (SMD), also caused by RPE degeneration, is a juvenile-onset inherent disease caused by mutations of ABCA4, ELOVL4, and PROM1. The phase I results in AMD and SMD patients showed that no tumorigenesis, immune rejection, and RPE outgrowth were observed after the engraftment of ESC-derived RPE sheets. Notably, long-term observation of these phase I/II trials revealed that transplanted RPE cells were integrated with the layer of photoreceptors and significantly recovered the vision spectrum in recipients 65 –67 .
Clinical trials for AMD using iPSCs were carried out in Japan. The phase I trial began in Japan in September 2014, initiated by the Kyoto University’s Center for iPSC Research and Application, Center for Developmental Biology RIKEN (RIKEN CDB), and Kobe City Medical Center General Hospital. This was the world’s first clinical trial using patient’s own iPSCs for autologous transplantation 68,69 . Long-term observations revealed no tumorigenesis or immune rejection. In addition, transplantation of the derived RPE sheets ameliorated the vision deterioration in the patients. More clinical patients will be recruited for the safety evaluation of iPSC-based cell therapy.
In the above-mentioned clinical trials, most applied hESCs were allogenic. Allogeneic transplantations can only be applied in some immune privilege sites, like the eyes and spinal cord. Some special strategies need to be considered to avoid immune rejection 61 . Personalized iPSC lines can resolve immune-related problems. However, generating and characterizing personalized clinical-grade iPSC lines are expensive and time consuming. Allogenic transplantation with human leukocyte antigen (HLA)-matching donors is a standard procedure of organ and bone marrow transplantation. It is an ideal strategy to establish and bank a few HLA-homozygous super-donor iPSC lines for clinical applications without the drawback of severe immune rejection (Fig. 3).

The source of human pluripotent stem cells (hPSCs) for clinical therapies. (A) Currently, most clinical trials use allogeneic human embryonic stem cells (hESCs) for transplantation without human leukocyte antigen (HLA) matching. (B, C) In Japan, two clinical trials used autologous and HLA matching allogeneic-induced pluripotent stem cells (iPSCs) as cell sources. For burden consideration, HLA homogenous iPSC banking is the optimal cell supplement source.
Genome Editing Tools Expand the Application Horizon of iPSC-Based Research and Disease Modeling
PSCs provide an in vitro model to recapitulate human development processes and cell lineage differentiation, especially for the early embryo stage. With genome modifications, researchers can delineate the cell fate, gene expression timing, and niche environment regulation at different developmental stages and 3D organoid architecture. Differentiating PSCs also can serve as a platform to address single or multiple gene functions and their physiological roles in vitro by gene mutations, deletions, or replacement 19,20,50,70 –81 (Fig. 4).
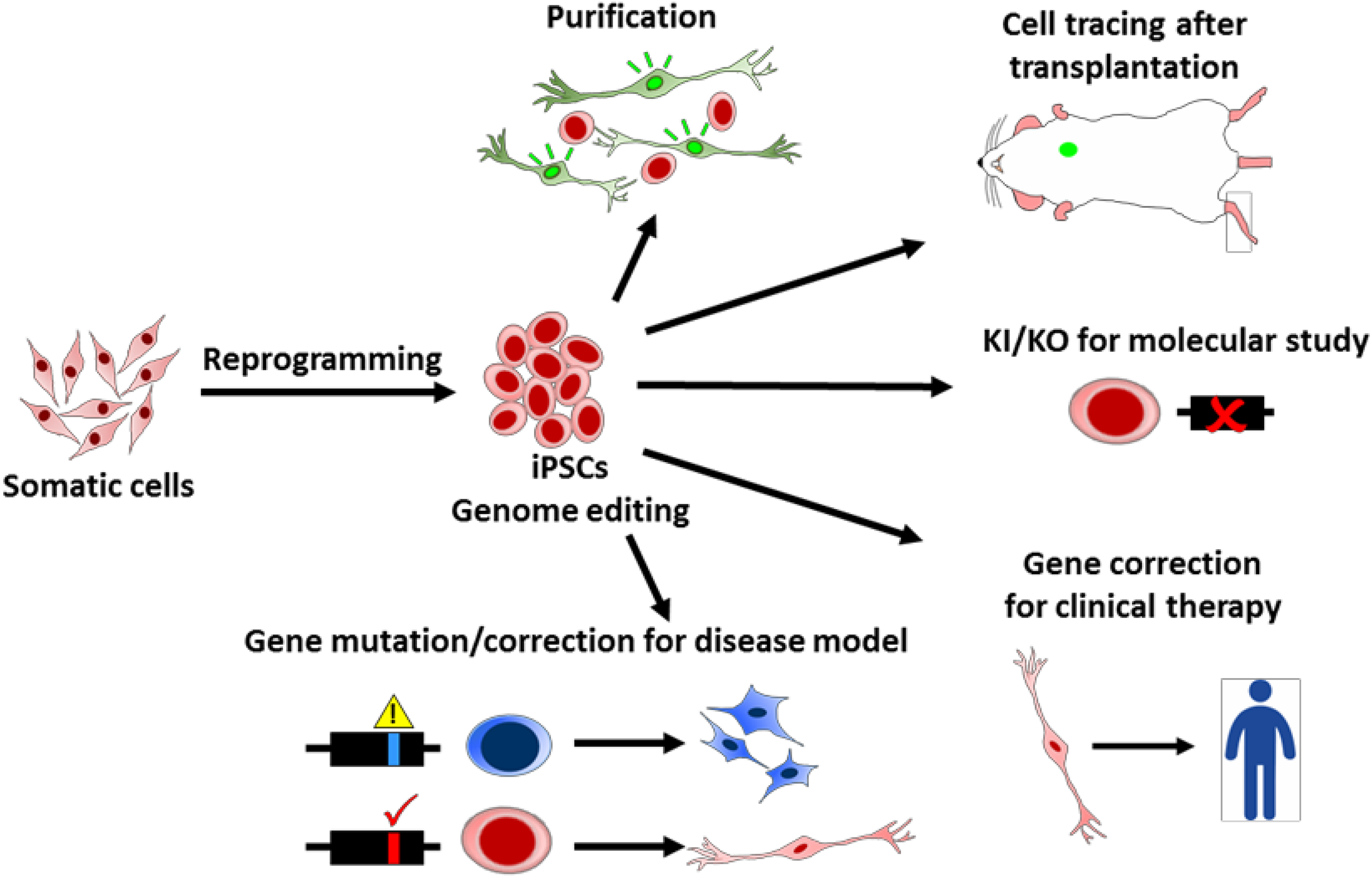
Applications of genome editing tools for induced pluripotent stem cell (iPSC) technology. Genetic technologies provide kinds of application potentials for iPSC research. For example, genome editing tools are used to generate reporter cells for cell purification and tracing, knock-in and knockout (KO) for molecular studies, mutations or corrections for disease modeling, and clinical therapies.
Genome editing tools also provide the feasibility to generate isogenic iPSCs for accurate disease modeling. Specific mutations of disease iPSCs can be artificially fabricated or corrected by genome editing tools to generate isogenic disease iPSCs without the interference of cell resources, random mutations and epigenetic variation during the iPSC establishment. This genotype/phenotype validation by SSN-mediated gene correction or introduction consolidates the roles of candidate genetic loci in patients-derived iPSCs.
SSN-mediated genetic corrections of patient-derived PSCs are applied to Parkinson’s disease 82 , Niemann–Pick type C (NPC) disease 83 , sickle cell disease 84,85 , β-thalassemia 86 –88 , Rett syndrome 89 , cystic fibrosis 90 –92 , and α1-antitrypsin deficiency 93 . CRISPR/Cas9 was also used to correct mutations of MYO15A in hair cell-like cell deficiency 94 , chromosome 7q deletion in myelodysplastic syndromes 95 , and COL7A1 in recessive dystrophic epidermolysis bullosa (RDEB) 96 . For Down syndrome trisomy 21 correction, an inducible XIST transgene was introduced into the DYRK1A locus of chromosome 21 with a ZFN-editing tool. This modification successfully silences the chromosome 21 and forms the chromosome 21 Barr-body to maintain genetic expression balances 97 . Combining of these two technologies also provide us to generate gene corrected iPSCs of large-scale chromosome abnormal diseases like Duchenne muscular dystrophy (DMD) 98,99 . The above examples demonstrate that SSN- mediated genetically corrected hPSCs not only provide research materials but also potentially serve as healthy cell sources for autologous grafting.
Future Direction of SSN/hPSC-Combined Applications
Integrating both fast-evolved genome editing tools and standardized hPSCs fundamentally accelerates the scientific progress on the human developmental studies, disease modeling, specific cell tracing/isolation and clinical cell therapy 19,20,50,70 –81 . The exogenous factors-directed differentiation from PSCs basically follows the developmental principles and reflects early embryonic formation, enabling the hPSC as an ideal model to investigate early human development. Given the high efficacy of CRISPR/Cas9-mediated on single gene knockout, the impact of a disrupted gene can be traced in the hPSCs and their derivatives. Investigating the mutant effects in spherical 3D organoid culture of PSCs can further broaden the affected spectrum of the cell–cell interaction, such as cell migration, niche environment and tissue organization 100 –103 . By the stable expression of inducible Cas9 at the AAVS1 locus, similar to the ROSA26 locus in mice, precisely inactivating the gene expression at desired developmental stages can also be achieved 104 . This constitutive expression of Cas9 endonuclease (iCRISPR) significantly improves the efficacy of multiple gene inactivation and facilitate the exploration of orthologous genes simultaneously in developmental studies. Moreover, the CRISPR/Cas9 system can be further modified to be a gene repressor or converted to be a gene activator module to delineate the regulatory complex and gene function in specific cell types 105,106 . These highly evolved and versatile genetic editing tools will substantially shape the hPSCs as a new platform for early human developmental study.
Accumulative evidence indicates that familial genetic mutations in the patient-derived iPSCs are high penetrable and can faithfully recapitulate the disease phenotype in the derivative target cells from iPSCs 100,107 –122 . The disease cytopathology can be corrected by the TALEN or CRISPR/Cas9-mediated HR 123 –127 . Given the power of CRISPR/Cas9 system, researchers can modify the disease progress and severity by introducing gene deletion or repair using synthetic oligonucleotides in isogenic iPSCs. The interaction network of risk factors can also be delineated by multiplex gene knockout. The combined CRISPR/Cas9 and PSCs also provide a valuable system to investigate the accumulation effects of common but low-risk genetic factors for the progress of non-familial and idiopathic diseases, especially for the neurodegeneration, diabetes, atherosclerosis and cancers. Recently, whole-genome gRNA libraries (genome-wide CRISPR knockout screen [GeCKO]) have been established to dissect gene function in several cancer lines and hPSCs 128,129 . These innovative approaches will continuously provide fundamental and comprehensive understanding for the etiology of multiple-hit idiopathic diseases, paving the way for an optimized treatment strategy for patients.
Fabricating PSCs with genetic editing tools can extend the application of precision medicine by introducing specific tag on the cells and correcting gene mutation in PSC-derived cells. Expressing single or multiple specific reporter proteins under cell specific promoters can assist the cell tracing and cell sorting for further cell analysis and transplantation 19,20,70 –75 . Currently the clinically applied PSC-derived RPE sheet for AMD is manually isolated without cell sorting. Genetic labeling of PSC-derived cells may be required in the future for enriching desired cell population, such as dopaminergic neuron precursors and hematopoietic stem cells, to facilitate the tracing of cell fate, cell survival rates and tumorigenesis in vivo.
The advantage of ease, cost, efficacy and versatility warrants CRISPR/Cas9 system as a suitable tool for human gene therapy. Several clinical trials based on genome editing and PSC technologies also reveal the possibilities to bring SSN/PSC-combined transplantation therapies to the real world. Not only for genetic disease, SSN/PSC-combined technologies also have potential for HIV and cancer therapies 130 . However, to apply PSCs and genome editing tools for clinical use, there are still some serious challenges on the road ahead that need to be overcome.
Large-scale production of clinical grade, high-purity engineered specific cell lineages is the first challenge for clinical applications. iPSC generation, specific cell lineages differentiation and genome editing are effort and time consuming, also a high cost processes. Especially in current Good Manufacture Practices (cGMP) norm. To streamline the procedure of SSN/PSC-based therapies, cGMP grade HLA iPSC banking is an imperative approach for the next decade. For clinical hPSC preparation, a bioreactor capable of large-scale amplification of high quality hPSCs is needed. Moreover, an efficient differentiation protocol to generate specific cell lineages is another big challenge 131 . For this issue, fluorescence-based cell sorting is an ideal option to purify specific cell lineages. For example, the surface marker, Corin and leucine rich repeats and transmembrane domains 1 (LRTM1) were used for dopaminergic neuron precursor purification 132,133 . As previously descripted, SSN-mediated cell lineage specific reporter can also provide an ideal solution for cell purification.
For clinical therapy, the most concerned issue is safety. The innate tumorigenic property of hPSCs is still a challenge. For clinical applications, how to identify low-tumorigenesis-risk iPSC clones and eliminate undifferentiated iPSCs before transplantation still has room to improve. Besides, the off-target risk of CRISPR/Cas9 is another challenge of safety. The high tolerance of mismatches for target recognition and strong DSB activity can create considerable off-targeting mutation, which dramatically hinders the clinical application on gene therapy. The researches using CRISPR/Cas9 for primary human CD4+ T-cells showed promising CCR5 specific targeting, negligible off-target mutation and HIV resistance 134,135 . However, testing CRISPR/Cas9 for homologous recombination-directed repair in tripronuclear embryos observed high unintended mutations in the genome of zygotes 136 . Although several approaches have developed to reduce the off-targeting drawbacks of CRISPR/Cas9 system, such as aforementioned Cas9-nickase, dCas9-FokI dimers and truncated sgRNA oligomer 54,56 , the clinical-grade gene-editing tools with high fidelity and low undesired mutations have to be developed and validated before the clinical applications in embryo or in PSC-based cell transplantation.
Recently, the discover of naïve-state hPSCs may benefit the differentiation of primordial germ cells (PGCs) 137 . Researchers also applied CRISPR/Cas9 to edit human embryo’s genome 136 . However, as technology advances, some ethical debates will arise. The SSN/PSC-based technologies may not only allow us to apply to transplantation therapies, but also allow us to generate modified germ cells. It is an important ethical concern that needed to be detailed discussed before applying these techniques for human germline modification and affect our next generation.
Conclusion
In conclusion, combining iPSCs and SSN genome targeting tools provides new insights for disease modeling and transplantation therapies. The tumorigenic and off-targeting issues of SSN/PSC-based therapies may be overcome in a near future and SSN/PSC-based therapies may be routinely clinical therapies. The future road map is challenging but hopeful.
Footnotes
Declaration of Conflicting Interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported by Bio-innovation Center and financially supported by Buddhist Tzu Chi General Hospital (grant no. TCRD105-16).