
Abstract
Over a century ago, clinicians started to use the human amniotic membrane for coverage of wounds and burn injuries. To date, literally thousands of different clinical applications exist for this biomaterial almost exclusively in a decellularized or denuded form. Recent reconsiderations for the use of vital human amniotic membrane for clinical applications would take advantage of the versatile cells of embryonic origin including the entirety of their cell organelles. Recently, more and more evidence was found, showing mitochondria to be involved in most fundamental cellular processes, such as differentiation and cell death. In this study, we focused on specific properties of mitochondria of vital human amniotic membrane and characterized bioenergetical parameters of 2 subregions of the human amniotic membrane, the placental and reflected amnion. We found significantly different levels of adenosine triphosphate (ATP) and extracellular reactive oxygen species, concentrations of succinate dehydrogenase, and lactate upon inhibition of ATP synthase in placental and reflected amnion. We also found significantly different rates of mitochondrial respiration in isolated human amniotic epithelial cells and human amniotic mesenchymal stromal cells, according to the subregions. Differences in metabolic activities were inversely related to mitochondrial DNA copy numbers in isolated cells of placental and reflected amnion. Based on significant differences of several key parameters of energy metabolism in 2 subregions of vital amnion, we propose that these metabolic differences of vital placental and reflected amnion could have critical impact on therapeutic applications. Inclusion of region-specific metabolic properties could optimize and fine-tune the clinical application of the human amniotic membrane and improve the outcome significantly.
Keywords
Introduction
Over a century ago, J. S. Davis reported the potential surgical use of the human amniotic membrane (hAM) or amnion as skin coverage material for wounds 1 . Soon thereafter, clinicians started to use the hAM for tissue repair such as wound coverage material after burn injuries 2,3 , followed by applications in reconstructive medicine 4 –6 and ophthalmology 7,8 . Over time, it became apparent that components of the hAM contribute to enhance tissue regeneration on multiple levels.
In clinics, hAM is usually employed in a decellularized or a devitalized form, relying upon the fact that its extracellular matrix provides a supportive microenvironment for tissue regenerative processes. Only in recent years, there has been growing interest toward the application of hAM in its vital form. By taking advantage of vital cells of the hAM, the tissue regenerative capacity of hAM could be exploited to its full extent, which may improve the clinical outcome. Thereby, the application of hAM could be optimized on various levels, executing different strategies tailored specifically for each clinical setting. Since cellular metabolism is involved in all cellular processes, the metabolic properties of the hAM should be investigated on cellular, subcellular, and extracellular levels.
Tissue regeneration requires energy that comes from aerobic (mitochondria) or anaerobic (glycolysis) processes. On a subcellular level, the pivotal role that mitochondria play for a number of tissue regenerative processes has become more and more evident in recent years 9 –11 . The fact that certain diseases such as age-related macular degeneration 12 and retinitis pigmentosa are caused by mitochondrial mutation 13 moved mitochondria also into focus for the treatment of degenerative eye diseases. This puts current concepts of (stem) cell therapy into new perspective, as functional mitochondria seem to be required for regeneration of diseased tissue.
In a previous study, we demonstrated differential mitochondrial respiration in biopsies of 2 amniotic subregions, the placental and reflected amnion 14 . Recent studies demonstrate the impact of specific metabolic pathways, such as mitochondrial energy metabolism and the generation of reactive oxygen species (ROS) on stem cell fate 15,16 . Since the hAM contains cells with stem cell characteristics, we investigated further parameters linked to cellular metabolism in hAM and hAM-derived cells. We hypothesize that if mitochondrial activity and generation of ROS are significantly different in cells obtained from the 2 subregions of the hAM, this could impact the therapeutic and regenerative potential of the hAM in clinical applications.
Therefore, we determined adenosine triphosphate (ATP) concentrations, lactate concentrations, succinate dehydrogenase, ROS, and mitochondrial respiration of vital cells in 2 subregions of the hAM, placental, and reflected amnion. By piecing together all these features, we aim to define and comprehend the state of energy metabolism of cells of vital hAM and their dormant regenerative potential representing a basis for improved clinical transplant quality.
Materials and Methods
Separation of Placental and Reflected Regions of the hAM
Placentae of cesarean sections from patients having signed informed consent were obtained with approval of the local ethics committee. This study adheres to the tenets of the Declaration of Helsinki. All placentae were derived from planned cesarean sections at term. Cesarean sections from premature deliveries and emergency cesarean sections were excluded. Placentae were transported in 500 mL Ringer solution, supplemented with 0.25 µg/mL amphotericin B, 100 µg/mL streptomycin, and 60 µg/mL penicillin G, and transportation of the placentae never exceeded 4 h. Placentae with detached or largely detached amniotic membranes were excluded from the study. The placental and reflected regions of the hAM (Fig. 1) were separated from each other as previously described 14 .

Human amniotic membrane before preparation. Placental amnion (P) covers the placenta, reflected amnion (RA) is located opposite of the placenta.
ATP Measurement
Liquid nitrogen frozen biopsies of hAM (Ø 8 mm) of day 0 were homogenized in Precellys tubes with ceramic beads (Keramik-kit 1.4 mm; Peqlab VWR, Wilmington, DE, USA) in a ball mill (CryoMill MM301; Retsch, Haan, Germany) with 500 µL of Tris-HCl buffer (20 mM Tris, 135 mM KCl, pH 7.4). Four hundred microliters of boiling 100 mM Tris/4 mM ethylenediaminetetraacetic acid (EDTA) buffer (pH 7.75) were added to 100 µL homogenate, incubated for 2 min at 100 °C, and centrifuged at 1000g for 2 min. ATP was determined by ATP Bioluminescence Assay kit CLS II (Roche, Basel, Switzerland) using luciferase reagent with Luminat LB 9507 (Berthold, Bad Wildbad, Germany).
Lactate Measurement
ATP synthase was inhibited with oligomycin (0.001 mM). Biopsies of hAM (Ø 26 mm) of day 0 were incubated in 2 mL Dulbecco’s modified Eagle’s medium (DMEM) supplemented with 10% fetal calf serum (FCS), 1%
Succinate Dehydrogenase
Liquid nitrogen frozen biopsies of hAM (Ø 8 mm) of day 0 were recorded by Miniscope MS 200 electron paramagnetic resonance (EPR) spectrometer (Magnettech Ltd, Berlin, Germany) at −196 °C (modulation frequency 100 kHz, microwave frequency 9.429 GHz, microwave power 1 mW, modulation amplitude 5G). Intensities of signals were recorded at 3200 ± 1000 G. The magnitude of the peak-to-peak amplitude of g = 1.93 signal (metalloprotein of electron transfer system [ETS], succinate dehydrogenase) 17 was calculated and expressed in arbitrary units (AU).
Extracellular ROS of hAM Biopsies
Biopsies of hAM (Ø 8 mm, 3 biopsies per sample) of day 0 were incubated for 1.5 h at 37 °C, 5% CO2 in 500 µL culture medium with 500 µM cyclic hydroxylamine 1-hydroxy-3-carboxy-2,2,5,5-tetramethylpyrrolidine hydrochloride (CP-H; Noxygen, Elzach, Germany). Subsequently, 30 µL of supernatants were placed in micropipettes (Blaubrand®; Brand GmbH & Co. KG, Wertheim, Germany), and EPR spectra were recorded at room temperature with the MiniScope MS 200 EPR spectrometer (Magnettech Ltd). The general settings were as follows: modulation frequency 100 kHz, microwave frequency 9.425 GHz, microwave power 8.3 mW, modulation amplitude 4 G, and gain 200. The magnitude of the peak-to-peak amplitude of oxidized CP-H (3-CP) signals was calculated and expressed in AU.
Measurement of ROS in Isolated Cells
Freshly isolated human amniotic epithelial cells (hAECs) and human amniotic mesenchymal stromal cells (hAMSCs) were seeded in duplicates into 12-well plates (Corning Incorporated, Midland, NC, USA) at a density of 500,000 cells/well in DMEM, supplemented with 10% FCS, 2 mM
Nicotinamide Adenine Dinucleotide Phosphate Oxidase
To determine the contribution of nicotinamide adenine dinucleotide phosphate (NADPH) oxidase to cellular ROS levels, we detected nonmitochondrial oxygen consumption using high-resolution respirometer Oxygraph-2 k (Oroboros Instruments, Innsbruck, Austria) with or without NADPH oxidase inhibitor diphenyleneiodonium (0.1 mM). Mitochondrial oxygen consumption was inhibited by the addition of 20 µM myxothiazol to fresh biopsies (14 biopsies per measurement).
Isolation of Human Amniotic Epithelial Cells
The isolation of hAECs was performed on the day of the cesarean section as previously described
18
. Briefly, placental and reflected hAM were cut into 2 × 2 cm pieces and incubated in 0.05% trypsin/EDTA solution and shaken at 37 °C for 20 min. Digestion was stopped with cold PBS, undigested pieces of hAM were collected and digested again for 20 min. The procedure was repeated once again. The digested cell suspensions were filtered through 100 µm cell strainers and centrifuged at 4 °C for 9 min at 400g. The cells were resuspended in DMEM supplemented with 10% FCS, 2 mM
Isolation of Human Amniotic Mesenchymal Stromal Cells
The isolation of hAMSCs was performed on the day of the cesarean section as previously described
19
. Placental and reflected amnion were cut into 2 × 2 cm pieces and incubated in 1 mg/mL collagenase type I solution and shaken at 37 °C for 2 h. Digestion was stopped with cold PBS, and the cell suspension was filtered through 100 µm cell strainers and centrifuged at 4 °C for 9 min at 400g. The cell pellet was resuspended in DMEM supplemented with 10% FCS, 2 mM
Measurement of Mitochondrial Respiration of Isolated hAECs and hAMSCs
Mitochondrial respiratory parameters were measured with high-resolution respirometer (Oxygraph-2k; Oroboros Instruments). Measurements were performed at day 0. Routine respiration of intact hAECs and hAMSCs was measured by incubating 4 × 106 cells in DMEM and 5 mg/mL fatty acid-free bovine serum albumin (BSA) (Sigma-Aldrich) at pH 7.2 and 37 °C. LEAK respiration was measured by inhibition of ATP synthase with 1 µM oligomycin (Sigma-Aldrich). Maximum ETS capacity was induced by titration of carbonyl cyanide-4-(trifluoromethoxy) phenylhydrazone (Sigma-Aldrich) in steps of 0.5 µM. Oxidative phosphorylation (OXPHOS) was calculated by subtraction of LEAK respiration from routine respiration.
Laser Scanning Microscopy
Isolated hAECs and hAMSCs were seeded on chamber slides with borosilicate bottom plates and incubated for 24 h. Mitochondria were stained with 25 nM MitoTracker Green FM (Molecular Probes, Eugene OR, USA; staining mitochondria) and 50 nM tetramethylrhodamine-methylester (TMRM, Heidelberg, Germany interrogating mitochondrial membrane potential) and analyzed with LSM 510 (Carl Zeiss, Oberkochen, Germany).
Measurement of Mitochondrial DNA Copy Numbers
DNA was extracted from a pellet of 10,000 hAMSCs or hAECs stored at −20 °C using the Tissue and Cell Genomic DNA Purification kit in accordance with the manufacturer’s protocol (GMbiolab Co, Taichung City, Taiwan). Quantitation of the cellular mitochondrial DNA (mtDNA) content was performed by targeting a highly conserved region of the mitochondrial MT-ND1 gene. MtDNA copy numbers were counted using chip-based digital polymerase chain reaction. For detailed information see Supplemental Material.
Statistical Analysis
Statistical analysis was performed with GraphPad Prism, version 5.01 (GraphPad Software, La Jolla, CA, USA), using t tests. In all tests, n (sample size) represents biological replicates (donors). Significance level was set at 0.05 and is indicated as *P < 0.05, **P < 0.01, ***P < 0.001. Wilcoxon-matched pairs test was used to analyze subregional differences in mtDNA copy numbers in hAECs and hAMSCs. Differences in mtDNA copy numbers between hAECs and hAMSCs were tested with the Mann–Whitney test.
Results
Measurements Performed in Biopsies
ATP concentrations higher in placental amnion
We measured ATP concentrations in placental and reflected regions of fresh hAM biopsies and found significantly higher ATP concentrations in placental amnion compared to reflected amnion (Fig. 2A; P < 0.05, n = 4).

ATP and lactate concentrations of fresh hAM. Placental amnion showed higher concentrations of ATP compared to reflected amnion. Each measurement was performed with 3 biopsies (Ø 8 mm; A; n = 4, mean ± standard deviation). Inhibition of ATP synthase with oligomycin stimulated cells of the placental amnion to generate ATP from glycolysis as indicated by elevated levels of lactate. In contrast, reflected amnion showed only very weak response to the inhibition with oligomycin (B). Supernatants of 3 biopsies (Ø 26 mm) were taken for each measurement (B; n = 4, mean ± standard deviation). ATP, adenosine triphosphate; hAM, human amniotic membrane; P, placental amnion; RA, reflected amnion.
Responsiveness to inhibition of ATP synthase more pronounced in placental amnion
Inhibition of ATP synthase led to elevated lactate levels in the placental region (P < 0.01) of the hAM, reflecting a switch to increased glycolysis. In contrast, reflected amnion did not perform this metabolic shift (Fig. 2B; n = 4).
Succinate dehydrogenase levels higher in placental amnion
Measurement of the iron–sulfur center of succinate dehydrogenase (g = 1.93; Fig. 3A) with EPR showed higher levels in placental amnion compared to reflected amnion (Fig. 3B; P < 0.05, n = 4), although it may also reflect the changes in intracellular redox potential 20 .

EPR spectra and quantification of the metalloprotein succinate dehydrogenase. EPR spectrum of hAM recorded at liquid nitrogen temperature (−196 °C). Insert represents typical tracing of the signal at g = 1.93, predominantly coming from succinate dehydrogenase (A). Quantification of EPR spectra showed higher levels of metalloprotein succinate dehydrogenase in fresh placental amnion (B; n = 4, mean ± standard deviation). Each measurement was performed with 3 biopsies (Ø 8 mm). AU, arbitrary units; EPR, electron paramagnetic resonance; hAM, human amniotic membrane; P, placental amnion; RA, reflected amnion.
Extracellular ROS levels higher in placental amnion
Levels of extracellular ROS in the supernatant of the placental region of hAM biopsies were higher compared to the reflected region (Fig. 4A; P = 0.064, n = 4). Further analysis of isolated hAMSCs showed differences in intra- and extracellular ROS levels in the amniotic subregions (Supplemental Fig. 1A), whereas isolated hAECs did not show any subregional differences (Supplemental Fig. 1B).

Levels of ROS and activity of NADPH oxidase of fresh hAM. Intensities of oxidized CP-H (3-CP) signals showed lower levels of extracellular ROS in reflected amnion compared to placental amnion (A; n = 4, mean ± standard deviation). Indirect measurement of NADPH oxidase activity by addition of the NADPH oxidase inhibitor diphenyleneiodonium revealed significantly higher NADPH oxidase activity in placental amnion compared to reflected amnion (B; n = 4, mean ± standard deviation). AU, arbitrary units; 3-CP, 3-carboxy- 2,2,5,5-tetramethylpyrrolidine; hAM, human amniotic membrane; NADPH, nicotinamide adenine dinucleotide phosphate; P, placental amnion; RA, reflected amnion; ROS, reactive oxygen species.
NADPH oxidase higher in biopsies of placental amnion
Calculation of the oxygen consumption of NADPH oxidase showed a significantly higher activity of NADPH oxidase in placental amnion compared to reflected amnion (Fig. 4B; P < 0.05, n = 4).
Measurements Performed in Isolated Cells
Mitochondrial respiration higher in isolated cells of placental amnion
Placental hAECs showed significantly higher routine respiration (P < 0.01), maximum ETS capacity (P < 0.01), and OXPHOS (P < 0.001) compared to reflected hAECs (Fig. 5A; n = 5). Isolated placental hAMSCs showed significantly higher routine respiration (P < 0.01), LEAK (P < 0.05), maximum ETS capacity (P < 0.01), and OXPHOS (P < 0.05) compared to reflected hAMSCs (Fig. 5B; n = 5).
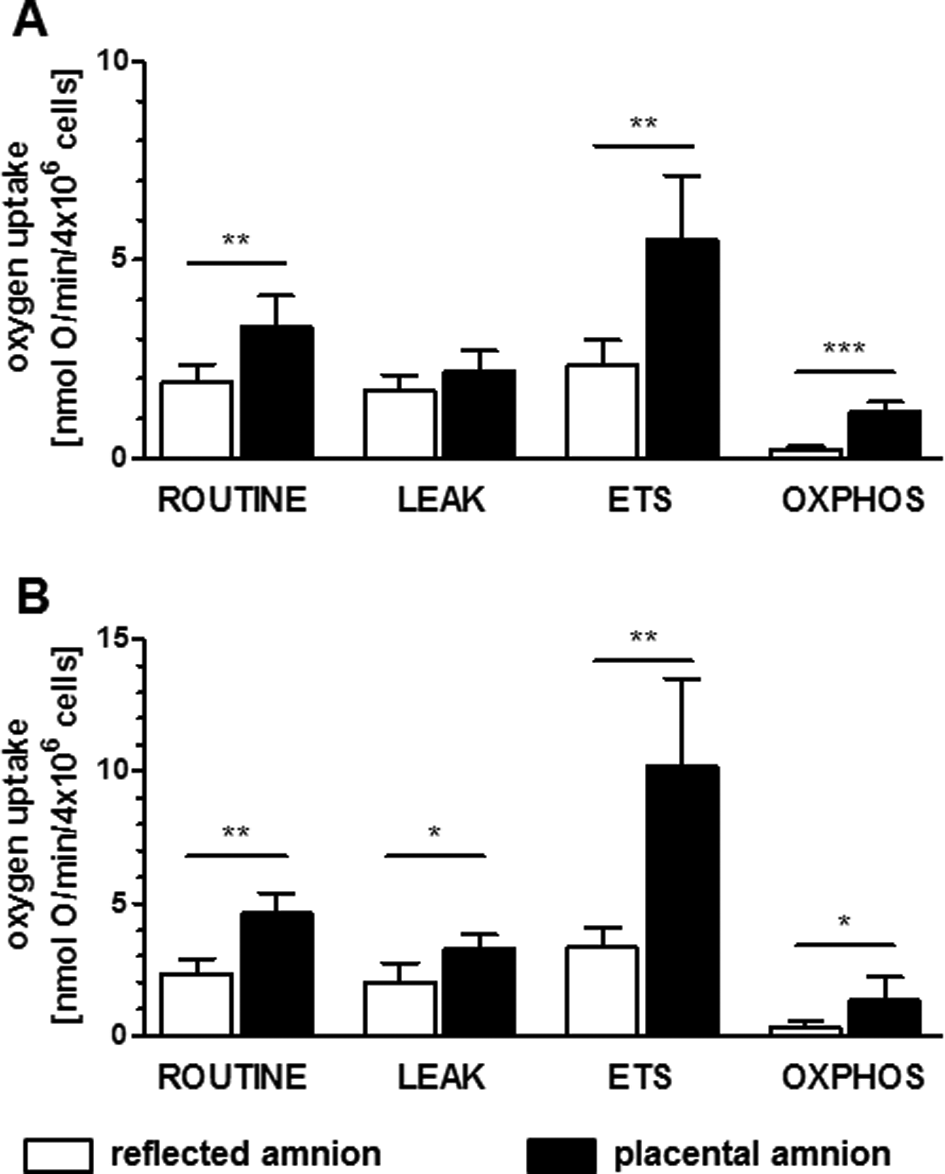
Mitochondrial respiration of isolated human amniotic cells. Isolated hAEC (A) and hAMSC (B) were measured with Oxygraph-2 k. Comparison of placental and reflected amnion revealed routine respiration, maximum ETS capacity and OXPHOS of hAECs (A) and hAMSC (B) were significantly higher in placental amnion. LEAK state was significantly higher in hAMSCs but not in hAMSC. (n = 5, mean ± standard deviation). ETS, electron transfer system; hAEC, human amnion epithelial cells; hAMSC, human amnion mesenchymal stromal cells; OXPHOS, oxidative phosphorylation; P, placental amnion; RA, reflected amnion.
mtDNA copy number higher in isolated cells of reflected amnion
We observed higher mtDNA copy numbers in cells of reflected amnion of both cell types hAECs (Fig. 6A; P = 0.06) and hAMSCs (Fig. 6B; P = 0.06) compared to placental amnion. The comparison of cellular mtDNA copy numbers across cell types showed higher levels of mtDNA in hAECs of reflected amnion (Fig. 6C; P < 0.01) and placental amnion (Fig. 6D; P < 0.01) compared to the respective regions of hAMSC (n = 5).

Mitochondrial DNA copy number in isolated amnion-derived cells. Higher mtDNA copy numbers were found in cells of reflected amnion of both cell types hAECs (A) and hAMSCs (B) compared to placental amnion. MtDNA copy numbers were higher in hAECs in reflected amnion (C) and placental amnion (D) compared to hAMSC, n = 5. Data in C and D are depicted as (min-to-max) box-and-whiskers plots. Quantitation was performed by quantitative PCR (empty symbols) or digital PCR (filled symbol). hAECs, human amniotic epithelial cells; hAMSCs, human amniotic mesenchymal stromal cells; mtDNA, mitochondrial DNA.
Isolated hAECs of placental amnion are more heterogeneous than of reflected amnion
Placental hAECs were more heterogeneous showing higher mitochondrial membrane potential (TMRM, red) than reflected hAECs. Isolated hAECs of placental amnion also showed a higher frequency of adherence to the collagen-coated surface of borosilicate chamber slides (Fig. 7, representative image of n = 3). Differences in heterogeneity and attachment behavior were not observed between hAMSCs of the 2 subregions (Fig. 8, representative image of n = 3).
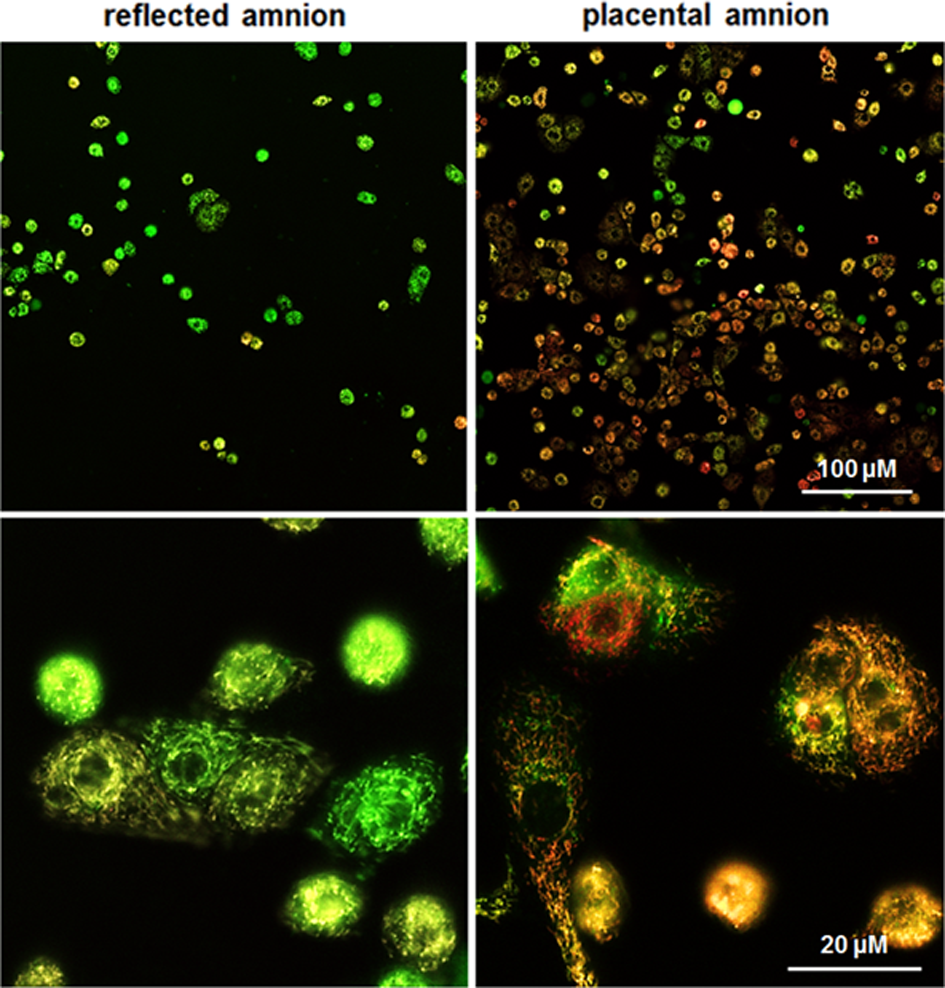
Isolated hAEC. MitoTracker Green FM (green, staining mitochondria) and TMRM (red, interrogating mitochondrial membrane potential) were used for laser scanning confocal microscopy. The population of hAECs of placental amnion appears more heterogeneous than that of reflected amnion. Cells of placental amnion adhere more readily to collagen-coated surface. Moreover, hAECs of placental amnion seem to have higher membrane potentials indicated by more mitochondria-stained positive for TMRM. Images were taken 1 d after cell isolation and seeding. Representative figures of merged MitoTracker Green and TMRM staining. hAEC, human amniotic epithelial cells; TMRM, tetramethylrhodamine-methylester.
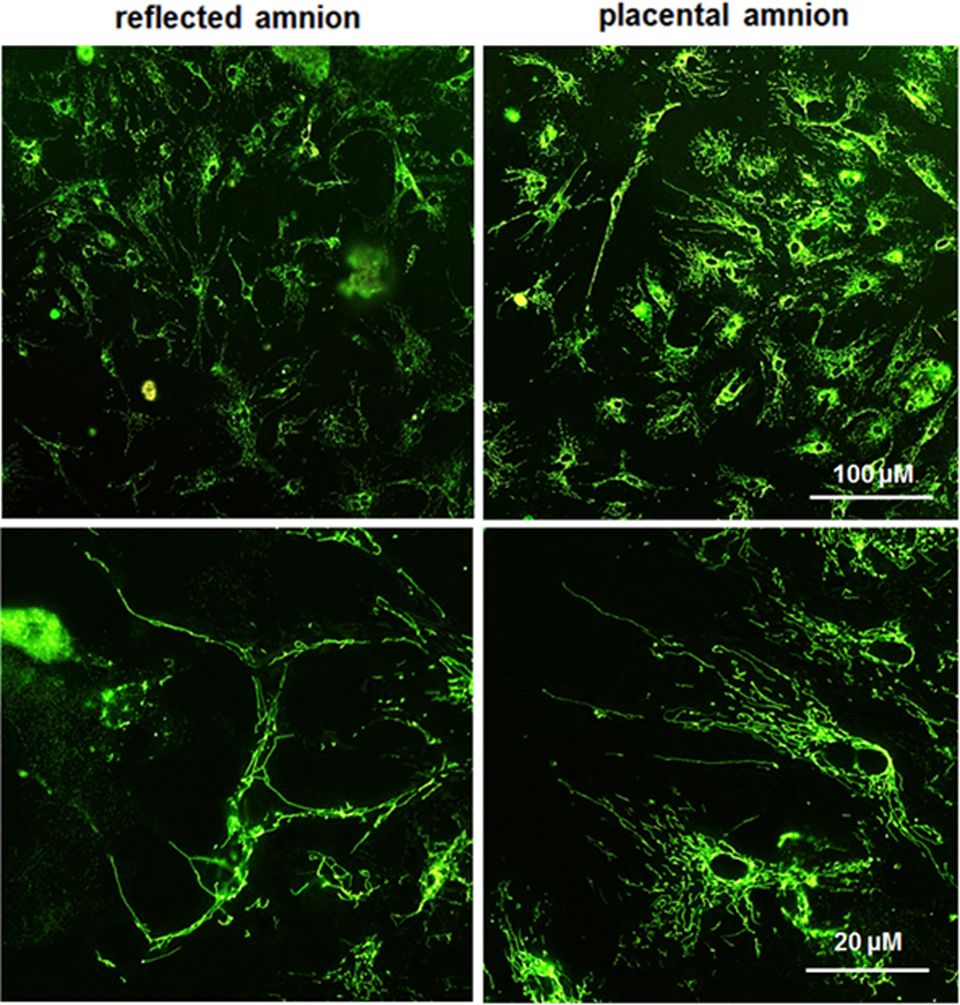
Isolated hAMSC. MitoTracker Green FM (green, staining mitochondria) and TMRM (red, interrogating mitochondrial membrane potential) were used for laser scanning confocal microscopy. Morphologically, hAMSCs do not appear to be different in the respective amniotic regions. Placental and reflected amnion show highly elongated mitochondria 1 d after cell isolation and seeding. Representative figures of merged MitoTracker Green and TMRM staining. hAMSCs, human amniotic mesenchymal stromal cells; TMRM, tetramethylrhodamine-methylester.
Discussion
The hAM is a readily accessible biomaterial. Its application techniques have undergone advancements on different levels, depending on current understandings of tissue repair processes and regulatory standards (reviewed by Silini et al. 21 ). Effective strategies for regenerative medicine require profound examination of the hAM. This also includes cell organelles such as mitochondria and their coplayers. Furthermore, increasing evidence has established the central role of mitochondrial energy metabolism on cell fate. OXPHOS generates significantly more ATP from glucose than glycolysis, and yet, certain cells rely on glycolysis rather than OXPHOS for ATP generation, in particular, certain types of stem cells and cancer cells 22 .
Recently, we have shown differential mitochondrial activities in 2 subregions of the hAM 14 . In the present study, we aimed to characterize more specifically key parameters connected to mitochondria in 2 subregions of the hAM.
We found ATP concentrations to be significantly higher in placental amnion compared to reflected amnion. These data corroborate the results of mitochondrial activity as previously shown 14 . In general, cellular ATP levels reflect the metabolic state of a vital tissue. However, since ATP is consumed by a range of cellular processes, it should be taken into account that ATP levels do not necessarily exclusively correlate with mitochondrial activity 23,24 .
Inhibiting the ATP generation of the ETS can stimulate the cell to detour ATP production via glycolysis. We found cells of the placental region of hAM to respond to the inhibition of ATP synthase by increased glycolysis rates as indicated by elevated lactate levels. In contrast, cells of the reflected amnion only showed a weak response to the inhibition of ATP synthase. One explanation could be that cells of reflected amnion produce ATP mostly via glycolysis rather than via OXPHOS, which would be in line with lower levels of ATP and the lower mitochondrial activity of these cells. Accordingly, these cells would not need to compensate for an inhibition of the ATP synthase. Of note, it has previously been shown that human embryonic stem cells generate ATP via glycolysis 22,25 . In this context, it should be mentioned that the hAM emerges at a very early developmental stage, and at term, many of the hAECs 18,26 and hAMSCs 27 still express markers of pluripotency. Another explanation could be that cells of reflected amnion have a lower responsiveness to the inhibition of the ATP synthase. This could in consequence mean that these cells have a lower capacity to cope with external stress factors. Particularly at the site of a therapeutic transplantation, it is crucial that cells have the ability to adopt their energy metabolism according to the energy “supply and demand”. A site of injury usually represents a harsh microenvironment with a high degree of inflammatory factors, immune cells, and hypoxia.
Recently, a novel mechanism has been found that indicates complex II of the mitochondrial ETS, or succinate dehydrogenase, as an important factor that allows the cell to react to increased energy demand 28 . This is insofar interesting as complex II, a component of the mitochondrial ETS, is also an enzyme of the Krebs cycle. We found higher levels of succinate dehydrogenase in placental amnion compared to reflected amnion. This is in line with the results of our previous study of mitochondrial differences in placental and reflected amnion measured with high-resolution respirometry 14 . Furthermore, this supports the assumption that the placental region of the hAM might have a higher capacity to cope with changing energy demand.
Mitochondria are not only an important metabolic interface, cellular respiration also produces ROS, which are important cellular signaling molecules (reviewed by Serrander et al. and Karlsson et al. 29,30 ). In this study, we found that placental amnion is characterized by higher levels of extracellular ROS. This is particularly interesting, as we have previously shown lower intracellular ROS levels in placental amnion 14 . Interestingly, the same differences were also found in isolated hAMSCs but not in isolated hAECs. In this context, it should be taken into account that ROS can also be produced by NADPH oxidase. Depending on the involved receptor, NADPH oxidase–derived ROS can be released intracellularly 31 –33 as well as extracellularly 32 –34 . The assessment of the NADPH oxidase activity showed a significantly higher contribution of NADPH oxidase activity in placental amnion compared to reflected amnion. Since NADPH oxidase–derived ROS evidently play an important role in the immune response 35,36 , we speculate that the determined NADPH oxidase activity in hAM may contribute to its reported antimicrobial effect 37 .
Moreover, ROS seem to be critical factors for the regulation of cell fate 15,16 , and the signaling function of ROS interacts with many cellular processes 38,39 . Considering these fundamental roles of ROS, the subregional differences in extracellular and intracellular ROS levels suggest different cellular functions in the 2 amniotic subregions.
Measurement of mitochondrial activity of isolated cells of the hAM provided information on how much each cell type, hAECs and hAMSCs, contribute to the differential mitochondrial activity of amnion biopsies. Independent of the cell type, respiratory activity was significantly higher in amniotic cells of the placental region in almost all respiratory states. Of note, OXPHOS state, reflecting mitochondrial ATP synthesis, is low compared to the other respiratory states in both cell types. This is in line with recent studies showing that ATP in (pluripotent) stem cells is mainly generated by glycolysis, whereas OXPHOS activity is low 22,25 .
It seems reasonable to speculate that the differential mitochondrial respiratory activity could be connected to the anatomic location. In addition, results of mtDNA analysis suggest that differences in mitochondrial activity are a possible response to the physiological requirements of the specific region within the amnion. If placental and reflected regions have different tissue oxygen levels, it is likely that cells of these locations adapt their mitochondrial activity accordingly.
Finally, visualizing the mitochondria of isolated cells with 2 different mitochondrial stains showed that hAECs of placental amnion are more heterogeneous compared to hAECs isolated from reflected amnion. This difference was not visible with hAMSCs of the 2 subregions. This could be attributed to the fact that hAECs are exposed directly to the amniotic fluid as opposed to hAMSCs. This exposure may be the explanation for the heterogeneity of hAECs, enabling the cells to react to any changes in their immediate environment.
Taken together, these mitochondria-linked processes contribute to the current metabolic state of a cell, and thus to the potential to adapt to a given microenvironment. Gaining knowledge on these specific interplays is indispensable for deeper insights into the capacity of a cell to adapt to a diseased environment in terms of survival, proliferation, and regenerative actions such as differentiation. At this point, it cannot be stated how and by which mechanisms these assessed regional and cellular properties of the hAM may impact tissue regeneration in clinical applications. Further efforts are required to understand the underlying mechanisms and regulations of tissue repair, in particular in a hypoxic or inflamed environment. We will focus on these issues in our future studies.
We propose that taking advantage of subregional differences of vital hAM could further improve efficacy in tissue regeneration. We are aware of the fact that using vital hAM implies further standardization processes concerning donor variability and safety for patients and requires critical evaluation of the pathological situation. Regarding donor variability, one should take into account that human tissue is prone to variation, and larger cohorts may further confirm our results. Furthermore, although all placentae were obtained at term, there still might be variations in gestational weeks, which could also have impacted the results. Nevertheless, exploring the metabolic landscape of vital hAM led us to the conclusion that vital hAM should be considered an attractive alternative to current decellularized applications for improving tissue regeneration.
Supplemental Material
Supplemental Material - Cellular and Site-Specific Mitochondrial Characterization of Vital Human Amniotic Membrane
Supplemental Material for Cellular and Site-Specific Mitochondrial Characterization of Vital Human Amniotic Membrane by Asmita Banerjee, Andrea Lindenmair, Simone Hennerbichler, Philipp Steindorf, Ralf Steinborn, Andrey V. Kozlov, Heinz Redl, Susanne Wolbank, and Adelheid Weidinger in Cell Transplantation
Footnotes
Ethical Approval
Placentae from caesarean sections were obtained with approval of the local ethics committee (No. 200 by the ‘Ethikkommission des Landes Oberösterreich’). This study adheres to the tenets of the Declaration of Helsinki.
Statement of Human and Animal Rights
Specimens of human placentae were obtained from patients having signed an informed consent according to the local ethics committee.
Statement of Informed Consent
Specimens of human placentae were obtained from patients having signed an informed consent according to the local ethics committee.
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) received no financial support for the research, authorship, and/or publication of this article.
Supplemental Material
Supplementary material for this article is available online.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.