
Abstract
A lot of phenolic compounds are widespread in industrial effluents and they are considerable environmental pollutants. Being a compound commercially available, the effect of a bearing-wastewater phenolic compound 3,4-dimethylphenol (3,4-DMP) on Ca2+ homeostasis and its related physiology has not been explored in cultured human kidney cell models. The aim of this study was to explore the effect of 3,4-DMP on [Ca2+]i and viability in HK-2 human proximal renal tubular epithelial cells. In terms of Ca2+ signaling, 3,4-DMP (5–100 μM) induced [Ca2+]i rises only in HK-2 cells and Ca2+ removal reduced the signal by 40%. In Ca2+-containing medium, 3,4-DMP-induced Ca2+ entry was inhibited by 20% by a modulator of store-operated Ca2+ channels (2-APB), and by a PKC activator (PMA) and inhibitor (GF109203X). Moreover, 3,4-DMP-induced Mn2+ influx suggesting of Ca2+ entry. In Ca2+-free medium, inhibition of PLC with U73122 abolished 3,4-DMP-induced [Ca2+]i rises. Furthermore, treatment with the endoplasmic reticulum Ca2+ pump inhibitor thapsigargin abolished 3,4-DMP-evoked [Ca2+]i rises. Conversely, treatment with 3,4-DMP abolished thapsigargin-evoked [Ca2+]i rises. Regarding to cell viability, 3,4-DMP (60–140 μM) killed cells in a concentration-dependent fashion in HK-2 cells. Chelation of cytosolic Ca2+ with BAPTA-AM partially reversed cytotoxicity of 3,4-DMP. Collectively, our data suggest that in HK-2 cells, 3,4-DMP-induced [Ca2+]i rises by evoking Ca2+ entry via PKC-sensitive store-operated Ca2+ entry and PLC-dependent Ca2+ release from the endoplasmic reticulum. 3,4-DMP also caused cytotoxicity that was linked to preceding [Ca2+]i rises. Our findings provide new insight into the cytotoxic effects of 3,4-DMP and the possible mechanisms underlying these effects.
Keywords
Introduction
Phenolic compounds are widespread in industrial effluents originating from many different sources. They are major constituents of wastewater from coal conversion processes, coke ovens, petroleum refineries and petrochemical industries, resin and fiberglass manufacturing and herbicide production. 1 Toxicity is higher in substituted phenols. 2 Among substituted phenols, dimethylphenols are highly reactive in the atmosphere. 3 A few of the effect of dimethylphenols on physiology have been reported. Previous studies have shown that the mechanism and kinetics of the atmospheric oxidative degradation of dimethylphenol isomers were initiated by OH radical. 3 Furthermore, dimethylphenol derivatives were determined in vitro antioxidant and radical scavenging activities. 4 3,4-Dimethylphenol (3,4-DMP), a compound of dimethylphenol derivative, belongs to phenolic compounds of bearing-wastewaters. A previous study has reported that defensive secretions of the carabid beetle Chlaenius cordicollis contained many chemical components that include 3,4-DMP. 5 Moreover, 3,4-DMP and its derivatives acted as potent α(1)-adrenoceptor antagonists in rat anococcygeus muscles. 6 Despite being commercially available, however, the effect of 3,4-DMP on Ca2+ signal transduction in kidney cell models was unclear.
Ca2+ ions play a crucial role as a second messenger in cellular signal transduction. A transient rise in intracellular free Ca2+ level ([Ca2+]i) can regulate many cellular responses including fertilization, apoptosis, gene expression, secretion, contraction, protein folding, etc. 7 Due to its importance, Ca2+ signaling is tightly regulated by diverse molecules in the cell. [Ca2+]i rises could be contributed by Ca2+ entry from extracellular medium and/or Ca2+ release from organelles such as the endoplasmic reticulum. 8 Therefore, it is important to explore the mechanisms of an agent-induced Ca2+ entry and Ca2+ release in order to understand the impact of this agent on physiology of the cells.
Since dimethylphenol derivatives including 3,4-DMP are associated with environmental pollutants and may provide toxic perspective for human, the effect of use of 3,4-DMP on human physiology should be cautioned. The aim of this study was to investigate the effect of 3,4-DMP on Ca2+ homeostasis and its related physiology in HK-2 human proximal renal tubular epithelial cells. The HK-2 cell, an immortalized proximal tubule epithelial cell lines from normal adult human kidney, was used because it regulates significant Ca2+ handling upon pharmacological stimulation. It has been shown that in this cell, [Ca2+]i rises can be modulated by various chemicals such as 2,3,5-tris(Glutathion-S-yl)hydroquinone 9 and cyclosporin A. 10 Fura-2 was used as a Ca2+-sensitive dye to measure [Ca2+]i. The [Ca2+]i rises were characterized, the concentration-response plots were established, the mechanisms underlying 3,4-DMP-evoked Ca2+ release and Ca2+ entry was examined. The effect of 3,4-DMP on cell viability was also explored in HK-2 cell models.
Materials and methods
Chemicals and agents
3,4-DMP (Figure 1(a)) were from Sigma-Aldrich® (St. Louis, MO, USA). Phorbol 12-myristate 13 acetate [PMA, 1 nM, a protein kinase C (PKC) activator], GF109203X (2 μM, a PKC inhibitor), modulators of store-operated Ca2+ channels (econazole, 0.5 μM; 2-APB, 20 μM; SKF96365, 5 μM), U73122 [2 μM, a phospholipase C (PLC) inhibitor], adenosine triphosphate (ATP, 4 μM, a PLC-dependent agonist), or thapsigargin (TG, 1 μM, an endoplasmic reticulum Ca2+ pump inhibitor) were from Sigma-Aldrich® (St. Louis, MO, USA) unless otherwise indicated. Aminopolycarboxylic acid/acetoxy methyl (fura-2-AM, 2 μM) and 1,2-bis(2-aminophenoxy)ethane-N, N, N’,N’-tetra acetic acid/acetoxy methyl (BAPTA-AM, 5 μM) were from Molecular Probes® (Eugene, OR, USA). The reagents for cell culture were from Gibco® (Gaithersburg, MD, USA).

Effect of 3,4-DMP on [Ca2+]i in fura-2-loaded kidney cell models. (a) The chemical structure of 3,4-DMP. (b) In HK-2 cells, 3,4-DMP was added at 25 s. The concentration of 3,4-DMP was indicated. The experiments were performed in Ca2+-containing medium. Y axis is the [Ca2+]i induced by 3,4-DMP in Ca2+-containing medium. (c) In HK-2 cells, effect of 3,4-DMP on [Ca2+]i in the absence of extracellular Ca2+. 3,4-DMP was added at 25 s in Ca2+-free medium. Y axis is the [Ca2+]i rise induced by 3,4-DMP in Ca2+-free medium. (d) In HK-2 cells, concentration-response plots of 3,4-DMP-induced [Ca2+]i rises in the presence or absence of extracellular Ca2+. Y axis is the percentage of the net (baseline subtracted) area under the curve (25–250 s) of the [Ca2+]i rises induced by 100 μM 3,4-DMP in Ca2+-containing medium. Data are mean ± SD of three independent experiments. *p < 0.05 compared to open circles. 3,4-dimethylphenol: 3,4-DMP; Ca2+: calcium ion; SD: standard deviation.
Solutions used in [Ca2+]i measurements
Ca2+-containing medium (pH 7.4) had 140 mM NaCl, 5 mM KCl, 1 mM MgCl2, 2 mM CaCl2, 10 mM 4-(2-hydroxyethyl)-1-piperazineethanesulfonic acid (HEPES), and 5 mM glucose. Ca2+-free medium contained similar chemicals as Ca2+-containing medium except that CaCl2 was replaced with 0.3 mM ethylene glycol tetra acetic acid (EGTA) and 2 mM MgCl2. 3,4-DMP was dissolved in absolute alcohol as a 0.1 M stock solution. The other chemicals were dissolved in water, ethanol or dimethyl sulfoxide (DMSO). The concentration of organic solvents in the experimental solutions did not exceed 0.1%, and did not affect viability or basal [Ca2+]i.
Cell culture
The HK-2 cells were purchased from American Type Culture Collections (ATCC, Rockville, MD, USA). Cells are cultured in Dulbecco’s Modified Eagle Medium/Nutrient Mixture F-12 (DMEM/F12) medium or Minimum Essential Media (MEM) medium. The medium contains fetal bovine serum (10%) and penicillin (100 U ml−1)-streptomycin (100 µg ml−1). The cells were maintained in a humidified incubator at 37°C in 5% CO2 and 95% air.
Study design
In terms of Ca2+ homeostasis, the experiment was divided four groups: no treatment group (only cells), 3,4-DMP group (3,4-DMP-treated cells), agents’ group (Ca2+ modulators-treated cells), combination of agents and 3,4-DMP group (combination of Ca2+ modulators- and 3,4-DMP-treated cells). In terms of cell viability, the experiment was divided four groups: no treatment group (only cells), 3,4-DMP group [3,4-DMP (0–140 μM)-treated cells], BAPTA-AM group [BAPTA-AM (5 μM)-treated cells], combination of BAPTA-AM and 3,4-DMP group (combination of BAPTA-AM- and 3,4-DMP-treated cells). In experiments using combination of BAPTA-AM and 3,4-DMP, cells were treated with BAPTA-AM 1 h prior to incubation with 3,4-DMP for 24 h.
[Ca2+]i measurements
Confluent cells grown on 6 cm dishes were trypsinized and made into a suspension in culture medium at a concentration of 106 ml−1. Cell viability was determined by trypan blue exclusion. 11 The viability was greater than 95% after the treatment. Cells were subsequently loaded with 2 μM fura-2-AM for 30 min at 25°C in the same medium. After loading, cells were washed with Ca2+-containing medium twice and was made into a suspension in Ca2+-containing medium at a concentration of 107 ml−1. Fura-2 fluorescence measurements were performed in a water-jacketed cuvette (25°C) with continuous stirring; the cuvette contained 1 ml of medium and 0.5 million cells. Fluorescence was monitored with a Shimadzu RF-5301PC spectrofluorophotometer immediately after 0.1 ml cell suspension was added to 0.9 ml Ca2+-containing or Ca2+-free medium, by recording excitation signals at 340 nm and 380 nm and emission signal at 510 nm at 1-s intervals. During the recording, reagents were added to the cuvette by pausing the recording for 2 s to open and close the cuvette-containing chamber. For calibration of [Ca2+]i, after completion of the experiments, the detergent Triton X-100 (0.1%) and CaCl2 (5 mM) were added to the cuvette to obtain the maximal fura-2 fluorescence. Then the Ca2+ chelator EGTA (10 mM) was added to chelate Ca2+ in the cuvette to obtain the minimal fura-2 fluorescence. Control experiments showed that cells bathed in a cuvette had a viability of 95% after 20 min of fluorescence measurements. [Ca2+]i was calculated as previously described. 12
Cell viability analyses
The measurement of cell viability was based on the ability of cells to cleave tetrazolium salts by dehydrogenases. Increases in the amount of developed color correlated proportionally with the number of live cells. Assays were performed according to manufacturer’s instructions (Roche Molecular Biochemical, Indianapolis, IN, USA). Cells were seeded in 96-well plates at a density of 104 cells per well in culture medium at 37°C for 24 h in the presence of 3,4-DMP (0–140 μM). The cell viability detecting reagent 4-[3-[4-lodophenyl]-2-4(4-nitrophenyl)-2H-5-tetrazolio-1,3-benzene disulfonate] (WST-1; 10 µl pure solution) was added to samples after 3,4-DMP treatment, and cells were incubated at 37°C for 30 min in a humidified atmosphere. The cells were incubated at 37°C with/without 3,4-DMP for 24 h. In the experiments using BAPTA/AM to chelate cytosolic Ca2+ to inhibit cytosolic [Ca2+]i rises, cells were treated with 5 μM BAPTA-AM at 37°C for 1 h prior to incubation with 3,4-DMP. The cells were washed once with Ca2+-containing medium and incubated with/without 3,4-DMP for 24 h. The absorbance of samples (A450) was determined using a multiwall plate reader. Absolute optical density was normalized to the absorbance of unstimulated cells in each plate and expressed as a percentage of the control value.
Manganese ions (Mn2+) quenching measurements
Mn2+ quenching of fura-2 fluorescence was performed in Ca2+-containing medium containing 50 μM MnCl2. MnCl2 was added to cell suspension in the cuvette 30 s before the fluorescence recoding was started. Data were recorded at excitation signal at 360 nm (Ca2+-insensitive) and emission signal at 510 nm at 1-s intervals as described previously. 13
Statistics
Data are reported as mean standard deviation (mean ± SD) of three independent experiments (n = 3). Data were analyzed by one-way analysis of variances (ANOVA) using the Statistical Analysis System (SAS®, SAS Institute Inc., Cary, NC, USA). Multiple comparisons between group means were performed by post-hoc analysis using the Tukey’s HSD (honestly significantly difference) procedure. A p-value less than 0.05 was considered significant.
Results
3,4-DMP-induced [Ca2+]i rises in a Ca2+-containing or -free medium in HK-2 cells
The effect of 3,4-DMP on basal [Ca2+]i was investigated in HK-2 cells (Figure 1(b)). Figure 1(b) shows that the basal [Ca2+]i level was 51 ± 2 nM. At 5–100 μM, 3,4-DMP-induced concentration-dependent rises in [Ca2+]i. At a concentration of 100 μM, 3,4-DMP-induced [Ca2+]i rises of 82 ± 5 nM (n = 3). This signal was saturated at 100 μM because 150 μM 3,4-DMP did not induce a larger response (not shown). In Ca2+-free medium, 3,4-DMP also induced concentration-dependent rises in [Ca2+]i at 5–100 μM. At 100 μM, 3,4-DMP induced rises in [Ca2+]i of 51 ± 3 nM (Figure 1(c)). Figure 1(d) shows the concentration-response relationship. The EC50 value was 20 ± 3 μM or 50 ± 2 μM in Ca2+-containing medium or Ca2+-free medium, respectively, by fitting to a Hill equation. Ca2+ removal reduced the Ca2+ signal by approximately 40%.
BAPTA-AM treatment partially reversed 3,4-DMP-induced cytotoxicity in HK-2 cells
Given that acute incubation with 3,4-DMP-induced substantial [Ca2+]i rises, and that unregulated [Ca2+]i rises often alter cell viability, 14 experiments were performed to examine the effect of 3,4-DMP on cell viability. Cells were treated with 0–140 μM 3,4-DMP for 24 h, and the tetrazolium assay was performed. In the presence of 60–140 μM 3,4-DMP, cell viability decreased concentration-dependently (Figure 2). Therefore, the concentration between 60 μM and 140 μM was chosen to determine the effect of 3,4-DMP on cytotoxicity. The next question was whether the 3,4-DMP-induced cytotoxicity was caused by preceding [Ca2+]i rises. The intracellular Ca2+ chelator BAPTA-AM was used to prevent [Ca2+]i rises during 3,4-DMP treatment. 15 After treatment with 5 μM BAPTA-AM, 100 μM 3,4-DMP failed to evoke [Ca2+]i rises (Figure 2(a)). This suggests that BAPTA-AM effectively chelated cytosolic Ca2+. Figure 2(b) shows that 5 μM BAPTA-AM loading did not alter the control value of cell viability. Therefore, in the presence of 60–140 μM 3,4-DMP, BAPTA/AM loading partially reversed 3,4-DMP-induced cell death only in HK-2 cells by 9.2 ± 0.5%, 10.5 ± 0.6%, 11.1 ± 0.8%, 9.9 ± 0.6% or 9.2 ± 0.3% (p < 0.05) (n = 3), respectively (Figure 2(b)). Therefore, the findings implicate that 3,4-DMP-induced cytotoxicity was caused by [Ca2+]i rises in HK-2 cells.

Cytotoxic effect of 3,4-DMP in kidney cell models. (a) Following BAPTA-AM treatment, cells were incubated with fura-2-AM as described in “Materials and methods.” Then [Ca2+]i measurements were conducted in Ca2+-containing medium. 3,4-DMP (5–100 μM) was added as indicated. (b) Cells were treated with 0–140 μM 3,4-DMP for 24 h, and then cell viability assay was conducted in HK-2 cells. Data are mean ± SD of three independent experiments. Each treatment had six replicates (wells). Data are expressed as percentage of control that is the increase in cell numbers in 3,4-DMP-free groups. Control had 11,567 ± 325 cells/well before experiments, and 13,123 ± 333 cells per well after incubation for 24 h. *p < 0.05 compared to control. In each group, the Ca2+ chelator BAPTA-AM (5 μM) was added to cells followed by treatment with 3,4-DMP in Ca2+-containing medium. Cell viability assay was subsequently performed. #p < 0.05 compared to the pairing group. 3,4-dimethylphenol: 3,4-DMP; BAPTA-AM: 1,2-bis(2-aminophenoxy)ethane-N, N, N, N-tetra acetic acid tetrakis (acetoxymethyl ester); Ca2+: calcium ion; SD: standard deviation.
3,4-DMP-induced PKC- or 2-APB-sensitive store-operated Ca2+ entry in HK-2 cells
Because 3,4-DMP-induced Ca2+ response saturated at 100 μM, in the following experiments the response induced by 100 μM 3,4-DMP was used as control. PMA (1 nM), GF109203X (2 μM), econzaole (0.5 μM), 2-APB (20 μM) or SKF96365 (5 μM) were applied 1 min before 3,4-DMP (100 μM), in Ca2+-containing medium.16–18 Except econazole and SKF96365, PMA, GF109203X or 2-APB inhibited 3,4-DMP-induced [Ca2+]i rises in Ca2+-containing medium by 22 ± 2%, 22 ± 3%, or 20 ± 2%, respectively (p < 0.05) (n = 3) (Figure 3). This suggests that PKC- or 2-APB-sensitive store-operated Ca2+ entry were involved in 3,4-DMP-induced [Ca2+]i rises.

Effect of Ca2+ channel modulators on 3,4-DMP-induced [Ca2+]i rises. In blocker- or modulator-treated groups, the reagent was added 1 min before 3,4-DMP (100 μM). The concentration was 10 nM for PMA, 2 μM for GF109203X, 0.5 μM for econazole, 20 μM for 2-APB, or 5 μM for SKF96365. Data are expressed as the percentage of control (first column) that is the area under the curve (25–200 s) of 100 μM 3,4-DMP-induced [Ca2+]i rises in Ca2+-containing medium, and are mean ± SD of three independent experiments. *p < 0.05 compared to the first column. 3,4-dimethylphenol: 3,4-DMP; 2-APB: aminoethoxydiphenyl borate; PMA: phorbol 12- myristate 13-acetate; Ca2+: calcium ion; SD: standard deviation.
3,4-DMP-induced Mn2+ influx in HK-2 cells
The Mn2+ quenching experiment was to explore whether Ca2+ influx occurred during 3,4-DMP incubation. Figure 4 shows that 100 μM 3,4-DMP evoked an instant decrease in the 360 nm excitation signal that reached a value of 220 ± 2 arbitrary units at 250 s, therefore it appears that Ca2+ influx participates in 3,4-DMP-triggered [Ca2+]i rises.

Effect of 3,4-DMP on Ca2+ influx by measuring Mn2+ quenching of fura-2 fluorescence. Experiments were performed in Ca2+-containing medium. MnCl2 (50 μM) was added to cells 1 min before fluorescence measurements. The y axis is fluorescence intensity (in arbitrary units) measured at the Ca2+-insensitive excitation wavelength of 360 nm and the emission wavelength of 510 nm. Trace a: control, without 3,4-DMP. Trace b: 3,4-DMP (100 μM) was added as indicated. Data are mean ± SD of three independent experiments. 3,4-dimethylphenol: 3,4-DMP; Mn2+: manganese ion; Ca2+: calcium ion; SD: standard deviation.
PLC was involved in 3,4-DMP-induced [Ca2+]i rises in HK-2 cells
To exclude the contribution of Ca2+ influx, the experiments were conducted in Ca2+-free medium. U73122 (2 μM) was applied to explore if the activation of PLC was required for 3,4-DMP-induced Ca2+ release.19,20 ATP (4 μM) is a PLC-dependent agonist of [Ca2+]i rises in most cell types including kidney cells.20,21 Figure 5(a) shows that 4 μM ATP-induced [Ca2+]i rises of 51 ± 2 nM. Figure 5(b) shows that incubation with 2 μM U73122 did not change basal [Ca2+]i but abolished ATP-induced [Ca2+]i rises. This suggests that U73122 effectively suppressed PLC activity. In addition, incubation with U73122 inhibited 100 μM 3,4-DMP-induced [Ca2+]i rises and the combination of U73122 and ATP had the same effect (n = 3). U73343 (2 μM), a U73122 analog, failed to have an inhibition on ATP-induced [Ca2+]i rises (not shown). U73343 is a PLC-insensitive structural analog of U73122 and is often used as a control for U73122 activity. This suggests that U73122 inhibited 3,4-DMP-induced [Ca2+]i rises through acting on PLC.
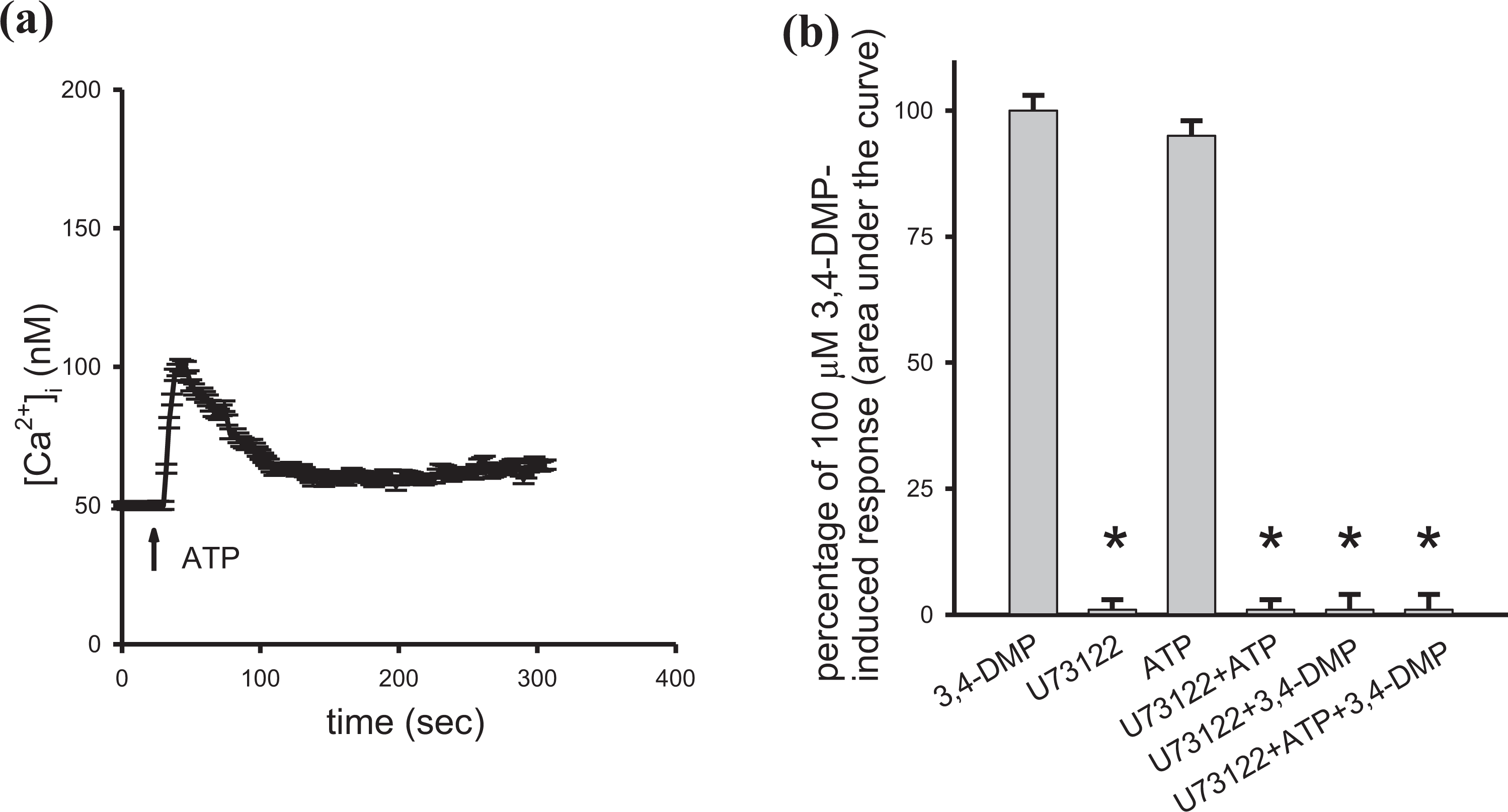
Effect of U73122 on 3,4-DMP-induced Ca2+ release. Experiments were performed in Ca2+-free medium. (a) ATP (4 μM) was added at 25 s. (b) First column is 100 μM 3,4-DMP-induced [Ca2+]i rises. Second column shows that 2 μM U73122 did not alter basal [Ca2+]i (*p < 0.05 compared to first column). Third column shows ATP-induced [Ca2+]i rises. Fourth column shows that U73122 pretreatment for 60 s abolished ATP-induced [Ca2+]i rises (*p < 0.05 compared to third column). Fifth column shows that U73122 pretreatment for 200 s abolished 100 μM 3,4-DMP-induced [Ca2+]i rises (*p < 0.05 compared to first column). Sixth column shows that U73122 (incubation for 60 s) and ATP (incubation for 30 s) pretreatment inhibited 100 μM 3,4-DMP-induced [Ca2+]i rises (*p < 0.05 compared to first column). Data are mean ± SD of three independent experiments. 3,4-dimethylphenol: 3,4-DMP; ATP: adenosine triphosphate; U73122: 1-[6-[((17b)-3-methoxyestra-1,3,5[10]-trien-17-yl)amino]-hexyl]-1H-pyrrole-2,5-dion; Ca2+: calcium ion; SD: standard deviation.
3,4-DMP-induced Ca2+ release from endoplasmic reticulum in HK-2 cells
The source of 3,4-DMP-induced Ca2+ release was conducted in Ca2+-free medium. Figure 6(a) shows that after 3,4-DMP (100 μM) induced a Ca2+ signal, addition of 1 μM TG failed to induce [Ca2+]i rises. 22 Figure 6(b) shows that 1 μM TG-induced [Ca2+]i rises of 22 ± 1 nM. 3,4-DMP (100 μM) added afterward at 500 s did not induce a Ca2+ signal. This suggests that 3,4-DMP-induced [Ca2+]i rises by releasing Ca2+ from the endoplasmic reticulum.

Effect of thapsigargin on 3,4-DMP-induced Ca2+ release. ((a) and (b)) Thapsigargin (TG; 1 μM) and 3,4-DMP (100 μM) were added at time points indicated. Experiments were performed in Ca2+-free medium. Data are mean ± SD of three independent experiments. 3,4-dimethylphenol: 3,4-DMP; TG: thapsigargin; Ca2+: calcium ion; SD: standard deviation.
Discussion
Ca2+ signaling regulates physiology in most cell models including kidney cell models. 13 This study explored the effect of 3,4-DMP on Ca2+ signaling and cell viability in kidney cell models. It has been shown that phenolic compounds such as caffeic acid, ferulic acid or polyphenolic grape extracts affect Ca2+ signaling in various human epithelial cell models.23,24 Our data show that 3,4-DMP increased [Ca2+]i only in HK-2 cells. Because cell types derived from different origins may have different cell function, the Ca2+ signal responses induced by 3,4-DMP appear to vary among different models.
Cell viability could be altered in a Ca2+-dependent or -independent manner.25,26 In our study, 3,4-DMP-induced cytotoxic effect appears to be associated with preceding [Ca2+]i rises because BAPTA/AM pretreatment inhibited 3,4-DMP-induced [Ca2+]i rises partially reversing cytotoxicity in HK-2 cells. Therefore, it suggests that 3,4-DMP induced a Ca2+-associated cell death in HK-2 cells. Because 3,4-DMP-induced Ca2+-associated cell death in HK-2 cells, it might interfere with numerous Ca2+-sensitive processes that orchestrate to alter physiology of kidney models. 14
In terms of [Ca2+]i rises induced by 3,4-DMP in HK-2 cells, the Ca2+ signal was composed of Ca2+ entry and Ca2+ release because the signal was reduced by 40% by removing extracellular Ca2+. The activity of many protein kinases is known to associate with Ca2+ homeostasis.27,28 A previous study has shown that Ca2+ oscillations, Ca2+ sensitization, and contraction were activated by PKC in small airway smooth muscle. 29 Conversely, PKC negatively regulated ADP-induced Ca2+ mobilization and thromboxane generation in platelets. 30 Our data show that 3,4-DMP-evoked [Ca2+]i rises were inhibited by 20% by enhancing or inhibiting PKC activity. It appears that a normally maintained PKC activity is crucial for the maximal function of this enzyme.
In HK-2 cells, the most common Ca2+ entry pathway is the store-operated Ca2+ entry. 9 Our findings show that 3,4-DMP-evoked [Ca2+]i rises were inhibited by 20% by 2-APB but not by econazole or SKF96365. These three compounds have been used to inhibit store-operated Ca2+ entry in various cell models, although there are so far no selective inhibitors for this entry.16–18 Therefore, 3,4-DMP appears to cause Ca2+ entry through 2-APB-sensitive store-operated Ca2+ entry which is induced by depletion of intracellular Ca2+ stores, 31 based on the inhibition of 3,4-DMP-induced [Ca2+]i rises by 2-APB by 20%. 3,4-DMP-evoked [Ca2+]i rises involved Ca2+ influx was confirmed in Mn2+ quenching experiments. Mn2+ enters cells through similar mechanisms as Ca2+ but quenches fura-2 fluorescence at all excitation wavelengths. 12 Therefore, quenching of fura-2 fluorescence excited at the Ca2+-insensitive excitation wavelength of 360 nm by Mn2+ implicates Ca2+ influx. Together, 3,4-DMP induced a PKC-dependent [Ca2+]i signal which was coupled to store-operated Ca2+ entry.
The mechanism underlying the other 20% Ca2+ influx remains to be investigated. One possible pathway involved in 3,4-DMP-induced [Ca2+]i rises is transient receptor potential (TRP) channels. Naturally occurring organic compounds, such as phenolic compounds, modulate or directly activate a variety of TRP channels. 32 A phenolic compound such as carvacrol was shown to modulate Ca2+ homeostasis by activating transient receptor potential vanilloid-3 (TRPV3) channels or transient receptor potential ankyrin-1 (TRPA1) channels in tongue epithelial cells 33 but inhibiting transient receptor potential melastatin 7 (TRPM7) channels in hippocampal brain neurons. 34 Since HK-2 cells have been shown to express the transient receptor potential canonical (TRPC) channels that are Ca2+-permeable, nonselective cation channels with various functions, 35 TRPC channels may have participated in 3,4-DMP-induced [Ca2+]i rises in HK-2 cells. However, so far, there are no selective blockers for TRPC channels. Therefore, it is worthy to further explore this issue.
Because PLC is one of the crucial proteins that modulate the release of Ca2+ from the endoplasmic reticulum, the role of PLC in this process was explored. PLC triggers hydrolysis of phosphatidylinositol 4,5-bisphosphate to inositol trisphosphate (IP3) and diacylglycerol (DAG).7,14 The increased DAG concentration leads to the activation of PKC while IP3 binds to the IP3 receptor (IP3R), an intracellular Ca2+-release channel, thereby inducing Ca2+ release from internal stores.7,14 The data further show that the Ca2+ release was via a PLC-dependent mechanism, given the release was inhibited when PLC activity was inhibited.
Since the endoplasmic reticulum has been shown to be a dominant Ca2+ store in most cell models including endothelial cells,14,36 the role of the endoplasmic reticulum in 3,4-DMP-evoked Ca2+ release was examined. Regarding the Ca2+ stores involved in 3,4-DMP-evoked Ca2+ release, the TG-sensitive endoplasmic reticulum store seemed to be the dominant one. One possible mechanism for 3,4-DMP to release the endoplasmic reticulum Ca2+ is that 3,4-DMP acts similarly to TG by inhibiting the endoplasmic reticulum Ca2+-ATP pump.
Since dimethylphenolic compounds have not been tested in patients, the achievable plasma level is unknown. However, previous studies have explored the cytotoxic effects of dimethylphenolic compounds in animals.37,38 The plasma level of dimethylphenolic compounds may reach 10–20 μM. 39 This level may be expected to go much higher in animals with liver or kidney disorders. 39 Our data show that 3,4-DMP at a concentration of 60 μM induced slight cell death. Therefore, our data may be relevant to in vivo cases.
Together, in HK-2 cells, 3,4-DMP evoked Ca2+ influx via PKC-sensitive store-operated Ca2+ entry and also Ca2+ release from the endoplasmic reticulum in a PLC-dependent manner. 3,4-DMP also caused Ca2+-associated cell death. Because rises in [Ca2+]i can interfere with many cellular processes, caution should be exercised in using 3,4-DMP for other in vitro research, and it should be noted that 3,4-DMP at around 100 μM may be cytotoxic to normal cell models. This study provides a basis for further studies of the role of 3,4-DMP in human health management, and a strong rationale for clinical evaluation of this compound. To this end, it is important to extend the knowledge about the action mechanisms of this compound and to perform additional preclinical studies using in vitro and in vivo models.
Footnotes
Author contributions
K.-J.C. and W.-Z.L.: conceptualization, methodology, formal analysis, resources, data curation and project administration. W.-Z.L.: writing-original draft preparation, writing-reviewing and editing, supervision, visualization.
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This study was supported by Kaohsiung Municipal United Hospital (KMUH109-10).