
Abstract
Objectives:
Our work was aimed at investigating the impact and regulatory mechanism of coenzyme Q10 (CoQ10) on exogenous scrotal heat stress (HS)-induced testicular injuries in rats.
Methods:
The rats (n = 32) were assigned into four groups: control, HS control, HS+CoQ10, and CoQ10. To induce HS, rats’ testicles were immersed in a water bath at 43°C for 20 min, every other day for 8 weeks. Moreover, treatment with CoQ10 (10 mg/kg) immediately started before inducing HS and continued for 8 weeks.
Key findings:
HS decreased the activity of the testicular antioxidant system, superoxide dismutase, glutathione peroxidase, and catalase, while the amount of lipid peroxidation (malondialdehyde) was increased. The index of apoptosis and mRNA expression of caspase 3 and Bax were increased, while the mRNA expression levels of Bcl-2, 3β-HSD, and 17β-HSD3 decreased after HS. Exposure to HS decreased the serum testosterone level but increased the activation of pro-inflammatory cytokines (interleukin 1 beta and tumor necrosis factor-alpha). Deleterious effects of HS on the mentioned parameters were reduced when the rats were treated with CoQ10.
Conclusions:
CoQ10 could suppress the degenerative effects following testicular hyperthermia via its antiapoptotic, anti-inflammation, antioxidative, and androgen synthesis effects.
Introduction
Normal spermatogenesis occurs when the testicular temperature is 2–4°C lower than the temperature of the core body. 1 Several factors play an important role in the regulation of the testicular temperature, including scant hair distribution, thin skin, sweat glands, cremaster and dartos muscles, counter-current heat interchange between testicular veins, the pampiniform plexus, and arterial blood. 1,2
It has been reported that scrotal heat stress (HS) can produce reactive oxygen species (ROS).
2
Overproduction of ROS induced by HS can result in the disruption of seminiferous epithelium, testicular germinal atrophy, and spermatogenic arrest, although ROSs such as hypochlorite, hydroxyl (•OH), and superoxide anion radicals (
It has been demonstrated that when testicles are exposed to HS, the level of hydrogen peroxide is increased. Subsequently, lipid peroxidation is enhanced through decreasing enzymatic antioxidant activity, including catalase (CAT), superoxide dismutase (SOD), and glutathione peroxidase (GPx). 1,2
On the other hand, intrinsic (mitochondrial-dependent) and extrinsic apoptotic pathways, which are induced in response to HS, may lead to cell cycle arrest and germ cell death. 1,5 In addition, oxidative stress induced by HS could activate transcription factors such as nuclear factor κB (NF-κB), which is involved in testicular inflammation and male infertility. 6
Several studies have indicated that natural antioxidants attenuate the destructive effects of free radicals on cellular functions by binding to them. 7,8 Hence, they can possibly be used for the mitigation of the adverse effects of HS on the male reproductive system. It has been reported that coenzyme Q10 (CoQ10, 2,3-dimethoxy-5-ethyl-6-multiprenyl-1,4-benzoquinone) (Figure 1), known as ubiquinone, is able to protect cells against free radicals as a major intracellular antioxidant. 9,10 Moreover, various food sources contain CoQ10, including pork, egg, yogurt, soybean, corn, olive, peanut, and sesame oil, spinach, broccoli, avocado, grapefruit, and so on. 11
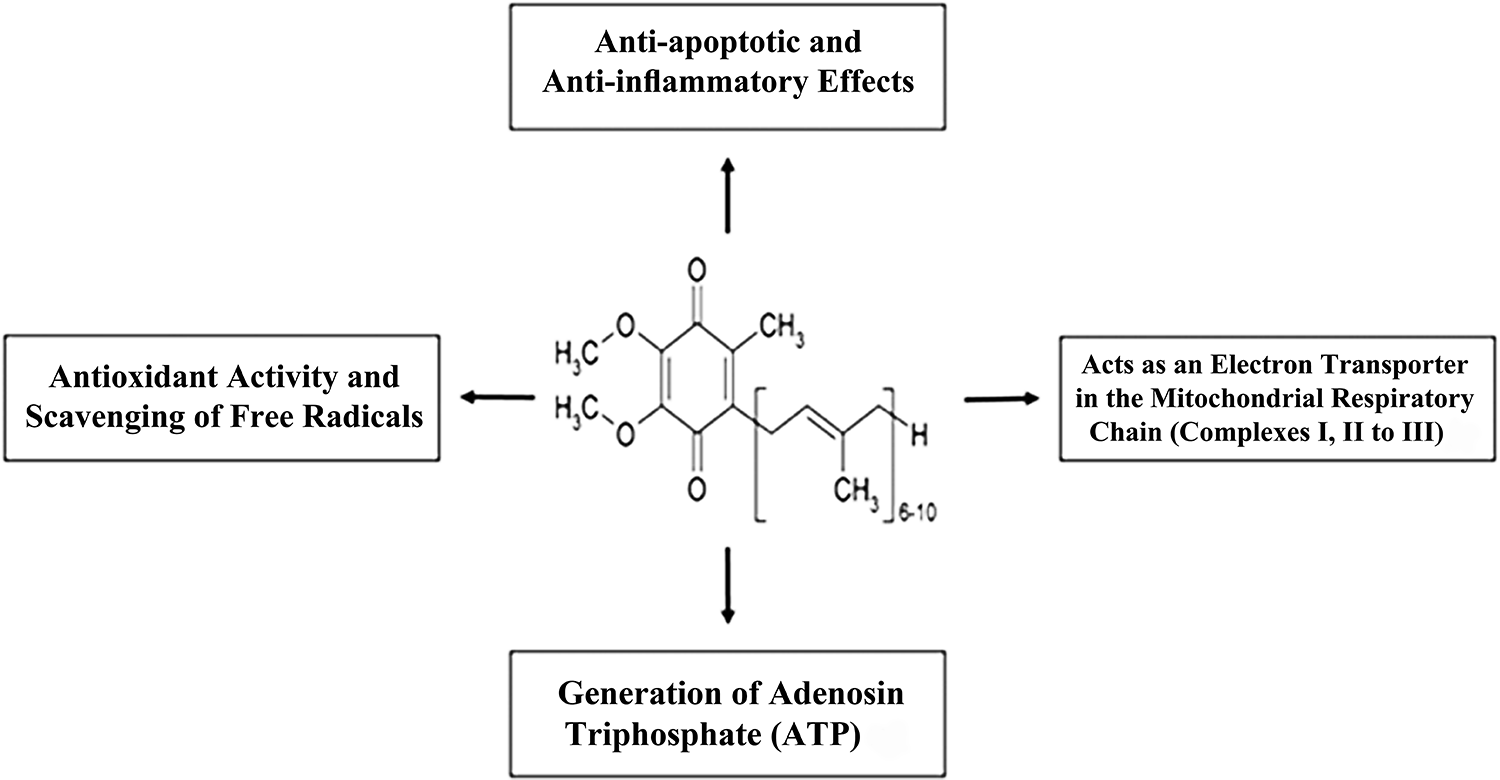
The biological activities and chemical structure of CoQ10. CoQ10: coenzyme Q10.
As a component of the electron-transport chain, CoQ10 plays a significant role in the synthesis of adenosine triphosphate production in mitochondria. 12 The antioxidative property of CoQ10 has been recently reported in our laboratory by decreasing the intracellular levels of ROS in metaphase two (MII) oocytes retrieved from cyclophosphamide-treated mice. 9 In line with these investigations, CoQ10 supplementation can not only decrease oxidative DNA damage and facilitate the process of DNA repair but improve mitochondrial energetics of oocytes. 13,14 A review has reported that the administration of CoQ10 could decrease ROS content and increase the levels of testosterone after testicular damage by a chemical toxicant. 15 One study showed that the administration of CoQ10 protects pancreatic β-cell line MIN6 against staurosporine by suppressing the activation of caspase 3 and the release of cytochrome C from mitochondria. 16 The potent antioxidant capacity of CoQ10 was confirmed in a study conducted by AL-Megrin and colleagues; the results showed that CoQ10 was able to elicit a significant elevation in Nrf2 and HO-1 expression against lead acetate (PbAc)-induced oxidative stress in the kidney. 17
In the light of these pieces of literature, no study has been performed so far to elucidate the impact of CoQ10 on the adverse effects of HS in testicular tissue. Therefore, the aim of this study was to investigate the effects of CoQ10 on apoptosis-related genes (Bax, Bcl-2, caspase 3), steroidogenic genes (3β-HSD and 17β-HSD3), histomorphometry, antioxidant status, and pro-inflammatory cytokines (tumor necrosis factor-alpha (TNF-α) and interleukin 1 beta (IL-1β)) in the testicles of rats following heat exposure.
Materials and methods
Animals and work design
In this study, 32 adult male Wistar rats (weighing 250–300 g) were housed in a room with standard conditions (22 ± 2°C temperature and 40–50% humidity) on a 12-h light/12-h dark cycle. This experimental study received an ethical code (AFG.KIMC.REC.1398.78) from the Ethical Committee of Kabul University of Medical Sciences, Afghanistan. Rats were randomly separated into four groups (n = 8/group), including healthy control group receiving sesame oil (0.2 mL, as same as CoQ10 group), HS control group, HS group receiving CoQ10 (Sigma-Aldrich, Germany, CAS Number: 303-98-0) (10 mg/kg, 18 intraperitoneal (IP) injection, every other day) for 8 weeks (HS+CoQ10 group), and the healthy control group receiving CoQ10 (10 mg/kg, 18 IP injection, every other day) (CoQ10 group). Scrotal HS was induced based on a previous study; briefly, first, the rats were anesthetized by IP injection of ketamine/xylazine (10 mg/kg) (Sigma Aldrich, Germany, CAS numbers: 1867-66-9 and 7361-61-7); then, the testicles of rats were immersed in a water bath at 43°C for 20 min, every other day. 19 Treatment with CoQ10 dissolved in sesame oil immediately started before heat exposure and continued on alternate days for 8 weeks. It has been reported that the half-life of CoQ10 in the blood is about 33 h. 20 After the treatment period, all of the rats were anesthetized as earlier mentioned, and then, testes and serum blood samples were collected and stored (−80°C) for further biochemical and molecular evaluations.
Histopathological assessments
The testis samples (right) were fixed in Bouin’s solution at 4°C (48 h), and routine histological techniques were performed. Afterward, paraffin-embedded tissues were serially sectioned at 5 µm thickness, deparaffinized, and then hematoxylin and eosin (H&E) staining was performed. The prepared sections were examined under a light optical microscope (Nikon, Japan) at 200× magnification. For each seminiferous tubule, spermatogenesis was evaluated by mean Johnson’s score (MJS) (ranging 1–10) to assess the testicular injury. 7 Moreover, 30 cross sections of round-shaped seminiferous tubules were photographed. The epithelium height (EH) and tubular diameter (TD) of each seminiferous tubule were measured by Image J 1.51 software based on our previous studies. 7,21,22
Biochemical surveys
The measurement of oxidative stress markers, including malondialdehyde (MDA), SOD, GPx, and CAT, was carried out in the testis tissues of rats according to our previous studies. 7,21 Bradford method was used to determine the concentration of protein in testicular tissues. Lipid peroxidation was evaluated using the thiobarbituric acid-reactive species and quantified as MDA levels; samples homogenized in 5.1% potassium chloride solution to obtain 1:10 (w/v) of whole homogenates, and finally, the level of MDA calculated using a thiobarbituric acid reaction based on the Uchiyama and Mihara method. 23 SOD and GPx activity was assayed by means of commercial kits (Ransod and Ransel, Randox Com, UK) based on the method of Sun et al. 24 and Paglia and Valentine, 25 respectively. Also, the activity of CAT was determined using a commercial kit (Beyotime Biotechnology, Shanghai, China) by detecting the absorbance of the colored product N-(4-antipyrine)-3-chloro-5-sulfonate-p-benzoquinoneimine at 530 nm using a spectrophotometer. 26
TUNEL staining
Cellular apoptosis was evaluated by the TUNEL peroxidase kit (Roche, Germany). In brief, the sections were deparaffinized, dehydrated, and treated with proteinase K for 30 min at 37°C. 27 Then, the TUNEL reaction mixture was added to samples (50 µL for each sample), which were incubated for 1 h at 37°C. After incubation, the samples were three times washed with phosphate-buffered saline and incubated with converter-POD for 30 min at 37°C. Afterward, they were treated with 3,3-diaminobenzidine for 10 min and finally counterstained with hematoxylin for 1 min. The index of apoptosis was considered as the percentage of cells that were TUNEL-positive. 7,27
Measurement of testosterone level
To measure the testosterone (T) hormone, blood was centrifuged at 1000 × g for 14 min, 28 and then, the serum levels of the hormone were was assayed using a T ELISA kit (Demeditec Diagnostics, Germany) at an absorbance of 450 nm. 27
Determination of IL-1β and TNF-α levels
The concentrations of IL-1β and TNF-α in testicular tissues were evaluated using commercial kits (Sigma-Aldrich, Germany) at an absorbance of 450 nm. 6
RNA extraction, complementary DNA synthesis, and real-time quantitative reverse transcription (qRT)-PCR
Total RNA was extracted from testicles (left) using TRIzol reagent (Invitrogen, Paisley, UK, CAS Number: 9048-46-8) according to the manufacturer’s instructions. To eliminate the DNA contamination, DNase I (Invitrogen Life Technologies, Eugene, Oregon, USA, CAS Number: 7732-18-5) treatment was performed based on previous studies.
7,21
The RNA concentration was determined using the ratio of 260-nm and 280-nm absorbance, the obtained RNA was diluted, and complementary DNA (cDNA) generated using the commercial kit (Thermo Scientific, Germany, EU, CAS Number: 7732-18-5).
7,21
The sequences of specific primers for Bax, Bcl-2, caspase 3, 3β-HSD, 17β-HSD3, and glyceraldehyde-3-phosphate dehydrogenase as the endogenous control are presented in Table 1.
29,30
qRT-PCR was performed in a 20-µL reaction containing 7-mL RNase/DNase free water, 1-mL cDNA, 10-mL SYBR Green Master Mix (Sigma-Aldrich, Germany), and 2-mL forward and reverse primers.
7,21
Subsequently, the quantitative analysis of the mentioned-genes was done using the
Primers for qRT-PCR.

GAPDH: glyceraldehyde-3-phosphate dehydrogenase; qRT-PCR: real-time quantitative reverse transcription-PCR.
Results
Testicular weight
Table 2 presents the testicular weight in all the study groups. In the HS group, the weight of testicles was significantly decreased (68.32 ± 4.1%) relative to that of the control group (p < 0.03). In the HS group treated with CoQ10, the weight of testicles was significantly increased (53.21 ± 3.7%) compared to the HS-untreated group (p < 0.05). However, the comparison between the two healthy control groups showed that there was no significant difference in this regard (p > 0.05).
Testicular weight and quantitative histological evaluation of seminiferous tubules in each group of the study.a

SD: standard deviation; CoQ10: coenzyme Q10; HS: heat stress; MJS: mean Johnson’s score; EH: epithelium height; TD: tubular diameter.
a This table shows a comparison of testicular weight and evaluating the EH, TD, and testicular MJS between all groups of the study.
b p ≤ 0.03, Statistically significant from the control group.
c p < 0.05, Statistically significant from the HS group.
Histopathological evaluations on testicles
H&E staining (Figure 2) of testicles showed normal morphology of the seminiferous tubule epithelium with dynamic spermatogenesis in the two healthy control groups, namely normal and CoQ10-treated rats (Figure 2(a) and (d)). In rats subjected to severe HS, damage in the seminiferous tubules, including abnormal morphological structure and irregularity in germ cells, was observed (Figure 2(b)). Accordingly, disarray and degeneration were observed in all types of spermatogenic cells, including spermatogonia, spermatocytes, and spermatids when compared to the normal. After the administration of CoQ10 to heat-treated groups (Figure 2(c)), the testicular structure was markedly improved compared to the HS-untreated group. On the other hand, the results of TD and EH of seminiferous tubules as well as MJS are presented in Table 2. According to Table 2, TD, EH, and MJS were significantly decreased in the HS group relative to the control group (p ≤ 0.03), while in the treated group, that is, HS-treated rats with CoQ10, the mentioned parameters were significantly increased relative to the HS-untreated group (p < 0.05). Moreover, a comparison between the two healthy groups, that is, normal and CoQ10-treated healthy rats, showed that there was no significant difference for the abovementioned factors (p > 0.05).

Photomicrographs of testicular histology stained with H&E in all the study groups. (a) The healthy control group in which normal architecture of seminiferous tubule and intact spermatogenesis are evident. (b) The HS group in which massive testicular damage, including germ cell destruction, is apparent. (c) The HS group treated with CoQ10 in which a significant improvement relative to the HS group is observed. (d) The healthy control group treated with CoQ10 in which, similar to the control group, normal seminiferous tubule architecture and intact spermatogenesis are observed. Magnification: ×200 (50 µm). H&E: hematoxylin and eosin; CoQ10: coenzyme Q10; HS: heat stress.
Evaluation of CoQ10 effect on antioxidant status
Biochemical parameters including SOD, GPx, MDA, and CAT in the testicles (right) of control and experimental rats were measured to assess the efficacy of CoQ10 against HS, which are presented in Figure 3. According to Figure 3, the activity of SOD, GPx, and CAT antioxidant enzymes was significantly reduced in the testicles of rats exposed to HS compared to the normal, respectively, while the activities of these endogenous antioxidants were significantly increased in rats under HS treated with CoQ10 compared to the HS-untreated group (p < 0.03). On the other hand, although HS increased the level of MDA in testicular tissues relative to that of the control group (p < 0.01), the administration of CoQ10 to rats under HS led to a reduction in the levels of lipid peroxidation as compared to the untreated-HS group (p < 0.05). Moreover, the comparison between the two healthy groups, namely normal and CoQ10-treated healthy rats, showed that there was no significant difference for any of the mentioned factors (p > 0.05).

The effect of HS and CoQ10 on the levels of antioxidant factors in the testicle of rats. (a) SOD, (b) GPx, and (d) CAT graphs showing a significant reduction of the mentioned parameters in both HS groups (HS and HS+CoQ10) compared to the control group (*p < 0.01), while in the HS-treated group (HS+CoQ10), there is a significant improvement (# p < 0.03) compared to the HS-untreated group. Moreover, (c) MDA graph shows that although in testicles exposed to HS (HS and HS+CoQ10 groups), the level of MDA significantly increased compared to the normal control group (*p < 0.01), treatment with CoQ10 (HS+CoQ10 group) significantly decreased the level of MDA (# p ≤ 0.03), compared to the HS group. HS: heat stress; CoQ10: coenzyme Q10; SOD: superoxide dismutase; GPx: glutathione peroxidase; CAT: catalase; MDA: malondialdehyde.
Evaluation of CoQ10 on mRNA expression of apoptotic and steroidogenic genes
qRT-PCR was performed to investigate the impact of CoQ10 on mRNA expression of steroidogenic (3β-HSD and 17β-HSD3) and apoptosis-related (Bax, Bcl-2, and caspase 3) genes in testicular tissue of rats following HS. According to Table 3, the Bax and caspase 3 mRNA levels in testicles of rats exposed to HS were significantly greater than those of the control group (p ≤ 0.001), while the mRNA levels of mentioned-genes (Bax and caspase 3) decreased in the HS-treated rats with CoQ10 (p < 0.003). On the other hand, the mRNA expression of Bax was significantly decreased in the HS condition, while a significant increase was observed in the mRNA expression of Bax when the animals were treated with HS and CoQ10 as compared to the HS group (p < 0.003). In addition, the Bax/Bcl-2 ratio (as an index for the cells to decide whether to survive or die by apoptosis) was significantly increased in rats’ testes exposed to HS compared to the control group (p ≤ 0.001), while it was meaningfully lower in HS-treated group than HS group (p < 0.003). Although HS resulted in the downregulation of key steroidogenic factors (including 17β-HSD3 and 3β-HSD) compared to the control group (p ≤ 0.001), the treatment with CoQ10 significantly enhanced the mRNA expression of them compared to the HS group (p < 0.003). There was no remarkable difference for any of the mentioned genes (Bax, Bcl-2, caspase 3, 3β-HSD, and 17β-HSD3) between the normal and CoQ10-treated normal groups (p > 0.05) (Table 3).
Relative mRNA expression of apoptosis-related and steroidogenic genes analyzed by qRT-PCR.a
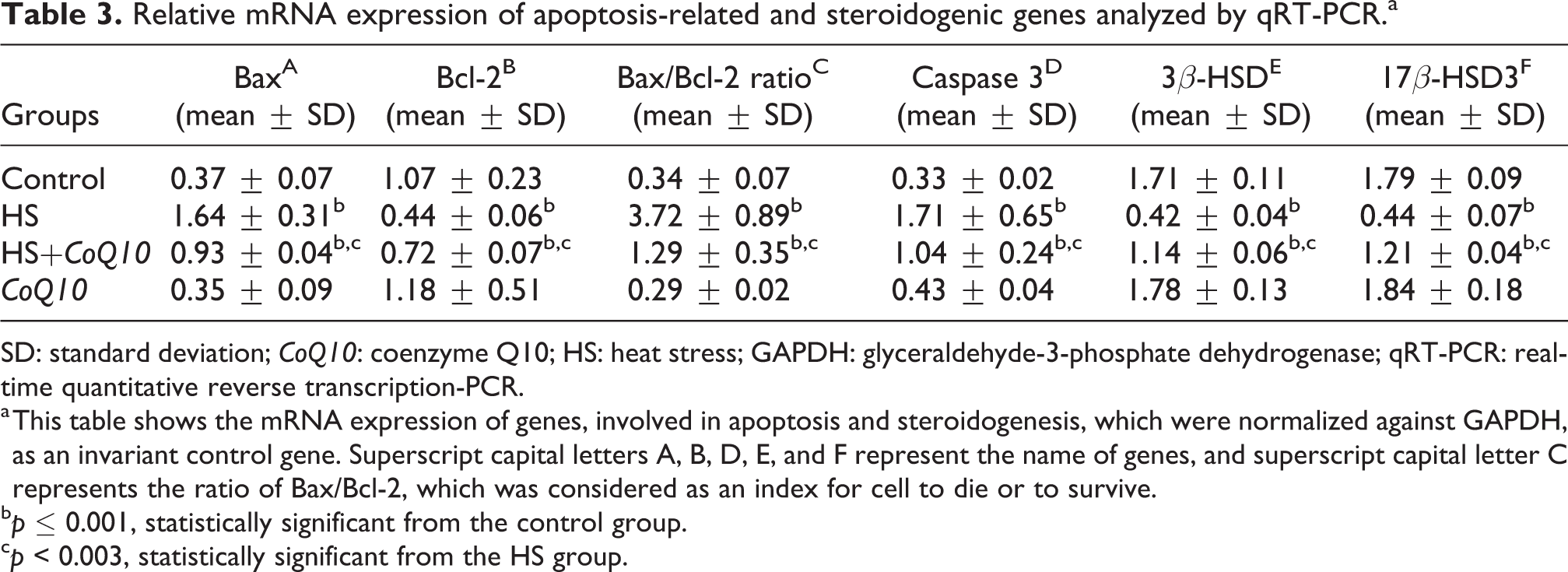
SD: standard deviation; CoQ10: coenzyme Q10; HS: heat stress; GAPDH: glyceraldehyde-3-phosphate dehydrogenase; qRT-PCR: real-time quantitative reverse transcription-PCR.
a This table shows the mRNA expression of genes, involved in apoptosis and steroidogenesis, which were normalized against GAPDH, as an invariant control gene. Superscript capital letters A, B, D, E, and F represent the name of genes, and superscript capital letter C represents the ratio of Bax/Bcl-2, which was considered as an index for cell to die or to survive.
b p ≤ 0.001, statistically significant from the control group.
c p < 0.003, statistically significant from the HS group.
The serum concentration of testosterone
HS led to a statistically significant decrease in serum testosterone concentration relative to the normal control group (p < 0.03) (Figure 4). Furthermore, when the rats were treated with HS and CoQ10, the concentration of T was remarkably increased relative to the HS group (p < 0.05). However, the comparison between the two healthy control groups indicated that there was no significant difference in this regard (p > 0.05) (Figure 4).

Serum testosterone (T) level (ng/mL) in all groups of the study. The graph shows a significant reduction in both HS groups (HS and HS+CoQ10) compared to the control group (*p < 0.03), while serum testosterone level significantly increased in the CoQ10-treated HS group compared to the HS group (# p < 0.05). CoQ10: coenzyme Q10; HS: heat stress.
Evaluation of CoQ10 on pro-inflammatory cytokines in testicles
According to Table 4, HS led to a significant increase in levels of some pro-inflammatory cytokines such as TNF-α and IL-1β relative to the normal control group (p < 0.04). Treatment with CoQ10 remarkably decreased the levels of both cytokines in the HS-exposed state relative to the HS group (p < 0.05). However, a comparison between the two healthy control groups revealed that there was no significant difference in this regard (p > 0.05) (Table 4).
The effects of HS and CoQ10 on the levels of TNF-α and IL-1β in the testis tissue.

SD: standard deviation; CoQ10: coenzyme Q10; HS: heat stress; TNF-α: tumor necrosis factor-alpha; IL-1β: interleukin 1 beta.
a p < 0.04, The amount of TNF-α and IL-1β was significantly increased under HS compared to the control group.
b p < 0.05, Administration of CoQ10 decreased both levels of TNF-α and IL-1β compared to the HS group.
Evaluation of CoQ10 on apoptosis index
TUNEL staining was directed to examine the impact of CoQ10 on testicular germ cell apoptosis. According to Figure 5, HS significantly increased the number of TUNEL-positive cells (the index of apoptosis, Figure 6) relative to the normal control group (p < 0.02). Treatment with CoQ10 markedly reduced the number of apoptotic germ cells in the HS-exposed state compared to the HS-untreated group (p < 0.05). In addition, no significant difference was detected between any of the normal groups (i.e. control and CoQ10) (p > 0.05).

The impact of HS and CoQ10 on germ cell apoptosis in testis. (a) The healthy control group that received sesame oil, (b) HS group, (c) CoQ10-treated HS group, and (d) healthy control group that received CoQ10. Arrows show TUNEL-positive cells. Magnification: ×400 (20 μm). HS: heat stress; CoQ10: coenzyme Q10.
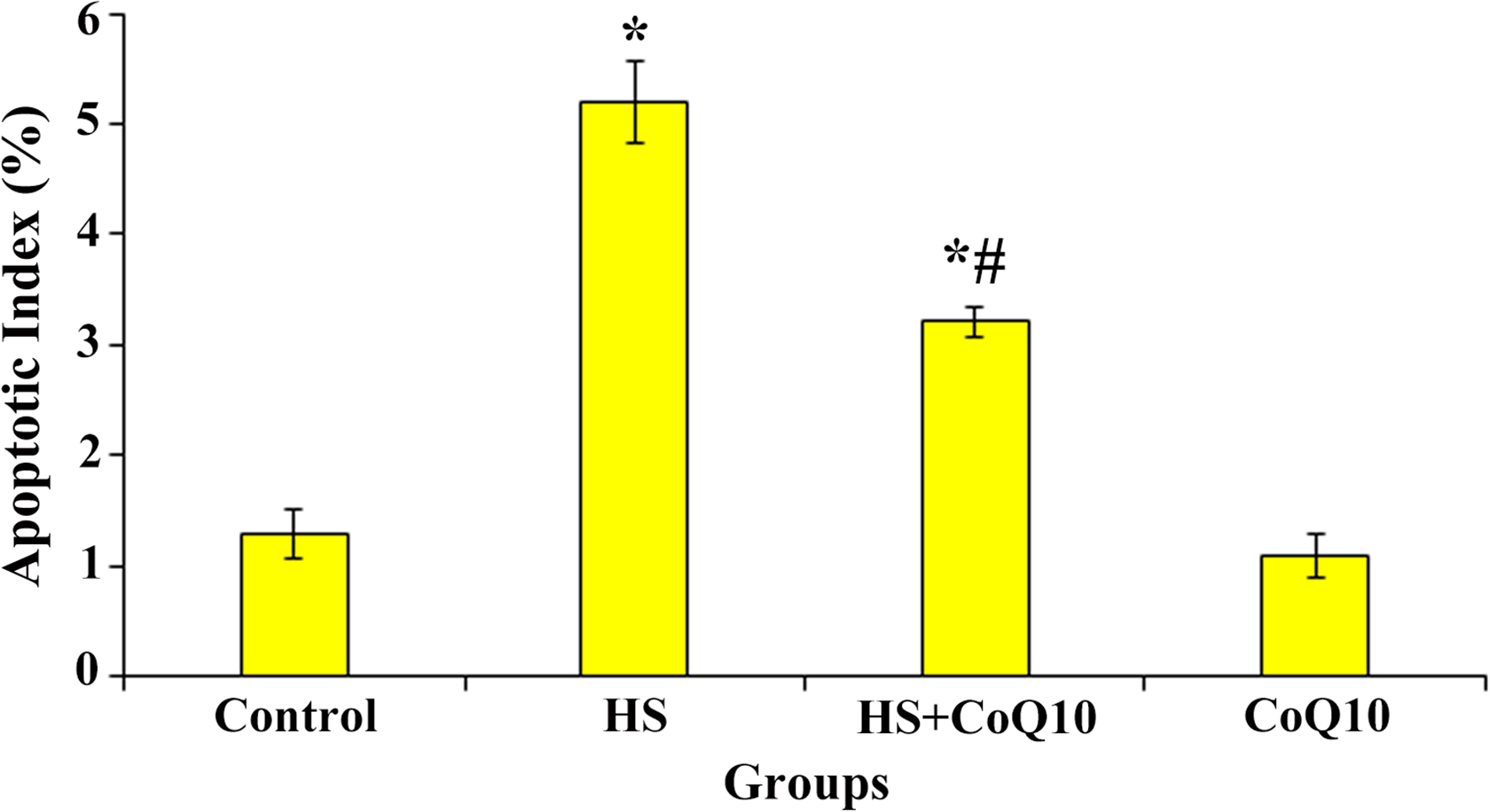
The index of apoptosis in different groups of the study. The graph showing that HS significantly increased the apoptotic index compared to the control group (*p < 0.02), while in the CoQ10-treated HS group, the index of apoptosis remarkably decreased compared to the HS group (# p < 0.05). CoQ10: coenzyme Q10; HS: heat stress.
Discussion
This study is probably the first investigation to show that the deleterious effects of hyperthermia in the testicles can be eliminated by the CoQ10 administration. It has been reported that hyperthermia could reduce testicular weight as well as enhancing germ cells apoptosis in the seminiferous tubules. 1,28
In this study, there was a significant difference between the treated and untreated rats (groups 3 and 2) when the testicle weight was measured. The weight of testicles under scrotal HS decreased compared with the control rats, while in those rats treated with CoQ10, the reduction in testicular weight was significantly recovered.
Interestingly, in the HS group, histopathological images revealed that HS led to the degeneration in testicular tissue, including disturbance of the cellular arrangement in seminiferous tubules as well as germ cell loss, while the administration of CoQ10 induced repaired morphology patterns. In one study, the light microscopy of testicular tissue under scrotal hyperthermia indicated that the order of the cells in seminiferous tubules was disrupted, the seminiferous tubules were atrophied, and the germinal epithelial cells were divided from each other. 31 Therefore, the high rate of weight loss in testicles under HS may be due to testicular damage. Moreover, parameters including the diameter of seminiferous tubules, mean Johnson’s scores, and the height of seminiferous epithelium were markedly reduced under HS, while these histological changes were mitigated in CoQ10-treated group. On the other hand, CoQ10 had no negative effect on the testicular tissue because typical histological features were observed in the CoQ10-healthy control group.
Our biochemical findings showed that scrotal HS increased the MDA level but decreased the activities of SOD, GPx, and CAT in the testicles of rats.
In our research, a high level of MDA has been considered as an index for the status of lipid peroxidation and oxidative stress. Furthermore, the determination of the grade of testicular tissue damage could be evaluated by measuring the MDA level.
7
Increased lipid peroxidation leads to the formation of free radicals, including hydroxyl (OH•), superoxide (
Another part of our results showed that the mRNA expression of genes (Bax and caspase 3) involved in apoptosis decreased compared to the healthy control group.
It has been suggested that ROS-induced oxidative stress regulates the phosphorylation of Bcl-2 family of proteins, which play significant roles in the programmed cell death pathways with individual proapoptotic (i.e. Bax and Bad) and antiapoptotic (i.e. Bcl-xL and Bcl-2) member genes. 7,21,34 ROS induced by HS can also cause protein and DNA damages in testicles of the male gender and enhance the rate of cellular injury and apoptosis (Figure 7). 31 One study showed that in the testicles under HS, the mRNA expression of Bcl-xL decreased, while the mRNA expression of caspase 3 increased, which is involved in caspase-dependent apoptosis initiating molecular pathways. 28 In this regard, our histopathological results also indicated that the number of TUNEL-positive cells was significantly higher in HS-group than normal group. Alternatively, treatment with CoQ10 could prevent those decreases (Bax and caspase-3) in rats subject to HS. It has been reported that CoQ10 prevents apoptosis by inhibiting mitochondrial depolarization. 35 Güleş et al. demonstrated that treatment with CoQ10 (10 mg/kg/daily) decreased the number of TUNEL-positive cells and testicular MDA levels in rats in which testicular toxicity was induced by bisphenol-A. 36 Interestingly, one study showed that daily treatment with CoQ10 (10 mg/kg, IP injection) for 1 week could reduce the mRNA expression of Bax and caspase 3, while increase the Bcl-2 expression in the kidney of rats, in which nephrotoxicity was induced by PbAc. 17 Also, Guo et al. showed that HS was able to enhance the rate of apoptosis through the Fas/FasL pathway. 37

The impact of HS and CoQ10 on the mitochondrial function and the expression of apoptosis-related genes. In response to HS, the exogenous generation of ROS could possibly alter the function of mitochondria via preventing the CoQ10 synthesis, which is involved in the activity of respiratory chain complexes and ATP synthase (F0F1). However, this claim must be further studied. On the one hand, it has been proved that endogenous ROS leads to the activation of apoptosis through the intrinsic apoptosis pathway (mitochondrial pathway). Then, this pathway stimulates mitochondrial outer membrane permeabilization resulting in the release of cytochrome C from the intermembrane space into the cytosol. By binding the cytochrome C to apoptotic protease-activating factor 1, the apoptosome complex is formed, which subsequently leads to the initiation of the caspase cascade, caspase 3 activation, and cell death. Treatment with CoQ10 could decrease ROS and therefore decrease the mRNA expression of Bax and caspase 3 genes possibly via its antioxidant and antiapoptotic effects. CoQ10: coenzyme Q10; HS: heat stress; ATP: adenosine triphosphate; ROS: reactive oxygen species.
On the other hand, the mRNA expression of Bcl-2 as an antiapoptotic protein was significantly decreased in the testes following HS, while in HS group fed with CoQ10, it was remarkably amplified compared to the HS-untreated group.
It has been reported that the antioxidant defense system is indirectly regulated by Bcl-2, leading to a decrease in apoptosis of germ cells. 7 Interestingly, the Bax/Bcl-2 ratio is a predictive marker of apoptosis guiding the cells to decide whether to die or to survive. 21 In the current study, the mentioned ratio was meaningfully higher in the HS-untreated group than the control group, while the administration of CoQ10 decreased such a ratio in the treated-HS group. An investigation showed that pretreatment of rats with CoQ10 decreased cardiac apoptosis induced by ischemia-reperfusion injury via upregulation of Bcl-2 mRNA expression. 38 In this regard, our histopathological examination also indicated that the number of TUNEL-positive cells was remarkably decreased in the treated-HS group than the HS-untreated group. Therefore, it seems that CoQ10 protects the testicular germ cells against the adverse effects of scrotal hyperthermia probably through its antioxidant and antiapoptotic properties.
HS exposure can significantly decrease the serum testosterone level and mRNA expression of 3β-HSD and 17β-HSD3, while the levels of the abovementioned parameters were significantly higher in the treated-HS group.
In the male, a large amount of testosterone is secreted by Leydig cells in testicles, while the suprarenal glands in both sexes can produce smaller amounts of it. 39 However, in Leydig cells, steroidogenesis begins when cholesterol is delivered into the inner membrane of the mitochondrial, which is mediated by the steroidogenic acute regulatory (StAR) protein. 40 Inside the mitochondria, the cytochrome P450 cholesterol side-chain cleavage enzyme (P450scc) converts the cholesterol to pregnenolone. 40 Afterward, pregnenolone is metabolized to progesterone, 17α-hydroxyprogesterone, androstenedione, and testosterone by 3β-HSD, 17α-hydroxylase/C17-20 lyase (P450c17), and 17β-hydroxysteroid dehydrogenase, respectively. 40 It has been stated that the expression of highly regulated genes, including 17β-HSD3 and 3β-HSD, is of high importance for the accurate synthesis of testosterone. 28,40,41 One study indicated that an antioxidant source (curcumin) modulated the 3β-HSD gene and prevented HS-induced damage in Leydig cells, promoting androgen biosynthesis that is essential for normal spermatogenesis. 28 Therefore, in our study, these two genes (17β-HSD3 and 3β-HSD) were used to investigate some of the testosterone synthesis pathways. We also suggest that the other pathways involved in T secretion should be investigated in the future to find how CoQ10 prevents the decrease of T in the HS condition.
Another part of our results concerning the levels of TNF-α and IL-1β indicated the significant increase in the group exposed to HS compared to the normal group, while in HS group fed with CoQ10, these levels decreased compared to the untreated-HS group.
While pro-inflammatory cytokines play key roles in normal homeostasis of testicles, testicular dysfunction could occur following promotion in their expression levels. 42 Elevated TNF-α and IL-1β levels can decrease the serum testosterone levels in human patients with sepsis and/or burns (Cheol Yi Hong). An in vitro study demonstrated that the treatment with TNF-α or IL-1 resulted in an inhibition of steroidogenesis in Leydig cells 43 by downregulating the expression of StAR, 3β-HSD, P450scc, and P450c17, which are involved in testosterone secretion. A number of studies have reported that pro-inflammatory cytokines, such as TNF-α, IL-1β, and IL-6, can activate NF-κB. 6,43 Activated NF-κB inhibits the transactivation of orphan nuclear receptors, including steroidogenic factor 1 as well as Nur77 (nerve growth factor IB), leading to a decrease in serum T level. 42,43 It seems that the decrease in T levels in our study may also be due to these facts. However, we suggest that CoQ10 exerts its anti-inflammatory effects by decreasing the level of TNF-α and IL-1β. Nevertheless, the possible effects of CoQ10 on the NF-κB signaling pathway should be investigated in future studies for a more detailed evaluation of the anti-inflammatory impact of CoQ10.
Conclusion
Our results indicate that the administration of CoQ10 can prevent testicular dysfunction through its antioxidative, anti-inflammatory, and antiapoptotic properties, which were induced by scrotal hyperthermia.
Footnotes
Acknowledgments
The authors would like to thank Dr Mahdi Mohaqiq from US Wake Forest University.
Author contributions
All authors participated in the design, interpretation of the studies and analysis of the data and review of the manuscript; HSH, AD, MD, MM, MAM, and VN have collected the data; HSH, FA, MMHT, and SHAR have written the paper; YKD, HSH, and MNGH have contributed data or analysis tools; HSH, AF, and MD have performed the analysis; and HSH, MD, and MMHT have conceived and designed the analysis.
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) received no financial support for the research, authorship, and/or publication of this article.