
Abstract
The study was to investigate the effects of ligustrazine on rats with cerebral ischemia–reperfusion (I/R) injury and to explore the potential mechanism. Transient focal cerebral ischemia Wistar rat model was established through middle cerebral artery occlusion. The cerebral I/R injury rats were treated with intraperitoneal injection of ligustrazine (1, 3, and 10 mg/kg). Human amniotic epithelial cells (HAECs) were treated with ligustrazine (1, 10, 100 μM) and PI3K inhibitor wortmannin (100 μM), following oxygen–glucose deprivation (OGD) treatment. The expression levels of protein kinase B (PKB or AKT), phospho-Akt (p-Akt), endothelial nitric oxide synthase (eNOS), and phosphor-eNOS (p-eNOS) in HAECs and brains of rats were measured by Western blot. The levels of nitric oxide (NO) in HAECs were measured by Griess method using NO2−/NO3− Assay Kit. Infarct volume and neurological deficits were evaluated 24 h after reperfusion. The levels of NO, p-Akt/Akt, and p-eNOS/eNOS in HAECs were significantly reduced after OGD, but ligustrazine treatment increased the levels of those factors in a dose-dependent manner, while those increases were reversed by PI3K inhibitor wortmannin. Similarly, p-Akt/Akt and p-eNOS/eNOS in brain tissue of rats with I/R were significantly reduced compared with control group (p < 0.05), but ligustrazine treatment increased the levels of p-Akt and p-eNOS in a dose-dependent manner (p < 0.05), while those increases were also reversed by using wortmannin. Ligustrazine also improved the damage of rat brain tissue caused by I/R, but wortmannin reversed the improvement. Ligustrazine plays a neuroprotective role in rats with cerebral I/R injury through the activation of PI3K/Akt pathway.
Introduction
Cerebral ischemia as a cerebrovascular disease is characterized by reduced blood flow to the brain accompanied by pathological changes in brain tissue. 1 In recent years, the incidence of cerebral ischemia has increased significantly due to the change of people’s lifestyle and the quickening of life rhythm. 2 It’s generally believed that more than 80% of stroke cases are caused by cerebral ischemia. 3 Injection of plasminogen activator for thrombolysis is the most effective way for the treatment of cerebral ischemia; however, the application strategy is still challenged by its severe complications, such as cerebral ischemia–reperfusion (I/R) injury. 4 Cerebral I/R injury refers to the aggravated nerve dysfunction after reperfusion. 5 Most of the available treatments including anti-oxidative therapy have been proved to be ineffective in the treatment of cerebral (I/R) injury and its complications. 6 To this end, the study aimed to identify new drugs with higher efficiency and safety to improve the treatment outcomes of cerebral (I/R) injury.
The studies have revealed that the occurrence of cerebral I/R injury is closely related to the degree of inflammatory response. 7 Therefore, how to control the progression of inflammatory response during cerebral I/R injury has been a main task for the treatment of this disease. In recent years, some studies have shown that phosphoinositide 3-kinase (PI3K) and serine/threonine kinase (AKT) signaling pathway is an important pathway mediating cell survival and differentiation, proliferation, apoptosis, and metastasis, which may be related with cerebral diseases. 8,9 Therefore, further understanding of signaling pathways during I/R could provide detailed information about mechanisms.
Ligustrazine, also known as tetramethylpyrazine (TMP), is a traditional Chinese medicine with the functions of activating blood circulation, removing blood stasis, relieving pain, anti-platelet aggregation, and expanding blood vessels, which can be found in fermented cocoa beans and natto. 10 It has been used in the treatment of cardiovascular and cerebrovascular diseases and has achieved good results. Some experimental studies in rats have showed that ligustrazine can play an anti-inflammatory role in different diseases through different pathways. Some studies have shown that ligustrazine can inactivate cAMP/PKA/CREB pathway to improve memory impairments caused by scopolamine. 11 Besides that, cellular inflammatory responses were significantly reduced with ligustrazine treatment in rats with permanent focal cerebral ischemia. 12 And also, ligustrazine showed a promising therapeutic treatment efficacy in the treatment of various I/R injuries including cerebral (I/R) injury. 13,14 However, the mechanism and function of ligustrazine in the treatment of cerebral (I/R) injury still hasn’t been fully understood.
In this study, the effects of ligustrazine on cerebral (I/R) injury were observed at cellular level and in transient focal cerebral ischemia rat model. Simultaneously, the interaction between ligustrazine and PI3K pathway was also explored.
Materials and methods
Cell culture and treatment
Human amniotic epithelial cells (HAECs) were purchased from ATCC (Manassas, VA, USA). Cells were cultured with Endothelial Cell Growth Medium 2 (Heidelberg, Germany) supplemented with gentamicin/amphotericin B, 5% fetal bovine serum, and growth supplements in an incubator (37°C, 5% CO2) for overnight to reach 80–90% confluent. In oxygen–glucose deprivation (OGD) experiments, cells were incubated with OGD buffer pre-deoxygenated for 30 min using nitrogen in an incubator (5% CO2, 95% N2). Reoxygenation was performed by replacing the media. Cells that cultured in regular media under non-hypoxic conditions were used as the control. In the treatment group, ligustrazine (1, 10, 100 μM, Sigma-Aldrich, St Louis, MO, USA) was added at the beginning. Then PI3K inhibitor wortmannin (100 μM, Sigma-Aldrich, St Louis, MO, USA) and
NO assay
The supernatant of HAECs culture medium was collected. NO in the supernatant was detected using NO2−/NO3− Assay Kit (Sigma-Aldrich, St Louis, MO, USA). All operations were performed in strict accordance with the instructions provided by the manufacturer.
Establishment of transient focal cerebral ischemia model
All Wistar rats were from Shanghai Laboratory Animal Center, Chinese Academy of Sciences. Middle cerebral artery occlusion (MCAO) was performed to establish transient focal cerebral ischemia model using the same method described in the previous study. 15 Briefly, after anesthesia with 10% chloral hydrate at a dose of 350 mg/kg, left external and common carotid arteries were separated and ligated. Then a nylon monofilament was inserted into common carotid artery to enter internal carotid artery to block the origin of the middle cerebral artery. Transcranial laser-Doppler (Perimed, Jarfalla, Sweden) was used to monitor the regional cerebral blood flow (rCBF), and the blood flow lower than 20% of baseline indicated the successful occlusion. Reperfusion was performed 2 h later, and the evaluation of neurological deficit was performed 24 h after reperfusion using the method described in a previous study. 16 The rats were anesthetized by intraperitoneal injection of 3% sodium pentobarbital (50 mg/kg). They were perfused with normal saline and decapitated. Their brains were stored in liquid nitrogen for the subsequent experiments.
Animal grouping
Rats were randomly divided into I/R group, ligustrazine group, and sham operation group. Rats in I/R group were treated with intraperitoneal injection of 1% dimethyl sulfoxide at 2 h and 12 h after operation, while intraperitoneal injection of ligustrazine (1, 3, and 10 mg/kg) was performed for rats in ligustrazine group. Intraperitoneal injection of wortmannin (100 nM) was also performed for rats with or without ligustrazine treatment at 30 min before MCAO to plore the mechanism of the endothelial nitric oxide synthase (eNOS) phosphorylation after ligustrazine treatment. Animal experiments were proved by the Animal Ethics Committee of our hospital (No. 2017-194).
Evaluation of infarction volume and neurological deficits
Infarct volume and neurological deficits were evaluated 24 h after reperfusion according to the methods described by Liu et al. 16 Referring to Longa’s 5-level 4-point method, the symptoms of neurological deficit were scored and recorded when rats woke up for 24 h after anesthesia. The scoring criteria were as follows: 0 points, no neurological injury symptoms; 1 point, the contralateral forelimb of the lesion could not be fully straightened when lifting the tail; 2 points, rats turn to the paralyzed side while walking; 3 points, rats fall to the opposite side of the lesion while walking; 4 points, unable to walk spontaneously and lose consciousness. Neurological symptoms were scored by single-blind method, that is, the observers did not know the experimental group.
HE staining
Rat brain tissues were immobilized in 4% paraformaldehyde solution, embedded in paraffin wax after dehydration, and sectioned continuously with a thickness of 5 μm. The sections were dewaxed with xylene and hydrated with ethanol at all levels. The brain sections were stained with hematoxylin (Solarbio, Beijing, China) for 5 min and washed with tap water. They were differentiated by ethanol hydrochloride for 30 s and immersed in tap water for 15 min. Then they were placed in eosin dye (Solarbio, Beijing, China) solution for 2 min. Finally, they were dehydrated, transparent, and sealed. The pathological changes of cerebral cortex ischemic area in rats were observed under 400× optical microscope (Olympus BX51, Japan Olympus Company).
Western blot
Protein extraction from HAECs and brain tissues were performed by conventional method, and protein quality was checked by Bicinchoninic acid (BCA) method (Sigma-Aldrich, St Louis, MO, USA). Then, 12% sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE) gel electrophoresis was carried out to separate different proteins. The proteins were transferred to PVDF through transmembrane. Blocking was performed using 5% skimmed milk. After washing, membranes were incubated with corresponding antibodies including rabbit anti-eNOS antibody (1: 2000, ab5589, Abcam), rabbit anti-p-eNOS (S1177) antibody (1:2000, ab195944, Abcam), rabbit anti-AKT (S1177) antibody (1:2000, ab8805, Abcam), rabbit anti-AKT (S473) antibody (1:2000, ab81283, Abcam), and anti-GAPDH (1:1000, ab9845, Abcam) overnight at 4°C. The membranes were washed, and then incubated with anti-rabbit IgG-HRP secondary antibody (1:1000, MBS435036, MyBioSource) at room temperature for 1 h. Signals detection was performed using electrochemiluminescence (ECL) method (Sigma-Aldrich, USA). Relative expression levels of each protein were normalized to endogenous control GAPDH using Image J software.
Statistical analysis
SPSS19.0 software was used in this study to perform all statistical analyses. Normal distribution data were expressed as x¯ ± SD, and comparisons between two groups were performed using t-test. Abnormal distribution data were compared between groups using one-way ANOVA. p < 0.05 was considered to be statistically significant.
Results
Ligustrazine treatment increased the phosphorylation of eNOS and AKT and the production of NO in HAECs after OGD
As shown in Figure 1, compared with control group, phosphorylation levels of eNOS and AKT (normalized to eNOS and AKT) and the production of NO were significant reduced in OGD group cells (p = 0.00). Compared with OGD group, phosphorylation levels of eNOS and AKT and the production of NO were significant increased by ligustrazine treatment, and the increase showed a dose-dependent manner (p = 0.03 or p = 0.00). Those data suggest that ligustrazine treatment could activate AKT pathway and promote the production of NO in HAECs after OGD.
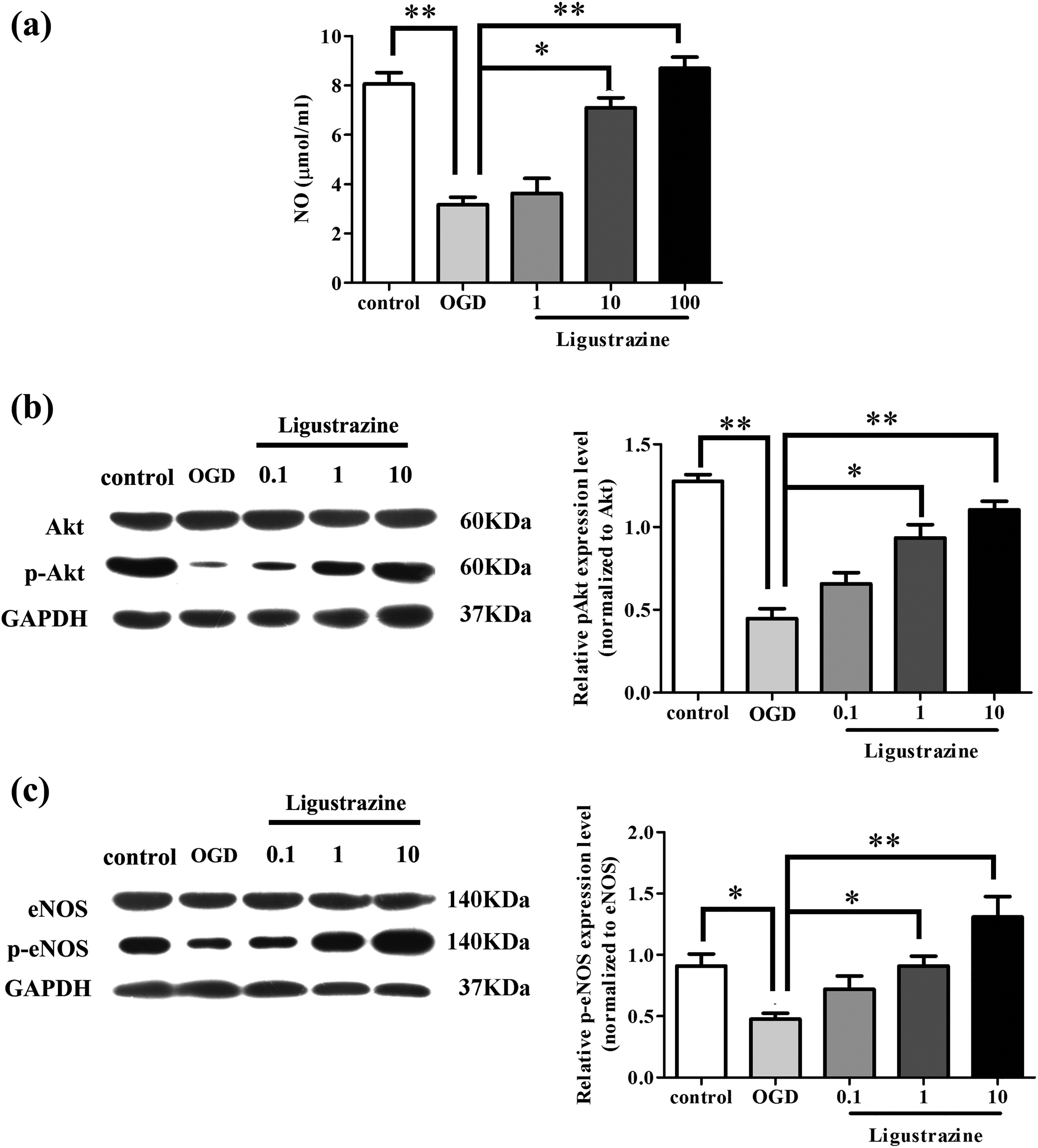
Effect of ligustrazine on PI3K/AKT pathway and NO production in HAECs after OGD. (a) Levels of NO in cells of each group; (b) levels of Akt and p-Akt in cells of different groups tested by Western blot; and (c) levels of eNOS and p-eNOS in cells of different groups measured by Western blot. *p < 0.05; **p < 0.01. NO: nitric oxide; OGD: oxygen–glucose deprivation; eNOS: endothelial nitric oxide synthase; HAEC: human amniotic epithelial cell; p-eNOS: phosphor-eNOS.
Ligustrazine treatment increased the phosphorylation of eNOS in HAECs through the PI3K/AKT pathway
In order to investigate the mechanism of the increased phosphorylation of eNOS in HAECs after OGD caused by ligustrazine treatment, PI3K inhibitor wortmannin was also used to treat HAECs. As shown in Figure 2, compared with control group, ligustrazine treatment alone failed to significantly increase the levels of p-Akt/Akt and phosphor-eNOS (p-eNOS)/eNOS, indicating that ligustrazine has no significant effects on PI3K/Akt pathway and NO production in normal HAECs. Compared with OGD cells with ligustrazine treatment alone, levels of p-Akt/Akt and p-eNOS/eNOS were significantly decreased in OGD cells with both ligustrazine treatment and wortmannin treatment (p = 0.03, p = 0.00). Those data suggest that the decrease in the level of p-eNOS in HAECs after OGD is achieved through the activation of PI3K/Akt pathway.

Ligustrazine treatment increased the phosphorylation of eNOS through the interaction with PI3K/AKT pathway in HAECs. (a) Levels of Akt and p-Akt in cells with different treatments and (b) levels of eNOS and p-eNOS in cells with different treatments. *p < 0.05; **p < 0.01. eNOS: endothelial nitric oxide synthase; HAEC: human amniotic epithelial cell; p-eNOS: phosphor-eNOS.
Ligustrazine treatment increased the phosphorylation of eNOS and AKT in I/R rats
Similar to the results obtained from HAECs after OGD, as shown in Figure 3, ligustrazine treatment significantly increased the levels of p-eNOS and p-AKT in brain tissue of I/R rats (p = 0.04, p = 0.00). The increase also showed a dose-dependent manner. Those data suggest that ligustrazine treatment can activate PI3K/Akt pathway and increased the levels of p-eNOS to induce the production of NO in I/R rats.

Effects of ligustrazine treatment on the phosphorylation of eNOS and Akt in rats with I/R. (a) Levels of eNOS and p-eNOS in rats of different groups and (b) levels of Akt and p-Akt in rats of different groups. n = 6, *p < 0.05; **p < 0.01. I/R: ischemia–reperfusion; eNOS: endothelial nitric oxide synthase; p-eNOS: phosphor-eNOS.
Ligustrazine treatment increased the phosphorylation of eNOS in brain tissue of I/R rats through the interaction with PI3K/AKT pathway
As shown in Figure 4(a), compared with control group, neurological damage was significantly increased in cerebral I/R rats (p = 0.00). Compared with cerebral I/R rats, ligustrazine treatment significantly improved the neurological damage (p = 0.02), while this improvement was almost reversed by the treatment of PI3K inhibitor wortmannin. As shown in Figure 4(b) and (c), compared with cerebral I/R rats only treated by ligustrazine, levels of p-eNOS and p-Akt were significantly reduced in I/R rats with both ligustrazine treatment and wortmannin treatment. Besides that, ligustrazine treatment also significantly improved the conditions of brain tissue from cerebral I/R rats; however, PI3K inhibitor reverted the effect of ligustrazine (shown in Figure 4(d)). Those data suggest that ligustrazine can play a neuroprotective role in rats with cerebral I/R by increasing the production of NO by activating PI3K/Akt pathway.

Ligustrazine treatment increased the phosphorylation of eNOS in brain tissue of rats with I/R through the interaction with PI3K/AKT pathway. (a) Neurological scores of rats in each group; (b) levels of p-eNOS in each group; (c) levels of p-Akt in each group; and (d) representative results of coronal brain section staining with 2,3,5-triphenyltetrazolium chloride. Data were expressed as mean ± SEM. n = 6, *p < 0.05; **p < 0.01. I/R: ischemia–reperfusion; eNOS: endothelial nitric oxide synthase; p-eNOS: phosphor-eNOS.
Morphological effects of Ligustrazine on I/R rats
As shown in Figure 5, the cells in Sham group had normal morphology, complete nuclear membrane, clear nucleolus, no pyknosis, no infiltration of cells in the gap, and they were arranged neatly. In the model group, the cells morphology was changed seriously, and their volumes were reduced. The nucleuses were deep-stained and the edemas were serious. The cell infiltration occurred in the gap, and the pathological changes and necrosis were common in nerve cells. However, after treatment, the pathological morphology and edema of the cells were improved, and the deep staining and shrinkage of the cells were gradually alleviated.

Morphological effects of ligustrazine on I/R rats (×400). (a) Sham; (b) I/R; (c) ligustrazine 1 mg/kg; (d) ligustrazine 3 mg/kg; and (e) ligustrazine 10 mg/kg. I/R: ischemia–reperfusion.
Discussion
Ligustrazine is a chemical compound which usually can be found in fermented cocoa beans and natto. 10 In China, ligustrazine isolated from Chinese herb medicine chuanxiong has been widely used for a long time to treat various diseases including asthma. 17,18 Based on the anti-inflammatory and neuro-protective effect, ligustrazine has also been used to treat different types of I/R injuries. Feng et al. have shown that ligustrazine can improve renal dysfunction caused I/R of the kidney, which is an unavoidable complication of renal transplantation. 19 In another study, the same research group showed that ligustrazine could inhibit oxidative stress, reduce oxidative stress, decrease neutrophils infiltration, and reduce the expression levels of ICAM-1 and TNF-α in rats, which in turn protects kidney from the injuries caused by I/R. 20 In the study of myocardial I/R injury, Chen et al. showed that ligustrazine could induce the expression of heme oxygenase-1 to reduce the I/R injuries. 21 Ligustrazine also showed therapeutic effects in the treatment of cerebral I/R injury. Recently, ligustrazine was found to be able to decrease the increased striatal concentrations of neurotransmitters and neurological deficits caused by cerebral I/R injury. 22 Consistent results were found in our study in which ligustrazine treatment significantly improved the neurological deficits in rats caused by MCAO. These results further confirmed the therapeutic effects of ligustrazine in the treatment of cerebral I/R injury.
The development and progression of cerebral I/R injury is a complex process with various signaling transduction pathways involved. The eNOS/NO signaling pathway plays pivotal roles in the regulation of cerebral blood flow, especially under conditions of fluctuating arterial pressure. Therefore, eNOS/NO signaling transduction is essential in maintenance of sufficient cerebral blood flow. 23 In this study, levels of NO and p-eNOS was significant reduced in HAECs after OGD compared with normal control cells, while ligustrazine treatment significantly increased the levels of p-eNOS and NO in HAECs. In addition, the reduced level of p-eNOS in rat brain tissue caused by I/R was also increased by ligustrazine treatment. Those results suggest that ligustrazine treatment can activate eNOS/NO signaling pathway to increase the production of NO in HAECs after OGD. PI3K/Akt signaling pathway plays essential roles in various biological processes including cell growth, proliferation, and survial. 24 Numerous studies have showed that the activation of Akt by phosphorylation can play a protective role to reduce apoptosis induced by I/R. 25 It has been proved that PI3K/Akt pathway can activate eNOS/NO signaling pathway to perform its biological functions. 26 In this study, the activation of PI3K/Akt pathway was also found to be inhibited in rats with I/R and HAECs with OGD, while ligustrazine treatment significantly increased the level of p-Akt, indicating that ligustrazine can activate PI3K/Akt pathway to protect against I/R injury. Treatment with PI3K inhibitor wortmannin reversed the increased levels of NO, p-Akt, and p-eNOS after ligustrazine. In addition, wortmannin also reversed the improvement of neurological function in I/R rats after the treatment of ligustrazine. Those results suggest that the activation of PI3K/Akt pathway is essential for the function of ligustrazine in activating eNOS/NO signaling pathway in both HAECs with OGD and rat with I/R injury.
Safety is also a critical point for the application of a drug in the treatment of certain diseases. In this study, ligustrazine showed no significant effects on neurological function and levels of p-Akt and p-eNOS in normal rats, which indicated that ligustrazine will not disturb normal signal transduction and neurological function. Therefore, ligustrazine could potentially be effective drug with high safety for the treatment of cerebral I/R injury in human.
Conclusions
In conclusion, our study showed that ligustrazine could protect against cerebral I/R injury in both HAECs with OGD and rat cerebral I/R injury model. The function of ligustrazine in cerebral I/R injury is very likely to be achieved by activating eNOS/NO signaling pathway to increase the production of NO through the activation of PI3K/Akt pathway. Our study provided the basis for future studies on the application of ligustrazine in the treatment of cerebral I/R injury in human. However, clinical studies are lacking in this study. Therefore, further clinical studies are still needed to further confirm the conclusions made in this study.
Footnotes
Author contributions
Yongbo Ding and Juan Du contributed equally to this article.
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) received no financial support for the research, authorship, and/or publication of this article.