
Abstract
Most organisms feature an endogenous circadian clock capable of synchronization with their environment. The most well-known synchronizing agents are light and temperature. The circadian clock of mosquitoes, vectors of many pathogens, drives important behaviors related to vectoral capacity, including oviposition, host seeking, and hematophagy. Main clock gene expression, as well as locomotor activity patterns, has been identified in Aedes aegypti and Culex quinquefasciatus under artificial light-dark cycles. Given that these mosquito species thrive in tropical areas, it is reasonable to speculate that temperature plays an important role in the circadian clock. Here, we provide data supporting a different hierarchy of light and temperature as zeitgebers of two mosquito species. We recorded their locomotor activity and quantified mRNA expression of the main clock genes in several combinations of light and temperature cycles. We observed that A. aegypti is more sensitive to temperature, while C. quinquefasciatus is more responsive to light. These variations in clock gene expression and locomotor activity may have affected the mosquito species’ metabolism, energy expenditure, fitness cost, and pathogen transmission efficiency. Our findings are relevant to chronobiology studies and also have epidemiological implications.
Many organisms are exposed to dramatic cyclical variations in their environment; consequently, such organisms must adjust their metabolism and behavior to properly establish a temporal niche. Insects are notably adaptable to new niches, which is crucial to survival. Insects are poikilothermic, which requires that they anticipate and restrict their activity to mild temperatures as well as avoid heat and desiccation. The endogenous machinery of almost all living organisms, also known as the circadian clock, adjusts their physiologies and behaviors according to their surrounding environments. This endogenous clock is self-sustaining even in constant environmental conditions and governs the overall rhythmicity of an organism (Aschoff, 1963; Dunlap and Loros, 2004). According to Aschoff (1963), the endogenous system exhibits a range of entrainment that depends on the characteristics of the driven system and the strength of the driving agent.
Light and temperature cycles are the most important synchronizers, or zeitgebers (reviewed in Saunders, 2002). Yoshii et al. (2009) observed that combining both external cues resulted in an improved amplitude of molecular and behavioral rhythmicity in the fruit fly Drosophila melanogaster compared with the results of isolating these two zeitgebers. Moreover, a genome-wide expression analysis of D. melanogaster showed that hundreds of genes were influenced by light and temperature (Boothroyd et al., 2007). Some of these genes were dependent on the circadian clock machinery; others were uniquely influenced by light or temperature and thus bypassed the circadian clock (Boothroyd et al., 2007). The importance of light to organism entrainment is significant, but in recent years many reports have demonstrated that the importance of temperature surpassed that of light for circadian clock entrainment. Previous analysis of D. melanogaster under light-dark (LD) and temperature cycles (TC) in opposite phases provided evidence that light is the main zeitgeber of larger delays, whereas temperature dictates the entrainment of smaller delays (Yoshii et al., 2010). Under more natural conditions, temperature was essential to the entrainment of fruit flies, distinct from observations in laboratory-controlled conditions (Vanin et al., 2012).
Both zeitgebers can directly or indirectly influence behavior via the molecular clock machinery, composed of several interlocked transcription factors (TFs) responsible for driving clock gene expression (reviewed in Mendoza-Viveros et al., 2016). In Drosophila, CLOCK (CLK) and CYCLE (CYC) are positive elements that activate the rhythmic transcription of hundreds of genes in multiple feedback loops, including period (per), timeless (tim), vrille (vri), and Par domain protein 1 (Pdp1), all of which are negative regulators (reviewed in Rivas et al., 2016).
The expression patterns of orthologous clock genes in dipterans and Drosophila are remarkably similar (Meireles-Filho and Kyriacou, 2013); however, the expression profiles of Clock and cycle are dissimilar in fruit flies, mosquitoes, and sand flies (Meireles-Filho et al., 2006a; Meireles-Filho et al., 2006b; Gentile et al., 2009; Rund et al., 2011; Rund et al., 2013; Leming et al., 2014).
Aedes aegypti and Culex quinquefasciatus are known to exhibit different habits (diurnal and nocturnal, respectively); however, the mosquito species share circadian transcription profiles for most canonical clock genes (Gentile et al., 2009). This is especially noteworthy considering that the species may have diverged between 52 and 22 million years ago (Foley et al., 1998). That being said, the cryptochrome 2 (cry2) gene has a bimodal pattern of expression in A. aegypti and only a unimodal pattern of expression in Culex. The evolutionary history of cry genes has been characterized by gene loss and duplication (Yuan et al., 2007) and their fundamental roles in circadian repression, temporal mating, magneto sensitivity, and sun compass orientation (Yuan et al., 2007; Zhu et al., 2005; An et al., 2004; Gegear et al., 2008; Zhu et al., 2008). Accordingly, cryptochrome genes should be considered as excellent research candidates to improve our understanding of species-specific behavioral differences.
The circadian clock dictates host-seeking rhythmicity, which is crucial to insect vector efficiency (Das and Dimopoulos, 2008; Gentile et al., 2013; Lima-Camara et al., 2014). Moreover, the environment can affect vector capacity. Recent studies have observed the influence of temperature on the extrinsic incubation period and transmission of arthropod-borne virus (Lambrechts et al., 2011; Carrington et al., 2013; Alto et al., 2014; Zouache et al., 2014; Christofferson and Mores, 2016; Danforth et al., 2016). Some reports have shown that the dispersal and adaptation of many insect vectors into temperate regions are due to climate change (Rogers et al., 2014; Chadee and Martinez, 2016). Thus, it is imperative to study the effects of the environment on the behavior and gene expression of insect vectors in order to understand how the environment can modulate the circadian clock and drive vector competency.
This study aims to observe the locomotor activity and circadian clock gene expression of Aedes aegypti and Culex quinquefasciatus, the main vectors of Dengue, Chikungunya, and Zika arboviruses and West-Nile virus, respectively. Both mosquito species’ daily locomotion and clock gene expression were evaluated under various artificial light-dark regimes and temperature cycles. Our results concur with previous findings and reveal notable differences regarding the behavior and gene expression of Aedes and Culex. Our data show how both species respond distinctly to light and temperature cues. This research suggests that alternative hierarchies of zeitgebers exist between different species of insects.
Methods
Mosquitoes
All experiments were conducted with female mosquitoes from laboratory colonies of A. aegypti (Rockefeller strain) and C. quinquefasciatus (IBEX strain) maintained at IBEX (Instituto de Biologia do Exército, Rio de Janeiro, Brazil). The Rockefeller strain was derived from mosquitoes collected in Cuba, Nigeria, Philippines, and Puerto Rico, interbred in the United States in the 1930s, and distributed worldwide (Kuno, 2010). The C. quinquefasciatus mosquitoes were collected in Rio de Janeiro, and the colony was established at IBEX in 2000 (Belinato et al., 2013). The pupa-stage mosquitoes were synchronized from pupa stage to 12 h of light and 12 h of dark (LD) under a constant 25 °C, and the newly emerged, virgin females were separated from the males.
Locomotor Activity Recording
The insects were individually transferred to 1 × 7 cm glass tubes with a cotton plug soaked in 10% sucrose solution at one end, and both ends were properly sealed with Parafilm M (Sigma-Aldrich, St. Louis, MO). Locomotor activity analyses were performed with a larger version of the Drosophila DAM5 Activity Monitoring System (Trikinetics Inc., Waltham, MA), as described in previous studies (Gentile et al., 2009; Gentile et al., 2013). The activity of the 2- to 3-day-old female mosquitoes was monitored under different artificial photoperiod (12:12 LD) and temperature (daily fluctuation of 10 °C or 20 °C constant) regimens (see Results section for details) in a Precision Scientific Incubator Mod. 818 (Thermo Scientific, Waltham, MA). Each insect’s movement was detected by an infrared interruption method, and daily locomotion was recorded during 30-min intervals using computer software (DAMSystem3 Software; Trikinetics Inc). All experimental procedures were repeated twice. To calculate the mean locomotor activity of each time interval, we first transformed the data to logarithm values, calculated the arithmetic means, and then reconverted these values to numbers, based on the Williams mean method (Wm) (Williams, 1937). This procedure has been used by our group to correct the effects of high activity displayed by some specimens that can produce biases in mean activity (e.g., Gentile et al., 2009; Lima-Camara et al., 2011). All graph comparisons were made with Excel (Microsoft, Redmond, WA). The actograms were double-plotted with ActogramJ Software (Schmid et al., 2011).
To conduct our analysis of constant darkness conditions (see Results for details), we measured the free-running period of 10 consecutive days using a χ2 periodogram algorithm with ActogramJ, as used previously (Gentile et al., 2013; Pavan et al., 2016). Mosquitoes with visible activity throughout the entire experiment and a χ2 periodogram with a peak above the confidence level were considered rhythmic and were used for actogram plotting and free-running period estimation. The rhythmic consistency of both species was measured by calculating the power, which is defined as the amplitude from the top of the peak to the confidence level in the χ2 periodogram (Liu et al., 1991). This method has been proven to effectively evaluate the rhythm strength of different insect species (Moriyama et al., 2008; Uryu et al., 2013; Gentile et al., 2013; Pavan et al., 2016).
Sample Collection, Processing, and Gene Expression Analysis
Female mosquitoes of both species were entrained for 2 days under different artificial regimens: (1) LDTC: 12:12 LD with temperature cycles of 30 and 20 °C; (2) LDCT: 12:12 LD with temperature cycles of 20 and 30 °C; (3) LLTC: constant light with temperature cycles of 30 and 20 °C; (4) DDTC: constant darkness with temperature cycles of 30 and 20 °C. On the following day (day 3), 10 females were collected and beheaded every 2 h for a 24-h period, as previously described (Gentile et al., 2006). Each experiment represented 12 time point samples, and this procedure was repeated 4 times. The total RNA extraction used the TRIzol method (Invitrogen, Carlsbad, CA), and the cDNA synthesis of the TaqMan Reverse Transcription Reagents (Applied Biosystems, Foster City, CA) followed the methods described by Gentile et al. (2009). Each time point sample had a final cDNA concentration of 1 ng/µL. All samples were used to determine the relative mRNA abundance via quantitative real-time PCR (qPCR), using the Power SYBR Green PCR Master Mix (Thermo Fisher, Waltham, MA) in a StepOnePlus Real-Time PCR System (Thermo Fisher). We conducted these experiments using the same oligonucleotides designed by Gentile et al. (2009), and we measured the relative mRNA abundance with the comparative CT method (Pfaffl, 2001) using the rp49 gene as a constitutive control (Gentile et al., 2006, 2009). All values of relative mRNA abundance respective to all genes studied here were illustrated in graphs via Excel or heat maps with dendrograms using the packages gplots v. 3.0 and RColorBrewer v. 1.2 in R environment (https://cran.r-project.org/).
Statistical Analysis
To perform the statistical analyses, we first tested whether all parameters followed a Gaussian distribution with the Shapiro-Wilk normality test (α = 0.05). Moreover, the proportions of diurnal and nocturnal activity were normalized by transforming these values with the arcsine square root, prior to statistical analysis. Behavioral results were statistically evaluated using the Student t test or its nonparametric analogue (Mann-Whitney test). We also measured whether the mRNA relative abundance of each gene in both species varied significantly throughout the 24-h period of each artificial regimen. We considered that a certain gene had a rhythmic expression only if the mRNA abundance differed significantly among the 12 time point samples using a 1-way ANOVA (α = 0.05). A nonparametric test (Mann-Whitney U test) was performed to determine the significance of the difference of mRNA abundance between species at a specific time point (ZT). All statistical analyses were conducted with the GraphPad Prism 5 (Prism, La Jolla, CA).
Results
Daily Locomotor Activity Rhythms with Temperature Cycles and with or without Light-dark Cycles
At first, we artificially maintained both species under combined light-dark regimens and temperature cycles (LDTC—light-dark with temperature cycles of 30 and 20 °C) during 4 consecutive days (Fig. 1). Under these conditions, the diurnal and nocturnal patterns of Aedes and Culex were not affected, since most of their activities were sustained during their respective phase of day. Both species shared an evening peak in the early night (ZT 12-13); however, at this time point, the activity levels of Culex were higher than those of Aedes (total activity during ZT 12-13 for nonnormalized data: A. aegypti = 23.09 and C. quinquefasciatus = 59.20). Under light-dark cycles (at a constant temperature of 25 °C), A. aegypti repeatedly exhibited a bimodal pattern of activity with peaks in the morning and the evening (Gentile et al., 2009; Lima-Camara et al., 2011, 2014). Interestingly, we observed that the presence of a temperature cycle as an additional zeitgeber tended to suppress Aedes’s morning peak (Fig. 1A).
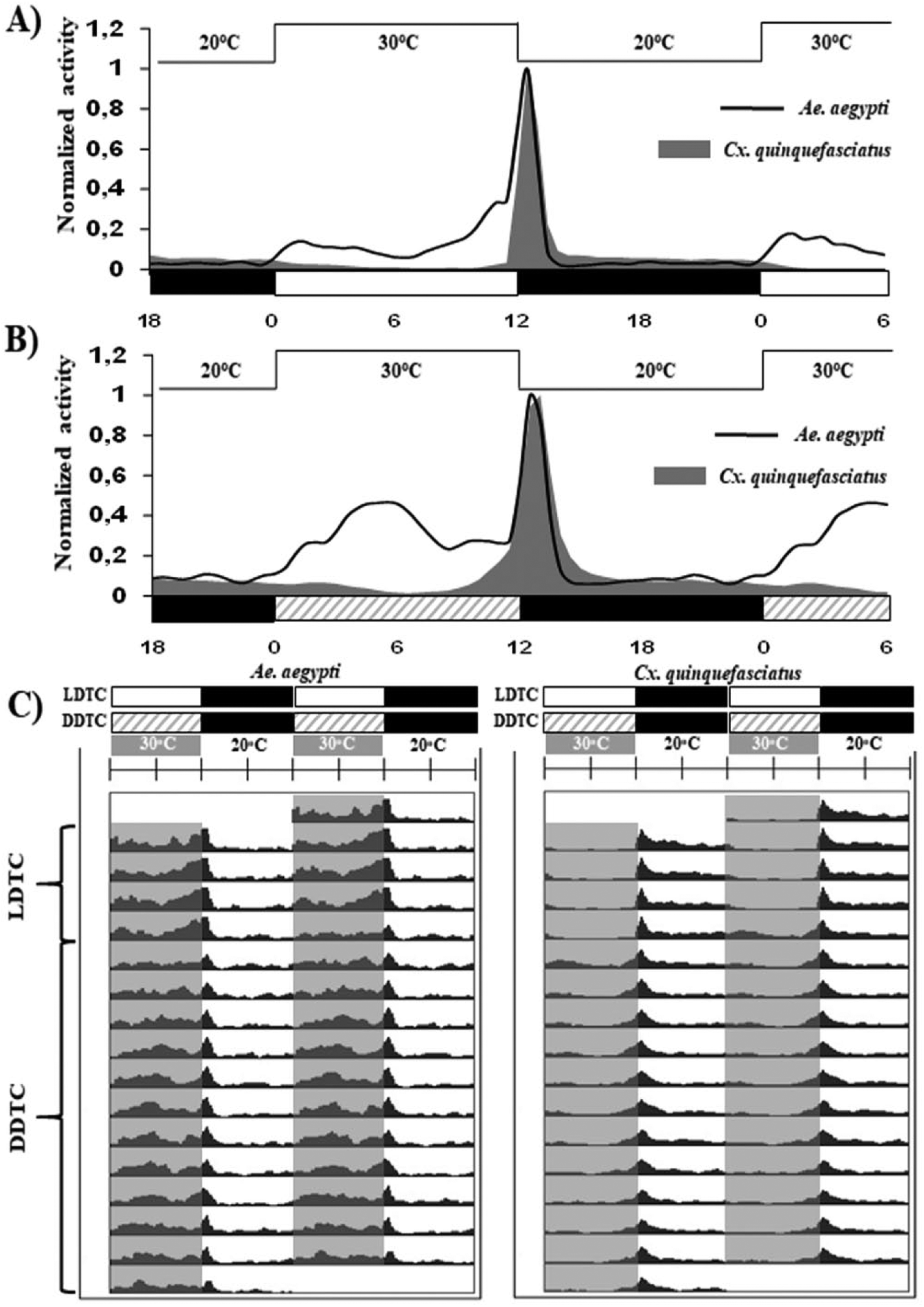
Locomotor activity of mosquitoes under 2 temperature cycle regimens. Displayed here are the average activity profiles of Aedes aegypti (black lines) and Culex quinquefasciatus (gray area) under (A) light-dark with temperature cycles (LDTC) and (B) constant darkness with temperature cycles (DDTC). In all conditions, the average activity profiles were normalized (the maximum was set to 1). (C) Average actograms of A. aegypti and C. quinquefasciatus. Both species were kept in LDTC for the first 4 days, and then the light-dark cycle was removed (DDTC) for 12 days. Bars below the average activities indicate the light regime: white indicates lights-on in LD cycles, diagonal gray stripes indicate lights-off in DD cycles (“subjective day”), and black indicates lights-off in LD or DD (“subjective night”). Areas in actograms indicate warm-cold cycles: gray indicates warm phase in LD cycles and white indicates cold phase. In the average activity profiles, the temperature was indicated for each phase. The lines above the graphs in A and B indicate the temperature cycles. The x-axis represents the zeitgeber time.
On the following day (day 5), we removed the light regimen and kept the temperature cycle as the main external cue in order to observe the exclusive effect of the temperature on locomotor activity. In Figure 1C, the evening peak of both species was dampened if compared with the previous regimen. Conversely, the general activity profile of Culex was barely affected in DDTC (constant darkness with temperature cycles of 30/20 °C). Whereas the evening activity was preceded by an anticipatory pattern just a few hours before the main peak, the early activity was strongly inhibited in the presence of light (under LDTC) (Fig. 1B). Without light-dark cycles, the temperature cycle alone was also able to sustain the evening peak phase of A. aegypti, similar to that of Culex; however, the temperature cycles produced midday activity that was not observed in the LDTC condition (Fig. 1A). This activity suggested a transient pattern during the first 6 days of the regimen that steadied in the final days of the experiment (Fig. 1C).
Daily Rhythms of Locomotion under Conflicting Light-dark and Temperature Cycles
We evaluated how the presence of 2 zeitgebers (light and temperature) in opposite phases affected the daily locomotion of both species. To synchronize the mosquitoes’ activities, we applied a light-dark regimen and temperature cycles in phase (LDTC: light at 30 °C and dark at 20 °C) for 5 consecutive days (Fig. 2). During the final 7 days of the experiment, the temperature cycle was phase-shifted by 12 h (LDCT: light at 20 °C and dark at 30 °C) (Fig. 2C). Both species promptly modified their activity in response to the temperature phase shift, but the responsiveness was species-specific (Fig. 2). The proportions of diurnal activity in A. aegypti (LDTC, 73.17%; LDCT, 38.5%) and nocturnal activity in C. quinquefasciatus (LDTC, 62.57%; LDCT, 54.03%) were significantly affected by both regimens (A. aegypti: LDTC vs. LDCT, t92 = 11.65, p < 0.0001; C. quinquefasciatus: LDTC vs. LDCT, t72 = 2.378, p < 0.05) (Fig. 2). However, these differences were more pronounced in A. aegypti, since most of their daily activity changed from the photophase to scotophase, during the LDCT regime. Moreover, Aedes’s main activity peak was observed at ZT 0 instead of ZT 12 (Fig. 2B). Culex’s evening peak remained consistent during the LDCT regime but was strongly diminished, which suggests that when the temperature cycle is the opposite of the light-dark cycle, it affects the activity levels more so than the temporal niche.
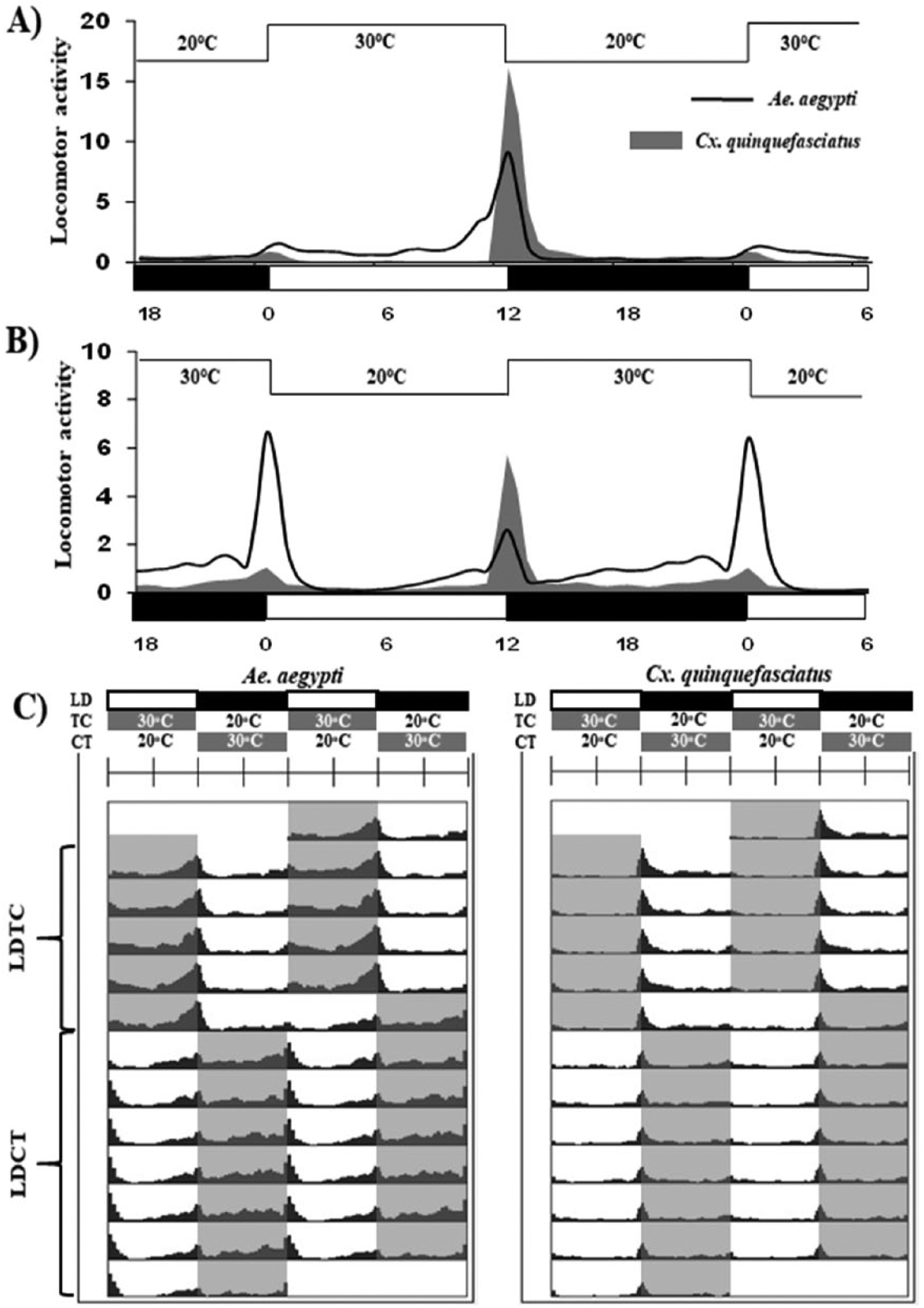
Locomotor activity of mosquitoes under reinforcing and conflicting regimens of light-dark and temperature cycles. Average activity profiles of Aedes aegypti (black lines) and Culex quinquefasciatus (gray area) under (A) reinforcing light-dark and temperature cycles (LDTC) and (B) conflicting light-dark and temperature cycles (LDCT). To normalize the average activity profiles, the maximum was set to 1. (C) Average actograms of A. aegypti and C. quinquefasciatus. First, the mosquitoes were entrained to LDTC for 5 days, and then the temperature cycles were shifted by 12 h and maintained in the “out of phase” condition (LDCT) for 7 days. Areas in actograms or bars below the average activity profiles indicate the temperature cycles: gray indicates 30 °C, white indicates 20 °C. In average activity profiles, the horizontal bars indicate light-dark cycles: white indicates lights-on and black indicates lights-off. The lines above graphs in A and B indicate the temperature cycles. The x-axis represents the zeitgeber time.
Effects of Constant Light on Mosquito Temperature Entrainment
It is expected that Culex, a nocturnal species, is more sensitive to light exposure than Aedes. Therefore, light avoidance could interfere with Culex’s temperature entrainment. We evaluated how constant light altered the locomotion of both species when the species were entrained by temperature cycles. It was important to investigate how much light intensity was necessary in order to inhibit activity and whether the chosen temperature cycle optimally maintained rhythmicity under these conditions. We conducted our experiments using a light intensity of 1000 lux, which is considered a bright light that sufficiently produces arrhythmic behavior in many organisms. Certainly, this could be an interesting strategy to test the power of the temperature zeitgeber on A. aegypti and C. quinquefasciatus. Under this LLTC regime, both species displayed locomotor rhythmicity using the temperature oscillations to restrict activity to the appropriate time of day, given that Aedes is more active during the warm phase and Culex exhibits more activity under low temperatures (Suppl. Fig. S1). As expected, both species became completely arrhythmic soon after removal of the temperature oscillation in the subsequent days (Suppl. Fig. S1).
Aedes specimens exhibited low activity levels during the cold phase (ZT 12-0) under this regimen (LLTC), which corresponded to the scotophase of light and dark cycles (LD). In turn, Culex tended to avoid the warm phase (ZT 0-12), which corresponded to the photophase of LD cycles, exhibiting a light avoidance behavior. To evaluate how light responsiveness could affect the temperature synchronization of Aedes and Culex, we decided to subject the insects to 2 independent experiments: (1) 6 days of LLTC entrainment, followed by 10 days of DD/20 °C; and (2) 6 days of DDTC entrainment, followed by 10 days in DD/20 °C (Fig. 3). After the insects spent 10 days in DD/20 °C, we measured their free-running periods and compared the results according to the previous entrainment regimens.
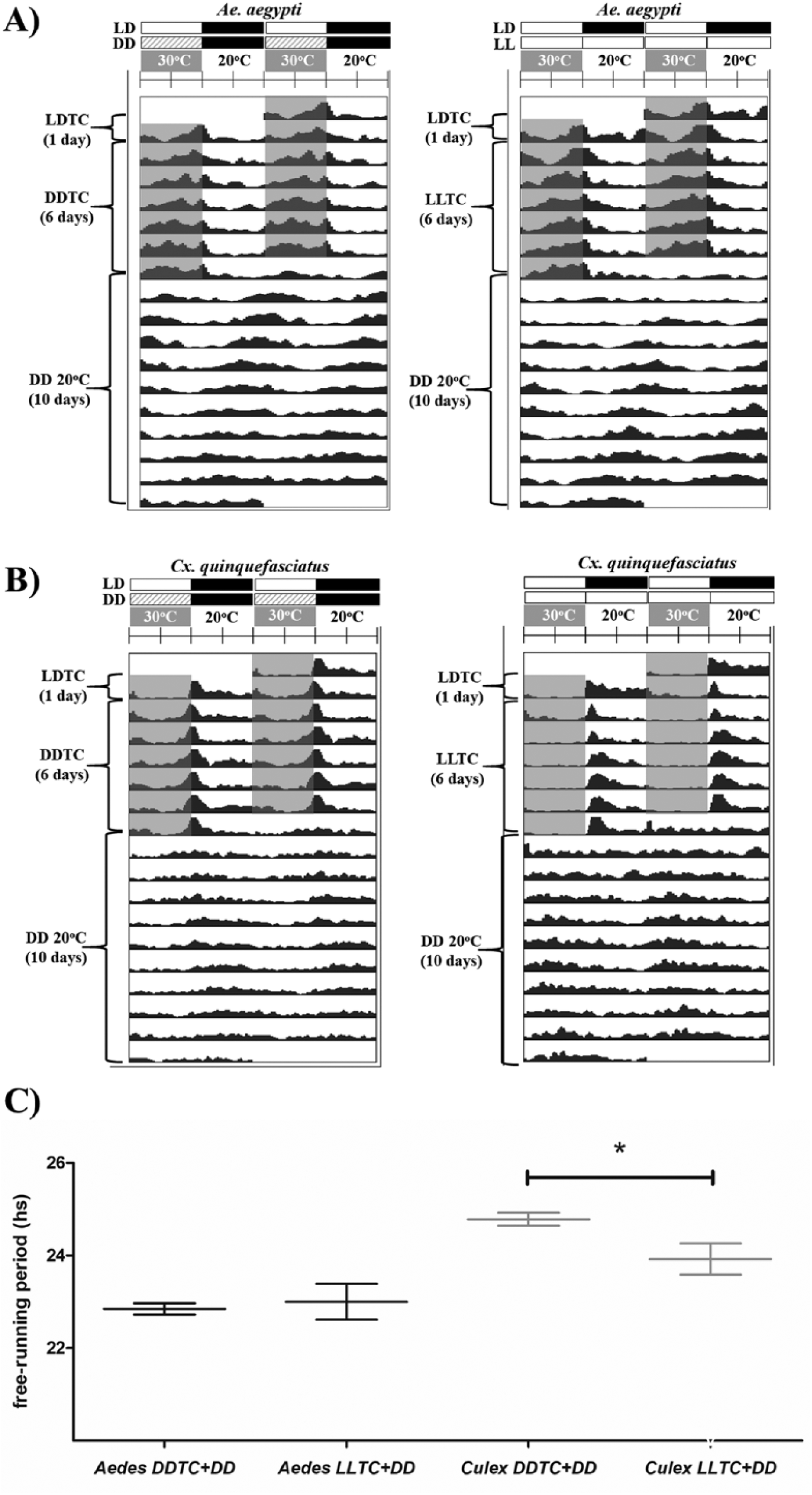
Free-running period of mosquitoes under constant darkness conditions, previously entrained by different photoperiodic regimes. Mosquitoes were entrained for 6 days of temperature cycles, which differed based on the absence or presence of constant light (DDTC and LLTC, respectively). Actograms represent the 17 days of the experiment under different artificial regimes of entrainment before the estimation of the free-running period under DD, at a constant 20 °C. (A) Aedes aegypti. (B) Culex quinquefasciatus. (C) Free running period length of activity in A. aegypti and C. quinquefasciatus was estimated during 10 days of experiment under DD/20 °C, after the first days of temperature entrainment with or without constant light. The asterisk marks a statistical difference detected by the Mann-Whitney U test (p < 0.05). The shaded area of all actograms represents the warm phase (30 °C) applied in this experiment.
Daily locomotion notably decreased when both species were under DD/20 °C, compared with the previous entrainment temperature cycles (DDTC and LLTC). Nevertheless, the activity under DD/20 °C was rhythmic throughout the experiment. Under temperature cycles and constant darkness (DDTC), both species exhibited the expected values of period length under DD/20 °C (Fig. 3 and Table 1). However, when temperature entrainment was combined with constant light, C. quinquefasciatus was more so affected, evidenced by the different period lengths when the 2 independent regimes (DDTC and LLTC) are compared (Fig. 3C and Table 1). Although A. aegypti mosquitoes did not show significant differences in period lengths, the presence of constant light during the entrainment days seemed to reduce the power levels under DD/20 °C, as observed in Culex (Table 1).
Free-running period of mosquitoes under constant darkness and temperature conditions, previously entrained by different photoperiodic regimes.
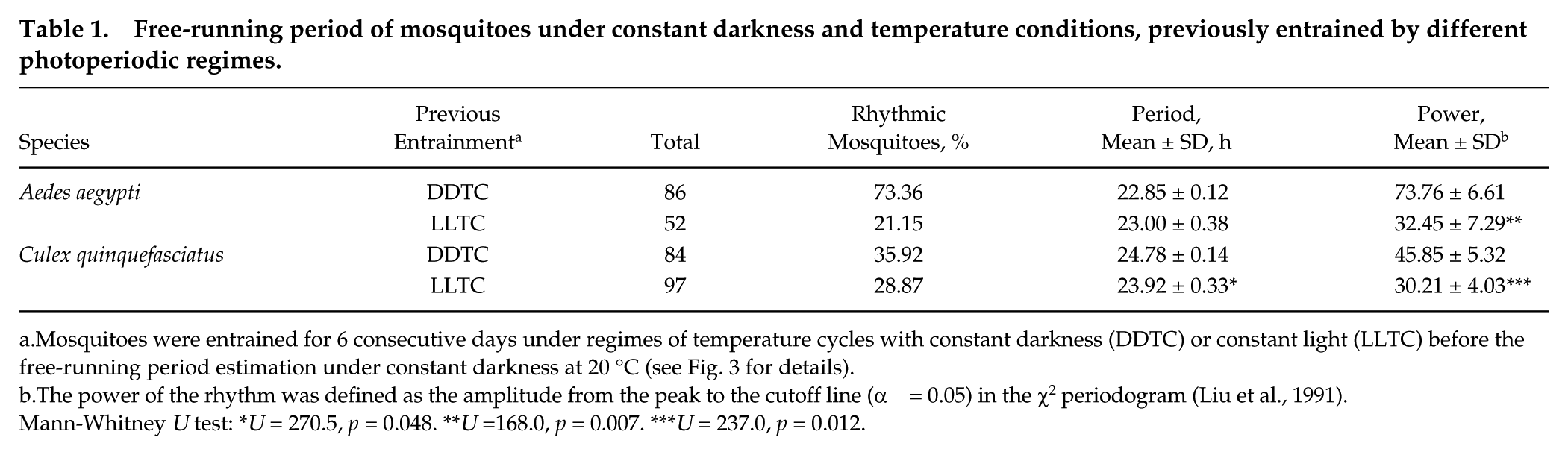
Mosquitoes were entrained for 6 consecutive days under regimes of temperature cycles with constant darkness (DDTC) or constant light (LLTC) before the free-running period estimation under constant darkness at 20 °C (see Fig. 3 for details).
The power of the rhythm was defined as the amplitude from the peak to the cutoff line (α = 0.05) in the χ2 periodogram (Liu et al., 1991).
Mann-Whitney U test: *U = 270.5, p = 0.048. **U =168.0, p = 0.007. ***U = 237.0, p = 0.012.
Clock Gene Expression Analysis under Different Light and Temperature Regimens
We previously observed distinct patterns of locomotion in A. aegypti and C. quinquefasciatus under LDCT and DDTC, compared with LD and DD regimes (Gentile et al., 2009). We suggested that the expression patterns of most canonical clock genes were conserved in both species, but the differences shown in cry2 expression could be crucial to sustaining different activity rhythms (Gentile et al., 2009). Interesting, although not significant, tim expression was different under LD and DD, which might suggest that genes other than cry2 display interspecific expression differences. Therefore, we decided to (a) evaluate the expression patterns of these orthologous clock genes using the regimens of the behavioral experiments, which could unveil some differences between these two species not previously observed under LD and DD, and (b) investigate whether the observed behavioral responsiveness was a consequence of the proportional modulation of clock gene expression.
Table 2 displays the statistical analyses of gene expression of per, tim, Clk, cyc, vri, Pdp1, and cry2. Most genes exhibited significant rhythmicity under the various regimens (Table 2). When gene expression is compared in a heat map form, the organization of gene clustering varied by regime (Suppl. Fig. S2). For instance, under LDTC, most of the orthologous genes clustered together (e.g., Ae_per and C_per) (Suppl. Fig. S2). However, under various artificial conditions (DDTC, LDCT, or LLTC), only some groups of orthologous genes, such as per, vri, and cyc, remained clustered. Under LDTC, for example, cry2 orthologs were clustered, and tim orthologs were not clustered, since Ae_tim expression peaked earlier in this species. To evaluate any putative differences observed in the heat maps, we superimposed the graphs of relative abundance of each gene for the interspecific comparison of mRNA expression (Fig. 4). Under LDTC, the expression of per was conserved in both species. As shown in the heat maps, per expression reached higher levels just after vri and tim in both species (tim and vri, ZT 9-13; per, ZT 13-19) (Figs. 4 and 5), while Ae_cyc and Ae_Clk reached coincident peaks in the early morning (ZT 1-7). Pdp1 mRNA abundance had a rhythmic pattern, and its level varied from the constant low of the photo-warm phase to high levels throughout most of the scoto-cold phase.
Statistical analysis of the circadian expression of clock genes in Aedes aegypti and Culex quinquefasciatus.
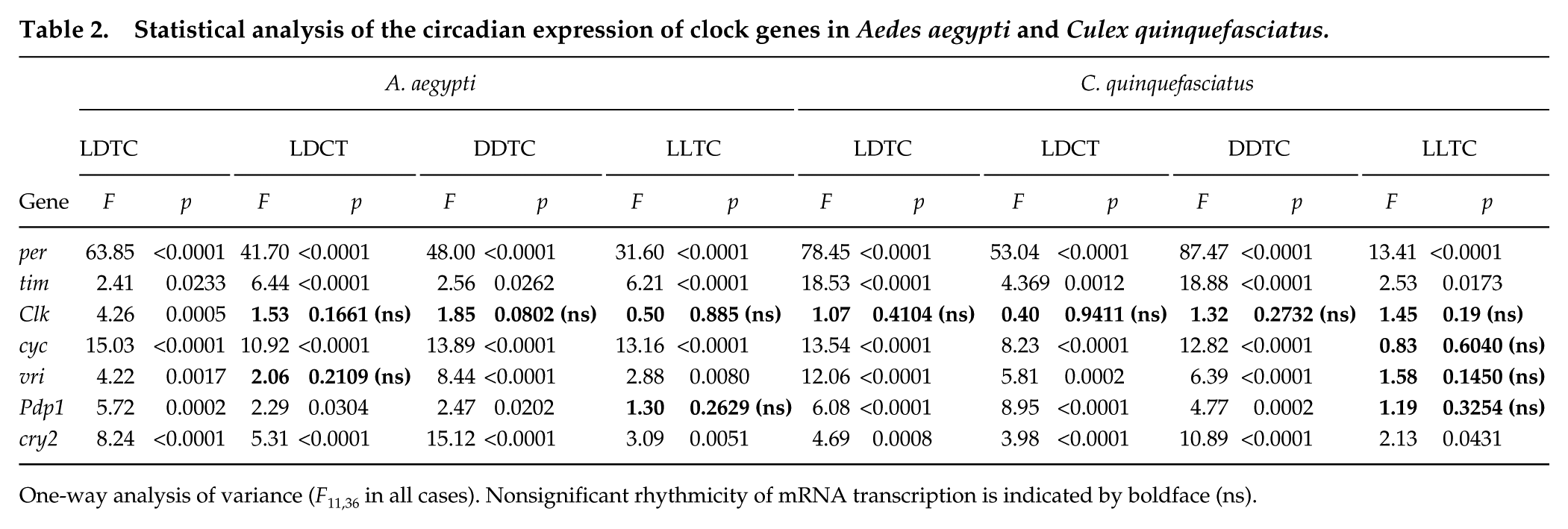
One-way analysis of variance (F11,36 in all cases). Nonsignificant rhythmicity of mRNA transcription is indicated by boldface (ns).
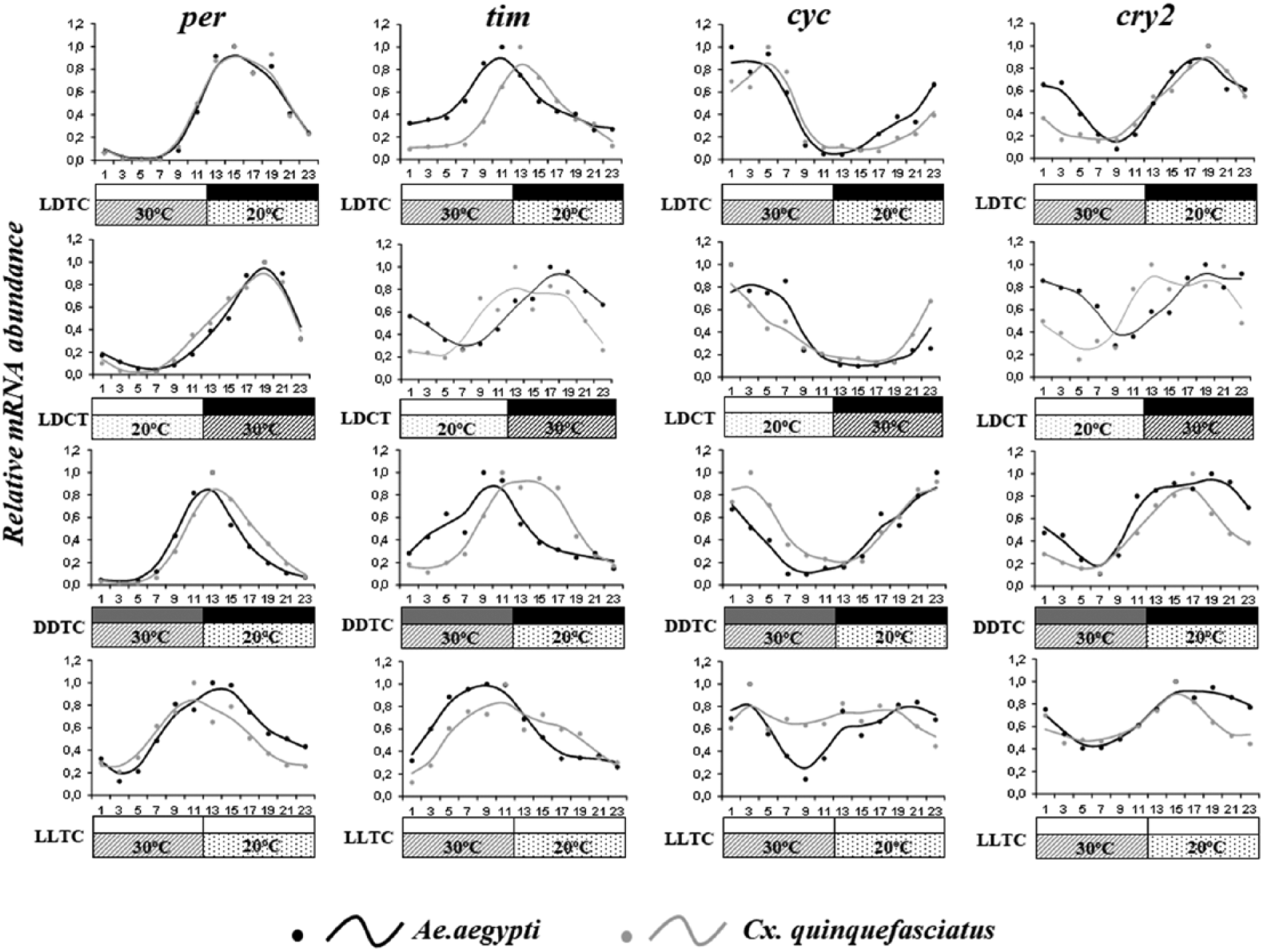
Circadian expression of clock genes in mosquitoes under different artificial regimes. The graphs show the expression of period (per), timeless (tim), cycle (cyc), and cryptochrome 2 (cry 2) in Aedes aegypti (black lines and dots) and Culex quinquefasciatus (gray lines and dots) under 4 different regimes, using qRT-PCR. The analysis was restricted to female heads in reinforcing light-dark and temperature cycles (LDTC), conflicting light-dark and temperature cycles (LDCT), as well as temperature cycles with constant darkness (DDTC) or constant light (LLTC). The bars below the graphs indicate different light conditions: white indicates lights-on, gray indicates “subjective day” in the DD cycle, and black indicates lights-off. Dashed bars indicate temperature. Dots depict the average value of each time point, and the line curves were obtained through a 3-point weighted moving average of these values (weight of 2 for the central time point). For clarity, error bars are not shown. The x-axis represents the zeitgeber time.
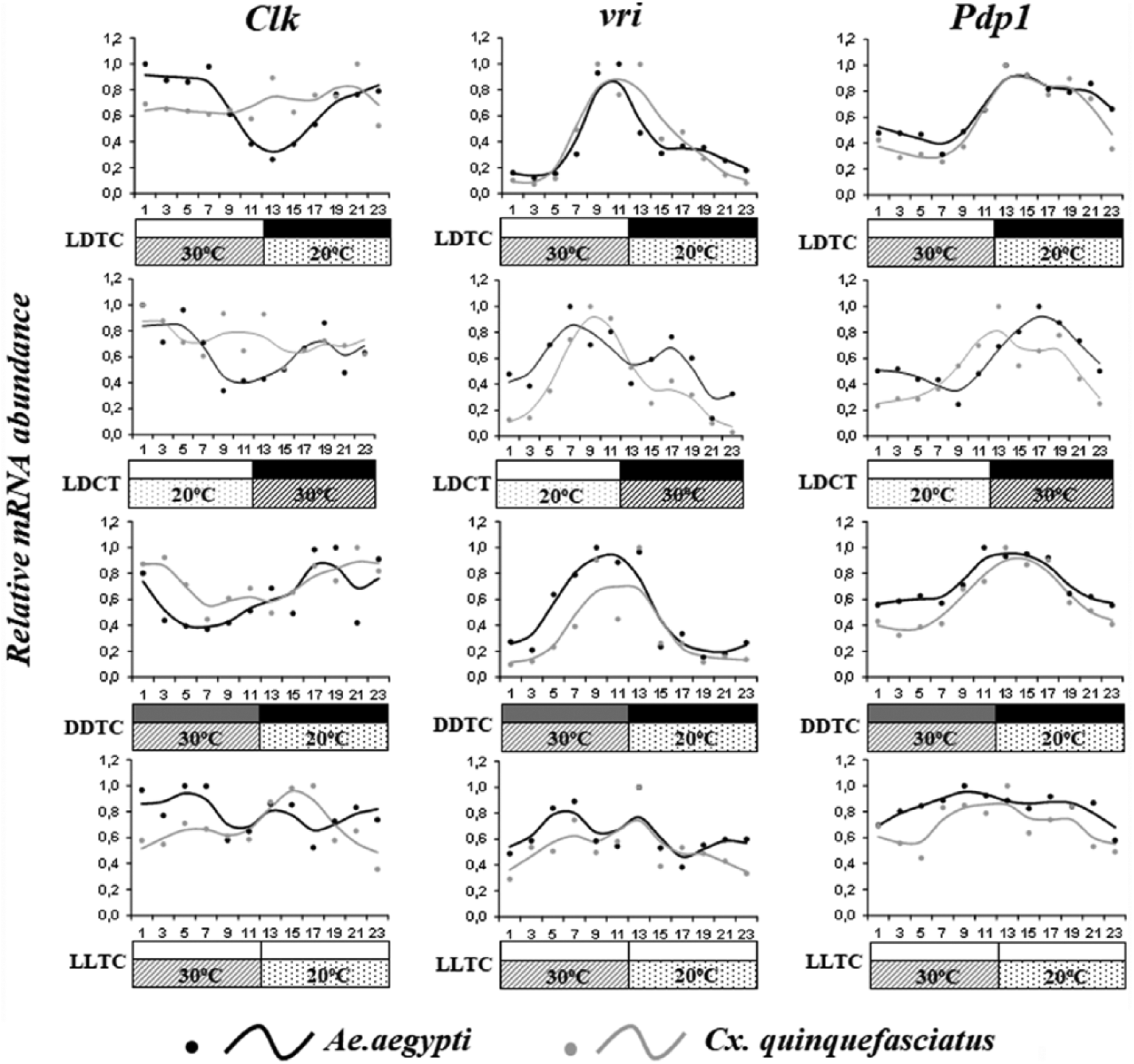
Circadian expression of Clk, vri, and Pdp1 in mosquitoes under different artificial regimes. The graphs show the expression of Clock (Clk), vrille (vri), and Par-domain-protein-1 (Pdp1) in Aedes aegypti (black lines and dots) and Culex quinquefasciatus (gray lines and dots) female heads. The mosquitoes were kept under same conditions described in Figure 4.
Besides the similar profiles of observed clock gene expression, per orthologues were similarly entrained by temperature in both mosquito species and all regimens used. Under LDCT, Ae_per and C_per had an mRNA peak later than under LDTC (ZT 17-21) and earlier when the temperature cycle was the only zeitgeber used (DDTC and LLTC; ZT 11-15). However, vri, Pdp1, and cycle expression patterns that were conserved among their orthologues under LDTC displayed slight differences between species under LDCT (Figs. 4 and 5). Under DDTC, the mRNA abundance of cyc showed a different expression pattern between species in the warm phase (ZT 3, ZT 5, ZT 7 U = 0.0, p < 0.05). Finally, C_cyc was surprisingly arrhythmic under LLTC (Fig. 4 and Table 2). Indeed, the presence of constant light affected the amplitude of gene expression and rhythmicity in Culex, since vri and Pdp1 were also arrhythmic under this regimen (Fig. 4 and Table 2).
In contrast to per, vri, Pdp1, and cyc expression under the LDTC regime, tim, Clk, and cry2 had dissimilar expression profiles between A. aegypti and C. quinquefasciatus (Figs. 4 and 5). We observed that Ae_Clk mRNA was cycling while C_Clk was arrhythmic under LDTC (Fig. 5). However, both species’ Clk expression was considerably affected when other regimes were used.
Under LDTC, Ae_tim exhibited a peak expression 2 h earlier than C_tim (Fig. 4). Interestingly, when the temperature cycle conflicted with the light-dark condition (LDCT), the mRNA peak of Ae_tim occurred later than the C_tim mRNA peak. The delay of Ae_tim expression reflected the mRNA abundance in the early morning, as observed by its increased levels at this time (ZT 1: U = 0.0, p < 0.05). Under DDTC, tim displayed a pattern of expression similar to LDTC, although the anticipation pattern of Ae_tim was more pronounced than C_tim (ZT 15-17: U = 4.0, p = 0.0019). Under constant light conditions (LLTC), temporal distinctions of tim peak expression were indistinguishable, and rhythmicity was maintained in both species (Table 2).
Under the LDTC regimen, Ae_cry2 mRNA expression displayed a bimodal pattern (ZT 3: U = 0.0, p < 0.05) (Fig. 4) similar to the pattern previously observed in LD and DD at 25 °C (Gentile et al., 2009). When the LDCT regimen was applied, we noted remarkable differences between Ae_cry2 and C_cry2 in the photophase (ZT 3: U = 1.0, p = 0.057; ZT 5: U = 0.0, p = 0.029; ZT 7: U = 0.0, p = 0.029). During the scotophase, C_cry2 mRNA expression seemed to reach the plateau at ZT13, while Ae_cry2 mRNA expression reached this plateau later, at ZT 19 (Fig. 4). Moreover, under DDTC and LLTC, when temperature was the only zeitgeber tested, we observed an increase of Ae_cry2 expression levels during the last portion of the cold phase in anticipation of the morning and evening peaks (Fig. 4). Under LDTC, DDTC, and LLTC, C_cry2 had a more accurate mRNA peak than Ae_cry2 when compared with the LDTC regimen (LDTC, ZT 19; DDTC, ZT 17; LLTC, ZT15). This pattern was also observed in the expression of per and tim genes, based on not only the mRNA peak but also the onset of mRNA abundance.
Interspecific Temperature Entrainment Driving Clock Gene Expression
Most orthologous mRNA profiles of both species clustered together on the heat maps; however, this pattern varied by applied regimen (Suppl. Fig. S2). These results were the first clue of a differential pattern of temperature entrainment between these species. In fact, when the expression of some orthologues was observed under LDCT, the mRNA abundance profiles varied slightly (Fig. 4). We further investigated these differences by observing how these two species were entrained by the temperature zeitgeber’s phase (LDTC or LDCT). We did not use the Clk and vri genes in this analysis because they did not cycle under LDCT (Table 2).
Figure 6 shows that both species modified their expression patterns according to the temperature cycle phases. The expression of per in both species was phase-delayed in LDCT compared with LDTC. This pattern was also observed in tim and Pdp1 but was more noticeable in Ae_tim; C_Pdp1 was barely affected by the temperature cycle inversion (Fig. 6). cycle and cry2 showed unique patterns of entrainment, because these genes were phase-advanced under LDTC in Culex but not in Aedes orthologous genes (Fig. 6). The comparison of both regimens demonstrates that Ae_cyc expression was slightly altered, but its mRNA peak was not considerably affected by the temperature phase shift. In contrast, Ae_cry2 mRNA levels were reduced under LDCT.
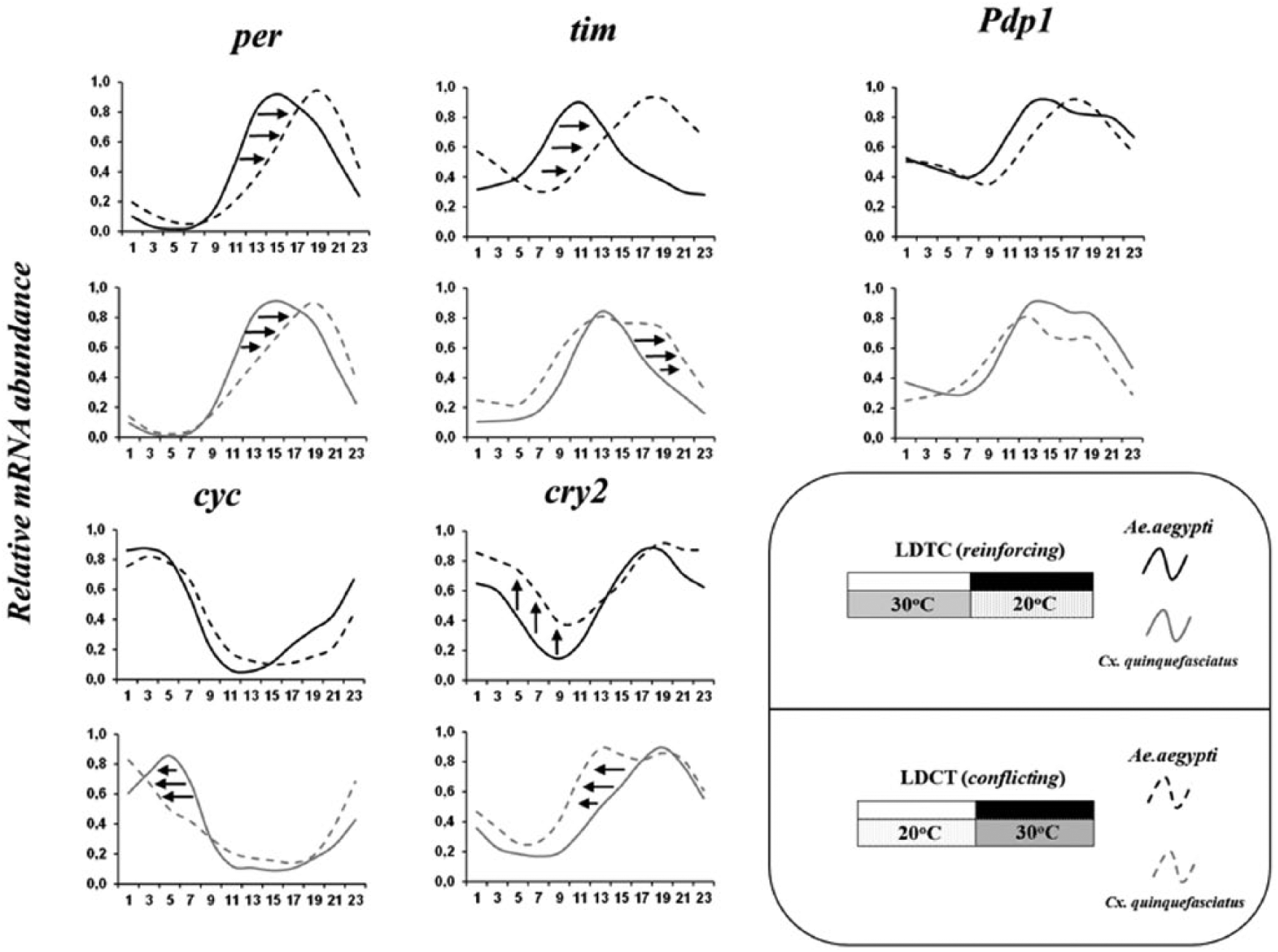
Modification of clock gene expression in mosquitoes under reinforcing or conflicting zeitgebers. Expression of per, tim, Pdp1, cyc, and cry2 in Aedes aegypti (black lines) and Culex quinquefasciatus (gray lines) female heads under reinforcing light-dark and temperature cycles (LDTC, continuous lines) or conflicting cycles (LDCT, dashed lines). Arrows indicate the direction of phase shift in LDCT compared with LDTC. The box shows a schematic representation of LDTC and LDCT regimes and of mosquito species graph lines.
Discussion
Herein, we analyzed the influence of temperature cycles on the daily activity rhythms and clock gene expression patterns of two mosquito species that exhibited distinct daily habits in constant temperatures (Gentile et al., 2009). We used these specimens in a series of artificial regimens that mimicked the daily environmental oscillations of light-dark and temperature in order to evaluate the patterns of locomotion and expression of clock genes. One of the most salient features observed was the unique daily activity pattern of each species when entrained by light-dark and temperature cycles (Figs. 1 and 2). A. aegypti changed its pattern of activity considerably, while C. quinquefasciatus exhibited only minor alterations in activity. Our findings suggest that the daily activity of A. aegypti is more influenced by temperature, whereas C. quinquefasciatus activity is strongly influenced by light sensitivity (Fig. 2). The Culex activity patterns under 12 h of misalignment between light and temperature (LDCT) resemble the profiles observed in fruit flies (Yoshii et al., 2010; Harper et al., 2016) and in kissing bugs (Lazzari, 1992), which tended to follow the light-dark cycles. On the contrary, the Aedes activity pattern under LDCT was unexpected and different than most insect species. Future studies using semi-natural light and temperature conditions can contribute to a more comprehensive understanding of mosquitoes’ circadian clocks.
Interspecific patterns of entrainment and light-temperature responsiveness were also observed in the mRNA levels of cry2, tim, Clk, and cyc. These results support the previous findings of Gentile et al. (2009); most clock gene expression profiles in these two species have been preserved throughout evolution, with the exception of cry2 (a bimodal pattern in Aedes vs. a unimodal pattern in Culex). These observations show how the mosquito’s behavior resonates with the expression of most clock genes to become synchronized to environmental surroundings.
In nature, multiple environmental factors in addition to light-dark and temperature simultaneously influence the endogenous clock. Social interactions, olfaction, feeding, and sexual volatile attractors are important cues that dictate behavior and modify the circadian expression of genes (Saunders, 2002; Clements, 1999). Natural selection has shaped the genomes of organisms to optimally adapt their daily rhythms. These genomic signatures can be found not only in canonical clock genes but likely in several other genes responsible for behavioral control as well. A comparison of the circadian transcriptomes of A. aegypti and the nocturnal species Anopheles gambiae has revealed temporally distinct expression of olfaction and vision genes (Rund et al., 2013).
In terms of vision, the eye structures of diurnal and nocturnal mosquitoes are distinct (Clements, 1999). Accordingly, different rhodopsins are expressed in different eye regions of A. aegypti and C. quinquefasciatus (Hu et al., 2009). These singularities are adaptive traits to the different light intensities that each species faces daily. The effect of light input is also distinct for each species’ habits and corresponding light responsiveness. As mentioned earlier, the resetting of the clock by light is dependent on TIM degradation regulated by CRY1 (reviewed in Hardin, 2011). Here, we observed a temporal difference in mRNA levels between tim orthologues when temperature was the only zeitgeber and when both zeitgebers were combined. We believe that this difference in tim expression could affect the temporal abundance of TIM protein in both species. When combined with interspecific signals of light input, it could reinforce the divergence of light entrainment between Aedes and Culex. Under conditions of bright light intensity, Culex should immediately respond by decreasing its activity, and the endogenous responsiveness of this effect may be caused by a proportional modulation of circadian gene expression. In fact, some of our results support this hypothesis, since C. quinquefasciatus had more genes that did not cycle under constant light compared with A. aegypti (Table 2, Fig. 4). These findings suggest that the transcription of these genes could be more responsive to light entrainment than that of the other clock genes.
Even so, we can still observe activity rhythms in Culex under LLTC. In this scenario, we suggest that the remaining activity under this regime is either driven by the circadian transcription of other genes (e.g., per, tim, and cry2) or a consequence of circadian protein activation (phosphorylation), regardless of whether the respective mRNA is cycling. Alternatively, it is known that light and temperature can directly influence rhythmicity independent of the molecular clock in fruit flies (Boothroyd et al., 2007). Activity directly affected by external cues, known as a “masking effect,” is important for the adjustment of daily activity (Mrosovsky, 1999). Although we cannot disregard the direct influence of temperature on output genes, we believe that most of the activity exhibited under LLTC is indeed a consequence of the expression of clock genes that normally occurs in Drosophila and mosquitoes under the same regimens. In this case, Drosophila sustains the circadian expression of the core genes as well as activity rhythms and therefore displays the unimodal pattern of activity (Glaser and Stanewsky, 2005) that occurred here in mosquitoes (Suppl. Fig. S1).
Gentile et al. (2009) previously observed that both cry2 orthologous gene rhythms in A. aegypti and C. quinquefasciatus shared a daily evening peak, but Ae_cry2 also had a morning peak that was not observed in C_cry2. Moreover, the transcriptome analyses of A. aegypti and A. gambiae by Rund et al. (2013) also suggested dissimilarities in the expression of cry2 orthologous genes, although this difference was not mentioned in the study. Here, we confirmed the bimodal pattern of expression in Ae_cry2 not only in the presence of temperature cycles but also when combined with light-dark cycles (Fig. 4). Under DDTC, the morning peak phase-shifted to the late hours of the subjective night (at 20 °C), which in turn made it difficult to detect the evening peak, which did not occur in C_cry2. The higher cry2 expression of diurnal mosquitoes in the early morning could be similar to the role of cry1 expression in mating isolation of sibling species of tephritid fruit flies that varied in their time of mating (day or dusk) (An et al., 2004). In the morning, cry1 expression was higher in “day mating” strains compared with “dusk mating” strains. We observed here a similar pattern in cry2 expression when comparing diurnal and nocturnal mosquitoes, which could be important for modulating the circadian transcription of output genes and the arousal of activity at different times. We did not evaluate cry1 expression here because previous reports on mosquitoes have already observed that their mRNA expression is arrhythmic in the head (Gentile et al., 2009; Rund et al., 2011). Moreover, a quick search for cry1 expression profiles in the database generated by Leming et al (2014) did not show any evidence of rhythmicity under LD and DD conditions (https://www3.nd.edu/~bioclock/aedes.html).
While cry1 expression in the mosquito’s head is probably not influenced by temperature since its mRNA does not change throughout the day, we cannot exclude the hypothesis that both cryptochromes may have similar roles in the modulation of daily activity, since cry1 can act as a repressor in the periphery of the clock in fruit flies (Collins et al., 2006). In the tephritid fruit fly, cry1 is constantly expressed in the head, but it shows rhythmicity in the brain and in antennae (An et al., 2004). Thus, it is possible that cry1 is cycling in the mosquito’s brain but not in the whole head. Except for Ae_Clk under LDTC, Clk is not cycling in either species (Table 2). We previously had similar difficulties detecting the rhythmicity of Clk mRNA levels in A. aegytpi and C. quinquefasciatus (Gentile et al., 2009). However, rhythmic mRNA levels of Clk were detected in A. gambiae by RT-qPCR but not by microarray analysis (Rund et al., 2011). Therefore, the detection of rhythmic Clk and cry1 mRNA expression would probably be circumvented by qPCR or brain in situ hybridization. Despite the limiting autofluorescence factor of A. aegytpi tissues (Zucker, 2006), our group successfully detected cyc and per circadian mRNA expression in the brain using nonfluorescent in situ hybridization assays (Chahad-Ehlers et al., 2013). Thus, further studies in this direction could establish a better understanding of clock gene expression in the central pacemaker of mosquitoes.
Most genes studied here in the head of mosquitoes displayed an expression profile that was modified by the presence of one zeitgeber or by the simultaneous effect of light and temperature. For instance, cyc, Pdp1, cry2, and per displayed the anticipated expression peaks under conditions when the temperature was the only zeitgeber used (DDTC and LLTC). More specifically, we observed an expression peak in per around ZT 13-15 under LDTC, while the peak occurred about 2 h earlier under DDTC and LLTC (Fig. 4). This anticipated pattern also occurred in cyc, Pdp1, and cry2, but it was more discrete. The locomotor activity of A. aegypti also followed the pattern of gene expression under DDTC and LLTC represented by an arousal of activity during the warm phase, which was contrary to the LDTC results (Fig. 1). The reason why C. quinquefasciatus did not exhibit significant modifications of activity is too complex to be explained by our results. The strong light avoidance may have masked the activity following entrainment of clock gene expression in this species. However, it is notable that clock gene expression in Culex can be influenced by light-dark and temperature cycles (Figs. 4-6).
In Drosophila, changes in clock gene expression by temperature in only a few subsets of neurons are sufficient to change the locomotor activity regardless of the phase of expression of other areas of the fly head (Yoshii et al., 2010). Thus, the relationship between patterns of clock gene expression in the whole head and the modulation of activity is too complex to be established with accuracy. Future experiments using in situ hybridization and immunocytochemistry in the brain will clarify this question. Nevertheless, our results provide evidence of the putative expression profiles of these areas in the mosquito brain. For instance, under a regimen of conflicting zeitgebers (LDCT), both species’ gene expression patterns were altered. When we compared the patterns of both species during reinforcing and conflicting regimens (LDTC and LDCT), the direction of transcriptional entrainment was sometimes the complete opposite (Fig. 6). This result, despite the fact that we used whole mosquito heads to detect changes of clock gene expression due to light or temperature, offers evidence of what is taking place in some areas of the brain that control mosquito activity. We believe that the patterns of clock gene expression observed here did not occur by chance. Instead, these findings reflect putative modifications in certain areas of the mosquito brain that influence locomotor activity.
Despite recent advances in our understanding of the mosquito clock machinery (Yuan et al., 2007, Gentile et al., 2009), many questions remain. One intriguing question is the role of cry2 in mosquitoes, which is also known to act as a repressor in many insects, including mosquitoes (Yuan et al., 2007). Our results give some insights into the role of cry2 in mosquitoes. For example, cyc and cry2 mRNA levels in Culex display unique patterns of entrainment and exhibit higher levels earlier under LDCT than under LDTC. Considering that the cry2 gene in mosquitoes has E-box motifs (CACGTG) in its promoter regions (Gentile et al., 2009), it is reasonable to speculate that the circadian transcription of cry2 could be influenced by cyc expression. In Drosophila, CLK/CYC can dictate the expression of several genes by its interaction with the E-box region (Hardin, 2011). Our results show that Ae_cry2 and Ae_cyc are unaltered in their evening and morning peak phases, respectively. However, we also observed a reduction of cry2 amplitude. One possible reason for this pattern is the expression observed in vri. Previously found were putative V/P-boxes in the promoter region of Ae_cry2, which are motifs that VRI and PDP1 bind to regulate transcription (Cyran et al., 2003). In A. aegypti, the lower amplitude of cry2 expression under LDCT seems to follow vri, which does not cycle significantly in this regime (Table 2). Therefore, our results represent how some core clock genes can influence the expression profiles of other canonical genes. These findings support the general idea that these genes and their corresponding proteins are interconnected, as is the case in Drosophila.
We showed here interesting examples of how environmental parameters modify daily activity and clock gene expression patterns of mosquitoes as well as how behavioral responsiveness to external cues is proportional to clock gene transcriptional modulation. At the transcriptional level, clock genes have often altered their phase or amplitude of expression, which in turn can influence the transcription of output genes and daily activity. Recent fruit fly studies under semi-natural conditions have unveiled new paradigms about the importance of the endogenous clock dictating activity in nature and the strong influence of temperature entrainment in the field (and in contrast to laboratory conditions) (Vanin et al., 2012; Menegazzi et al., 2012, 2013; De et al., 2013; Green et al., 2015).
Although much of what is known about synchronization with light-dark and temperature cycles is based on Drosophila, recent studies have also described the molecular mechanisms of synchronization in other organisms. For example, Dunlap and Loros (2017) reviewed the Neurospora clock. The synchronization of these fungi with light-dark and temperature cycles occurred mainly through mechanisms that regulate the expression of the gene frequency (frq). This gene exhibited temperature-dependent alternative splicing, similar to that of per and tim in Drosophila (Majercak et al., 1999; Majercak et al., 2004, Collins et al., 2004, Boothroyd et al., 2007, Dunlap and Loros, 2017). Interestingly, temperature cycles may have an even greater influence on the Neurospora clock than the light-dark cycle (Dunlap and Loros, 2017). The mechanism of synchronization with temperature cycles dependent on alternative splicing seems not to be restricted to Neurospora and Drosophila, as it has been found in some mammalian tissues (Gotic and Schibler, 2017). Thus, we believe that future studies focused on the investigation of possible alternative splicing in mosquito clock genes, as well as the analysis of the influence of temperature cycles on the different isoforms, may contribute significantly to our understanding of the clock of these vectors.
Our results corroborate classic assumptions about environmental influences on daily mosquito rhythms (reviewed in Clements, 1999) and include innovative findings on the subject. We intend to expand these studies with further analyses under semi-natural conditions to reinforce the findings described here. Moreover, the opportunity to use clock genes as molecular targets in mosquitoes (via RNAi or CRISPR gene editing) could reveal the influence of one single clock gene on the persistence of circadian rhythmicity of other clock genes. Certainly, this strategy would shed new light on the hierarchy of positive and negative elements of the interlocked feedback loops of mosquitoes. As observed here, the expression of some clock genes is more so influenced by light or temperature cycles. Under a regimen of conflicting zeitgebers (LDCT), we observed that A. aegypti is unexpectedly active during the night, while C. quinquefasciatus’s main activity peak was reduced (Fig. 2), which could be a consequence of a divergence in clock gene entrainment of both species (Fig. 6).
Certainly, the unusual locomotion patterns might interfere with several aspects of metabolism, energy expenditure, fitness cost, and pathogen transmission efficiency. Thus, the choice of clock genes and light and temperature receptors as molecular targets should be relevant to chronobiological studies and have epidemiological implications.
Supplemental Material
Figure_S1_-_Rivas_et_al_-_30nov17 – Supplemental material for Effects of Light and Temperature on Daily Activity and Clock Gene Expression in Two Mosquito Disease Vectors
Supplemental material, Figure_S1_-_Rivas_et_al_-_30nov17 for Effects of Light and Temperature on Daily Activity and Clock Gene Expression in Two Mosquito Disease Vectors by Gustavo B. S. Rivas, Rayane Teles-de-Freitas, Márcio G. Pavan, José B. P. Lima, Alexandre A. Peixoto and Rafaela Vieira Bruno in Journal of Biological Rhythms
Supplemental Material
figure_S2_-_Rivas_et_al_-_30nov17 – Supplemental material for Effects of Light and Temperature on Daily Activity and Clock Gene Expression in Two Mosquito Disease Vectors
Supplemental material, figure_S2_-_Rivas_et_al_-_30nov17 for Effects of Light and Temperature on Daily Activity and Clock Gene Expression in Two Mosquito Disease Vectors by Gustavo B. S. Rivas, Rayane Teles-de-Freitas, Márcio G. Pavan, José B. P. Lima, Alexandre A. Peixoto and Rafaela Vieira Bruno in Journal of Biological Rhythms
Footnotes
Acknowledgements
We thank Robson Costa da Silva for his expert technical assistance. G.B.S.R., M.G.P., and R.T.F. received scholarships from the Brazilian funding agencies FAPERJ, CNPq, and CAPES. These funders had no role in the study design, data collection and analysis, decision to publish, or preparation of the manuscript. We dedicate this paper to the memory of Dr. Alexandre Afranio Peixoto, a great mentor and friend. This work was supported by Fundação Carlos Chagas Filho de Amparo à Pesquisa do Estado do Rio de Janeiro (FAPERJ) (grant number E-26/102.274/2010) and Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq) (grant number 401.857/2014-0).
Author Contributions
G.B.S.R., R.V.B., and A.A.P. conceived and designed the study. G.B.S.R., R.T.F., M.G.P., J.B.P.L., and A.A.P. performed and analyzed data. G.B.S.R., R.T..F, M.G.P., and R.V.B. drafted the manuscript. All authors approved the final version of the manuscript.
Conflict of Interest Statement
The author(s) have no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Notes
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.