
Abstract
Short-wavelength light guides many behaviors that are crucial for an insect’s survival. In Drosophila melanogaster, short-wavelength light induces both attraction and avoidance behaviors. How light cues evoke two opposite valences of behavioral responses remains unclear. Here, we comprehensively examine the effects of (1) light intensity, (2) timing of light (duration of exposure, circadian time of day), and (3) phototransduction mechanisms processing light information that determine avoidance versus attraction behavior assayed at high spatiotemporal resolution in Drosophila. External opsin-based photoreceptors signal for attraction behavior in response to low-intensity ultraviolet (UV) light. In contrast, the cell-autonomous neuronal photoreceptors, CRYPTOCHROME (CRY) and RHODOPSIN 7 (RH7), signal avoidance responses to high-intensity UV light. In addition to binary attraction versus avoidance behavioral responses to UV light, flies show distinct clock-dependent spatial preference within a light environment coded by different light input channels.
Introduction
Light guides many important behaviors in insects, including positive phototaxis, circadian entrainment, arousal, and sleep (Miller et al., 1981; Sheeba et al., 2008; Fogle et al., 2015; Garbe et al., 2015; Baik et al., 2017; Baik et al., 2018). Flies display both attraction and avoidance to short-wavelength light. Attraction to light (positive phototaxis) is a well-known behavior. In contrast, avoiding short-wavelength light minimizes desiccation, ultraviolet (UV) light–induced DNA damage, and excessive heat exposure. Adult flies exhibit a circadian-modulated avoidance to chronic high-intensity UV light over a 12-h period that coincides with the peak of light intensity and heat in the middle of the day (Baik et al., 2018). In contrast, low-intensity light evokes immediate attraction to light within minutes of light onset (Heisenberg and Buchner, 1977; Gao et al., 2008; Yamaguchi et al., 2010; Baik et al., 2017). This suggests that UV light–evoked attraction/avoidance responses may depend on intensity and/or duration of exposure. However, it is unclear which phototransduction channels modulate these two opposite behavioral responses to light stimuli and their time-of-day variance in response. The question still remains, how does an animal spatially navigate and choose between different light environments? Here, we comprehensively examine the external and internal factors influencing the animal’s spatiotemporal choices in response to different light environments. External factors include the spectral quality and quantity of light, while internal factors include circadian timing and different phototransduction signaling channels (external opsins in the eyes and other structures and internal neuronal and eye structures CRYPTOCHROME and RHODOPSIN 7) in flies. We examined the contribution(s) of (1) the duration of exposure versus circadian time of day, (2) the intensity, and (3) the different phototransduction mechanisms on UV light–evoked attraction/avoidance behavior.
Both attraction and avoidance in insects are evoked by short-wavelength light (Heisenberg and Buchner, 1977; Miller et al., 1981; Gao et al., 2008; Yamaguchi et al., 2010; Baik et al., 2017; Baik et al., 2018). In Drosophila, short-wavelength light is processed through multiple phototransduction mechanisms: (1) external opsin-based photoreceptors and direct neuronal phototransduction pathways: (a) CRY/HK-based phototransduction and (b) RH7-mediated phototransduction (Feiler et al., 1992; Chou et al., 1996; Salcedo et al., 1999; Fogle et al., 2011; Fogle et al., 2015; Saint-Charles et al., 2016; Ni et al., 2017). All three phototransduction pathways mediate circadian entrainment to light (Emery et al., 1998; Stanewsky et al., 1998; Helfrich-Forster et al., 2001; Helfrich-Forster et al., 2002; Malpel et al., 2002; Rieger et al., 2003; Klarsfeld et al., 2004; Veleri et al., 2007; Sheeba et al., 2008; Kistenpfennig et al., 2012; Schlichting et al., 2014; Schlichting et al., 2015; Saint-Charles et al., 2016; Schlichting et al., 2016; Ni et al., 2017). Both CRY/HK- and RH7-mediated phototransduction evoke a rapid increase in neuronal action potential firing and resting membrane potential (Fogle et al., 2011; Fogle et al., 2015; Baik et al., 2017; Ni et al., 2017). This suggests that the integration of multiple phototransduction inputs may be crucial in coordinating a complex light-evoked behavior, such as avoidance and attraction.
Materials And Methods
Fly Strains
The following fly strains were obtained from Bloomington Stock Center: w1118 (BL5905) and glass60j (BL509). cry01 was from Jeff Hall (Brandeis University). timeless0 and clkOUT were from Joanna Chiu (University of California, Davis). norpAP24 and rh71 were provided by Craig Montell (University of California, Santa Barbara). hk-/- was provided by Ming Zhou (Baylor College of Medicine). glass60j-cry01 double mutant fly line was generated in Baik et al. (2017) by standard recombination genetic crosses of gl60j and cry01 lines.
Standard Light Choice Assay
Standard LD light choice assays were conducted as outlined in Baik et al. (2017). The locomotor activity of individual flies was measured using the TriKinetics Locomotor Activity Monitoring System via infrared beam-crossing, recording total crosses in 1-min bins. Percentage activity and statistics were measured using Microsoft Excel and Sigma Plot. Philips TL-D Blacklight UV source with a narrow peak wavelength of 365 nm and intensity of 400 μW/cm2 was used for high intensity, and 10 μW/cm2 was used for low intensity by using neutral-density filters. To determine acute light responses, 15-min pulses of UV light (365 nm, 400 μW/cm2 for high-intensity pulses or 10 μW/cm2 for low-intensity pulses) were used throughout the 12-h daylight phase at hourly intervals instead of a 12 h:12 h UV light:dark cycle. The experiment was carried out under a constant dark condition (DD), and light pulses were administered only during the subjective light phase in order to maintain the prior circadian entrainment.
Multibeam Light Choice Assay
We adapted the standard LD light choice assay outlined in Baik et al. (2017) using Trikinetics MultiBeam Monitors. Two monitors were aligned parallel to each other, with each tube containing a single fly measured by both monitors simultaneously. One of the two monitors was covered, providing the flies with a choice of a shaded environment (covered monitor) versus UV-exposed environment (uncovered monitor) during the 12 h of UV light in the light phase (ZT0-12). Each of the infrared beams was spaced every 3 mm. By using 2 monitors, each fly had 17 infrared beams on each light-exposed side and on the shaded side, with a total of 34 beams per fly. Locomotor activity, preference percentile, and statistics were measured using Microsoft Excel and Sigma Plot. Data are presented as mean ± SEM (*p < 0.05; **p < 0.01; ***p < 0.001).
Statistics
Data are presented as mean ± SEM. Values of n refer to the total number of tested flies. In all cases, the n values were obtained from at least three separate experiments. All statistical tests were performed with SigmaPlot 11. For comparisons of two sets of normally distributed variables, paired t-tests were performed; for nonnormally distributed variables, values were compared using signed rank test.
Results
UV Light–evoked Attraction/Avoidance Behavioral Response Is Intensity Dependent
Previously reported avoidance to high-intensity UV light was observed over a 12-h light phase simulating daytime at 1-h resolution (Baik et al., 2018), while UV-evoked positive phototaxis events occur within a few minutes (Baik et al., 2017). If UV light behavioral attraction/avoidance depends on the duration of UV exposure, 1-h resolution binning could miss rapid positive phototaxis responses. To determine whether flies exhibit a rapid positive phototaxis response at different intensities of UV light, we subjected adult flies to light choice assays collected and analyzed in 1-min bins. Wild-type control flies exhibit little-to-no attraction to high-intensity UV light (365 nm, 400 µW/cm2) in the first few minutes of the light phase (Fig. 1). Mutant flies lacking all external opsin-based photoreceptors (glass60j flies and norpAP24 flies) similarly lack clear positive phototaxis responses to the high-intensity UV light (Fig. 1A, B). In contrast, mutant flies lacking blue/UV-specific CRY/HK-mediated (cry-/- and hk-/-) or violet-peak RH7-mediated (rh71) phototransduction show strong, fast kinetic attraction responses to high-intensity UV light that slowly attenuate with time (Fig. 1D-F; Suppl. Fig. S1C).
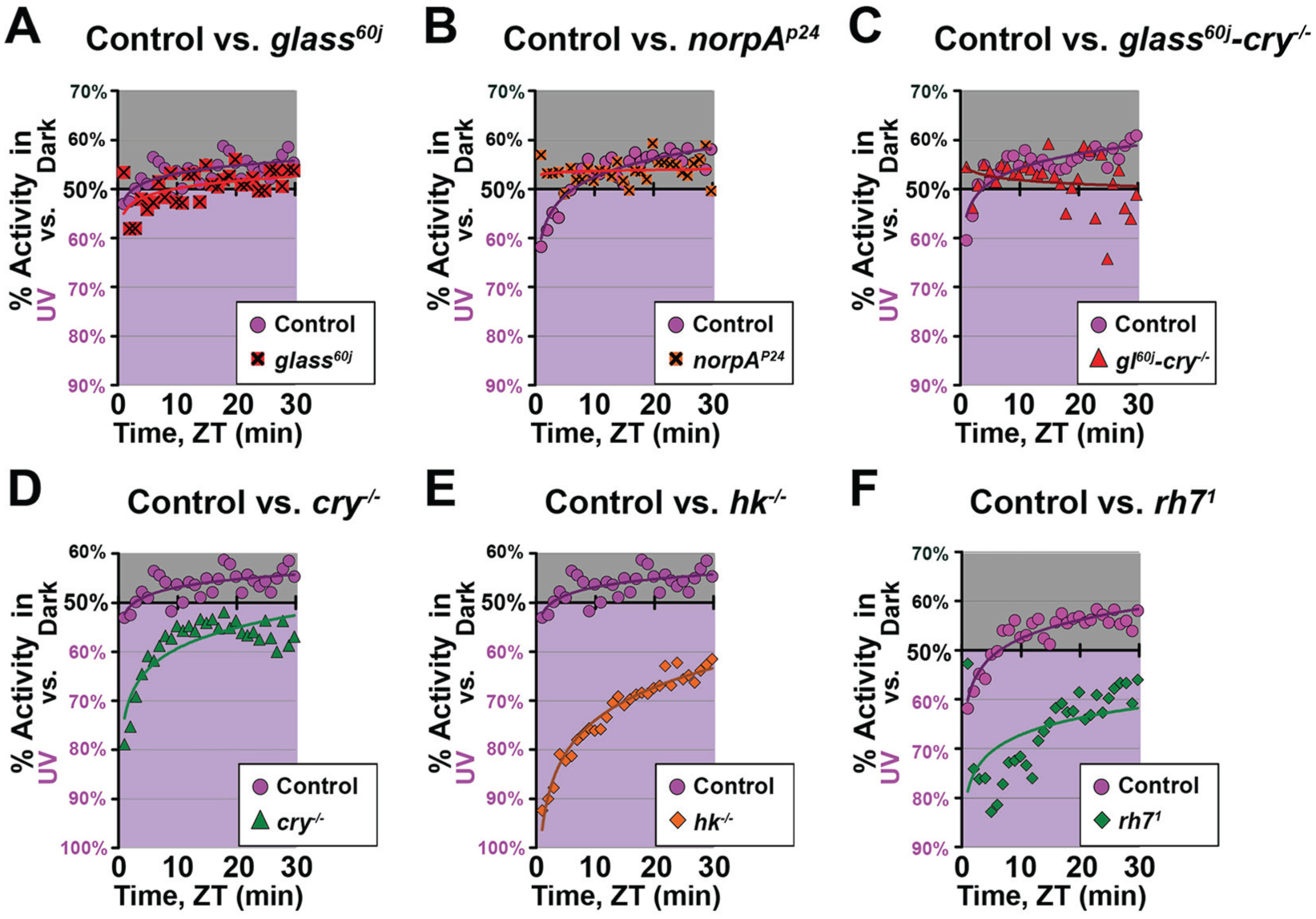
Internal neuronal photoreceptors are necessary for avoidance behavioral response to high-intensity ultraviolet (UV) light. UV attraction/avoidance behavior measured by preference for a shaded environment versus a high-intensity UV-exposed environment (365 nm, 400 μW/cm2) during the light phase of a standard 12 h:12 h light:dark cycle. Preference is calculated by percentage of activity in each environment over total activity for each time bin. (A-F) Preferences of wild-type control (w1118) versus mutant flies to high-intensity UV (365 nm, 400 μW/cm2) in ZT 0-30 min shown in 1-min bin. Similar to wild-type flies that show little to no attraction in the first few minutes of UV light exposure, (A) glass60j (n = 76 vs. control, n = 76), (B) norpAP24 (n = 80 vs. control, n = 79), and (C) glass60j-cry-/- (n = 55 vs. control, n = 95) mutant flies lack clear positive phototaxis responses. In contrast, (D) cry-/- (n = 78 vs. control, n = 76), (E) hk-/- (n = 77 vs. control, n = 76), and (F) rh71 (n = 42 vs. control, n = 79) mutant flies show strong fast positive phototaxis responses to high-intensity UV light.
To test if UV light intensity affects the choice between positive phototaxis and avoidance, we measured the choice behavior under low-intensity UV light (10 μW/cm2). Wild-type control flies show strong attraction to dim UV light that slowly attenuates (Fig. 2). Thus, attraction versus avoidance behavioral responses are dependent on UV light intensity. Mutant flies lacking CRY or HK also show attraction to low-intensity UV light (Fig. 2B, C, F, G). Similarly, mutant flies lacking RH7 show strong attraction to low-intensity UV light (Fig. 2D, H). In contrast, mutant flies lacking all external opsin-based photoreceptors (glass60j) are not attracted to low-intensity UV light (Fig. 2A). Flies lacking CRY/HK- or RH7-mediated phototransduction exhibit positive phototaxis, while flies lacking external opsin-based phototransduction exhibit very slight avoidance, regardless of the intensity of UV light exposure. Over the 12-h period of UV exposure during the daytime-simulating light phase, CRY-, HK-, and RH7-null mutant flies still exhibit time-of-day dependence (Fig. 2F-H), while external opsin-based photoreceptor mutant flies (glass60j and norpAP24) show a relative lack of circadian modulation in attraction/avoidance to UV light (Fig. 2E; Suppl. Fig. S1A). This suggests that CRY/HK- and RH7-mediated phototransduction is important for deciphering the intensity of UV light and thus signaling avoidance in response to high-intensity UV light. In contrast, external opsin-based phototransduction is necessary for evoking fast positive phototaxis response to low-intensity UV light and time-of-day–dependent modulation of attraction/avoidance. Double mutant flies lacking both opsin-based external photoreceptors and CRY-mediated phototransduction almost completely lack any preference for a UV-exposed versus a shaded environment (Fig. 1C; Suppl. Fig. S1B).
Ultraviolet (UV) light–evoked attraction/avoidance behavioral response is intensity dependent. UV attraction/avoidance behavior measured by the preference for a shaded environment versus a low-intensity UV-exposed environment (365 nm, 10 μW/cm2) during the light phase of a standard 12 h:12 h light:dark cycle. Preference is calculated by percentage of activity in each environment over total activity for each time bin. Preference of wild-type controls (w1118; n = 47) versus mutant flies for low-intensity UV (10 μW/cm2) in (A-D) ZT 0-30 min shown in 1-min bin and (E-H) in the light phase (ZT 0-12 h) shown in 1-h bin. Unlike wild-type control flies that show a strong attraction to low-intensity UV light, (A) glass60j flies (n = 31) lack attraction to low-intensity UV light in ZT 0-30 min. (B) cry-/- (n = 44), (C) hk-/- (n = 31), and (D) rh71 (n = 105) mutant flies show a strong attraction to low-intensity UV light in ZT 0-30 min. (E) glass60j flies (n = 31) sustain slight avoidance to low-intensity UV light throughout the ZT 0-12 h light phase. In contrast, wild-type control, (F) cry-/- (n = 44), (G) hk-/- (n = 31), and (H) rh71 (n = 105) mutant flies exhibit attraction to low-intensity UV light in the ZT 0-12 h light phase shown in 1-h bins.
High-intensity UV Light Evokes Avoidance in a Clock-dependent and Exposure Duration–independent Manner, while Low-intensity UV Light Evokes Fast Attraction Response in an External Opsin Photoreceptor-dependent Manner
Behavioral attraction to low-intensity UV light is observed most clearly in the first few minutes following light onset, followed by a gradual decrease in the degree of attraction within 15 min. Under a constant 12-h UV light phase, time-of-day–dependent changes in overall attraction/avoidance response behaviors are modulated by the circadian clock (Baik et al., 2018). Due to adaptation, it is difficult to determine the acute light responses using this chronic light choice assay. To determine acute light responses distributed throughout the day, we adapted the light choice assay to administer acute 15-min pulses of UV light (365 nm, 400 μW/cm2 for high-intensity pulses or 10 μW/cm2 for low-intensity pulses) throughout the 12-h light phase at hourly intervals. The acute light response was measured in flies already entrained to 12:12 LD prior to the experiment. The experiment was carried out under dark condition (DD), and light pulses were administered only during the subjective light phase to maintain prior circadian entrainment.
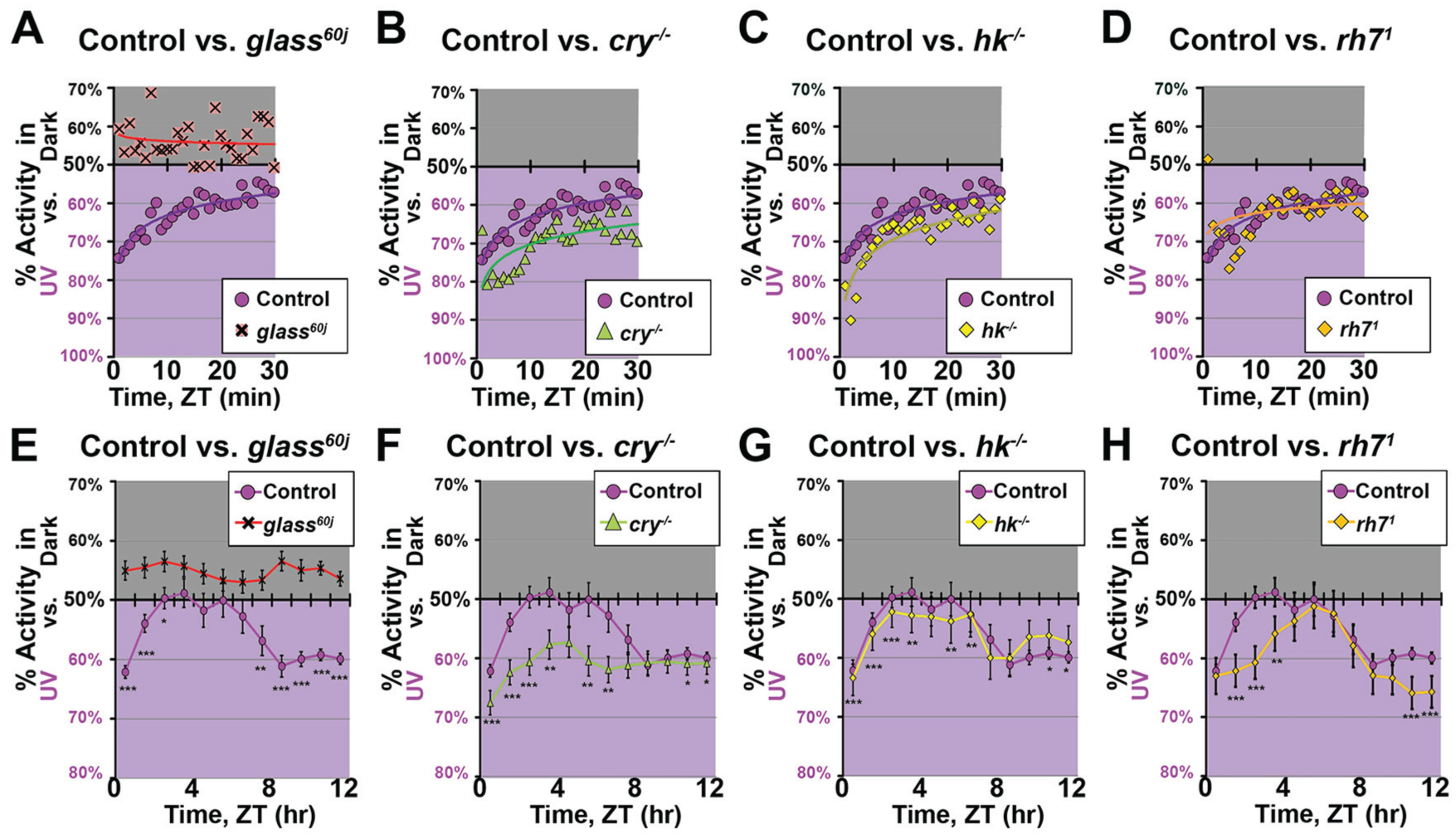
Under this acute repeated-pulse light choice assay, wild-type control flies still exhibit robust avoidance to high-intensity UV light in the early-mid light phase that shifts valence to light attraction toward the end of the light phase (Fig. 3A; Suppl. Fig. S2A). In contrast, circadian mutant flies, timeless-null (tim0) and clock-null (clkOUT), show a strong acute attraction to high-intensity UV pulses that do not vary with time of day (Fig. 3E, F). This shows that the avoidant behavioral response to acute high-intensity UV light pulses is modulated by the circadian clock, rather than by the simple duration of UV light exposure. Wild-type control flies exhibit a fast positive phototaxis response to acute low-intensity UV light pulses throughout the entire light phase (Fig. 4A; Suppl. Fig. S2A). Similarly, circadian mutants tim0 and clkOUT also show rapid attraction to low-intensity UV light at all times of the light phase (Fig. 4E, F). This suggests that the valance of UV light choice is gated by light intensity: avoidance is a chronic circadian clock–modulated response to high-intensity UV light, while attraction is an acute circadian clock–independent behavioral response to low-intensity UV light.
High-intensity ultraviolet (UV) light evokes avoidance modulated by the circadian clock that is independent of the duration of light exposure. UV attraction/avoidance behavior measured by the preference for a shaded environment versus a high-intensity UV-exposed environment (365 nm, 400 μW/cm2) during acute 15-min pulses of light throughout the 12-h light phase at hourly intervals. Preference is calculated by percentage of activity in each environment over total activity for each 1-min bin. (A) Wild-type control (w1118; n = 61) flies show avoidance to high-intensity UV light in the early-mid light phase. In contrast, (B) cry-/- (n = 63) and (C) hk-/- (n = 61) mutant flies show attraction to high-intensity 15-min UV light exposures at all time of day. (D) glass60j (n = 58) mutant flies lack clear preference at all times of the light phase. (E) timeless0 (n = 100) and (F) clockOUT (n = 102) circadian mutant flies show attraction to high-intensity 15-min UV light exposures regardless of time of day.
External opsin-based photoreceptors modulate attraction behavior in response to low-intensity ultraviolet (UV) light. UV attraction/avoidance behavior measured by the preference for a shaded environment versus a low-intensity UV-exposed environment (365 nm, 10 μW/cm2) during acute 15-min pulses of light throughout the 12-h light phase at hourly intervals. Preference is calculated by percentage of activity in each environment over total activity for each 1-min bin. (A) Wild-type control (w1118; n = 61), (B) cry-/- (n = 60), and (C) hk-/- (n = 62) flies show attraction to low-intensity 15-min UV light exposures at all times of day. In contrast, (D) glass60j (n = 62) mutant flies lack attraction. (E) timeless0 (n = 44) and (F) clockOUT (n = 40) circadian mutant flies show attraction to low-intensity 15-min UV light exposures regardless of time of day.
CRY-mediated electrical responses signal through an HK-dependent pathway, which couples light activation to potassium channel modulation in a redox-dependent mechanism (Fogle et al., 2015). Both mutant flies null for CRY/HK-mediated phototransduction show rapid acute attraction to both high- and low-intensity UV pulses that do not vary with time of day (Fig. 3B, C; Fig. 4B, C; Suppl. Fig. S2C, D), supporting earlier results showing that CRY codes for UV light avoidance responses (Baik et al., 2017; Baik et al., 2018). In contrast, mutant flies lacking external opsin-based photoreceptors (glass60j) lack clear preference (no clear attraction nor avoidance) throughout the light phase for both high- and low-intensity UV pulses (Fig. 3D; Fig.4D; Suppl. Fig. S2B). We conclude that high-intensity UV light evoked avoidance signals via a CRY/HK-mediated phototransduction mechanism, which is modulated by the circadian clock and appears to be independent of the duration of UV light exposure. In contrast, low-intensity UV light evokes acute attraction in a circadian clock–independent manner via external opsin-based photoreceptors. Thus, UV light intensity responses signal through different phototransduction channels in an intensity-dependent manner.
External and Internal Phototransduction Mechanisms Modulate Distinct Time-of-day–dependent Spatial Preference of Light Environment
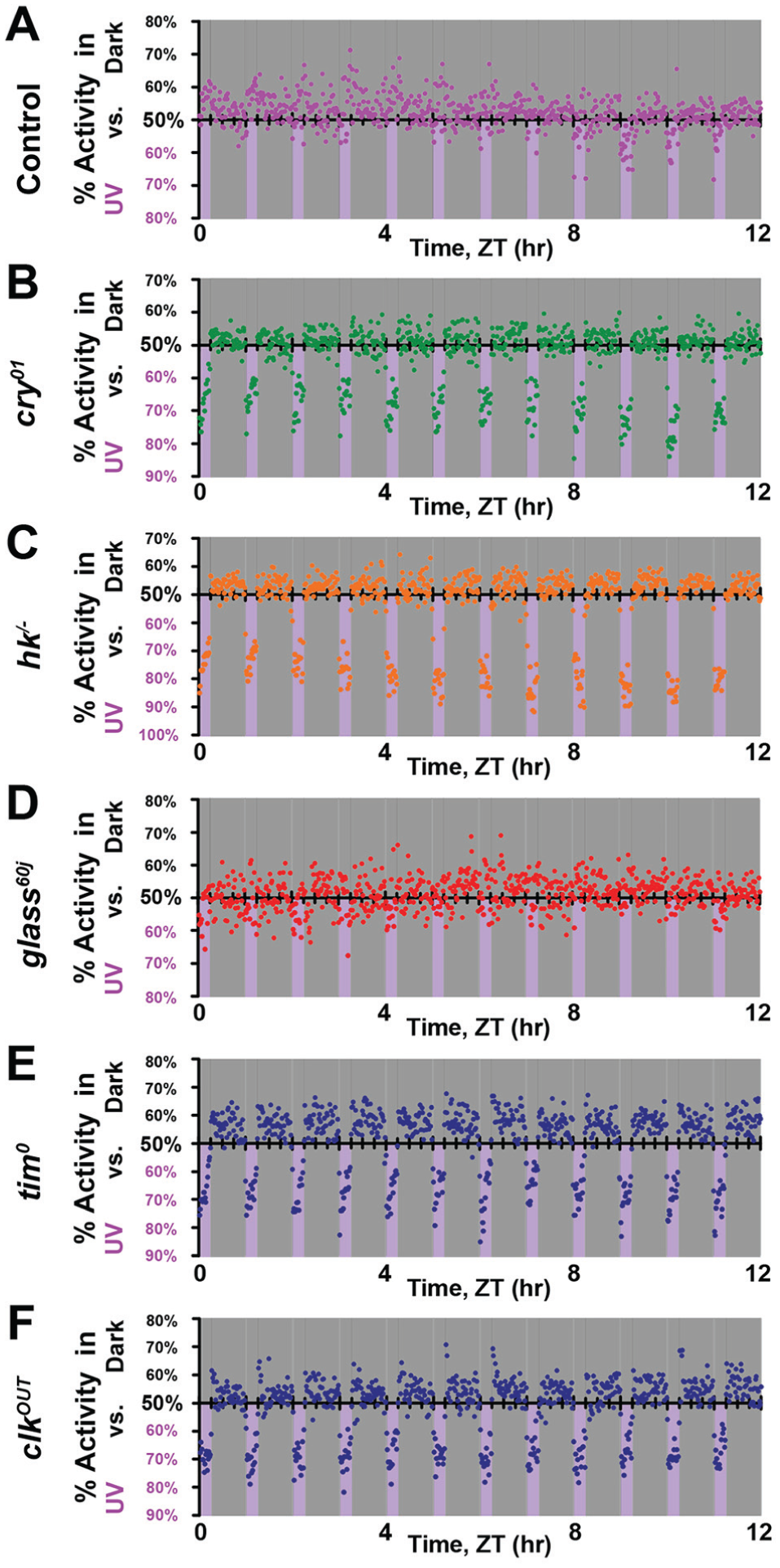
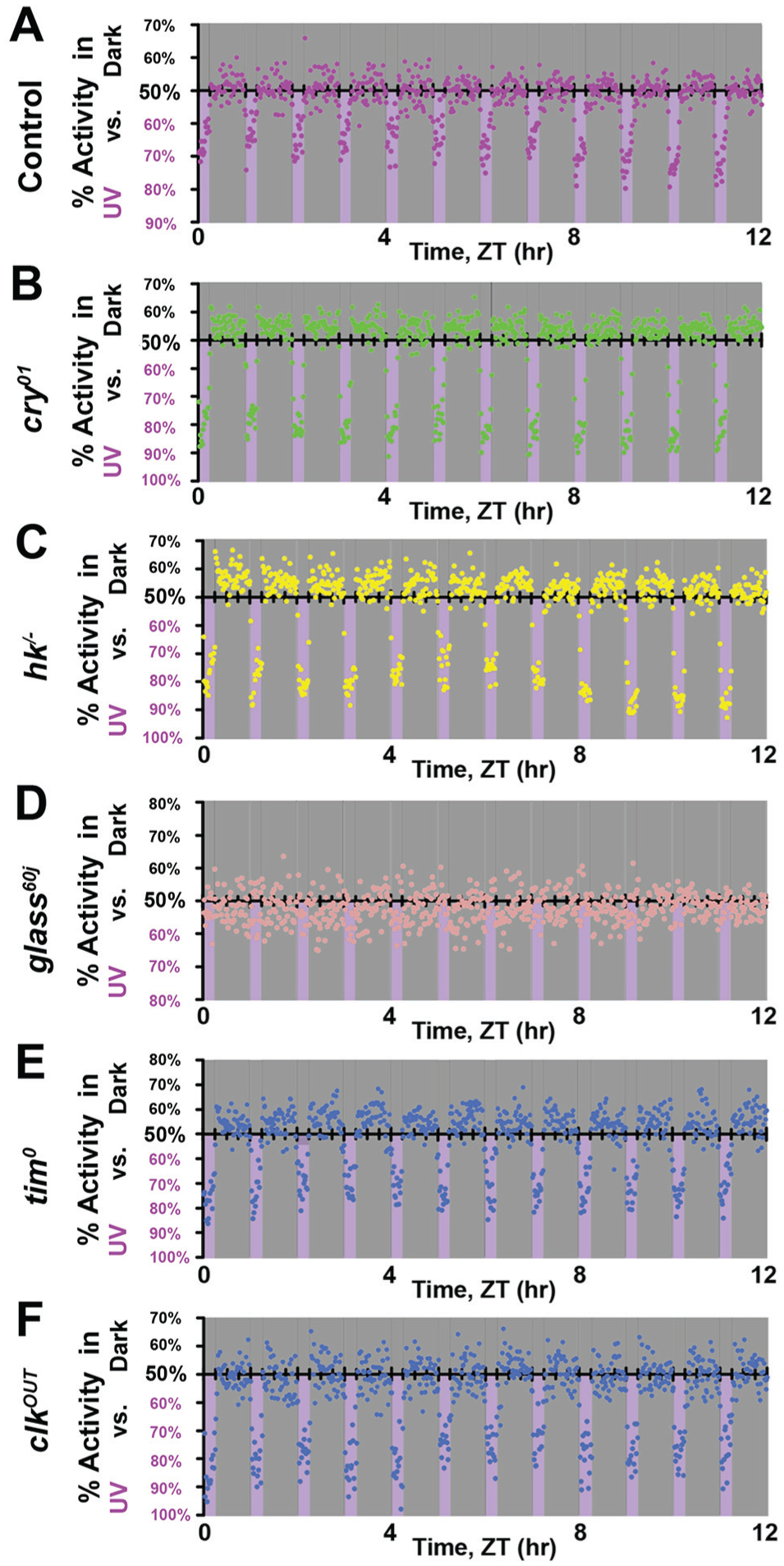
To resolve the behavioral responses to UV light at high spatial resolution, we used multibeam locomotor activity monitors (with infrared beams spaced every 3 mm, 17 beams on each light-exposed versus shaded side with a total of 34 beams per fly; Suppl. Fig. S3). Wild-type control flies strongly prefer shaded locations at the end of tubes that are the most distant from the UV-exposed light area. This behavior is subject to circadian-modulated time-of-day–driven preferences, with UV avoidance behavior that peaks during the middle of the light phase (Fig. 5A). Wild-type control flies venture out of the shade to UV-exposed areas at the beginning (ZT0-1) and the end (ZT10-12) of the light phase, even though UV light intensity does not vary throughout the 12-h light phase (Fig. 5A). During the simulated midday (ZT1-10), control flies strongly prefer to be at the edge farthest from the UV-exposed environment, close to the food. However, food location does not dictate this behavioral preference for location in wild-type flies. The time at which wild-type flies prefer the edge farthest from the UV-exposed environment coincides with the trough of the circadian feeding rhythm (Barber et al., 2016), and the other end of the tube on the UV-exposed side is not a strongly preferred location at any time of the light phase.
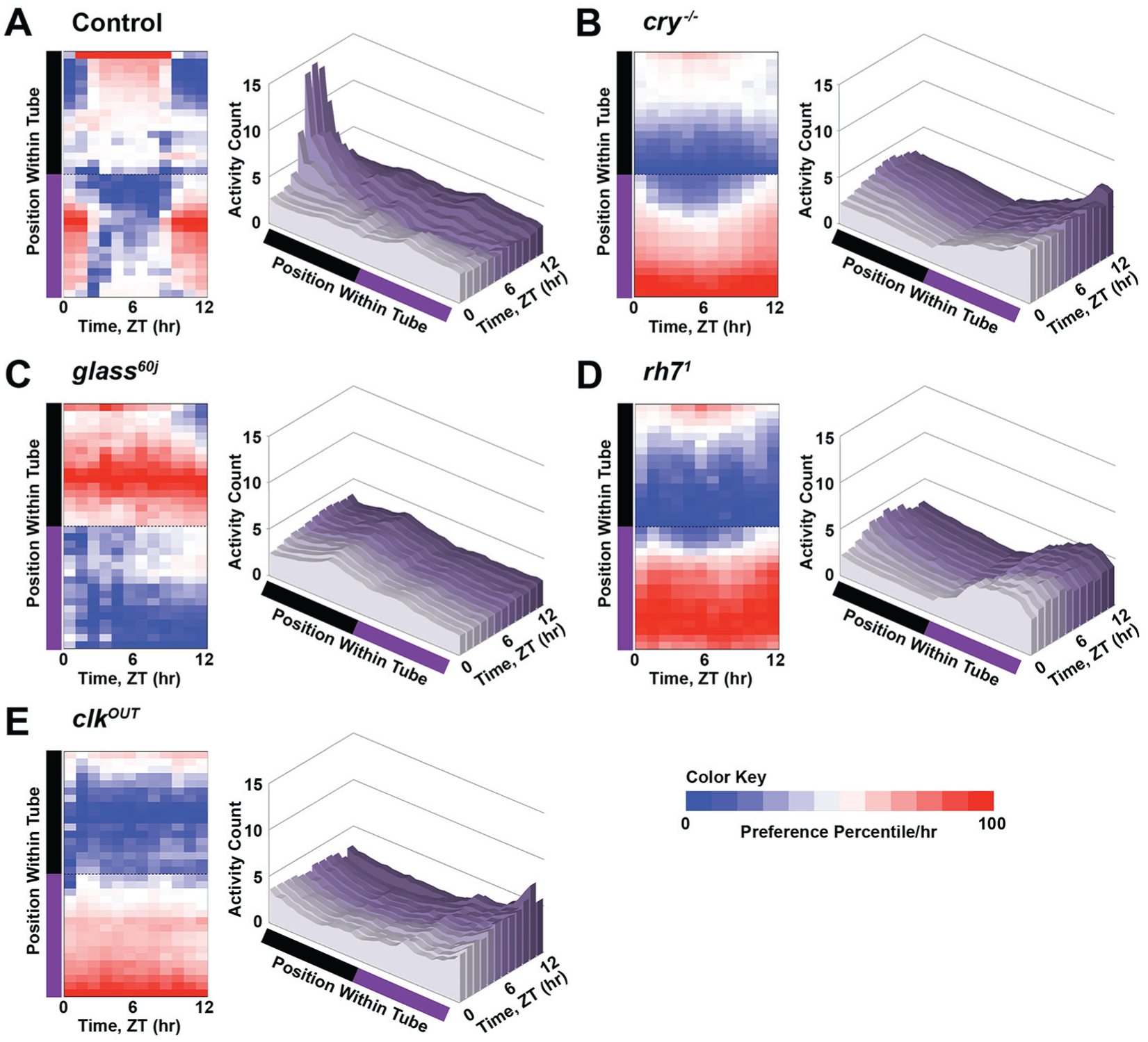
Avoidance/attraction behavior has distinct time-of-day–dependent spatial preference. (A-C) Time-of-day–dependent spatial preference of avoidance/attraction behavior in response to high-intensity ultraviolet (UV) light (365 nm, 400 μW/cm2) versus shade was monitored with high spatial and temporal resolutions using multibeam locomotor activity monitors. Heat map of preference for each 1-h bin (left) and activity count at each position for every 1-h bin (right) are shown. (A) Wild-type control (w1118; n = 144) flies prefer the mid-area of the UV-exposed environment during the early morning and the hours before simulated dusk. But during the midday, they prefer the edge of the shaded environment, farthest from the UV environment and close to the food. In contrast, (B) cry-/- (n = 61) flies strongly prefer the area farthest out in the UV-exposed environment at all times of the day. (C) glass60j (n = 64) flies prefer the mid-area within the shaded environment, regardless of the time of day. (D) rh71 (n = 46) flies strongly prefer the outer edge of UV-exposed environment at all times of the day. (E) clkOUT (n = 48) flies prefer the area farthest out in the UV-exposed environment close to the food at all times of the day.
In marked contrast to wild-type flies, cry-null flies strongly prefer the UV-exposed area (Fig. 5B) and avoid the mid-region between both the shaded- and the UV light–exposed environments. In additional detail, cry-null flies favor the edge closest to the food (Fig. 5B). With striking similarity, rh7-null flies strongly prefer the UV-exposed area, although not as close to the food, and like the cry-null flies, they avoid the mid-region between both the shaded and the UV light–exposed environments at all times of the day (Fig. 5D). ClkOUT circadian mutant flies prefer the UV light–exposed environment regardless of the time of day and prefer the edge closest to the food in the UV-exposed area. Unlike cry- or rh7-null flies, clkOUT mutants do not avoid the mid-region between the shaded and the UV light–exposed environments (Fig. 5E). Glass60j mutant flies, which lack all external opsin-based photoreceptors but still express CRY and HK, prefer the shaded environment at all times of the light phase and lack clear time-of-day–dependent modulation. Even though they cannot “see” UV light with image-forming vision, glass60j flies sense and avoid UV light. Image-forming vision does have some spatio-behavioral impact. Unlike control flies, glass60j flies prefer the mid-region within the shaded environment at all times of the light phase rather than the edge farthest from the UV light (Fig. 5C).
DISCUSSION
Insect behavior, including circadian rhythmic behavior, is subject to the integration of multiple sensory inputs. The combination of these sensory cues in natural environments is sufficiently powerful to mask major clock mutations, which yield severe behavioral impairments in less complex laboratory conditions (Vanin et al., 2012). We demonstrate that flies’ complex spatiotemporal behavioral light responses integrate multiple photic inputs to navigate their light environment. Low-intensity UV light evokes rapid attraction, which diminishes quickly. This confirms its importance as a survival mechanism for escape behavior. Here, we show that this fast attraction behavior is induced by external opsin-based photoreceptors at all times of the day. High-intensity UV evokes avoidance via internal/direct neuronal photoreceptors, CRY and RH7. In addition to intensity of light, circadian time and light spectra are important for avoidance/attraction signaling (Baik et al., 2018). In contrast to the fast kinetics of short-wavelength light acute attraction, CRY- and RH7-evoked avoidance is slowly modulated by the circadian clock, changing over the span of hours during the daytime-simulating light phase (Baik et al., 2017; Baik et al., 2018; Suppl. Fig. S1).
External and internal photoreceptors contribute to light entrainment of the circadian locomotor activity rhythm (Emery et al., 1998; Stanewsky et al., 1998; Helfrich-Forster et al., 2001; Helfrich-Forster et al., 2002; Malpel et al., 2002; Rieger et al., 2003; Klarsfeld et al., 2004; Veleri et al., 2007; Sheeba et al., 2008; Kistenpfennig et al., 2012; Schlichting et al., 2014; Schlichting et al., 2015; Saint-Charles et al., 2016; Schlichting et al., 2016; Ni et al., 2017). Both internal photoreceptors, CRY and RH7, are expressed in the circadian pacemaker neurons in the Drosophila brain, including lateral ventral neurons (LNv; Benito et al., 2008; Yoshii et al., 2008; Ni et al., 2017). LNv circadian neurons have been shown to be necessary for having a normal short-wavelength light avoidance (Baik et al., 2018). Double knockout mutant flies lacking both external and internal photoreceptors, glass60j-cry-/-, almost completely lack any preference for light environment and thus do not display either avoidance or attraction to UV light (Fig. 1; Suppl. Fig. S1). The similarity of their behavioral responses supports the notion that CRY- and RH7- phototransduction signals may converge to mediate avoidance/attraction behavior. In addition to circadian neurons, CRY is expressed in the eyes and plays a role in visual sensitivity (Mazzotta et al., 2013).
Here, we implemented multibeam activity monitors to acquire high temporal and spatial resolution of UV-evoked avoidance/attraction behavior. Interestingly, both wild-type control and glass60j flies show minimal behavioral preference for the shaded food–containing edge toward the end of the light phase, even though it coincides with the peak time of the feeding rhythm (Barber et al., 2016). In contrast, cry-null and clkOUT mutant flies show a strong positional preference in the UV-exposed area close to the food at all times of the light phase, with overall less time-of-day–modulated positional preference as compared with wild-type controls. This suggests that observed spatial preference in the far edge in simulated midday is not due to the presence of food. We show that there are multiple levels of sensory processing even for a single type of input such as light. We conclude that multiple phototransduction mechanisms modulate a complex behavioral output depending on its spectra, intensity, and time/duration of exposure.
At the organismal level, integration of multiple types of sensory inputs, including light and temperature, is crucial to synchronize to the changing environment. Both light and temperature cycles shape complex behavioral output, including locomotor activity and sleep in Drosophila (Currie et al., 2009; Yoshii et al., 2009; Head et al., 2015; Harper et al., 2016; Parisky et al., 2016; Harper et al., 2017; Lamaze et al., 2017; Chen et al., 2018; Yadlapalli et al., 2018). Multisensory entrainment of the circadian clock occurs by integrating different sensory cues, including light inputs via CRY and thermal inputs from transient receptor potential A1 (TrpA1; Das et al., 2015; Green et al., 2015; Das et al., 2016; Harper et al., 2017). Rhythmic preference for both light and temperature environment further reinforces an intricate and complex behavioral output (Figs. 1-5; Stevenson, 1985; Kaneko et al., 2012; Head et al., 2015; Baik et al., 2017; Tang et al., 2017).
In conclusion, complex spatiotemporal behavioral responses to UV light are mediated through multiple phototransduction pathways. Acute high-intensity UV light exposure (minutes) in wild-type flies evokes a transient phototaxis attractive response, which is mediated by the opsin-based phototransduction in the eyes. With longer high intensity UV light exposure (tens of minutes), avoidance is the dominant response, which is mediated by the CRY/HK and RH7 pathways. Both rapid/acute and long term/chronic CRY/HK mediated behavioral responses vary by time of day, while external photoreceptor/opsin mediated responses do not appear to vary by time of day. Acute low intensity UV light exposure (minutes) in wild type flies evokes a longer lasting transient phototaxis attractive response that resolves to mostly neutral responses. Spatial responses to UV light are strongly influenced by both neuronal cell autonomous phototransduction (CRY/HK) and external opsin-based photoreceptor phototransduction, indicating integrated contributions from both major UV sensing photosystems.
Supplemental Material
FigS1_Baik_2.18.19 – Supplemental material for Multiple Phototransduction Inputs Integrate to Mediate UV Light–evoked Avoidance/Attraction Behavior in Drosophila
Supplemental material, FigS1_Baik_2.18.19 for Multiple Phototransduction Inputs Integrate to Mediate UV Light–evoked Avoidance/Attraction Behavior in Drosophila by Lisa Soyeon Baik, Yocelyn Recinos, Joshua A. Chevez, David D. Au and Todd C. Holmes in Journal of Biological Rhythms
Supplemental Material
FigS2_Strobe_15L_only_2.18.19 – Supplemental material for Multiple Phototransduction Inputs Integrate to Mediate UV Light–evoked Avoidance/Attraction Behavior in Drosophila
Supplemental material, FigS2_Strobe_15L_only_2.18.19 for Multiple Phototransduction Inputs Integrate to Mediate UV Light–evoked Avoidance/Attraction Behavior in Drosophila by Lisa Soyeon Baik, Yocelyn Recinos, Joshua A. Chevez, David D. Au and Todd C. Holmes in Journal of Biological Rhythms
Supplemental Material
FigS3_Baik_2.18.19 – Supplemental material for Multiple Phototransduction Inputs Integrate to Mediate UV Light–evoked Avoidance/Attraction Behavior in Drosophila
Supplemental material, FigS3_Baik_2.18.19 for Multiple Phototransduction Inputs Integrate to Mediate UV Light–evoked Avoidance/Attraction Behavior in Drosophila by Lisa Soyeon Baik, Yocelyn Recinos, Joshua A. Chevez, David D. Au and Todd C. Holmes in Journal of Biological Rhythms
Footnotes
Acknowledgements
We thank Jeff Hall (cry01), Joanna Chiu (tim0 and clkOUT), Craig Montell (rh71 and norpAP24), and Ming Zhou (hk-/-) for providing fly stocks and the Bloomington Stock Center for other lines; Ceazar Nave and Annika Barber for helpful discussions; and Anthony Tette, Janita Parpana, and Duke Park for excellent administrative support. Funding was provided by the U.S. Department of Health and Human Services, National Institutes of Health (GM127102).
Conflict Of Interest Statement
The authors have no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Notes
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.