
Abstract
Light is known to elicit non–image-forming responses, such as effects on alertness. This has been reported especially during light exposure at night. Nighttime results might not be translatable to the day. This article aims to provide an overview of (1) neural mechanisms regulating alertness, (2) ways of measuring and quantifying alertness, and (3) the current literature specifically regarding effects of different intensities of white light on various measures and correlates of alertness during the daytime. In general, the present literature provides inconclusive results on alerting effects of the intensity of white light during daytime, particularly for objective measures and correlates of alertness. However, the various research paradigms employed in earlier studies differed substantially, and most studies tested only a limited set of lighting conditions. Therefore, the alerting potential of exposure to more intense white light should be investigated in a systematic, dose-dependent manner with multiple correlates of alertness and within one experimental paradigm over the course of day.
Introduction
Light is known to elicit both image- and non–image-forming (NIF) responses, such as entrainment of the biological clock to a 24-h cycle (Golombek and Rosenstein, 2010; Hughes et al., 2015; Pittendrigh and Daan, 1976), suppression of the nocturnal hormone melatonin (Cajochen et al., 2000; Gooley et al., 2011; Zeitzer et al., 2000), and acute alerting effects of light (e.g., Cajochen, 2007; Cajochen et al., 2000, 2005, 2011; Chellappa et al., 2013; Lavoie et al., 2003; Najjar et al., 2014; Rüger et al., 2006; Smolders et al., 2012; Van Der Lely et al., 2015). This article aims to provide an overview of the current literature regarding daytime NIF effects of white light on alertness and serves as an introduction to 2 independently performed experiments investigating the dose-dependent relationship between the intensity of white light and markers (and correlates of) alertness during daytime (Lok et al., 2018 [this issue]; Smolders et al., 2018 [this issue]). Possible mechanisms involved in regulation of alertness and measures quantifying alertness will also be discussed.
Alertness is a construct associated with high levels of environmental awareness (Figueiro et al., 2009) and is defined as achieving and maintaining a state of high sensitivity to incoming stimuli (Posner, 2008). Sleepiness is often used to indicate the adverse state, although the definition differs slightly from the exact opposite of alertness. It has been defined as the perceived experience of the propensity to fall asleep (Moller et al., 2006). Alertness and arousal are closely related but distinct concepts that are sometimes used interchangeably. Arousal refers to nonspecific activation related to changes in sleep and wakefulness (Oken et al., 2006). It plays an important role in, among others, the regulation of alertness, which is a more specific form of activation and requires some cognitive processing (Oken et al., 2006).
There are important implications associated with maintaining alertness during daytime, since it is known to affect cognitive performance (Figueiro et al., 2016), perceptual skills (Curcio et al., 2001), reasoning abilities (Curcio et al., 2001), judgment and decision-making capabilities (Van Dongen et al., 2004), psychological and physiological well-being (Cajochen et al., 2003; Dijk et al., 1992; Hull et al., 2003), caloric intake (Pardi et al., 2016), and pain perception (Alexandre et al., 2017). However, contemporary developments, such as the emergence of the 24-h society, pose unique physiological and psychological challenges on alertness. Identifying a tool to modulate alertness during waking hours is critical to adapt to the 24-h society and prevent or alleviate potential problems associated with decreasing alertness.
Circuitries
Light is directly related to vision, which relies on classical photoreceptors, rods, and cones, with projections to the visual cortex (see, for instance, Horvath et al., 1999). The discovery of a novel class of photoreceptors, called intrinsically photosensitive retinal ganglion cells (ipRGCs; Provencio et al., 1998, 2000), led to the detection that these photoreceptors were not necessarily involved in image forming but rather NIF aspects, such as entrainment (Berson et al., 2002; Hattar et al., 2002; Hankins et al., 2008). Nowadays, it is generally accepted that ipRGCs are important for both image and NIF responses (Sonoda and Schmidt, 2016). It is well established that ipRGCs project light information to, among others, hypothalamic regions such as the suprachiasmatic nucleus (SCN; the master pacemaker in the brain) via the retinohypothalamic tract (Gooley et al., 2003; Hattar et al., 2002). Studies using a monochromatic narrowband light of ~460 nm, specifically stimulating ipRGCs, have generally shown significant improvements in alertness (e.g., Lockley et al., 2006; Vandewalle et al., 2007), coinciding with increases in SCN and thalamic activity (Aston-Jones, 2005; Vandewalle et al., 2006, 2007).
Multiple hypotheses have been generated to speculate on light-induced alertness regulation, since alertness is known to be under the control of hypothalamic-associated regions (Aston-Jones, 2005). First, research has established that light exposure affects SCN activity (Gooley et al., 2003; Hattar et al., 2002, 2006; see also Golombek and Rosenstein, 2010; Fisk et al., 2018). In fact, light is the most important time cue for the master pacemaker located in the SCN (Duffy and Wright, 2005; Vetter et al., 2011). It is crucial for aligning the internal clock, which has a rhythm of about 24 h, to environmental demands (Czeisler et al., 1980). As alertness is implicated to be under regulation of the SCN, it also follows (under entrained conditions) 24-h rhythmicity. Among day-active persons, relatively low levels of alertness have been reported during the night (when melatonin levels are high), whereas relatively high levels of alertness exist during the subjective day (when melatonin is virtually absent; Åkerstedt et al., 2017; Dijk et al., 1992; Hull et al., 2003).
In addition to SCN activation by light, various hypothalamic regions involved in the regulation of alertness are directly controlled through projections from the ipRGCs and/or indirectly controlled by light due to projections from the SCN (Aston-Jones, 2005; Gooley et al., 2003; Hattar et al., 2002, 2006; Perrin et al., 2004; Vandewalle et al., 2006, 2007, 2009). Examples of these hypothalamic areas are the ventral lateral preoptic area (VLPO) and locus coeruleus (LC). The VLPO is known for its distinct function in sleep regulation and arousal (Fort et al., 2009; Gvilia, 2006; Sherin et al., 1996), both of which influence one’s level of alertness. When neurotransmitter systems of the VLPO are active, ascending arousal systems are inhibited, resulting in promotion of sleep (Lu et al., 2002; Moore et al., 2012). If these inhibitory neurotransmitter systems are inactive, alertness is promoted (Lu et al., 2002; Moore et al., 2012). In rats, direct projections via ipRGCs and indirect projections from the SCN to the VLPO have been determined (Lu et al., 2000; Moore et al., 2012), indicating that light can play an important role in the regulation of modulations in this brain area (Chou et al., 2002; Gooley et al., 2003). Similar circuitries have also been implicated in humans (Gooley et al., 2003; Perrin et al., 2004; Hattar et al., 2006).
Another hypothalamic-associated area involved in the regulation of alertness is the LC, which is a dense cluster of norepinephrine neurons and a source of efferent projections to multiple central nervous system regions (Aston-Jones, 2005). When the LC is excited by injecting excitatory agents in it, there is an increased level of arousal (Aston-Jones, 2005). This coincides with increased electroencephalographic activity in the frontal neocortex and theta waves in the hippocampus, suggesting an increase in alertness (Foote and Berridge, 1991). The neuropeptide orexin has been shown to be one of these agents that can excite LC neurons strongly and therefore promote arousal and wakefulness (Hagan et al., 1999). In addition to direct excitatory effects on the LC, orexins have been shown to affect the SCN firing rate in the rat (Brown et al., 2008; Klisch et al., 2009). In both rats and primates, indirect projections have been identified from the SCN to the LC via the dorsomedial hypothalamic nucleus (Aston-Jones, 2005; Winsky-Sommerer, 2004). Direct effects of light have also been determined in humans, in which light induces modulations in brain activity in an area compatible with LC neurons (Vandewalle et al., 2009).
Both the LC and dorsal raphe (DR) have been shown to play an important role in wakefulness promotion (Lee et al., 2005; Mieda and Yanagisawa, 2002). High levels of alertness can be achieved when wakefulness is promoted through the phase relationship between the endogenous circadian timing system and the sleep-wake cycle (Borbely, 1982; Daan et al., 1984). The caudal raphe nuclei are innervated by projections from the LC (Hermann et al., 1997) and are thought to affect sympathetic function via serotonergic output (Allen and Cechetto, 1994; Jacobs et al., 2002). Sympathetic nervous system activity is associated with high alertness, whereas increases in parasympathetic nervous activity are associated with decreases in alertness (Pressman and Fry, 1989), indicating a role for the serotonergic (5-hydroxytryptamine) system in regulating alertness. Moreover, serotonergic neurons located in the DR fire extensively during wakefulness, while decreased firing rates occur in periods of sleep (McGinty and Harper, 1976; Trulson and Jacobs, 1979). Firing rates may therefore be associated with wakefulness and alertness promotion.
The SCN, VLPO, LC, and DR pathways described above, as well as other neural pathways (see, e.g., Gooley et al., 2003; Hattar et al., 2006; Vandewalle et al., 2009), might be involved in NIF responses caused by light and, in particular, effects of light on alertness.
Quantification of Alertness
Alertness can be quantified with self-report, task performance, and physiological measures (Curcio et al., 2001). A subjective measure of alertness is often the most readily accessible information (Zhou et al., 2012). Subjective measures usually are recorded with self-rating scales, such as the visual analogue scale (a 100-mm-long line on which subjects evaluate their own state by marking a point along the line; Aitken, 1969) and Likert-type scales, such as the Karolinska Sleepiness Scale (KSS; a 9-point anchored scale, on which participants indicate the description level that best reflects their experienced state; Åkerstedt and Gillberg, 1990).
There are multiple performance tasks reflecting alertness, which can be divided into sustained attention versus executive performance tasks. A commonly used performance measure of alertness in lighting research is the Psychomotor Vigilance Task (PVT; in which stimuli are presented continuously and participants respond to each stimulus). This task is defined as a sustained attention task, since it measures the ability to perform over longer periods of time (Drummond et al., 2005). In addition, multiple cognitive performance tasks have been employed to assess the effects of light on executive functioning. These tasks require alertness but also rely on other (higher-order) cognitive functions, such as working memory, inhibition of responses, and/or arithmetic ability. For instance, the Sustained Attention to Response Task (SART; in the standard version of the task, participants are asked to push a button every time a number appears on the screen, except for the number 3), N-back task (in which a sequence of stimuli is presented, and the task consists of indicating when the current stimulus matches the one from n steps earlier in the sequence), and addition tasks are employed in other studies. Reaction time tasks in which subjects have to distinguish targets from nontargets, such as the SART, but also the Wilkinson Auditory Vigilance Task and Go-NoGo task, measure sustained attention as well as the ability to inhibit responses (Bokura et al., 2001).
In addition to such behavioral measures, other ways of assessing alertness objectively are through measuring an individual’s physiological state. One example of a physiological assessment of alertness is electroencephalography (EEG), which reflects central nervous activity. The presence of theta, alpha, and beta rhythms can provide information about the psychophysiological state of alertness (Santamaria and Chiappa, 1987). Autonomic activity measures have also been used as physiological indicators of alertness. Autonomic nervous system (ANS) activity is hypothesized to vary in intensity along a continuum from vigorous activity, intense emotion, and high alertness to calmness and sleep (Lowenstein et al., 1963). The ANS can be divided into the parasympathetic and sympathetic nervous system, in which sympathetic nervous system activity is associated with high alertness, whereas increases in parasympathetic nervous activity are associated with decreases in alertness (Pressman and Fry, 1989). Excitatory impulses from the cerebral cortex traveling via the reticular activating system and hypothalamus can influence sympathetic as well as parasympathetic activity levels (Pressman and Fry, 1989). This may, for instance, lead to pupil fluctuations (measured with pupillography), in which a dilated pupil is associated with higher levels of alertness (Johnson et al., 2008; Ma et al., 2014; Yoss et al., 1970). When alertness decreases, parasympathetic activity is relatively high, which is reflected by a decrease in pupil size and large, slow pupillary oscillations (Johnson et al., 2008; Ma et al., 2014; Yoss et al., 1970) and increases in blink frequency, blink duration, and eyelid movements (measured with electrooculography; Berntson et al., 1997; Caffier et al., 2003). Other parameters of autonomic nervous activation processes are heart rate and heart rate variability, with higher variability with more pronounced sympathetic activation compared to parasympathetic activation (Acharya et al., 2006; Heneghan et al., 2014). Sympathetic dominance likely co-occurs with increased attention and sensitivity to incoming stimuli (e.g., Chua et al., 2012; Hansen et al., 2003; Luque-Casado et al., 2016). Yet it is important to note that the reliability of, for instance, the low-frequency/high-frequency ratio as a measure for sympathetic dominance has been challenged because of, among others, the role of respiratory parameters and the complex, nonlinear interaction between parasympathetic and sympathetic branches (see, e.g., Berntson et al., 1997; Billman, 2013). Body temperature fluctuations have also been suggested to correlate with subjective alertness and task performance (Monk et al., 1983; Wright et al., 2002). Thermosensitive neurons in the anterior hypothalamus and other brain areas have been proposed to be involved in the relationship between alertness and body temperature (Wright et al., 2002). Changes in activity level of these neurons presumably result in correlated changes in sleep propensity and skin vasodilatation and therefore skin temperature (Raymann and Van Someren, 2007). In fact, a causal relationship between skin temperature and arousal-regulating mechanisms was found (Ivanov and Aston-Jones, 2000). Manipulation of skin temperature can lead to changes in alertness and sleep (Raymann, 2005; Raymann et al., 2008; Raymann and Van Someren, 2007). Higher core body temperature is associated with higher self-rated alertness and better performance on PVT (Wright et al., 2002).
Each of the described measures or correlates of alertness have advantages as well as disadvantages. Subjective measures of alertness can give insight into an individual’s experienced level of alertness but only to a certain degree. Conflicts arise because of recall bias and placebo effects, which have to be taken into consideration when using these measures (Cajochen, 2007). Nevertheless, some of the subjective measures used to study alertness, such as the KSS, have been validated with other objective correlates of alertness (Kaida et al., 2006; Zhou et al., 2012). Correlations between subjective alertness and performance measures have been shown (see, for instance, Åkerstedt and Gillberg, 1990; Dorrian et al., 2003; Wright et al., 2002). However, subjective alertness might not reflect performance and vice versa, as reduced alertness does not always reflect the magnitude of performance impairment (Zhou et al., 2012). There is some evidence suggesting that subjective alertness and neurobehavioral performance may respond to the same intervention to different extents (Posner and Rafal, 1987; Rosekind et al., 1995). For instance, in response to accumulating sleep debt, neurobehavioral performance monotonically declines on both PVT performance and the digit symbol substitution task, while subjective sleepiness approaches a (temporary) plateau after an initial increase (Van Dongen et al., 2003). Some studies indicate that correlates between subjective alertness and physiological measures of alertness are stronger compared with performance tasks (Putilov et al., 2012). In the current literature on lighting research, a multimeasure approach is often used to assess the effect of a (light) intervention on alertness.
Effects of White Light During the Night
The acute NIF effects of light on various measures or correlates of alertness have been established especially during the biological night, when alertness levels are generally relatively low and endogenous levels of melatonin are relatively high. Both polychromatic white and monochromatic light (of especially ~460 nm) have been shown to be able to improve nighttime alertness on various indicators (e.g., Badia et al., 1991; Cajochen et al., 2003; Lockley et al., 2006; Rüger et al., 2006; Sahin and Figueiro, 2013; Van Der Lely et al., 2015; Cajochen et al., 2000, 2005, 2011; Chang et al., 2012; Chellappa et al., 2012; Figueiro et al., 2016; Lavoie et al., 2003). In this introductory review, we focus on studies investigating the effects of the intensity of white light on alertness. Studies investigating modulations in the intensity of monochromatic and narrowband light are not included in the overview, since we are generally exposed to polychromatic white light in everyday life. Moreover, it is questionable whether the reported effects of monochromatic or narrowband light can be directly translated to the effect of specific wavelengths in the spectrum of polychromatic light because of the potential opposing actions of different photoreceptor classes (Spitschan et al., 2014; Woelders et al., 2018). While interactions between different photoreceptors may also occur under monochromatic light exposure, potential inhibitory responses might be more pronounced under exposure to white light because of relatively strong activation of more—if not all—classes of photoreceptors. Studies investigating monochromatic or narrowband light have particularly tested wavelengths at or near the peak sensitivity of one class of photoreceptors (e.g., Lockley et al., 2006; Sahin and Figueiro, 2013), resulting in a relatively strong activation of one class of photoreceptors compared with the other photoreceptor classes. For an overview of the potential alertness-enhancing effects of the spectral composition of white light, see Smolders and de Kort (2017) and Souman et al. (2018).
Nocturnal light exposure has been indicated to influence alertness in a dose-dependent manner. In fact, a dose-response relationship between light intensity and different measures of alertness during the night has been determined in the study by Cajochen and colleagues (2000). Results revealed increased levels of alertness with increases in illuminance level according to a logistic function. The maximum response was obtained at about 1000 lx at eye level, and the half-maximum of the alerting effects of light were achieved with illuminances between 90 and 180 lux. The same study also showed that the half-maximum melatonin suppression occurred between 50 and 130 lux and revealed strong correlations between melatonin suppression and alerting effects of light (Cajochen et al., 2000). Several other studies also revealed effects of light on melatonin suppression coinciding with increases in alertness, suggesting that light-induced melatonin suppression might elicit effects on nocturnal alertness (e.g., Chellappa et al., 2011; Lowden et al., 2004). Attenuating SCN-dependent mechanisms responsible for promoting and maintaining cortical and behavioral arousal have been implicated (Dijk and Czeisler, 1995; Lavie, 1997). It is, however, important to note that while significant alerting effects have been reported at night, studies also reported null effects (Souman et al., 2018).
Effects of the Intensity of White Light on Alertness During Daytime
Because there are systematic changes over the course of the 24-h day in both melatonin and alertness levels, with relatively higher alertness and lower melatonin concentrations during daytime compared with nighttime (Cajochen et al., 2003; Gronfier et al., 2007; Lavoie, 1997; Wright et al., 2002; Wyatt et al., 1999), it is questionable whether nighttime results are directly translatable to daytime situations. Nevertheless, some studies have shown acute effects of light on alertness, which are likely not only driven via melatonin suppression (Figueiro et al., 2016, 2009; Plitnick et al., 2010; Van de Werken et al., 2013). Moreover, several studies have revealed acute alerting effects of bright white light during daytime (Badia et al., 1991; Borragán et al., 2017; Daurat et al., 1993; Huiberts et al., 2015, 2016, 2017; Iskra-Golec and Smith, 2008; Kaida et al., 2006; Leichtfried et al., 2015; Maierova et al., 2016; Phipps-Nelson et al., 2003; Rüger et al., 2006; Smolders and de Kort, 2014; Smolders et al., 2012; Vandewalle et al., 2006). Although effects of light on alertness during the day have been studied, results seem to be less conclusive compared with results reported at night. However, as there are important implications associated with improved daytime alertness, it is important to determine to what extent a generally easy accessible tool, such as light, could improve daytime alertness. Therefore, an overview of the literature studying the effects of polychromatic white light intensities on alertness during daytime has been made.
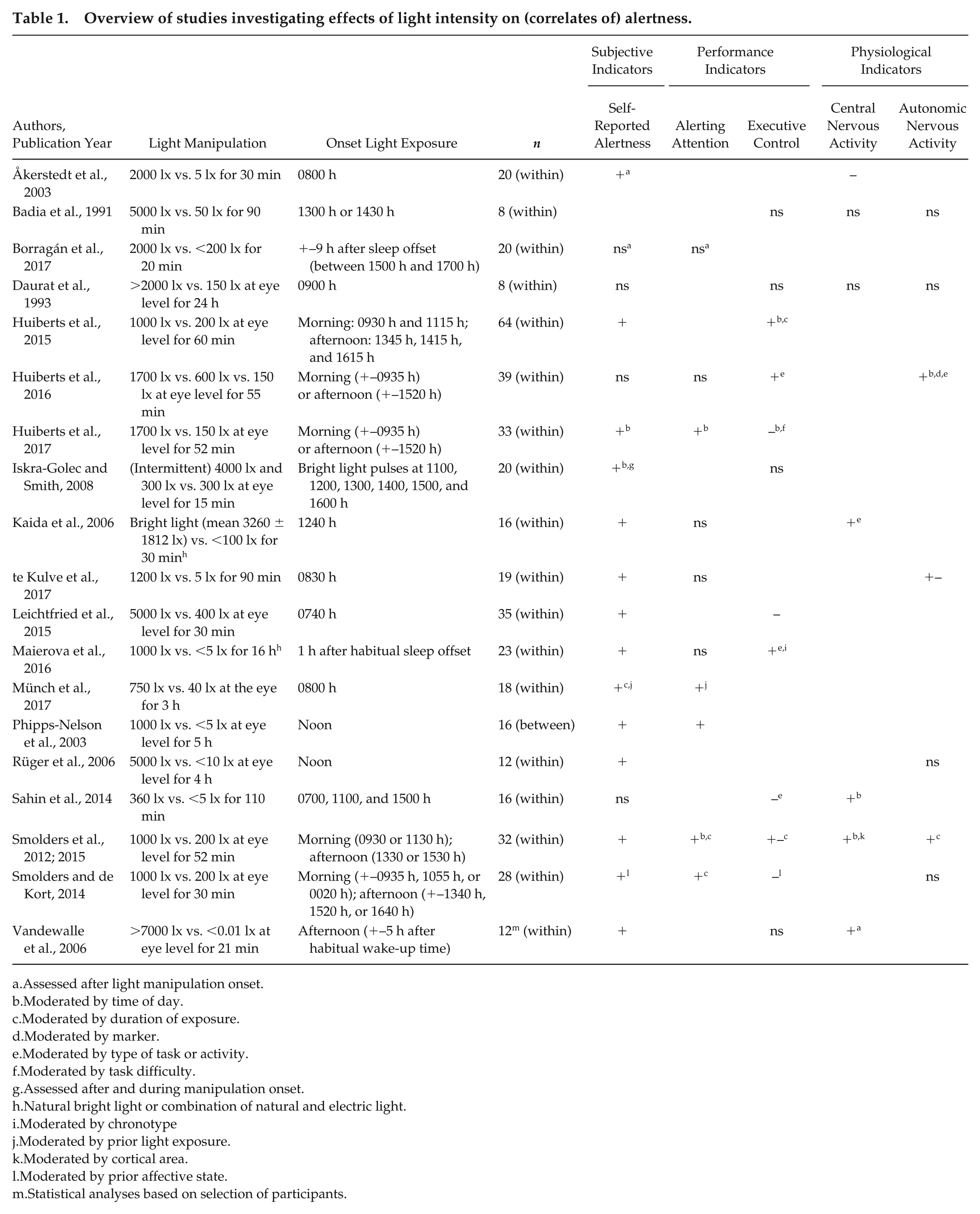
In total, 19 studies investigating diurnal NIF effects of the intensity of white light on alertness were included (Åkerstedt et al., 2003; Badia et al., 1991; Borragán et al., 2017; Daurat et al., 1993; Huiberts et al., 2015, 2016, 2017; Iskra-Golec and Smith, 2008; Kaida et al., 2006; te Kulve et al., 2017; Leichtfried et al., 2015; Maierova et al., 2016; Münch et al., 2017; Phipps-Nelson et al., 2003; Rüger et al., 2016; Sahin et al., 2014; Smolders and de Kort, 2014; Smolders et al., 2012; Vandewalle et al., 2006). Experiments (1) had to be performed during daytime and (2) had to investigate effects of different intensities of polychromatic white light on alertness (i.e., articles involving a comparison between a control condition and an experimental one, or between multiple levels of intensity during daytime). There were no other selection criteria (such as experimental design or subject inclusion criteria) to ensure a broad selection of studies.
Parameters of alertness were divided into the following categories: subjective indicators (self-reported alertness, as assessed with the KSS or visual analog scale); performance indicators, divided into sustained attention (performance on PVT) or executive control (e.g., performance on SART, N-back, Go-NoGo); and physiological indicators, which were split into central nervous activity (EEG) and autonomic nervous activity (skin temperature, core body temperature, heart rate, and heart rate variability). Effects of light on parameters of alertness are represented by “+” when a positive, significant effect of higher light intensity on the parameter was established, “–” when a negative effect of light was determined, “+/–” when mixed outcomes within the category were reported, and “ns” when nonsignificant effects of light intensities on alertness were established. Moreover, potential moderators, such as timing or duration of light exposure or prior conditions, are indicated by superscript letters in Table 1.
Overview of studies investigating effects of light intensity on (correlates of) alertness.
Assessed after light manipulation onset.
Moderated by time of day.
Moderated by duration of exposure.
Moderated by marker.
Moderated by type of task or activity.
Moderated by task difficulty.
Assessed after and during manipulation onset.
Natural bright light or combination of natural and electric light.
Moderated by chronotype
Moderated by prior light exposure.
Moderated by cortical area.
Moderated by prior affective state.
Statistical analyses based on selection of participants.
From all included studies, there were 3 studies that showed positive effects of higher intensities of light during daytime on all parameters and correlates of alertness used in that study (Huiberts et al., 2015; Münch et al., 2017, Phipps-Nelson et al., 2003). Thirteen other studies reported positive effects on some, but not all, parameters of alertness employed in the study (Åkerstedt et al., 2003; Huiberts et al., 2016, 2017; Iskra-Golec and Smith, 2008; Kaida et al., 2006; te Kulve et al., 2017; Leichtfried et al., 2015; Maierova et al., 2016; Sahin et al., 2014; Smolders and de Kort, 2014; Smolders et al., 2012; Vandewalle et al., 2006). Three studies reported no significant effects of bright light during the daytime on any of the measures included in the study (Badia et al., 1991; Borragán et al., 2017; Daurat et al., 1993).
In total, there were 18 studies using self-reported sleepiness, of which 14 reported significant effects and 4 nonsignificant effects of bright light exposure. Significant effects were reported to be moderated by time of day, duration of exposure, prior light exposure, and affective state. Performance measures were used in 17 of the included studies, of which 10 investigated the effects of light on alerting attention and 12 on executive control tasks. Five of the 10 studies using alerting attention tasks reported significant improvements in sustained attention under bright light, moderated by factors such as time of day, prior light exposure, and duration of exposure. The remaining 5 studies reported no significant effects. Outcomes on executive control tasks revealed mixed results, with positive (3 studies), nonsignificant (4 studies), mixed (1 study), and negative effects (4 studies). Results on performance measures for executive control were moderated by time of day, duration of light exposure, task difficulty, type of task or activity, chronotype, and/or prior affective state. Physiological parameters were used in 11 of the included studies, 4 investigating central nervous activity, 4 autonomic nervous activity, and 3 both central and autonomic nervous activity. Positive (4 studies), negative (1 study) or nonsignificant (2 studies) effects were reported on central nervous activity, moderated by time of day, marker, type of task or activity, duration of light exposure, and cortical area. Mixed results were found on parameters of autonomic nervous activity, with 2 studies reporting positive effects, 4 showing nonsignificant effects, and 1 reporting mixed effects. Results on autonomic nervous activity were moderated by time of day, duration of exposure, marker, and type of task or activity.
There are many differences between studies (such as experimental designs, light conditions, when and how alertness was assessed, and subject inclusion criteria) that have not been taken into account when including an article for this analysis. These factors might (positively or negatively) affect the outcome of a study. Possibly due to these differences in research paradigms, the overview of literature created seems to suggest that daytime effects of polychromatic white light on alertness are inconclusive in multiple studies using diverse experimental designs and lighting conditions. Nevertheless, the most consistent effect of daytime exposure to more intense light has been determined in subjective alertness, which shows a positive outcome in about three-fourths of the included studies (Åkerstedt et al., 2003; Huiberts et al., 2016, 2017; Iskra-Golec and Smith, 2008; Kaida et al., 2006; te Kulve et al., 2017; Leichtfried et al., 2015; Maierova et al., 2016; Phipps-Nelson et al., 2003; Rüger et al., 2006; Sahin et al., 2014; Smolders and de Kort, 2014; Smolders et al., 2012; Vandewalle et al., 2006).
Discussion and Conclusion
Light-induced effects on alertness have been studied extensively, especially during the night. However, humans have evolved as a diurnal species, and their physiological and psychological level of alertness is primarily lower in the late evening or at night compared with daytime hours (Cajochen et al., 2003; Dijk et al., 1992; Hull et al., 2003). Hormone levels, possibly affecting alertness, differ between night and day. Taking these factors into account, it might very well be that nighttime results of light on alertness are different from those during daytime. In fact, results found during the day are quite inconclusive, with mixed outcomes on various measures reported, particularly for the objective measures. As stated before, effects of light on subjective alertness seem to be the most conclusive based on the findings reported in the studies included in the overview. Moreover, performance on sustained attention and central nervous activation showed a more robust pattern than measures of performance on executive functioning tasks and autonomic nervous system activation. This might therefore also lead to the conclusion that alertness-enhancing effects of light are mostly reflected in subjective alertness, whereas effects on performance and physiology are less consistent. A recent literature review investigating alerting effects of light during both day- and nighttime also revealed that results are quite inconclusive, even during nighttime, especially in performance output measures (Souman et al., 2018). In fact, effects of light on subjective alertness may not always translate into statistically significant improvements in performance measures (e.g., Vandewalle et al., 2006; see also Fisk et al., 2018).
It is important to note that the performance measures for executive control and physiological measures for autonomic nervous activation employed in the studies were also heterogeneous. For instance, performance measures of executive control used in the studies included tasks probing inhibitory capacity and working memory, which also requires—in addition to alertness—other higher-order functions. It could also be that parameters other than subjective alertness are more vulnerable to differences in experimental design, in terms of timing and duration of the lighting manipulation, light history, and subject inclusion. On the other hand, a possible explanation for more consistent effects of light on subjective measures of alertness may be that recall bias and placebo effects (expectancy of outcomes and socially desirable behavior) influenced participants’ subjective ratings. This is particularly relevant given the fact that participants are generally not blind to a light manipulation. Since the 19 studies that have been included in this overview are inhomogeneous in terms of experimental design and intensity levels employed, this might contribute to overall inconclusive effects. For instance, differences in the spectral composition of white light may contribute to mixed results (e.g., facilitation or inhibition of alertness) due to potential activation of multiple neural pathways involved in NIF, as well as the image-forming processes, as a result of exposure to different (combinations) of wavelengths (Pilorz et al., 2016; Spitschan et al., 2014; Woelders et al., 2018). It is also important to mention that, despite reported inconsistent effects among measures within a specific study paradigm, publication bias toward studies including positive results could be expected. Sufficient or insufficient power could contribute to coincidental positive, null, or negative findings. The heterogeneity in research paradigms and substantial variation in power due to large differences in the number of participants and the number of measurements within participants make it difficult to compare the various studies and draw firm conclusions about the alerting potential of daytime exposure to bright light. In fact, this diversity calls for research investigating multiple intensity levels within one research paradigm with a relatively large sample size to determine the dose-dependent relationship between light intensity and alertness during daytime.
To date, the dose-response curve for alertness in response to light established at night has not been replicated yet (Cajochen et al., 2000). To our knowledge, there is, however, one study that fits a dose-response curve through existing subjective sleepiness data of both night- and daytime studies (Hommes and Giménez, 2015). Results of this study confirmed a dose-dependent relationship between subjective alertness and light intensity. Whether a similar pattern can be established for daytime effects only is still unknown. Most laboratory studies performed during daytime have investigated the effects of light intensity by comparing 2 or 3 light conditions. To determine whether light can indeed improve alertness during the course of the day in a comparable, dose-dependent manner as during the night, designing a systematic approach investigating the effects of a large range of light intensities on measures and correlates of alertness within one paradigm over the course of the day is one of the next steps in generating a dose-response curve for alertness. This has been done independently by both the Chronobiology Unit of the University of Groningen and the Human-Technology Interaction group at the Eindhoven University of Technology. Both research groups could not determine a clear dose-response relationship between light intensity and alertness. These research articles of Lok et al. and Smolders et al. can be found in this issue.
Footnotes
Conflict Of Interest Statement
The authors have no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.