
Abstract
Neurons in the suprachiasmatic nucleus (SCN) exhibit circadian variation in their neuronal activities. Here, we show that their input synapses undergo daily regulation. We discovered daily variation in the frequencies, but not the peak amplitudes, of miniature postsynaptic currents (mPSCs) for both glutamatergic and GABAergic synapses. In addition, the changes in the frequencies of mPSCs from glutamatergic and GABAergic synapses were antiphasic: An increase in the glutamatergic mPSC frequency and a decrease in the GABAergic mPSC frequency co-occurred during the dark phase of the light-dark cycle. Through optogenetic stimulation of presynaptic glutamatergic axonal terminals, we demonstrated that the elicited quantal size glutamatergic PSC could evoke action potentials in postsynaptic SCN neurons with higher probability during the light phase than during the dark phase. To investigate whether the intrinsic clock or the light-dark cycle mediates the daily rhythm of input synapses, we examined the mPSCs under light masking and skeleton photoperiod schemes. A prolonged light pulse (3 h) at the beginning of the subjective night suppressed the changes in mPSCs frequencies for both glutamatergic and GABAergic synapses; under the skeleton photoperiod, no significant differences were observed in mPSCs frequencies between the subjective day and night. Furthermore, we discovered that the plasticity of these concurrent quantal events occurred primarily in vasoactive intestinal peptide (VIP) neurons. This work provides evidence for light-dependent synaptic plasticity at glutamatergic and GABAergic input synapses of ventral SCN neurons. This concurrent synaptic plasticity could serve as one mechanism for regulating the responsiveness of SCN neurons to the wide range of light input over the day and night.
The suprachiasmatic nucleus (SCN) is considered the master clock for coordinating various biological oscillators in both the central nervous system and peripheral organs (Welsh et al., 2010; Dibner et al., 2010). The intrinsic rhythm of the SCN is maintained by intracellular autoregulatory transcription-translation feedback loops (Reppert and Weaver, 2002; Partch et al., 2014). To achieve a stable phase relationship between the master clock and daily environmental cycles (e.g., light, food, activities, etc.), the environmental input to the SCN adjusts the phase of the master oscillator, a process called entrainment (Daan, 2000). The major afferents to the SCN include retina, intergeniculate leaflet (IGL), and median raphe (Morin and Allen, 2006; Colwell, 2011). Retinal ganglion cells, primarily the melanopsin-expressing retinal ganglion cells (mRGCs), carry photic information of the environment and form glutamatergic synapses onto SCN neurons (Moore et al., 1995; Hattar et al., 2002; Panda et al., 2003). They also release pituitary adenylate cyclase activating polypeptide (PACAP) as a neuromodulator on a slow time scale (Hannibal, 2002; Morin and Allen, 2006). The IGL is considered to be a site where photic and nonphotic entrainment signals integrate, sending output to the ventral SCN via GABAergic synapses and coreleased neuropeptide Y (NPY) (Morin and Blanchard, 2001). The glutamatergic and GABAergic synapses impose transient changes in the membrane potential of the postsynaptic neurons, thus playing important roles in regulating the excitability and electrical output of SCN neurons. Activation of glutamatergic synapses in the SCN can induce phase shifts of circadian activity rhythms, while blockade of glutamate transmission in the SCN leads to impaired photic entrainment (Ebling, 1996; Gompf et al., 2015). GABA transmission in the SCN has 2 sources. One is from afferent nuclei like the IGL, and the other is from local SCN neurons, including feed-forward inhibition elicited by retinal input. Therefore, the functions of GABA transmission in the SCN are diverse. GABA can shift the phase of the clock (Ralph and Menaker, 1985; Albus et al., 2005) and reset and modulate synchrony of SCN neurons (Liu and Reppert, 2000; Albus et al., 2005; Han et al., 2012; Evans et al., 2013; Freeman et al., 2013).
One remarkable property of glutamatergic and GABAergic synaptic transmission in the SCN is that it exhibits circadian variation (Cui and Dyball, 1996; Lundkvist et al., 2002; Itri et al., 2004). More glutamatergic synaptic innervation has been observed during the subjective day compared with the night (Girardet et al., 2010), consistent with a higher frequency of spontaneous glutamatergic postsynaptic currents during the day (Lundkvist et al., 2002). In contrast, more NMDA components were found from glutamatergic synapses during the subjective night compared with the day (Pennartz et al., 2001). The frequency of spontaneous GABAergic responses peaked at the day-night transition (Itri et al., 2004), while GABAergic synapses may toggle between excitatory and inhibitory synapses within a day (Wagner et al., 1997; De Jeu and Pennartz, 2002; Choi et al., 2008). Most of the studies have examined spontaneous and evoked synaptic inputs to the SCN, but whether miniature synaptic events undergo circadian variation is unknown. Spontaneous miniature synaptic events provide background inputs to postsynaptic neurons. The functional roles of mini-events have been proposed as regulating the excitability of postsynaptic neurons (Lu and Trussel, 2000; Farrant and Nusser, 2005; Kombian et al., 2000), their spike output (Carter and Regehr, 2002; Sharma and Vijayaraghvan, 2003), and their synaptic efficacy in the absence of presynaptic action potentials (McKinney et al., 1999; Sutton and Schuman, 2006; Turrigiano, 2012). Modifications in the frequency and amplitude of mini-events could come from variation in presynaptic calcium levels (Madden and Kloot, 1978; Goswami et al., 2012; Shoudai et al., 2010; Llano et al., 2000), activity-dependent long-term plasticity (Jin et al., 2012; Hawkins, 2013), and neuromodulators (Sharma and Vijayaraghvan, 2003; Zhang et al., 2009; Jakawich et al., 2010).
In this study, we recorded miniature postsynaptic currents (mPSCs) of neurons in the ventral (core) part of the SCN, the input layer of the SCN (Moore and Silver, 1998). We examined the daily variation in mPSCs for both glutamatergic and GABAergic synapses and found distinct plasticity of quantal release frequency for the 2 types of synapses. Additionally, this plasticity of quantal events appeared only in VIP neurons of the ventral SCN. We further investigated whether light input or the intrinsic clock was responsible for this rhythmic synaptic plasticity. Our findings suggest that light modulates the frequency of quantal events at both excitatory and inhibitory synaptic inputs to the SCN.
Materials and Methods
Animals
All experimental protocols were approved by the institutional animal care and use committee of the University of Science and Technology of China (USTC). The C57/BL6 mice (n = 33; 2-3 months) were purchased from the laboratory animal facilities of AnHui Medical University. The VIP-ires-Cre::Rosa-GCaMP5 mice were bred by crossing VIP-ires-Cre (stock 010908; Jackson Laboratory, Bar Harbor, ME) with Rosa-GCaMP5 for at least 2 generations before use (kindly provided by Prof. Xiaohong Xu from the Institute of Neuroscience Shanghai). Upon arriving in our laboratory, control mice and VIP-ires-Cre::Rosa-GCaMP5 mice were housed with littermates on a 12/12-h light-dark (LD) cycle (lights on at 0400 h and off at 1600 h) with ad libitum access to food and water at room temperature (~25 °C). Because their original 12/12-h LD cycle was with lights on at 0800 h and off at 2000 h, the animals were allowed to acclimatize to the 4-h shift for at least a week. Animals in the negative masking group were given an additional 3-h exposure to light from ZT 13 to ZT 16. Recordings were done at ZT 17-20 on the same day. For the skeleton photoperiod group, mice were kept under darkness except for two 30-min pulses of light at ZT 0-0.5 and ZT 11.5-12 each day. They were allowed to acclimatize to this light scheme for at least 2 weeks, and 4 animals were given running wheels to monitor their locomotor activities during this period. The ambient light intensity in housing chambers was 1382 lux.
Brain Slice Preparation
Male and female mice (2-3 months) were anesthetized with 0.04% isoflurane and then decapitated. The brain was quickly extracted and placed in ice-cold N-methyl-
Electrophysiological Recordings
The acute SCN slice was transferred to the recording chamber with a constant perfusion of oxygenized ACSF at the rate of 3 mL/min. Neurons were visualized using an upright microscope (SS60, Scientifica, Clarksburg, NJ) with a 40× water immersion objective at room temperature (~25 °C). Neurons in the ventral SCN were chosen for electrophysiological recording. VIP-expressing neurons were visualized under a fluorescence microscope. Data were collected using Axopatch 700B patch clamp amplifier, Digidata-1440A data acquisition system, and pCLAMP 10.6 software (Axon CNS, Sunnyvale, CA). The patch pipette electrodes (OD = 1.5 mm, Sutter Instrument Co., Novato, CA) were pulled with Model P-1000 puller (Sutter Instrument) to a final tip resistance of 4 to 7 MΩ. For glutamatergic mPSCs recording, the electrodes were filled with a Cesium-based internal solution containing (in mM) 135 CsMethane sulfonate, 10 CsCl, 10 HEPES, 5 MgATP, 0.5 NaGTP, and 1 EGTA (pH was adjusted to 7.2 ± 0.1 using CsOH, and the osmolarity was adjusted to 310 ± 5 mOsmol/L using CsMethane sulfonate). The bath solution was ACSF with 50 µM Picrotoxin (PTX, Tocris, Abingdon, UK) and 400 nM tetrodotoxin (TTX, Sigma, St. Louis, MO). For GABAergic mPSC recording, the electrodes were filled with a high-chloride cesium-based internal solution containing (in mM) 140 CsCl, 10 HEPES, 5 Mg-ATP, 0.5 Na-GTP, and 1 EGTA (pH was adjusted to 7.2 ± 0.1 using CsOH, and the osmolarity was adjusted to 310 ± 5 mOsmol/L using CsCl). The bath solution was ACSF with 20 µM CNQX (Sigma), 20 µM AP-5 (Sigma), and 400 nm TTX. Cells were held at −70 mV and recorded in a whole-cell voltage-clamp mode. The pipette junction potential was not corrected. One sweep lasted 2 min, and between sweeps the access resistance and the leak current were checked. The access resistance was typically below 20 MΩ and the leak current was below 50 pA. If they exceeded these values, the recording would be terminated. Data were sampled at 20 kHz and low-pass filtered with cut-off frequency at 2 kHz.
Optogenetic Stimulation of Glutamatergic Axonal Terminals
Mice expressing Channelrhodopsin-2 (ChR2) on the membrane of mRGCs were bred by crossing OPN4-Cre (kindly offered by Prof. King-Wai Yau from John Hopkins University) with Rosa-ChR2 (RCL-CHR2(H134R)/EYFP) transgenic mice (Ai32, Stock No.012569; Jackson Laboratory) for at least 2 generations before use. Brain slices were prepared under dim red light. Blue LED light at 470 nm was given through the imaging objective (40×, NA 0.8). Slice visualization and patch clamp were done under infrared light with a CCD camera. When a whole cell voltage clamp configuration was obtained, a 2-msec-long light pulse with 87.98 mW/mm2 intensity was given onto the ventral SCN to activate ChR2. The light intensity was then adjusted to yield the minimum postsynaptic current with success rate over 50%. Once the minimum stimulation light intensity was determined, single pulse stimulations were repeated every 20 sec for at least 10 times. The recording mode was then changed to current-clamp mode to record membrane potential under the same light stimulation conditions.
Wheel-running
Two- to three-month-old male or female mice were individually housed in cages equipped with running-wheels and ad libitum access to food and water at room temperature (~25 °C). Cages were made of transparent-frosted acrylic (TFA) glass boards, so animals could see each other. Wheel-running data were collected by a custom-made device counting the number of wheel revolutions per second and then analyzed with custom-written software using Matlab (MathWorks, Natick, MA). The amount of wheel running activity was calculated as the total number of wheel revolutions in 5-min bins.
Data Analysis and Statistics
For detection of miniature events, recording traces were further low-pass filtered with a Gaussian filter to 1 kHz in Clampfit 10.7 (Axon CNS). An event template was generated by averaging 20 visually detected events, and then an event detection routine provided by Clampfit went through the whole trace and detected all the events that statistically matched the kinetics of the template. Following this automatic detection, we applied human visual inspection to evaluate the quality of the detection, and if the total number of false-positive and true-negative events exceeded 10% of the number of detected events, the entire process would be repeated with a new template. The peak amplitude of each detected event was measured as the local minimum of the peak from the baseline. The frequency of the miniature events was calculated by dividing the number of events by the total time. Data were presented as mean ± SEM. The n values indicated in the text were the number of cells recorded. The mean frequency of mPSCs was the average of miniature event frequencies from each cell. The mean mPSC amplitude was the average of the mean peak amplitudes of each cell. Evoked postsynaptic currents by activating ChR2 were detected as successful if the peak amplitude was larger than 2 times the standard deviation of the baseline. Only when the mean amplitude of the evoked synaptic currents was smaller than 10 pA were they considered as quantal-size EPSCs. To calculate the percentage of evoked action potentials by the quantal-size EPSCs, action potential occurrences were counted in a time window of 40 msec after the onset of the light stimulation. Wilcoxon rank tests were performed in Igor pro (WaveMetrics, Portland, OR), and the 2-tailed p value was presented. In the figures, asterisks are used to indicate the p values (*p < 0.05, **p < 0.01, and ***p < 0.001).
Results
Variation of Miniature Postsynaptic Responses over the Light-dark Cycle
We performed whole cell patch clamp recordings in neurons from acute SCN slices (Fig. 1A) during the light phase and dark phase of the LD cycle (Fig. 1B). Recordings of glutamatergic mPSCs were carried out in the presence of TTX and picrotoxin, a GABAA receptor antagonist. The frequency of glutamatergic mPSCs was significantly higher in the dark phase (0.57 ± 0.12 Hz, n = 27) than in the light phase (0.24 ± 0.05 Hz, n = 24). Their peak amplitudes were not significantly different between the light (7.29 ± 0.47 pA, n = 24) and dark phases (6.93 ± 0.51 pA, n = 27) (Fig. 1C). When GABAergic mPSCs were recorded, TTX and glutamate receptor antagonists (CNQX and AP5) were present in the bath. We raised the chloride concentration in the pipette solution to have inward and amplified chloride currents through ionotropic GABAA receptors. The frequency of GABAergic mPSCs was significantly reduced during the dark phase (1.14 ± 0.14 Hz, n = 29) compared with the light phase (3.56 ± 0.98 Hz, n = 13), and their peak amplitudes were not significantly different between the light (42.43 ± 6.38 pA, n = 13) and dark phases (35.10 ± 3.34 pA, n = 29) (Fig. 1D). Optogenetic stimulation of the axonal terminals from mRGCs in the ventral SCN resulted in evoked postsynaptic currents under the voltage clamp mode and postsynaptic potentials under the current clamp mode in SCN neurons. The minimum stimulation could evoke quantal size PSCs during the light and dark phases with similar success rate, while it evoked action potentials with an average probability of 16% during the light phase and 0% during the dark phase (Fig. 1E).
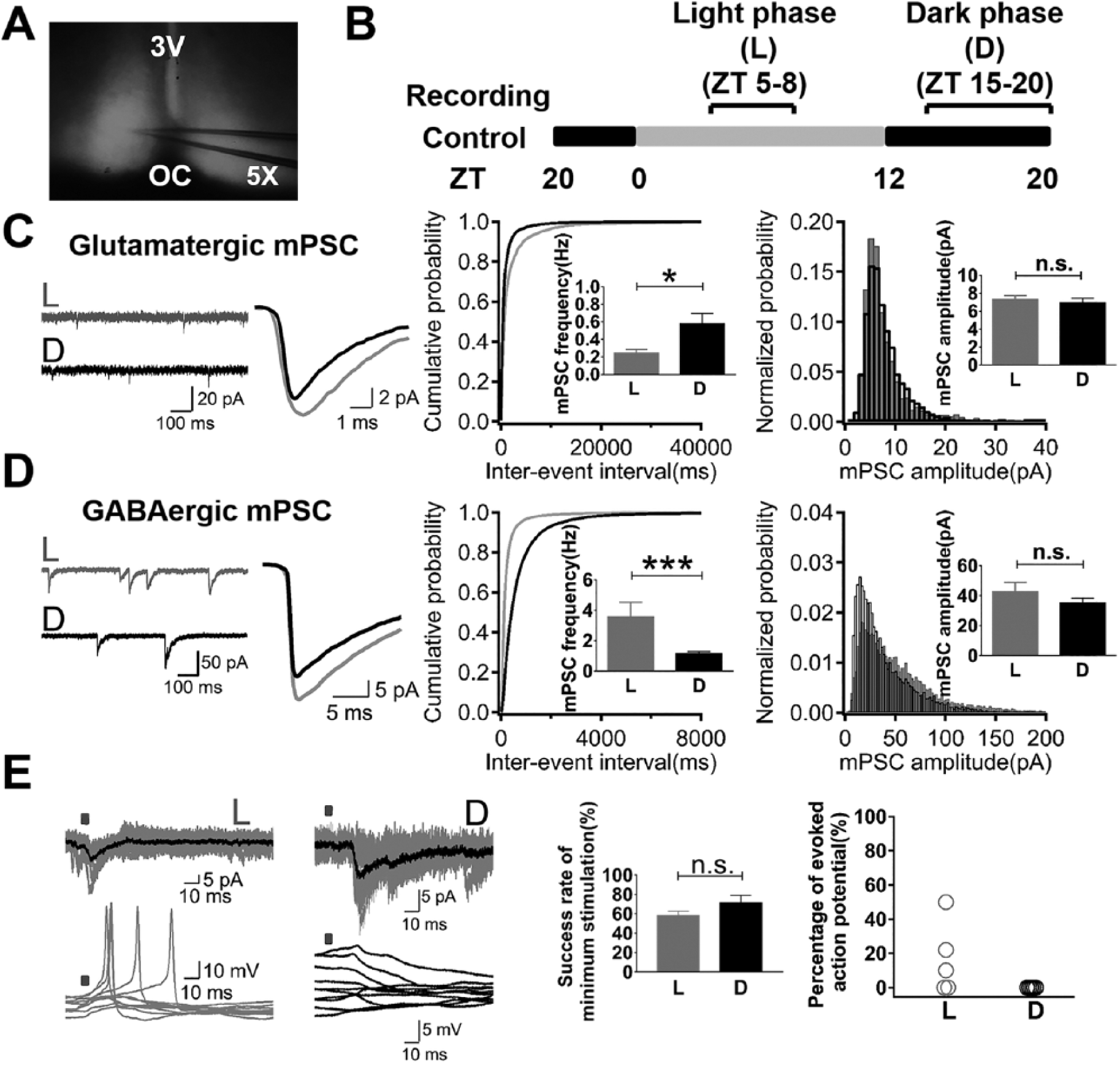
Increased glutamatergic quantal release and decreased GABAergic quantal release in the dark phase of the LD cycle. (A) A light transmission view of an acute SCN brain slice during the patch clamp recording. (B) Light phase and dark phase for recording in the LD cycle. (C) Recordings of glutamatergic mPSCs showed an increased frequency in the dark phase but no difference in the peak amplitude over the LD cycle. (D) Recordings of GABAergic mPSCs showed a decreased frequency in the dark phase but no difference in the peak amplitude over the LD cycle. (E) Optogenetic stimulation of mRGCs’ axonal terminals evoked miniature-size PSCs in SCN neurons. The minimum stimulation could evoke miniature-size PSCs with similar success rate between day and night; however, the probability of evoking action potentials in SCN neurons was higher in the light phase than the dark phase. 3V, third ventricle; OC, optic chiasm.
The Role of Light in Modulating the Plasticity of Synaptic Quantal Events
To test whether the plasticity of synaptic quantal events was a consequence of the LD cycle or an intrinsic circadian rhythm, we applied a 3-h light pulse to the animals from ZT 13 to ZT 16 (Fig. 2A). Animals exhibited negative masking (NM) of their locomotor activity during the light pulse (Fig. 2B). We recorded glutamatergic and GABAergic mPSCs from the acute SCN slices of these animals after the negative masking light pulse (ZT 17-20) and compared them with recordings from the similar time period without the light pulse. The frequency of glutamatergic mPSCs after the negative masking (NM: 0.11 ± 0.02 Hz, n = 13) was significantly lower than that from the normal dark phase (D: 0.48 ± 0.12 Hz, n = 14). No significant differences in the peak amplitudes were observed (D: 6.58 ± 0.72 pA, n = 14 vs. NM: 8.66 ± 1.03 pA, n = 13) (Fig. 2C). The frequency of GABAergic mPSCs after the negative masking (NM: 2.85 ± 0.58 Hz, n = 12) was increased to a level similar to that from the light phase (L: 3.56 ± 0.98 Hz, n = 13). Peak amplitudes were not significantly different compared with those recorded during the normal dark phase (D: 42.43 ± 6.38 pA, n = 13 vs. NM: 55.55 ± 7.88 pA, n = 12) (Fig. 2D). Thus, a prolonged acute light pulse given in the dark phase turned the frequency of glutamatergic mPSCs down and that of GABAergic mPSCs up to a similar level to that in the light phase.
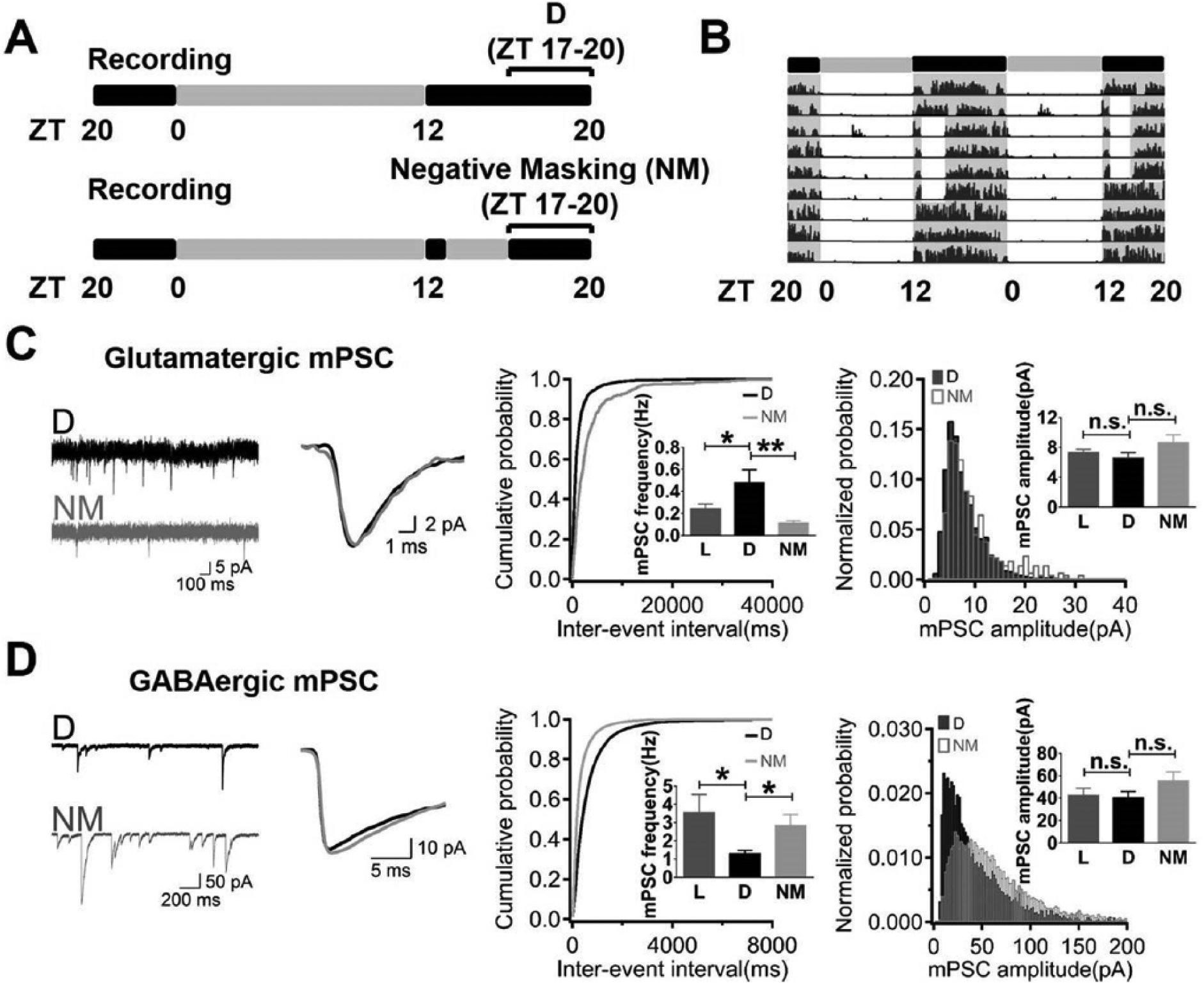
Prolonged light pulse during the subjective night restored synaptic quantal release to the level of the day. (A) Negative masking (NM) was implemented as a 3-h light pulse from ZT 13 to ZT 16. (B) A representative 48-h actogram showed the negative masking behavior during the light pulse in the subjective night and its recovery after returning to the LD cycle. (C) Recordings of glutamatergic mPSCs showed a decreased frequency after the light pulse but no difference in the peak amplitude. (D) Recordings of GABAergic mPSCs showed an increased frequency after the light pulse but no difference in the peak amplitude.
Because the early dark phase light pulse likely phase-delayed the circadian clock, we further tested the contribution of the LD cycle to the variation in synaptic plasticity by examining miniature postsynaptic responses of SCN neurons from animals entrained to a skeleton photoperiod (using two 30-min light pulses at ZT 0-0.5 and ZT 11.5-12) (Fig. 3A). When we compared the frequencies of mPSCs between subjective day (ZT 4-8) and night (ZT 16-20), no significant differences were found (glutamatergic mPSCs: 0.18 ± 0.03 Hz, n = 15 [ZT 4-8] vs. 0.35 ± 0.18 Hz, n = 12 [ZT 16-20]; GABAergic mPSCs: 2.12 ± 0.38 Hz, n = 14 [ZT 4-8] vs. 1.46 ± 0.27 Hz, n = 15 [ZT 16-20]) (Fig. 3, B and C). Their peak amplitudes were also similar between the subjective day and night (data not shown).
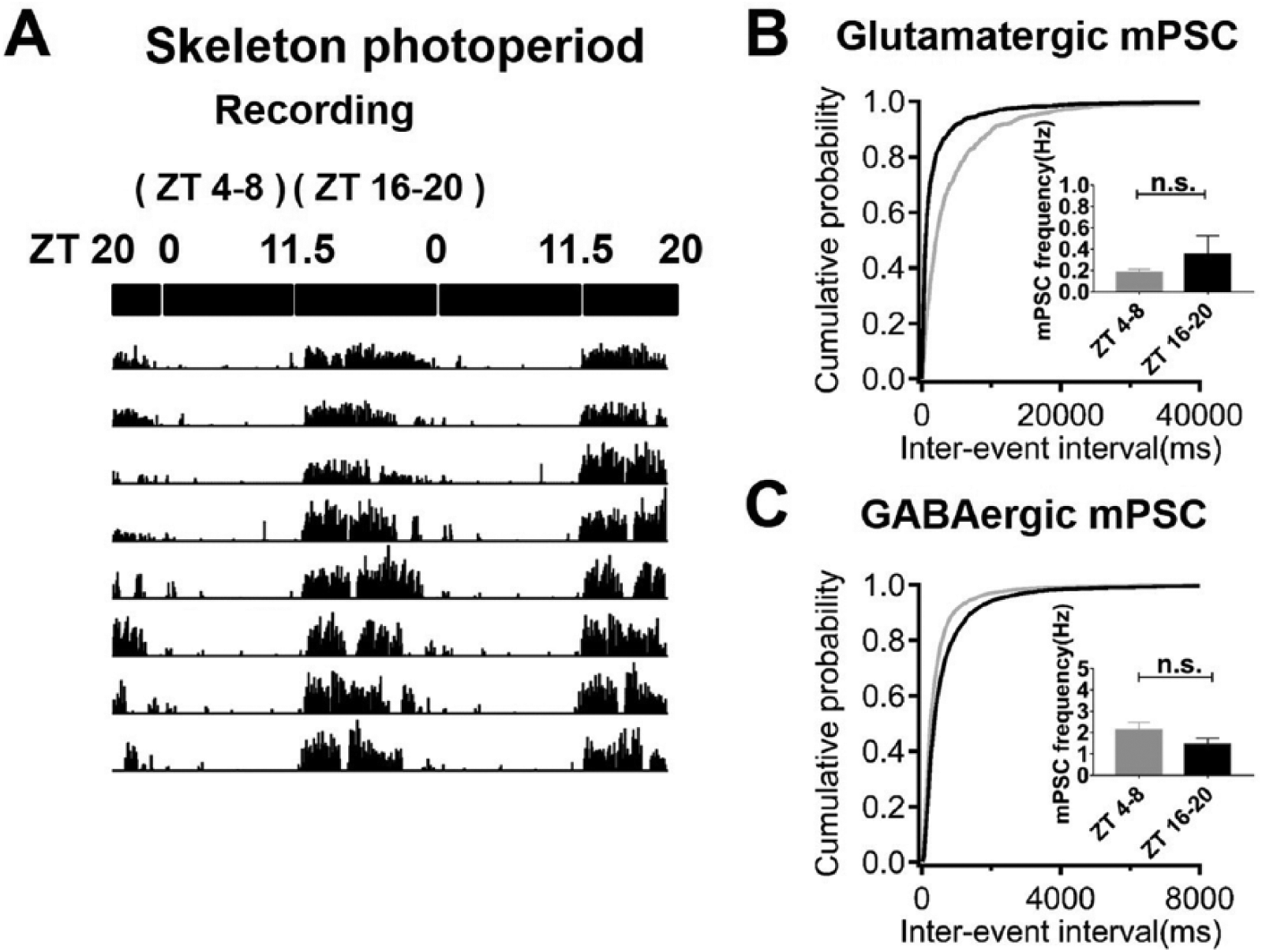
Lack of significant day-night differences in synaptic quantal release in the skeleton photoperiod. (A) The 48-h actogram showed the entrainment of locomotor activity to the skeleton photoperiod (light administered at ZT 0-0.5 and ZT 11.5-12). (B) The frequencies of glutamatergic mPSCs recorded at ZT 4-8 and ZT 16-20 were not significantly different. (C) The frequencies of GABAergic mPSCs recorded at ZT 4-8 and ZT 16-20 were not significantly different.
VIP Neurons Are Targets of Light-induced Quantal Event Plasticity
VIP neurons are the major input-receiving neurons in the ventral part of the SCN. We were able to identify VIP neurons using transgenic VIP-cre:GCaMP5 mice and record specifically from VIP neurons and non-VIP neurons in the ventral SCN. Under the LD cycle, VIP neurons exhibited a higher frequency of glutamatergic mPSCs and lower frequency of GABAergic mPSCs in the light phase compared with the dark phase (glutamatergic mPSCs: 0.42 ± 0.14 Hz, n = 14 [L] vs. 0.85 ± 0.28 pA, n = 11 [D]; GABAergic mPSCs: 1.01 ± 0.20 Hz, n = 11 [L] vs. 0.50 ± 0.10 Hz, n = 11 [D]), similar to the collective result from unidentified SCN neurons (Fig. 4, A and B). Their peak amplitudes did not show daily variation (data not shown). For non-VIP neurons, no significant differences were seen for either glutamatergic or GABAergic mPSCs over the LD cycle (Fig. 4, C and D).
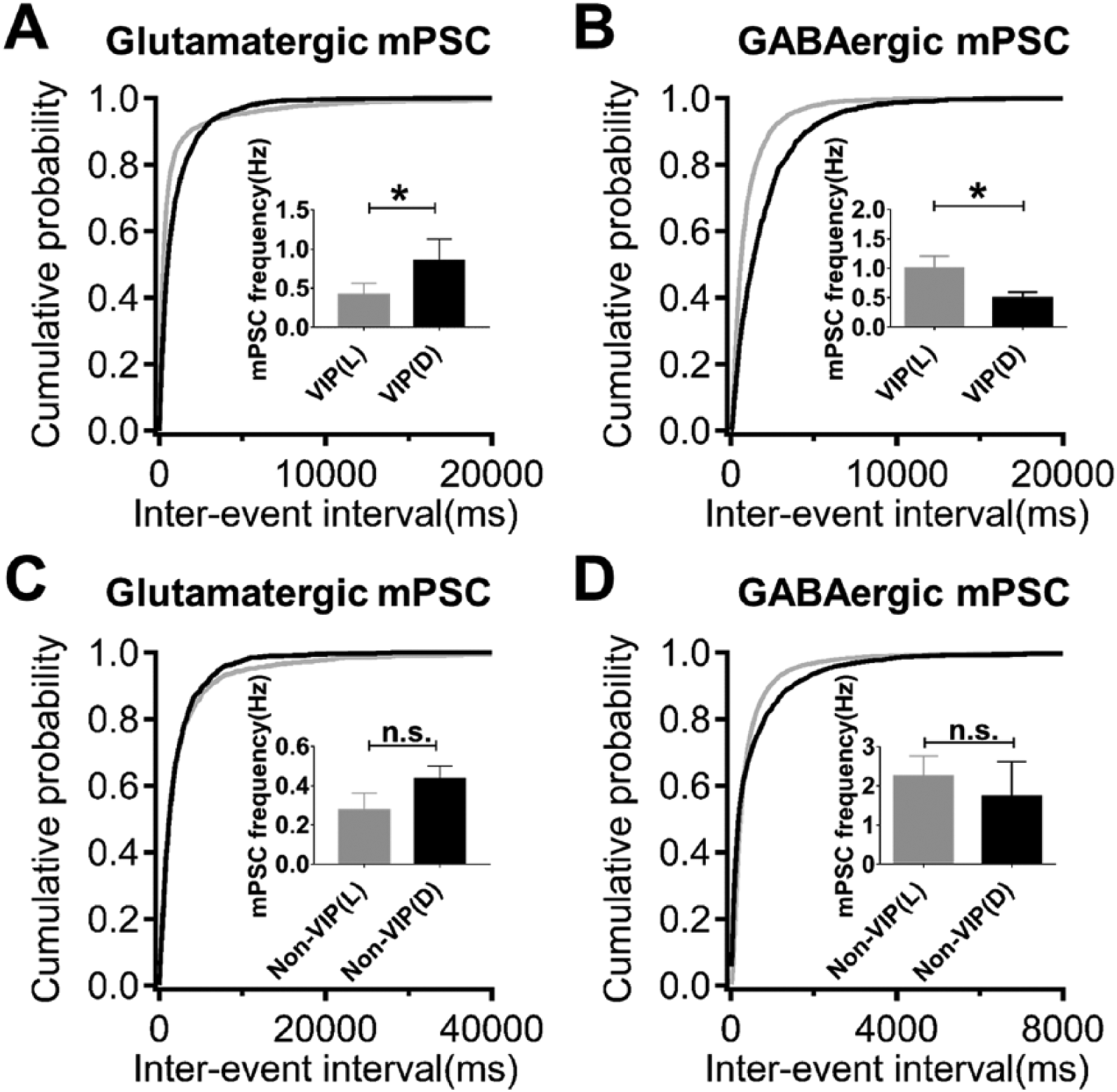
VIP neurons in the ventral SCN exhibit synaptic plasticity over the LD cycle but non-VIP neurons do not. (A) The frequency of glutamatergic mPSCs is higher in the light phase compared with the dark phase for VIP neurons in the ventral SCN. (B) The frequency of GABAergic mPSCs is lower in the dark phase compared with the light phase for VIP neurons. (C, D) The frequency of glutamatergic and GABAergic mPSCs did not differ over the LD cycle for non-VIP neurons.
Discussion
In this work, we have shown that the frequency of mPSCs of ventral SCN neurons exhibits daily variation, but their amplitudes do not vary across the day and night. We also demonstrated that miniature-size glutamatergic input could evoke action potentials in SCN neurons attributed to their high input resistance (Hermanstyne et al., 2016); therefore, plasticity of the quantal events could affect the output of SCN neurons over the daily cycle. Furthermore, the observed plasticity was antiphase for glutamatergic and GABAergic synapses, and they were both regulated by prolonged light input. VIP neurons are not the only neuronal type that receives retinal input (Fernandez et al., 2016); however, we found that light-induced synaptic quantal event plasticity appeared only in VIP neurons in the ventral SCN. The plasticity of quantal events described here could serve multiple functions in the processing of light information by the SCN, and it could also underlie the synaptic plasticity of evoked events. We will discuss the functional implications and potential mechanisms of this type of rhythmic plasticity.
Concurrent Plasticity of Quantal Events in Glutamatergic and GABAergic Synapses
Synaptic quantal release is a part of spontaneous release. It influences postsynaptic neurons transiently through changing the membrane potential, and it can induce long-term changes through tonic activation of calcium permeable postsynaptic receptors. The input resistance of VIP neurons is higher in the light phase compared with the dark phase (Hermanstyne et al., 2016), which is consistent with our finding that the quantal size glutamatergic input had a greater chance of evoking action potentials in the light phase than the dark phase. Therefore, the lower frequency of glutamatergic mPSCs during the day could improve the signal-to-noise ratio for excitatory photic input from the retina. SCN neurons exhibit a more quiescent state during the dark phase than the light phase (Schaap et al., 2003; Brown and Piggins, 2009), and they receive less spike-evoked excitatory inputs from the retino-hypothalamic tract (Girardet et al., 2010; Lundkvist et al., 2002). In the dark phase, SCN neurons need to be more sensitive to excitatory photic inputs, but their excitability is lower due to low input resistance. The increased frequency of excitatory quantal release may reflect an increase in the presynaptic release probability, thus enhancing the synaptic weight for photic input. In contrast, decreased GABAergic quantal release in the dark phase might indicate a decrease in both the background and evoked inhibition of SCN neurons, rendering them additionally more responsive to photic input. Some of the GABAergic synapses may turn excitatory during the night, which would further decrease the level of inhibition in the dark phase. In the light phase, the SCN receives a high level of photic input. The increased frequency of GABAergic inhibitory mPSCs could set a tonic inhibitory tone to balance the membrane potential of SCN neurons from the high level of excitation. It may also reflect an increase in evoked inhibition, especially feed-forward inhibition induced by photic input, which sets a gate for temporally coordinated photic input. Instead of being sensitive to weak light input during the dark phase, the SCN during the light phase needs to be selectively responsive to the change of light intensity; therefore, a coding strategy could use feed-forward inhibition to achieve a high dynamic range. Taken together, we speculate that the coherent plasticity of quantal events in glutamatergic and GABAergic synapses might act to modulate the level of inhibition onto the ventral SCN and regulate the responsiveness of SCN neurons, mainly VIP neurons, to the various levels of light input over the LD cycle. The possible significance of such a mechanism in the physiological context of active input from the retina will require further investigation.
Mechanisms Regulating the Two Types of Synapses
What are the mechanisms that coordinate the concurrent plasticity of glutamatergic and GABAergic synapses? Is there a common signal pathway? Is it the molecular clock or the input rhythm that plays a role? Our results suggest that light input is both necessary and sufficient for glutamatergic and GABAergic synaptic plasticity. Modifications in spontaneous quantal release frequencies could be the consequence of changes in the presynaptic vesicle release probability or the number of active synapses. Changes in the presynaptic release probability over the LD cycle are likely correlated with changes in membrane potential of the axonal terminals and their intracellular calcium levels (Awatramani et al., 2005). Activities of postsynaptic neurons can also modulate presynaptic release probability through retrograde signaling (Sharma and Vijayaraghvan, 2003; Zhang et al., 2009; Cheng et al., 2017). Particularly, brain-derived neurotrophic factor (BNDF) has been demonstrated to regulate the strength of photic glutamatergic signaling in the SCN (Liang et al., 1998; Michel et al., 2006; Baba et al., 2008; Jakawich et al., 2010). The glutamatergic and GABAergic presynaptic terminals in the ventral SCN presumably have higher activity levels and calcium concentrations in the light phase than the dark phase, attributed to the high levels of activity of retinal ganglion cells and SCN neurons in the light phase. Therefore, the increase of the GABAergic quantal release frequency in the light phase may be explained by presynaptic mechanisms, and the light-suppressed glutamatergic quantal release frequency is likely attributed to retrograde inhibition of presynaptic vesicle release. The expression of postsynaptic AMPA receptors can be regulated through spontaneous miniature synaptic release (McKinney et al. 1999; Sutton et al., 2004), resulting in changes in the synaptic efficacy or the number of active synapses. However, we did not observe changes in the size of the miniature quantal events over the LD cycle, suggesting AMPA receptor modification is unlikely to be involved in the rhythmic plasticity of synaptic quantal events. Under the skeleton photoperiod, the day-night rhythm of plasticity of synaptic quantal events was lost. We also observed that both the glutamatergic and GABAergic quantal release frequencies were low. Since the skeleton photoperiod experiments were performed 2 weeks after the acclimation of animals to this lighting schedule, there might be other homeostatic mechanisms that contribute to these synaptic properties.
Footnotes
Acknowledgements
We acknowledge support from the National Natural Science Foundation of China (31571073, 81401025 to J.B.; 81371066, 91432104 and 31322024 to T.X.; 81571293 to L.W.), from the Natural Science Foundation of Anhui Province (1508085MH185 to J.B.), and from the National Century Excellent Talents in University of China (gxbjZD2016034 to L.W).
Conflict of Interest Statement
The author(s) have no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.