
Abstract
The possession of a rhythm is usually described as an important adaptation to regular changing environmental conditions such as the light-dark cycle. However, recent studies have suggested plasticity in the expression of a rhythm depending on life history and environmental factors. Barn owl (Tyto alba) nestlings show variations in behavior and physiology in relation to the size of black feather spots, a trait associated with many behavioral and physiological phenotypes including the circadian expression of corticosterone and the regulation of body mass. This raises the possibility that individual spottiness could be associated with rhythmicity in sleep-wakefulness. Owlets showed ultradian rhythms in sleep-wakefulness, with a period length of 4.5 to 4.9 h. The period length of wakefulness and non-REM sleep was shorter in heavily compared to lightly spotted female nestlings, whereas in males, the opposite result was found. Furthermore, male and female nestlings displaying small black spots showed strong rhythmicity levels in wakefulness and REM sleep. This might be an advantage in a stable environment with predictable periodic changes in light, temperature, or social interactions. Heavily spotted nestlings displayed weak rhythms in wakefulness and REM sleep, which might enable them to be more flexible in reactions to unexpected events such as predation or might be a mechanism to save energy. These findings are consistent with previous findings showing that large-spotted nestlings switch more frequently between wakefulness and sleep, resulting in higher levels of vigilance compared to small-spotted conspecifics. Thus, nestlings with larger black feather spots might differently handle the trade-off between wakefulness and sleep, attention, and social interactions compared to nestlings with smaller black spots.
Keywords
Rhythmicity is occurring at every level in nature, from the rotation of the earth causing day-night patterns to the behavioral activity patterns of most animals, body temperature fluctuations, and rhythmic expression patterns of hormones such as melatonin and cortisol (Daan and Aschoff, 1975; Cassone et al., 1993; Zeman and Gwinner, 1993; Ingram et al., 1999; Foster and Kreitzmann, 2004; Williams et al., 2012). An adaptation to predictable changes in the environment allows animals to conduct appropriate behavior at the ecologically best time (Aschoff, 1981a), which can confer survival (DeCoursey et al., 2000; Spoelstra et al., 2016) and reproduction advantages (Greives et al., 2015).
Variability of rhythmicity in behavior and physiology has recently become the focus of many studies. For example, reindeer (Rangifer tarandus) living in the Arctic show a circadian rhythm of activity in periods when there is a light-dark cycle in spring and autumn but not when there is constant light (summer) or no sunlight (winter) 24 h a day (Van Oort et al., 2005). The suspension of rhythmicity in winter and summer might allow individuals to reduce energy expenditure in a nonrhythmic environment (Van Oort et al., 2005). Similarly, populations of the characin fish (Astyanax mexicanus) living in a completely dark cave environment all year long do not exhibit a circadian rhythm in metabolism in contrast to populations that do not live in caves. The absence of rhythmicity results in a net energy saving of up to 27% (Moran et al., 2014). Bumblebee (Bombus terrestris) queens display a circadian rhythm, but the intensity of this rhythm is dependent on life history (Eban-Rothschild et al., 2011). European blackbirds (Turdus merula) living in an urban habitat that is noisy and light polluted have a faster but less robust circadian rhythm of activity compared to conspecifics living in rural areas (Dominoni et al., 2013). These studies show the existence of plasticity in the expression of rhythms, which appear to be adaptive and related to life history and environmental conditions.
The existence of ultradian rhythms, that is, rhythms shorter than 24 h, and underlying physiological mechanisms are not well understood (Stephenson, 2013). Whereas circadian rhythms with a period length of about 24 h follow the light-dark cycle, ultradian rhythms do not correspond to any known environmental periodicity (Daan and Aschoff, 1981). Ultradian rhythms can be observed for behavior such as feeding, foraging, or locomotion, but they also occur in releasing patterns of hormones and in heartbeat frequency (Kimura and Tsai, 1984; Gerkema et al., 1993; Tornatzky et al., 1998). They are thought to closely express the internal physiological state of an individual (Berger et al., 2002). In mammals, the adjustment of circadian rhythms strongly relies on maturation of the hypothalamic suprachiasmatic nucleus (SCN), which is responsible for entrainment to the light-dark cycle, and on the development of corresponding neural and humoral signaling pathways (Weinert, 2005). However, the role of the SCN for the development of ultradian rhythms is not clear. In rats, a lesion of the SCN resulted in a complete loss of circadian and ultradian rhythms for wheel-running activity, indicating ultradian rhythms as submultiples of the circadian period (Wollnik and Turek, 1989). In contrast, ultradian rhythmicity in wheel-running activity and feeding of the common vole (Microtus arvalis) was still present after a lesion of the SCN, whereas the circadian rhythm was lost, indicating the presence of an autonomous ultradian oscillator outside the SCN (Gerkema and Daan, 1985). These two studies and further investigations in other species (Rusak and Zucker, 1979; Stupfel and Pavely, 1990; Ingram et al., 1999; Lumineau et al., 2001; Berger et al., 2002; Dowse et al., 2010) indicate that the understanding of the mechanism of ultradian rhythmicity is, to date, still unresolved.
Recent studies in the barn owl (Tyto alba) showed that nestlings differ not only in personality and physiology, depending on the degree of pigmentation (Ducrest et al., 2008; Van den Brink et al., 2012; Peleg et al., 2014), but also in the daily regulation of stress hormones (Roulin et al., 2010). When the mother was small spotted, nestlings had lower corticosterone levels in the morning and higher levels in the late afternoon (and vice versa when the mother was large spotted) (Roulin et al., 2010). Furthermore, in years characterized by poor rearing conditions, nestlings born from small-spotted mothers were heavier in the morning but lighter in the evening than nestlings from large-spotted mothers (Almasi and Roulin, 2015). Finally, nestlings born from small-spotted mothers switched less often from wakefulness to sleep and were less attentive towards the nest box’s entrance where parents predictably deliver food (Scriba et al., 2014). Sleep architecture and the daily regulation of several physiological and behavioral processes are therefore related to pigmentation of the individuals themselves (Roulin, 2009) and of their parents (Roulin et al., 2010; Almasi and Roulin, 2015; Scriba et al., 2014). This raises the question of whether rhythmicity is differently organized in differently colored individuals. Intrapopulation variations in rhythmicity have already been observed in mole rats (Heterocephalus sp.) in which some individuals display a strong rhythm while others are arrhythmic (Oosthuizen et al., 2003). Our aim in this study was to examine if rhythmicity is associated with melanin-based coloration.
Methods
Study Species and Area
We studied a population of wild barn owls in Switzerland breeding in nest boxes fixed to the external wall of barns. Hence, nestlings can perceive the light-dark cycle through the nest box’s entrance, and they can hear natural and human-made noise. Data were collected between May and October 2011 and between April and October 2012. We took a small blood sample to identify the nestling sex using molecular markers (Py et al., 2006), and the hatching date was determined using the wing length measured in the first days after hatching (Roulin, 2004b). To disentangle genetic and environmental effects on sleep states (Scriba et al., 2014), we exchanged the same number of eggs or hatchlings between randomly chosen pairs of nests with similar hatching dates (in 2011, we left some eggs in the nest of origin, while the other eggs of the clutch were exchanged between pairs of nests, whereas in 2012, we swapped entire clutches between pairs of nests). In 2011, 41 (20 males and 21 females) of the 66 nestlings were raised by foster parents, while 25 nestlings (11 males and 14 females) were not cross-fostered and hence raised by their biological parents. In 2012, 36 (11 males and 25 females) of the 42 nestlings were cross-fostered, and 6 (2 males and 4 females) nestlings stayed in their original nest. Barn owls in this area produce between 1 and 9 nestlings, which fledge at an age of approximately 55 days. Because incubation starts as soon as the first egg has been laid, nestlings hatch asynchronously about 2.5 days apart, generating a pronounced age hierarchy. The mother stays in the nest box throughout 24 h until the offspring are approximately 4 weeks of age, when they are able to thermoregulate and eat without maternal help. At that time, the parents come to the nest only at night to bring small mammals (Roulin, 2004a). Nestlings are active both during the light and dark period (Scriba et al., in press), and they spread their meals throughout 24 h by eating food that has been stored at night by their parents mainly during the first 4 h of the night (Roulin, 2001; Roulin, 2004a; Michelat and Giraudoux, 1992). Nestlings show complex social interactions not only by communicating vocally to negotiate priority access to food resources (Johnstone and Roulin, 2003) but also by sharing food actively and preening each other (Roulin et al., 2012, 2016). At night, each nestling produces in the absence of the parents, on average, 1279 ± 122 calls (up to 10,245 calls per night), implying that there is the potential for substantial noise disturbance (Roulin, 2002).
Assessment of Plumage Spottiness
Barn owls display eumelanin-based black spots of varying size on the tip of their ventral feathers. The diameter of black spots in a reference area (60 × 40 mm) on the breast was measured with a caliper to the nearest 0.1 mm at the age of about 55 days, when feathers are fully developed. This method is repeatable (0.84-0.92) (Roulin, 2004b). The mean spot diameter was calculated and used in statistical analyses.
Brain Activity Recordings
Brain activity was recorded in 2011 in 31 male and 35 female nestlings from 29 nests aged between 27 and 48 days (mean = 38.2 ± 0.5 days), and in 2012, an electroencephalogram (EEG) was recorded in 13 male and 29 female nestlings from 30 nests aged between 36 and 49 days (mean = 42.8 ± 0.6 days). We used minimally invasive subcutaneous wire electrodes in conjunction with a small neurologger (Neurologger 2; www.vyssotski.ch/neurologger2) (Vyssotski et al., 2009; Scriba et al., 2013a, 2013b). Briefly, part of the feathers on top of the head were removed, the skin was locally anesthetized (Gingicaine/tetracaine 754 mg/65 g; Gingi-Pak, Camarillo, CA, USA), and 5 stainless steel wire electrodes (0.13 mm diameter, with 2 mm of the insulation removed for exposure; Cooner Wire, Chatsworth, CA, USA) were inserted with a 23-gauge needle under the superficial layers of the skin. One electrode was placed over the posterior part of the visual hyperpallium (visual Wulst) of each brain hemisphere and referenced to a posterior electrode placed over the caudal nidopallium of the same hemisphere. The ground electrode was centered between the other electrodes. We connected all 5 electrodes to a data logger glued to the head and covered them with plastic protection to prevent the birds from removing the logger and electrodes. The data logger additionally contained a 3-dimensional accelerometer, which measured behavioral state–related head movements in all 3 axes (X, Y, Z). The logger was powered by 2 batteries (each 1.4 V and 310 mAh; zinc-air 13; Renata, Itingen, Switzerland) connected in series. The total weight was about 5 g, which represents 2.1% of the nestlings’ body mass. The EEG of both brain hemispheres was recorded at 200 Hz for up to 5 days; during the entire period, the nestlings stayed undisturbed together with siblings in the nest box and were naturally provisioned by their parents. The last 24-h period of the EEG was analyzed for each year by a different person using Somnologica software (Medcare, Reykjavik, Iceland). The EEG was scored for wakefulness, non-REM sleep, and REM sleep in 4-sec epochs (Scriba et al., 2013a, 2013b). As in earlier studies with barn owls, interhemispheric asymmetries in non-REM sleep intensity were very rarely observed (Scriba et al., 2013b), and usually, both hemispheres showed some level of non-REM sleep–related EEG activity simultaneously. Epochs with more than one state were scored according to the predominant state.
Rhythmicity Analysis
The time spent in each state (i.e., wakefulness, non-REM sleep, and REM sleep) was calculated for each 5-min period (rather than shorter periods of time) because this was shown to remove noise and clarify any periodicity in short time series (Dowse and Ringo, 1994; Dowse et al., 2010). The time series, consisting of the percentage of time spent in each state for each 5-min period over 24 h (Fig. 1), were then analyzed for the intensity of rhythmicity and the period length (τ, tau) of the sleep-wakefulness pattern by performing autocorrelations and maximum entropy spectral analysis (MESA) (Burg, 1967) implemented in a Fortran program (prediction error filter coefficient was set to 90) (Dowse, 2007). Autocorrelograms can be used to visualize periodic signals out of time series, and MESA is a high-resolution technique that is well suited for the analysis of short noisy time series (Ables, 1974; Dowse, 2009; Dowse et al., 2010). Tau values indicate the period length or duration of the rhythmic motive, that is, the time between 2 peak values of a parameter (Fig. 1). The maximal spectral density of the periodograms, produced by fast Fourier transformations with the software R (R Core Team, 2012), relates to the variance explained by a single frequency and hence is a measure of rhythmicity in time series, with higher values showing stronger single rhythmicity in the time series (Fig. 1). We used 24-h periods to determine ultradian rhythmicity in each individual. We did not analyze circadian periodicity since we only extracted 24-h periods of each recording.
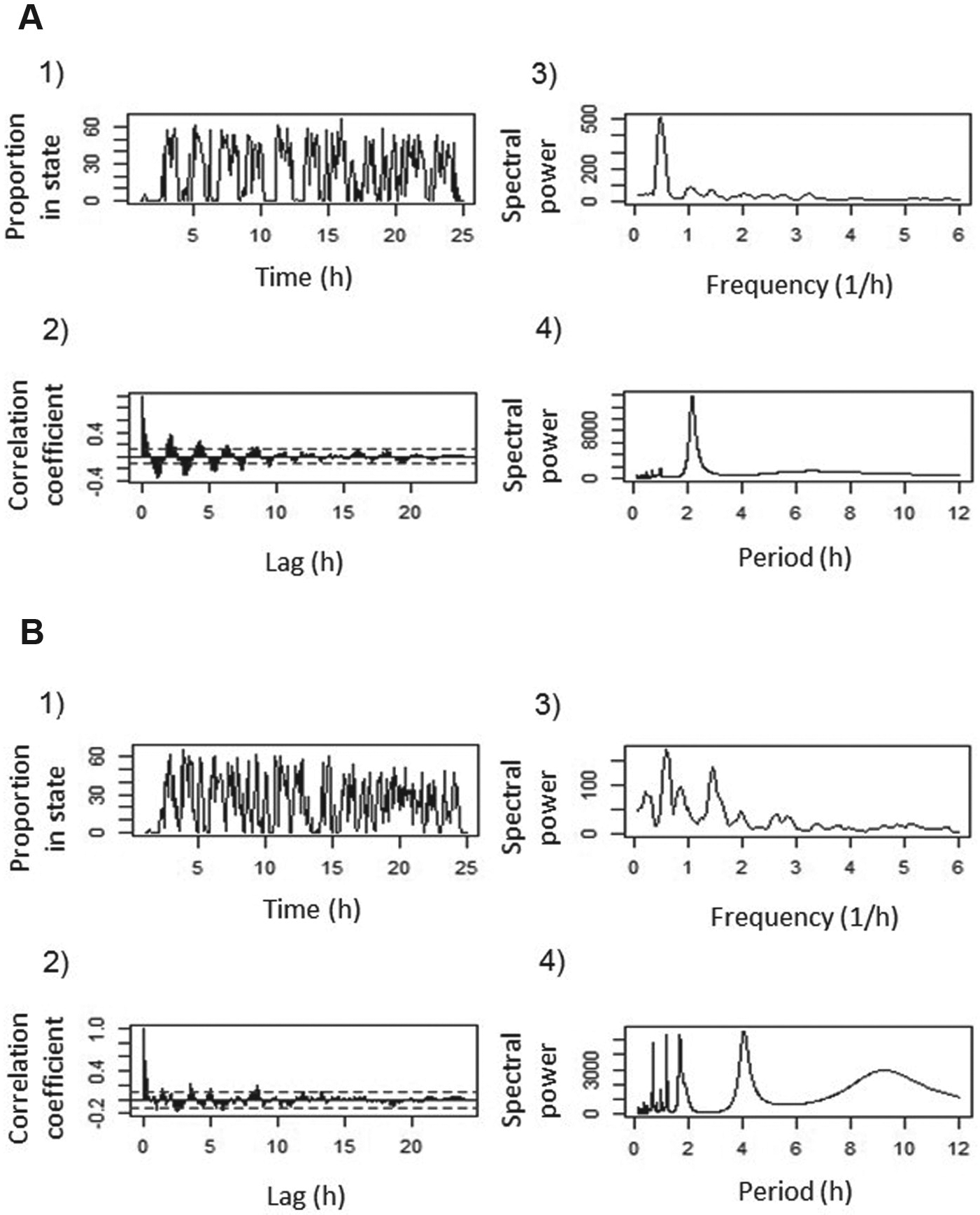
Non-REM sleep pattern (1) and corresponding autocorrelogram (2), periodogram (3), and MESA spectrogram (4) of a clearly rhythmic (A) and arrhythmic/multirhythmic (B) barn owl nestling. The period length tau can be determined from the autocorrelogram as the distance between 2 positive or negative peaks (for A, tau is 2.17 h, and for B, no single period length can be determined because several different oscillations appear to overlap). The spectral power of the highest peak in the periodogram represents the intensity of rhythmicity.
We considered a time series as significantly rhythmic when the maximal observed spectral density outranged the maximal spectral densities from 1000 data permutations of the same series. The calculation of data permutations was established as an effective method to detect periodicity in short time series (Dallmann et al., 2012; Krawczyk and Krapiec, 2010; Pardo-Iguzquiza and Rodriguez-Tovar, 2000; Ptitsyn et al., 2006; Mueller et al., 2012).
Statistical Procedure
Statistical analyses were conducted using the software JMP (version 10.0.0; SAS Institute, Cary, NC, USA). First, we compared rhythmicity values between years using the Student t test. We used Pearson correlations to look at the relationship between the period length and rhythm intensity of all 3 states but also to test whether these measures of rhythmicity are related to the percentage of time spent in each cerebral state, the mean duration of bouts, and the number of bouts performed during 24 h. We log transformed the period length and Box-Cox transformed rhythm intensity values (λ = 1.2) to obtain normally distributed model residuals. We performed linear mixed models with year and the nest of origin as random variables when we investigated the potential association between rhythmicity and plumage spottiness; this was necessary because we measured several siblings which have similar plumage spottiness (this trait is under strong genetic control). When testing whether nestlings sharing the same nest display a similar rhythm, we implemented the nest of rearing as a random factor. To control for potential effects of seasonal changes in the photoperiod on rhythmicity, we included the Julian date as a fixed effect in analyses. However, this variable was never significant, and hence, we removed it from the final models. Independent variables were nestling sex, age, number of siblings, the mean diameter of melanin-based black feather spots, and the interaction of plumage traits and sex. Because position in the within-brood age hierarchy and absolute age are significantly correlated (r = −0.58, n = 108, p < 0.0001), we did not include these 2 variables simultaneously in the same models. However, we verified that we obtain similar results if we replace absolute age with position in the within-brood age hierarchy; this was always the case. Note that the number of black spots was not related to the rhythmicity variables. We used only data of rhythmic individuals in the models with period length (as arrhythmic time series have none or several period lengths), whereas we used all individuals for analyses on rhythm intensity as it is measured on a continuous scale. We nevertheless compared plumage spottiness between rhythmic and arrhythmic individuals. Repeatability analyses were conducted using the method of Lessells and Boag (1987) to compare the rhythmicity of nest mates sharing the same nest. To standardize data, we applied the following formula: (values – mean)/SD. Tests were 2-tailed, with p < 0.05 considered significant. Values are expressed as mean ± SE.
Results
Rhythmicity in Relation to Cerebral States
To test whether sleep architecture (i.e., percentage of time spent in one cerebral state, bout duration, and number of bouts) (Table 1) was related to rhythmicity, we standardized values for each year so that we could pool the data obtained in 2011 and 2012. For all 3 states, the intensity of rhythmicity in each cerebral state was positively associated with the percentage of time spent in this state and the mean bout duration but negatively with the number of bouts per 24 h (Table 1). These relationships were particularly strong for REM sleep. In contrast, for all 3 cerebral states, the period length of rhythmicity was not significantly correlated with the 3 measures of sleep architecture (Table 2).
Sleep architecture in nestling barn owls.
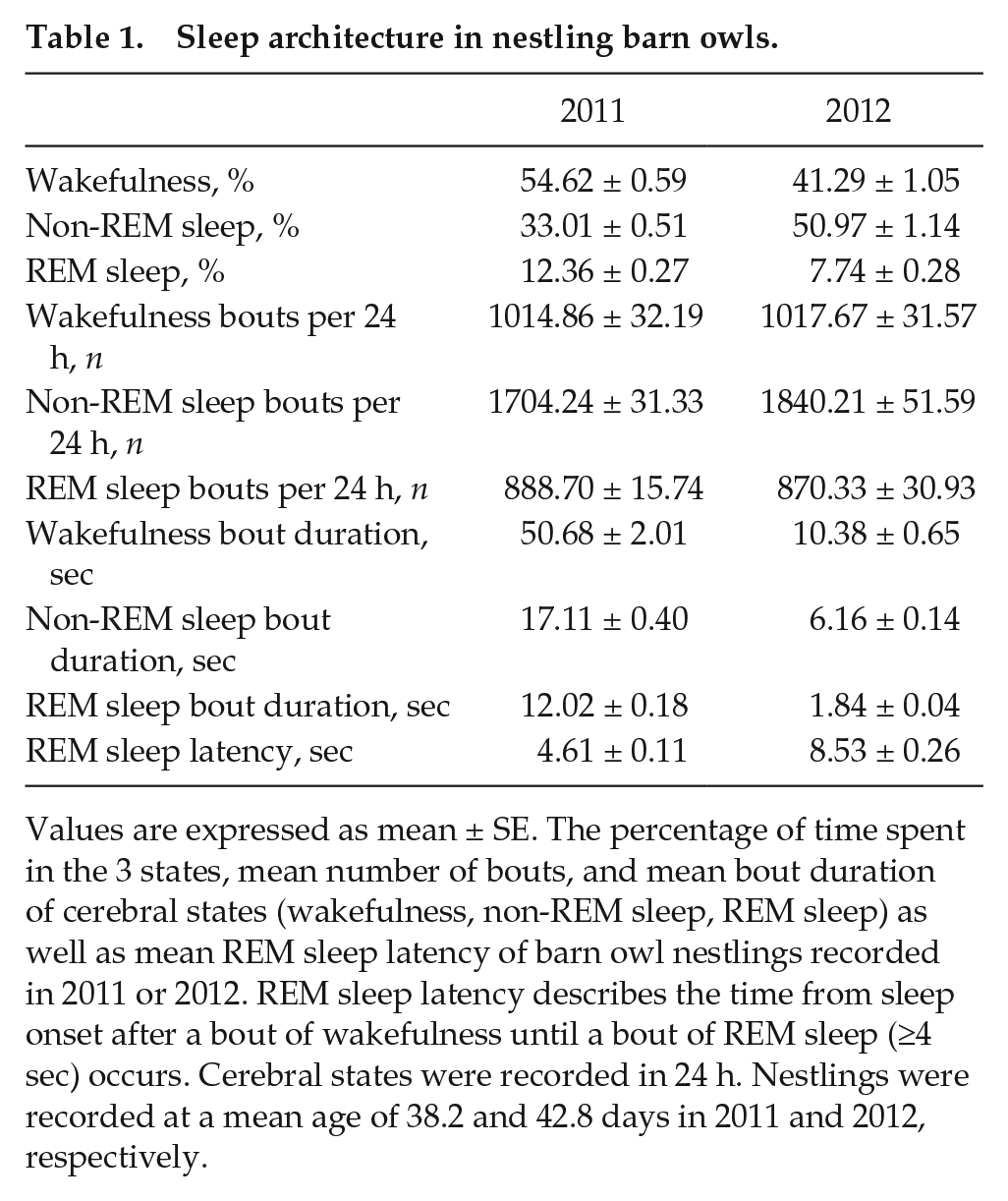
Values are expressed as mean ± SE. The percentage of time spent in the 3 states, mean number of bouts, and mean bout duration of cerebral states (wakefulness, non-REM sleep, REM sleep) as well as mean REM sleep latency of barn owl nestlings recorded in 2011 or 2012. REM sleep latency describes the time from sleep onset after a bout of wakefulness until a bout of REM sleep (≥4 sec) occurs. Cerebral states were recorded in 24 h. Nestlings were recorded at a mean age of 38.2 and 42.8 days in 2011 and 2012, respectively.
Pearson correlations between the intensity and period length of rhythmicity for the 3 cerebral states in nestling barn owls.
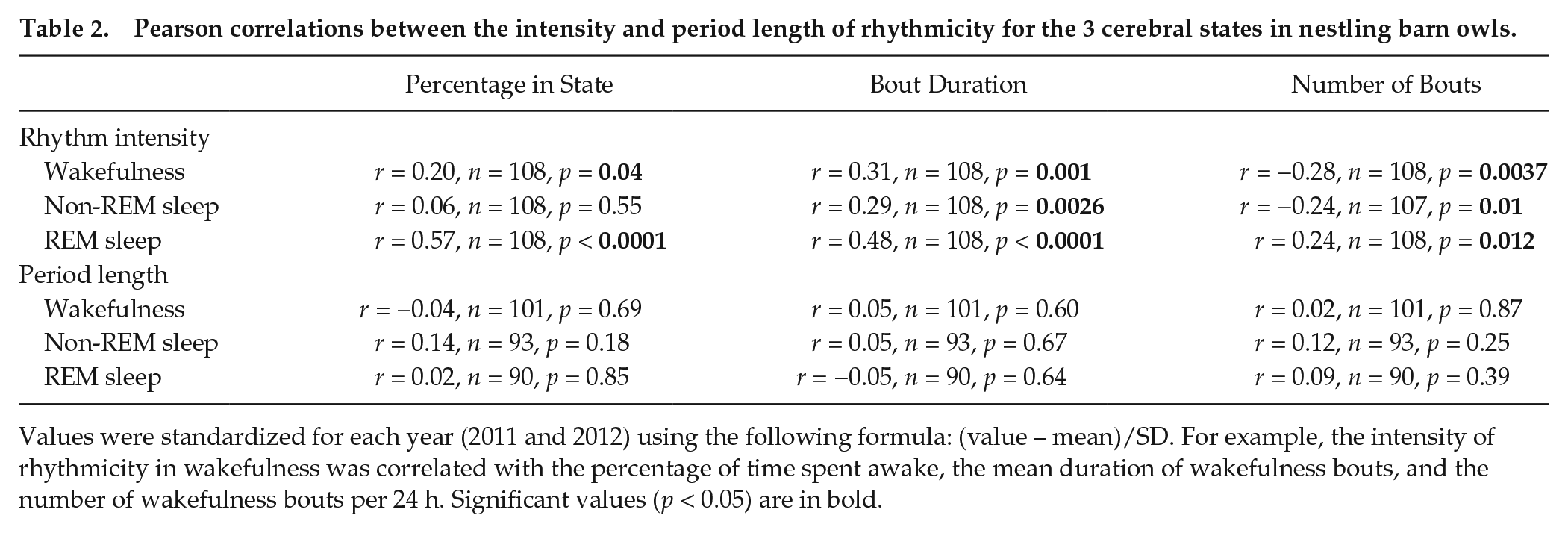
Values were standardized for each year (2011 and 2012) using the following formula: (value – mean)/SD. For example, the intensity of rhythmicity in wakefulness was correlated with the percentage of time spent awake, the mean duration of wakefulness bouts, and the number of wakefulness bouts per 24 h. Significant values (p < 0.05) are in bold.
Period Length of Rhythmic Brain Activity
Among the 66 nestlings recorded in 2011, 60 were rhythmic with respect to wakefulness, 59 to REM sleep, and 58 to non-REM sleep; overall, 44 individuals (67%) were rhythmic with respect to all 3 sleep-wakefulness components. From the 42 nestlings recorded in 2012, 41 were rhythmic in wakefulness, 39 in non-REM sleep, and 35 in REM sleep; overall, 32 nestlings (76%) were rhythmic in all 3 states. We combined the 2 years because period lengths measured in 2011 and 2012 were not significantly different (Student t test, p > 0.09). When considering only individuals showing rhythmicity, the period length of wakefulness was, on average, 4.50 ± 0.30 h (range = 1.1-12.0 h), of non-REM sleep was 4.62 ± 0.32 h (range = 1.1-12.0 h), and of REM sleep was 4.91 ± 0.31 h (range = 1.3-12.0 h) (Fig. 2). The period lengths of REM sleep and non-REM sleep were significantly associated (Pearson correlations on standardized log-transformed values, r = 0.46, n = 78 nestlings, p < 0.0001), as were REM sleep and wakefulness (r = 0.56, n = 84, p < 0.0001) and non-REM sleep and wakefulness (r = 0.91, n = 90, p < 0.0001). Nestlings raised in the same nest did not show similar period lengths for all 3 cerebral states (analysis of variance with the nest of rearing as a factor on standardized values, all p > 0.15), and similar results were obtained when considering siblings (with the nest of origin as a random variable instead of the nest of rearing).
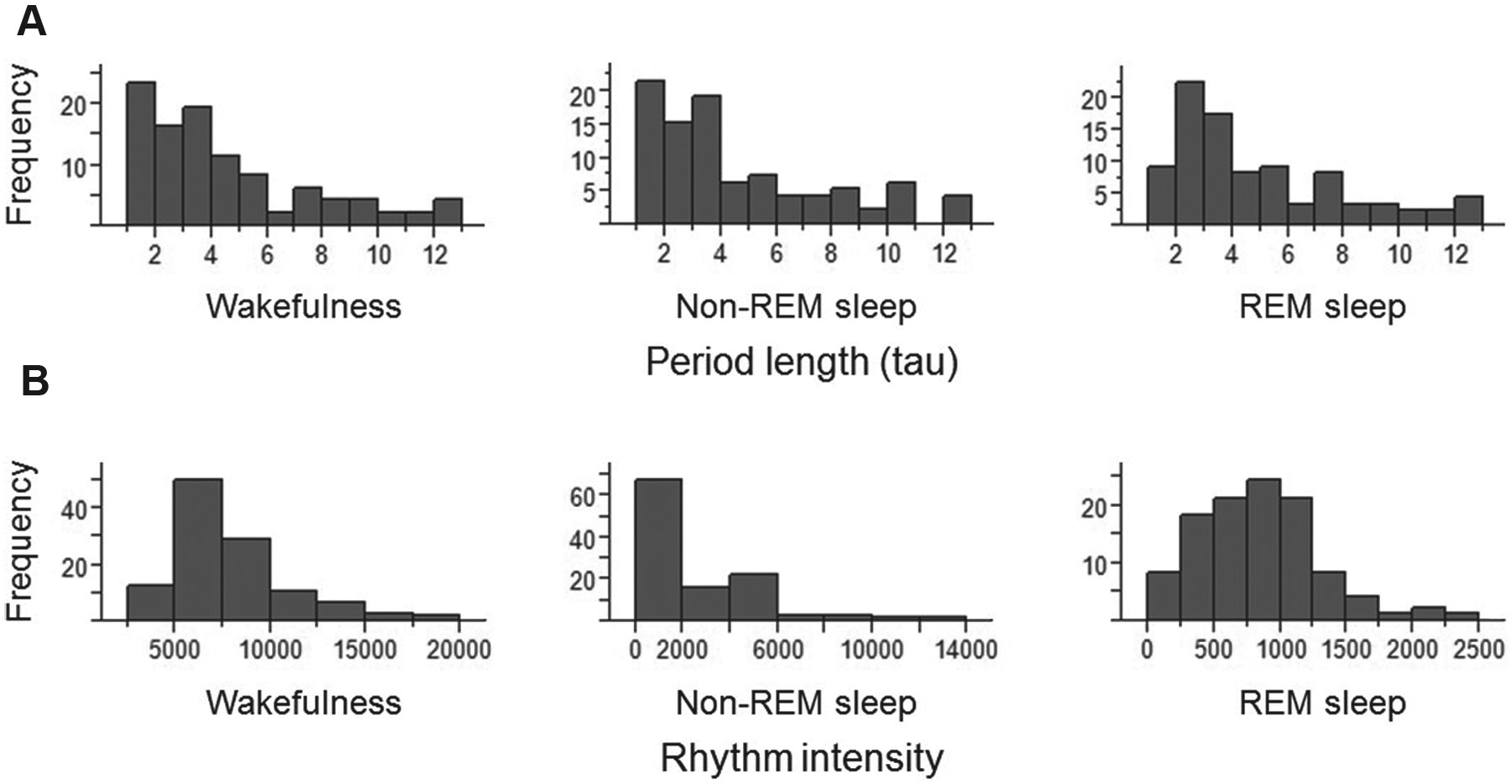
Histogram of the frequency (i.e., number of individuals) of the (A) period length and (B) intensity of rhythmicity in sleep-wakefulness states measured in barn owl nestlings.
Because the number of individuals that were rhythmic differed between cerebral states, we performed separate analyses to investigate whether the period lengths were associated with the size of black feather spots. In linear mixed models with the nest of origin as a random variable, the period length of wakefulness was significantly related to the interaction between nestling spot diameter and nestling sex (Table 3). Larger-spotted females showed a shorter period length of rhythmicity in wakefulness than smaller-spotted females (r = −0.33, n = 55, p = 0.014) (Fig. 3A), whereas in males, the opposite trend was detected (r = 0.24, n = 39, p = 0.14) (Fig. 3B). The period length of non-REM sleep was significantly related to the interaction between sex and spot diameter (Table 3). The period length of non-REM sleep was shorter in larger- than smaller-spotted females (r = −0.30, n = 49, p = 0.033) (Fig. 3D), whereas larger-spotted males showed a longer period length of non-REM sleep compared to smaller-spotted males (r = 0.40, n = 37, p = 0.015) (Fig. 3C). Finally, the period length of REM sleep was not associated with spot diameter alone or in interaction with sex (Table 3). The number of siblings was not related to the period length of all 3 sleep-wakefulness states (Pearson correlation between brood size and mean period lengths of nestlings sharing the same nest: all p > 0.28).
Linear mixed models for the relationships between period length and intensity of rhythmicity in sleep-wakefulness states and age, sex and spot diameter in barn owl nestlings.

Significant values (p < 0.05) are in bold. The nest of origin was a random variable to account for the measurement of several siblings.
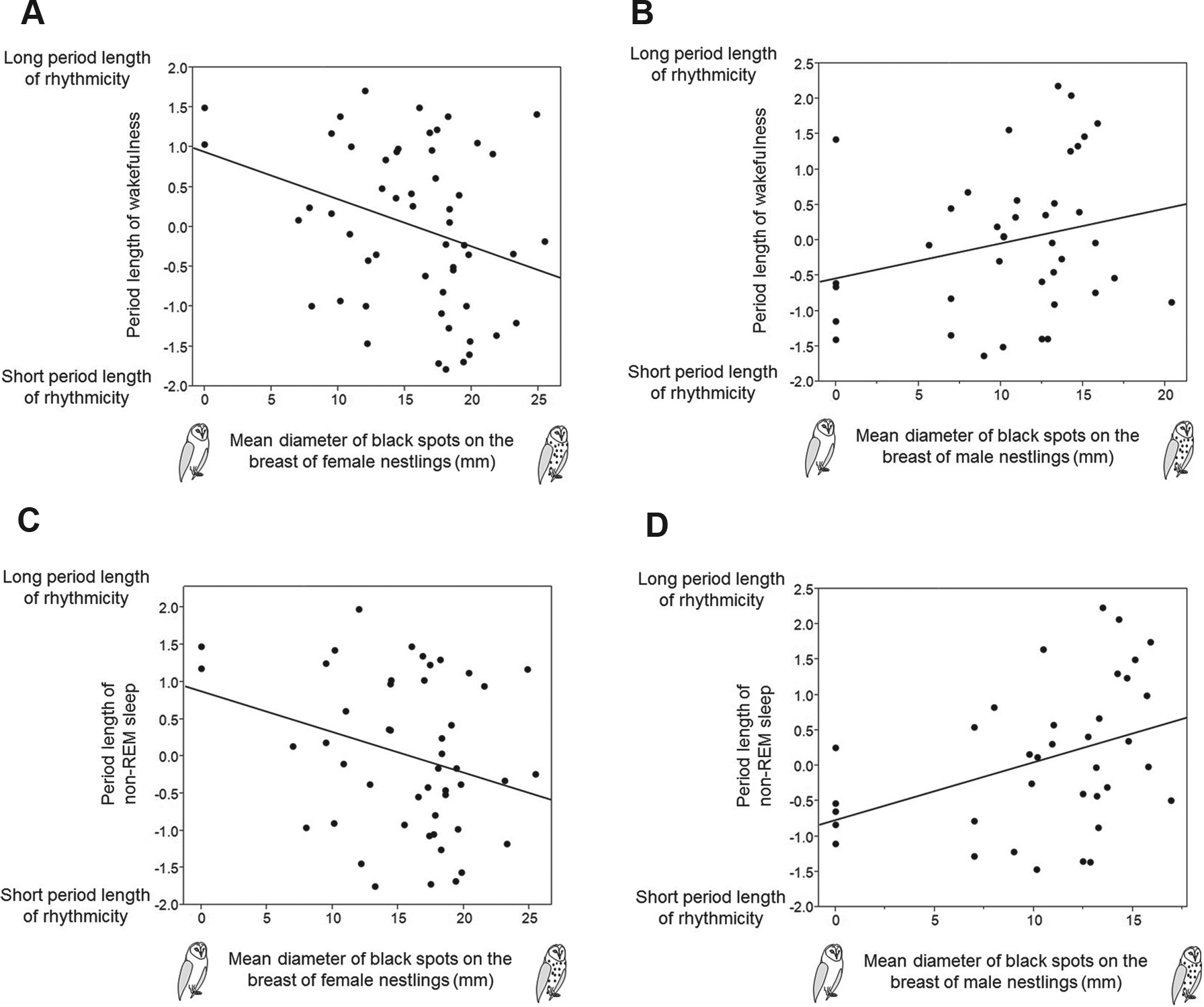
Relationship between the diameter of black feather spots and the period length of rhythmicity (log-transformed standardized tau values) of wakefulness (A, B) and non-REM sleep (C, D) in nestling male (B, D) and female (A, C) barn owls. Only rhythmic individuals were considered.
Intensity of Rhythmicity
The rhythm intensity in wakefulness did not differ between years (Student t test, t106 = −1.58, p = 0.12), whereas it was significantly higher in 2012 compared to 2011 for non-REM sleep (t106 = 35.13, p < 0.0001), and it was lower in 2012 compared to 2011 for REM sleep (t106 = −9.45, p < 0.0001). Because there were annual differences in the rhythm intensities of cerebral activities, we standardized the values for 2011 and separately for 2012. Nestlings raised in the same nest did not show similar rhythm intensities in the 3 cerebral states (analysis of covariance with the nest of rearing as a factor, p > 0.07). When replacing the nest of rearing with the nest of origin, we obtained similar results (p > 0.07). There was no link between the number of siblings and the intensity of rhythmicity for the 3 sleep-wakefulness states (p > 0.1).
The intensity of rhythmicity in wakefulness was positively correlated with the intensity of non-REM (r = 0.87, n = 108 nestlings, p < 0.0001) and REM sleep rhythms (r = 0.37, p < 0.0001), whereas the intensities of REM and non-REM sleep rhythms were not significantly correlated (r = 0.02, p = 0.84). In a linear mixed model with the nest of origin as a random factor, the intensity of rhythmicity in wakefulness was negatively related to nestling spot diameter (Table 3 and Fig. 4A). Adding to the model, the standardized percentage of time spent awake, the mean bout duration, or the number of bouts did not modify our conclusion. The intensity of rhythmicity in non-REM sleep increased with age (Fig. 5) but only tended to be associated with spot diameter (Table 3). The intensity of rhythmicity in REM sleep significantly decreased with spot diameter (Table 3 and Fig. 4B). Adding to the model, the standardized percentage of time spent asleep (non-REM sleep and REM sleep), the mean sleep bout duration, or the number of sleep bouts did not modify our conclusions.
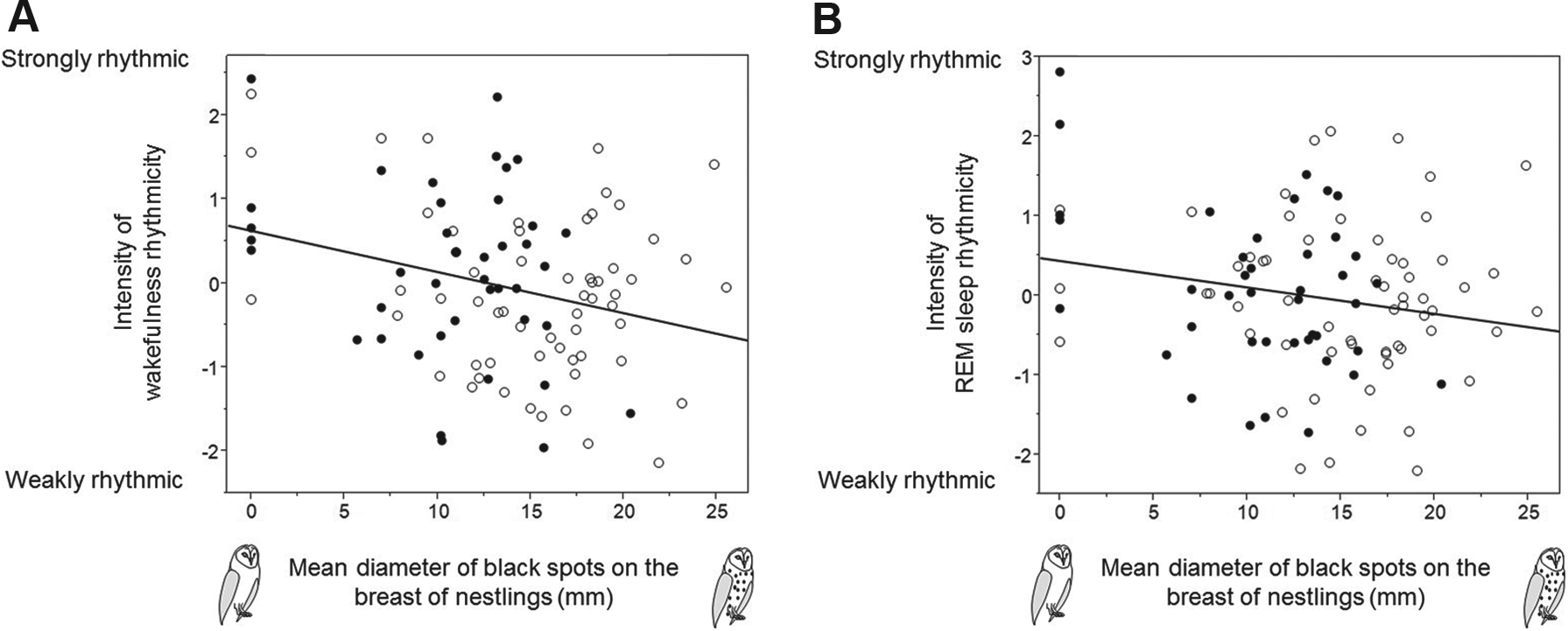
The intensity of rhythmicity in wakefulness and REM sleep in relation to spot diameter in male (closed circles) and female (open circles) barn owl nestlings. The intensity of rhythmicity in wakefulness (A) and REM sleep (B) declined with increasing spot size.
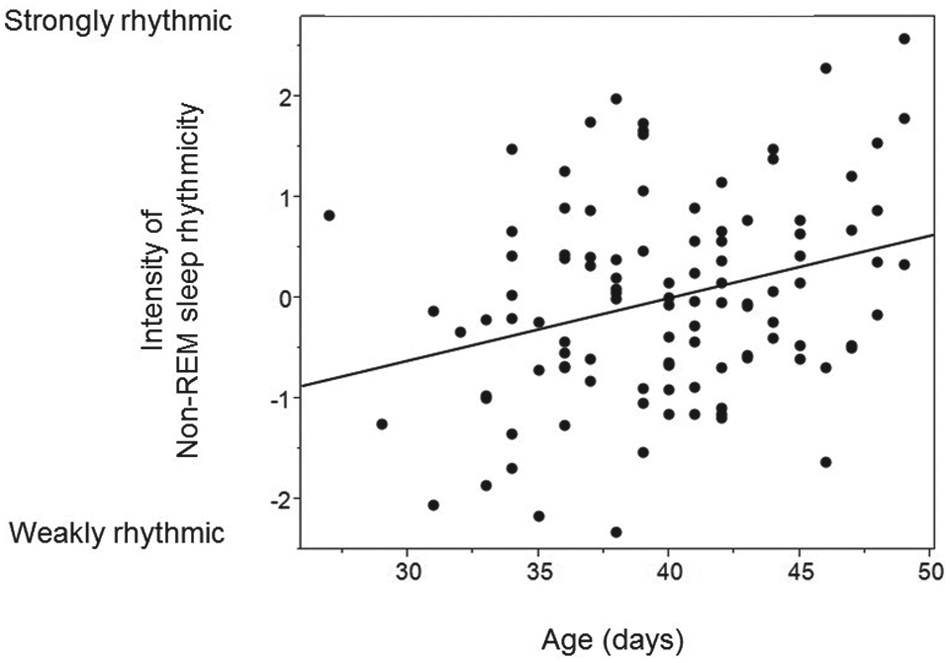
The intensity of rhythmicity in non-REM sleep increased with age in nestling barn owls.
In linear mixed models with the nest of origin and year as random variables, the size of black spots did not differ between rhythmic and arrhythmic individuals after controlling for sex (separate models for each cerebral state; wakefulness: F1,66.02 = 0.0002, p = 0.99; non-REM sleep: F1,58.94 = 0.29, p = 0.59; REM sleep: F1,66.33 = 0.58, p = 0.45).
Discussion
Barn owl nestlings were mostly rhythmic, with mean period lengths of 4.5 h for wakefulness, 4.6 h for non-REM sleep, and 4.9 h for REM sleep. Larger-spotted females had a shorter period length of wakefulness and non-REM sleep than smaller-spotted females, whereas larger-spotted males displayed a longer period length of non-REM sleep than smaller-spotted males. Regardless of sex, smaller-spotted nestlings were more strongly rhythmic with respect to wakefulness and REM sleep. The intensity of rhythmicity in non-REM sleep increased significantly with age, whereas the period length was not associated with age.
Development of Rhythmicity
In mammals and birds, sleep architecture changes during development. The time spent in REM sleep is highest in young animals and progressively decreases to reach low levels in adults (Roffwarg et al., 1966; Jouvet-Mounier et al., 1970; Frank and Heller, 1997; Thurber et al., 2008). This decrease is pronounced, and in the barn owl, we found in an earlier study a strong decline in the amount of REM sleep (from ~17% to 7%) over a relatively short period of time between 27 and 48 days of age, which was concomitant with an increase in the duration of wakefulness and non-REM sleep bouts (Scriba et al., 2013a). Here, we show that the intensity of rhythmicity, but not the period length, in non-REM sleep increased with age in barn owl nestlings. In Japanese quails (Coturnix japonica), the period length of ultradian rhythmicity in physical activity increased from day 2 to day 16 (Lumineau et al., 2001), and ultradian rhythmicity in sleep-wakefulness increased in young rats from day 17 to 12 weeks of age (Ibuka, 1984). Barn owl nestlings showed age-related increases in rhythm intensity only in non-REM sleep over their developmental period, but the period length did not change with age. Their ultradian period length was 4.5 to 4.9 h in sleep-wakefulness, which means that they showed reoccurring patterns of sleep-wakefulness every 4.5 to 4.9 h. These rhythms might occur as a result of an increase in sleep need about every 4.5 h in the nestlings. An endogenous ultradian oscillator might be responsible for the observed rhythm; further investigations are needed to test for an involvement of the SCN. Another hypothesis is that nestlings might feed about every 4 h, which could explain the period length of sleep-wakefulness states. In young rats, another altricial species, the period length of ultradian rhythm in sleep-wakefulness was 3 to 4 h at 2.5 weeks of age and increased to 8 to 12 h at the age of 4 weeks (Ibuka, 1984). Using video recordings, in adult blue tits, the period length of behavioral sleep-wakefulness at night was, on average, 2.14 h and in adult great tits was between 50 and 110 min (Mueller et al., 2012; Stuber et al., 2015). This raises the possibility that ultradian rhythms in nestlings are much shorter than in adults. Indeed, in altricial animals, young nestlings may differ considerably in their activity pattern compared to adults since they do not need to forage for food at this age. Overall, the differences in data collection (e.g., the different behaviors recorded to determine rhythmicity) and methodology prevent direct comparisons with other studies on rhythmicity. The differences in the time spent in the 3 sleep-wakefulness states between years (Table 1) can be explained not only by the mean age difference between the years (in 2012, older nestlings were recorded) but also by differences in scoring. Additionally, environmental factors could play a role.
Social Aspects of Rhythmicity
We found individual differences in the intensity of ultradian sleep-wakefulness rhythmicity, with some nestlings displaying an intense rhythm characterized by a high amplitude and other nestlings showing a less intense rhythm with a low amplitude of rhythmicity. The period length and intensity of rhythmicity in sleep-wakefulness states were different between nest mates. This indicates that brain activity is an individual-specific trait, which might be to a less degree heritable and weakly sensitive to the social environment. Furthermore, in adult blue tits (Cyanistes caeruleus), the intensity of rhythmicity in behavioral sleep was significantly repeatable (Mueller et al., 2012). It might be that rhythmicity in sleep-wakefulness is linked to personality types. Proactive and reactive strategies differ in their interaction and reaction to the environment (Koolhaas et al., 2001). Proactive individuals are more guided by internal mechanisms, whereas reactive ones explore faster, are more flexible, and interact more with the environment (Koolhaas et al., 2001). It might be that proactive individuals are more rhythmic, anticipating regular changes in the environment, whereas reactive ones might be less rhythmic and more flexible in their behavior. This hypothesis needs to be tested in future studies as well as the fitness benefits and costs of weakly and strongly rhythmic individuals.
Rhythm and Melanism
Previous studies showed that offspring born from larger-spotted mothers switch more frequently between wakefulness and sleep and are more vigilant to watch out for the arrival of a parent bringing food to the nest (Scriba et al., 2014). These results suggest that larger-spotted nestlings are better able to anticipate any change in environmental conditions than smaller-spotted individuals, raising the hypothesis that they are less rhythmic. Our results reported in the present study are consistent with this idea. Thus, what could be the benefit for smaller-spotted individuals to be more rhythmic? Being less behaviorally flexible might be advantageous when environmental conditions are stable. Another hypothesis is that larger-spotted owls are more often looking at the nest box’s entrance, and since this is the source of light in the nest, it might weaken the robustness of the rhythm. Since smaller-spotted nestlings are less attentive towards the nest box’s entrance, this may explain why they display a more robust rhythm. More data are required to discuss further the potential behavioral consequences of variations in the degree of rhythmicity.
We did not find a difference in spot size between rhythmic and arrhythmic individuals. It might be that we do not have enough power to detect an effect because from the 108 nestlings, only 7 were arrhythmic in wakefulness, 15 in non-REM sleep, and 18 in REM sleep. Also, we cannot distinguish between arrhythmic individuals and those showing several rhythms. Arrhythmic birds do not have a defined period length, and therefore, we cannot compare the period length of rhythmic and arrhythmic birds. We only compared the period length among rhythmic birds.
Barn owl nestlings with more black spots on the feathers have a lower body temperature and higher oxygen consumption compared to owlets with fewer spots (Dreiss et al., 2016). These physiological differences resulted in color-specific behavioral strategies, with heavily spotted nestlings huddling with siblings more rapidly to reduce heat loss compared to nestlings with fewer spots. Furthermore, larger-spotted individuals consumed less food under ad libitum conditions, but when food deprived, they lost less weight (Dreiss et al., 2010). These studies suggest that eumelanin-based coloration is associated with a better ability to maintain energy balance between food intake and energy expenditure (Dreiss et al., 2016; Ducrest et al., 2008; Roulin, 2009; Roulin et al., 2010). Therefore, large- and small-spotted owls may differentially handle the trade-off between rhythmicity and energy homeostasis because the maintenance of a rhythm may be energetically intense. Accordingly, in eyeless cavefish, the absence of a circadian rhythm in metabolism allows them to save energy (Moran et al., 2014). This raises the possibility that owlets with large spots might save energy by exhibiting a weak rhythm for sleep-wakefulness.
In red deer, the ultradian period length is longer in winter, while in all other seasons, when environmental conditions are less harsh, the period length of activity is shorter (Berger et al., 2002). The annual dynamics in period length has been interpreted as a nutritional strategy depending on food quality and quantity as in winter, food is scarce and of high fiber content, taking longer to digest (Berger et al., 2002). In owlets, we did not find any association with the date, but the period length was related to the size of black feather spots in interaction with sex. Smaller-spotted female nestlings showed a longer period length for wakefulness and non-REM sleep than larger-spotted females, whereas in male nestlings, the opposite relationship was observed for non-REM sleep. This rather surprising result could be interpreted in light of the fact that selection exerted on spot size is sex dependent, with selection being positive in females and negative in males (Roulin et al., 2010). Female nestlings are larger than males (Almasi and Roulin, 2015), and hence, the physiological requirements of differently spotted individuals may differ between males and females, with a possible result that individuals show different period lengths. Although the daily variation in temperature and human-made noise occurs predictably, on a smaller scale, parental visits as well as social interactions with siblings occur probably with unpredictable frequency at night. Therefore, female nestlings with large eumelanic spots might have a shorter period length to benefit from a higher investment in vigilance, which might bring the advantage of a faster reaction to parents bringing food. Finally, the different pattern of rhythmicity in differently spotted individuals might reflect differences in the maturation of neuronal tissue involved in rhythmicity.
Footnotes
Acknowledgements
This research was supported by the Swiss National Science Foundation, the Société Académique Vaudoise, and the Max Planck Society. The authors thank Paul Béziers and Céline Simon for their help in the field and laboratory. They are grateful to William J. Schwartz and to 2 anonymous reviewers for helpful comments on a previous version of the article.
Conflict of Interest Statement
The author(s) have no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Notes
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.