
Abstract
Our objectives were to investigate the period lengths (i.e., taus) of the endogenous core body temperature rhythm and melatonin rhythm in delayed sleep-wake phase disorder patients (DSWPD) and non-24-h sleep-wake rhythm disorder patients (N24SWD) compared with normally entrained individuals. Circadian rhythms were measured during an 80-h ultradian modified constant routine consisting of 80 ultrashort 1-h “days” in which participants had 20-min sleep opportunities alternating with 40 min of enforced wakefulness. We recruited a community-based sample of 26 DSWPD patients who met diagnostic criteria (17 males, 9 females; age, 21.85 ± 4.97 years) and 18 healthy controls (10 males, 8 females; age, 23.72 ± 5.10 years). Additionally, 4 full-sighted patients (3 males, 1 female; age, 25.75 ± 4.99 years) were diagnosed with N24SWD and included as a discrete study group. Ingestible core temperature capsules were used to record minute temperatures that were averaged to obtain 80 hourly data points. Salivary melatonin concentration was assessed every half-hour to determine time of dim light melatonin onset at the beginning and end of the 80-h protocol. DSWPD patients had significantly longer melatonin rhythm taus (24 h 34 min ± 17 min) than controls (24 h 22 min ± 15 min, p = 0.03, d = 0.70). These results were further supported by longer temperature rhythm taus in DSWPD patients (24 h 34 min ± 26 min) relative to controls (24 h 13 min ± 15 min, p = 0.01, d = 0.80). N24SWD patients had even longer melatonin (25 h ± 19 min) and temperature (24 h 52 min ± 17 min) taus than both DSWPD (p = 0.007, p = 0.06) and control participants (p < 0.001, p = 0.02, respectively). Between 12% and 19% of the variance in DSWPD patients’ sleep timing could be explained by longer taus. This indicates that longer taus of circadian rhythms may contribute to the DSWPD patients’ persistent tendency to delay, their frequent failure to respond to treatment, and their relapse following treatment. Additionally, other factors can contribute to misalignments in DSWPD and N24SWD disorders.
Circadian rhythms in physiological and behavioral processes are controlled by the biological body clock, which cycles at a near 24-h pace (Aschoff, 1970; Burgess and Eastman, 2008; Czeisler et al., 1999; Wever, 1979) in synchrony with the night-day cycle (Clodore et al., 1986; Duffy et al., 2001; Gradisar and Lack, 2004; Lack and Lushington, 1996). Although these endogenously generated patterns are often slightly longer than 24 h, slight deviations from the 24-h day are entrained by external light to synchronize the sleep-wake cycle, and other rhythms, to the day-night rotation of the earth (Czeisler et al., 1989; Freedman et al., 1999). However, endogenous circadian rhythms, including the sleep-wake cycle, can become desynchronized from the light-dark cycle. This desynchrony can lead to chronic sleep difficulties and daytime impairments. The problems arising from this desynchrony are collectively termed circadian rhythm sleep-wake disorders (CRSWD), of which delayed sleep-wake phase disorder (DSWPD) is the most common (American Academy of Sleep Medicine [AASM], 2014).
According to previous literature and the International Classification for Sleep Disorders–3 (ICSD-3; AASM, 2014), DSWPD is described as a 2- to 6-h delay in the circadian and major sleep period relative to “conventional” or socially desirable times (AASM, 2014; Dagan and Eisenstein, 1999; Regestein and Monk, 1995; Sack et al., 2007). Estimated to affect 1.1% of the general population (Lovato et al., 2013; Paine et al., 2014), DSWPD has a higher prevalence in subgroups of young adults and patients with insomnia, where prevalence ranges from 10% to 16% (AASM, 2014; Lovato et al., 2013; Paine et al., 2014). Non-24-h sleep-wake rhythm disorder (N24SWD) is a less common CRSWD and is readily observed in blind patients who cannot use light to synchronize their body with the external environment. Hence, typically patients’ sleep and wake times progressively delay each day as a result of their sleep-wake cycle being longer than 24 h (AASM, 2014).
A significant phase-delay of the circadian rhythm is regarded as the main etiological factor governing DSWPD. A continually delaying circadian rhythm is regarded as the main contributing factor to N24SWD (AASM, 2014; Sack et al., 2000; Sack et al., 2007; Uchiyama et al., 2000). In both conditions, attempts to adhere to more conventional sleep-wake times result in insufficient sleep that can have negative effects on patients’ physical and psychological health (AASM, 2014; Dagan and Eisenstein, 1999; Lovato et al., 2013; Regestein and Monk, 1995; Sack et al., 2000; Sack et al., 2007; Wilhelmsen-Langeland et al., 2014).
Hypothetically, appropriately timed morning light simulation should reset the delayed circadian rhythm of DSWPD to an earlier, more desirable target time. However, empirical literature to support efficacy of treatments is inconsistent, and recurrently failed responses to treatment are reported (Dagan and Eisenstein, 1999; Sack et al., 2007; Saxvig et al., 2014; Wilhelmsen-Langeland et al., 2013). As such, other mechanisms have been suggested to contribute to delays in both DSWPD and N24SWD, including a longer than normal period length or tau (i.e., a slower than normal circadian oscillator; Dagan and Eisenstein, 1999; Sack et al., 2007; Sack et al., 2000; Weitzman et al., 1981). For example, an individual may exhibit an endogenous tau that is closer to ~25 h than the conventional 24-h solar day. By taking significantly longer to complete 1 full circadian cycle, the patient’s circadian system will exert pressure to delay sleep timing by an additional hour each day. Therefore, confirming longer taus in DSWPD could provide insight into failed chronobiological treatments.
Only 2 studies, using 2 different experimental approaches, have previously investigated the notion of a longer circadian tau in DSWPD. Campbell and Murphy (2007) used the free-running protocol to investigate the taus in a single DSWPD patient. The investigators found that the patient had a circadian cycle or period length (i.e., tau) of core temperature rhythm that was almost an hour longer than their 3 healthy control sleepers. In a pilot study, we further investigated this premise using an ultradian routine (a separate sample, not embedded within the present group of participants) and found that 6 DSWPD patients also had a significantly longer temperature rhythm tau compared with 7 normally entrained sleepers (Micic et al., 2013). A longer tau has also been proposed in N24SWD in which the patient has a longer than 24-h sleep-wake cycle that has lost synchrony with the 24-h light-dark cycle (Kitamura et al., 2013; Uchiyama et al., 2000).
The purpose of the present study was to extend the previous findings with a larger sample and additional circadian measures. Contributing to the sparse knowledge of DSWPD etiology may enable more durable treatments. This is the largest sample of DSWPD taus in the literature to date and the second largest sample of N24SWD taus. Additionally, it is the first study to derive taus of melatonin circadian rhythms in DSWPD patients. In line with previous findings, it was hypothesized that individuals with DSWPD will exhibit a significantly longer tau as measured by core body temperature and melatonin compared with normally entrained controls. A third, serendipitous group of N24SWD patients were also included in the study. It was predicted that core body temperature and melatonin taus would be longer still in N24SWD compared with taus of DSWPD patients and controls.
Methods
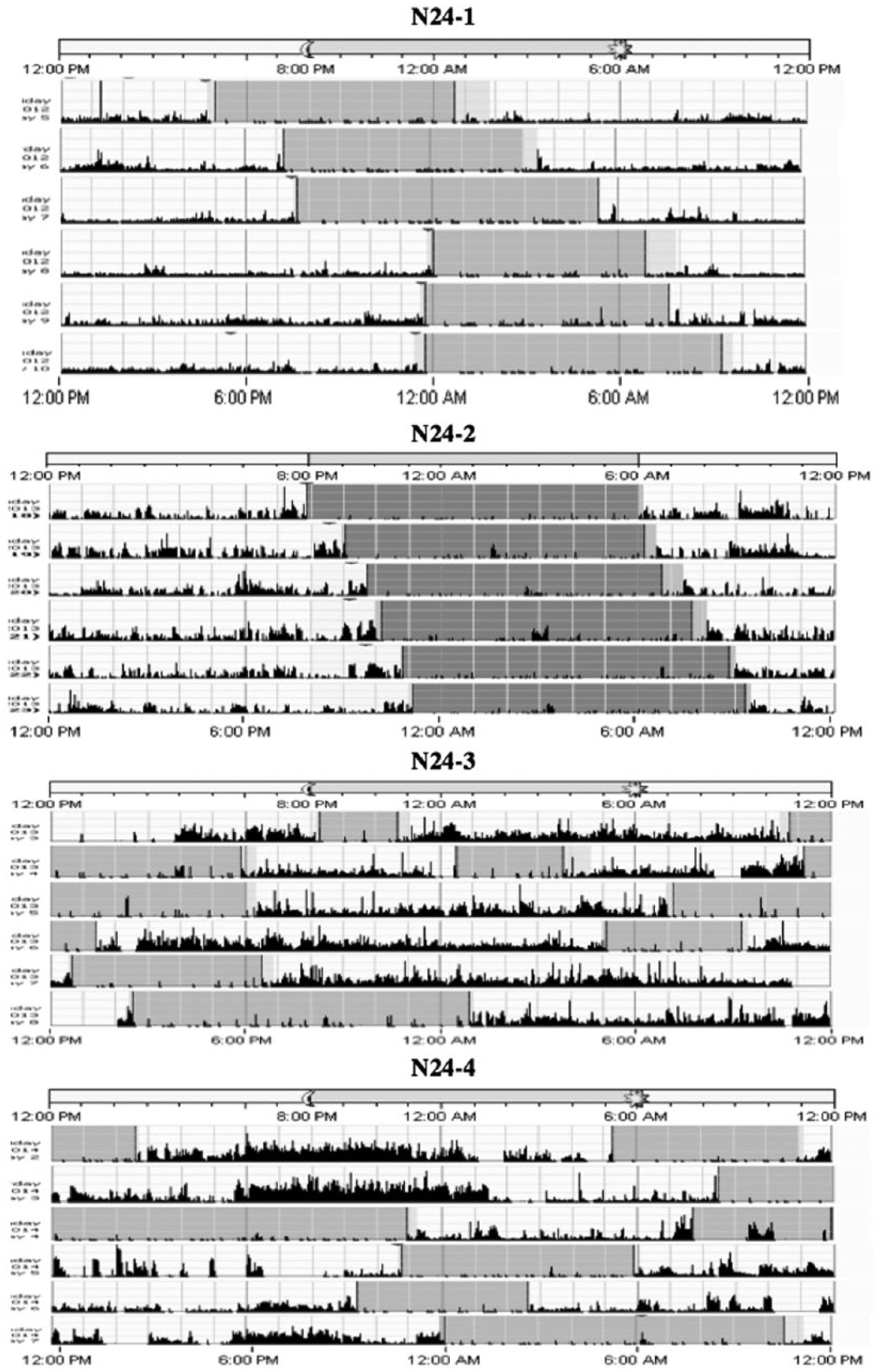
Sample Demographic
Twenty-six DSWPD patients (17 males, 9 females; age, 21.9 ± 5.0 years) were selected on the basis of ICSD-3 diagnostic criteria, as well as 18 good sleeping, normally timed controls (10 males, 8 females; age, 23.7 ± 5.1 years). An additional 4 full-sighted patients displayed instability in their delayed sleeping pattern during the screening process (see Fig. 1). They were diagnosed with N24SWD according to ICSD-3 criteria and were included as a separate study group (3 males, 1 female; age, 25.75 ± 4.99 years). The sample was community based, recruited via poster advertisements in local newspapers, television, radio, public notice boards, and educational institutions (i.e., 3 major Universities in South Australia) as well as online websites. Following a minimum 2-week screening process, suitable candidates were selected for an 80-h ultradian constant routine protocol at the Flinders University Sleep Laboratory. Informed consent was obtained prior to commencement of interviews, questionnaires, and laboratory procedures. The Southern Adelaide Clinical Human Research Ethics Committee granted ethics approval for the experiment. Upon completion, monetary compensation proportional to the time and contribution invested in the study was paid to participants. Participants who completed the entire study were reimbursed AUD$500.
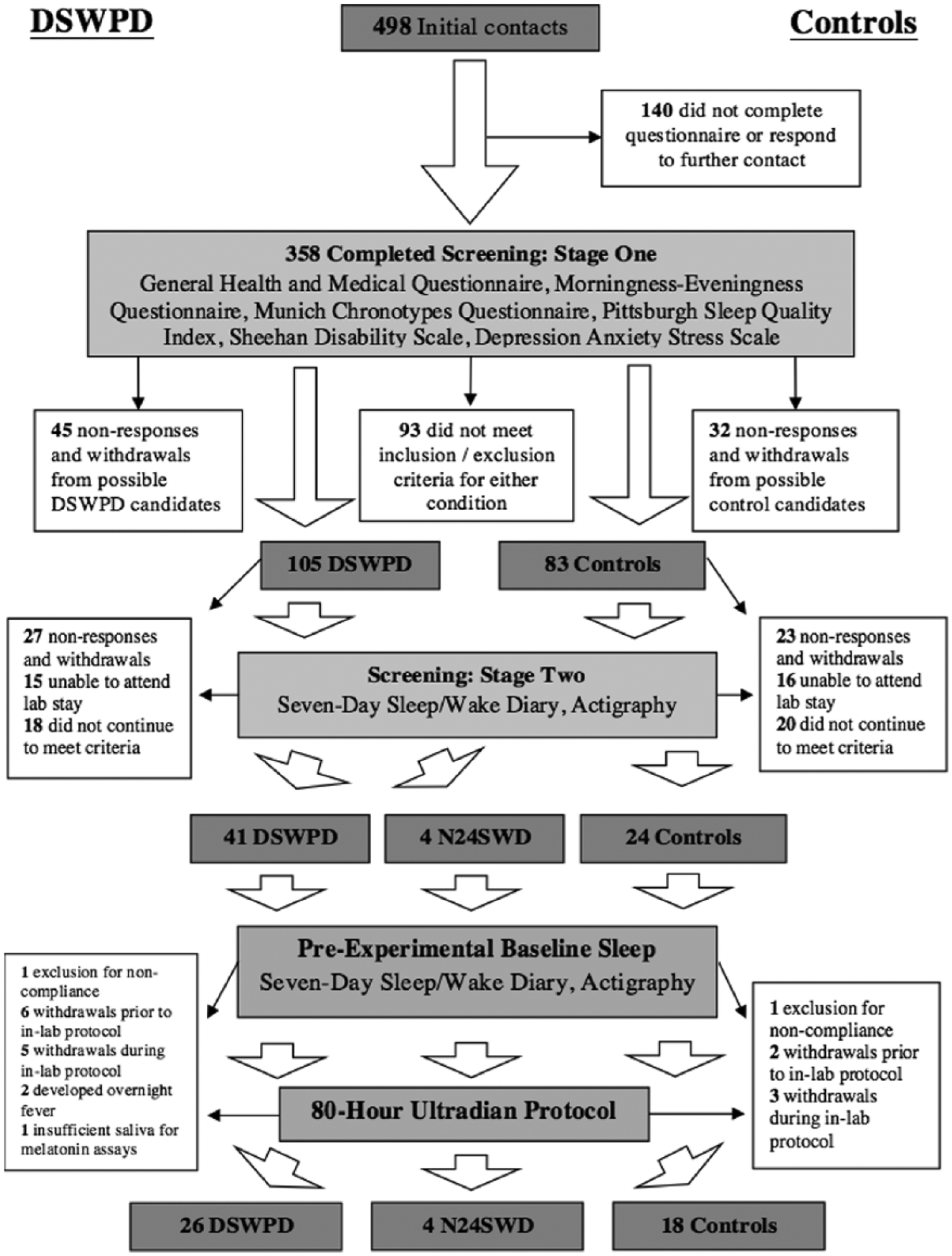
Participant flow through recruitment and involvement in the experiment before and during the 80-h ultradian modified constant routine. N24SWD patients became apparent following first week of sleep-wake diary and actigraphy monitoring.
Participant Recruitment
Screening materials included a semistructured clinical interview, a battery of psychological questionnaires, 2 weeks of sleep diaries, and wrist actigraphy. All participants were free from medical, psychiatric, and sleep disorders (except DSWPD and N24SWD). They did not take concurrent medication (including over-the-counter or herbal substances likely to affect sleep, alertness, or circadian rhythms) without approved discontinuation prior to enrolment. They were nonsmokers, were not shift-workers, were not pregnant or lactating, and reported consuming no illicit drug in the past 6 months. They had not traveled across 2 or more time zones in the 2 months prior to the study. Body mass index (BMI) was within the range of >18 and <32 kg/m2, to screen out anorexic and obese individuals with increased risk of sleep disorders. Habitual daily caffeine consumption was less than 250 mg and alcohol consumption was fewer than 14 standard drinks per week. Female participants’ menstrual cycles were noted, and those recruited either were in the follicular phase of the cycle during experimentation or used hormonal control in the form of Etonogestrel implant or the contraceptive pill.
A battery of questionnaires identified those with DSWPD and normally entrained good sleepers. The questionnaires consisted of the Munich Chronotype Questionnaire (MCTQ), Horne and Östberg’s Morningness-Eveningness Questionnaire (MEQ), Pittsburgh Sleep Quality Index (PSQI), DSWPD–Sleep Timing Questionnaire (DWSPD-STQ) (developed by the authors), Sheehan Disability Scale (SDS), and Depression Anxiety Stress Scale21 (DASS21). Patients with DSWPD were classified as evening-types (a score <42) on the MEQ. They indicated delayed sleep patterns of at least 2 h in contrast to “conventional” sleep times (i.e., sleep onset later than 0100 h) and inability to fall asleep at a preferred earlier time per the DSWPD-STQ. They further reported daytime dysfunction and lifestyle impairments as a result of their sleep pattern (i.e., a score ≥5 on any SDS scale) but otherwise had normal sleeping patterns (i.e., no other sleep disturbances) when sleeping at their usual delayed time (i.e., ~6 on the PSQI scale with no indication of comorbidities with DSWPD). Good sleepers were selected on the basis of being healthy, good sleepers at conventional sleep times (i.e., sleep onset between 2200 and 2400 h) who were neither morning nor evening types (MEQ score, 42-58); thus, they had a “typical” sleep pattern and showed no daytime impairments as a result of their sleep (i.e., <5 on any SDS scale and ≤6 PSQI).
DSWPD-STQ
This questionnaire was developed to diagnose patients who wanted to phase-advance their sleep periods by more than 2 h earlier as per the ICSD-3 criteria (AASM, 2014). Questions consisted of current sleep and wake time according to patients’ perceived body clock sleep-timing as well as their preferred sleep timing according to social and daytime obligations. To be classified as having DSWPD, patients were required to report a more than 2-h discrepancy between their current sleep time and an earlier preferred sleep schedule. For healthy, control sleepers this discrepancy had to be less than 30 min. Five-point, Likert-type scales (0 = not at all difficult; 4 = extremely difficult) also were used to assess patients’ difficulty with sleeping and waking at preferred times.
DSWPD criteria indicate the presence of recurrent difficulties falling asleep and waking up at conventional times; thus, DSWPD patients were required to report some difficulty with sleep onset and offset (score >1 at both ends) at their preferred times (AASM, 2014). This measure also indicated that control sleepers had no difficulty sleeping and awakening at necessary or conventional times.
Seven-day sleep diaries and wrist-worn activity motion monitors (AW64 Mini Mitter Actiwatch, Oregon, WA, USA) were used to further verify the patients’ sleep patterns. A qualified clinician (senior author, L.L.) confirmed participants’ eligibility for the study after reviewing all previously described screening information.
Prelaboratory Procedures
Those selected were notified and were invited to the Flinders University Sleep Laboratory for familiarization with the laboratory environment and experimental procedures. Participants signed a second consent for involvement in the 80-h laboratory session once fully informed of protocol and procedures. One week prior to the laboratory stay, participants were asked to maintain their habitual sleeping pattern, and compliance was again checked by sleep diaries and activity monitors. On the day of entry into the 80-h laboratory session, participants were instructed to refrain from napping, undertaking strenuous exercise, and consuming caffeine, alcohol, or foods thought to impede habitual melatonin secretion (e.g., chocolate, bananas, tomatoes).
Design of Protocol
The study data collection occurred over 3 years, during the Southern Hemisphere wintertime between April and August. Two or 3 participants were scheduled each week during this period and were required to enter the sleep laboratory at 1500 h Thursday for polysomnographic (PSG) and core temperature recording preparation. From 1700 h, participants remained in private, zeitgeber-free and temperature-controlled (20 °C) bedrooms, in dim-light conditions (<10 lux). The protocol comprised 80 consecutive hours of a “1-h day,” ultradian modified-constant routine protocol with alternating 20-min sleep opportunities and 40 min of enforced wakefulness (see Fig. 2). These “days” were spread evenly throughout the protocol to minimize accumulation of homeostatic sleep drive across the 80-h routine. Although the PSG data were recorded to measure the amount of sleep obtained within each sleep opportunity, it was not a primary outcome measure for the main aims of this study. After an hour of habituation, the protocol formally commenced at 1800 h with a 20-min sleep opportunity during which lighting was reduced to <1 lux. Participants were awoken after the interval, lights were turned on to less than 10 lux (~8 lux measured at angle of gaze), and enforced wakefulness was maintained for the remaining 40 min of the hour. This pattern continued until the protocol ceased at 0200 h on the subsequent Monday morning (i.e., for 80 h total).
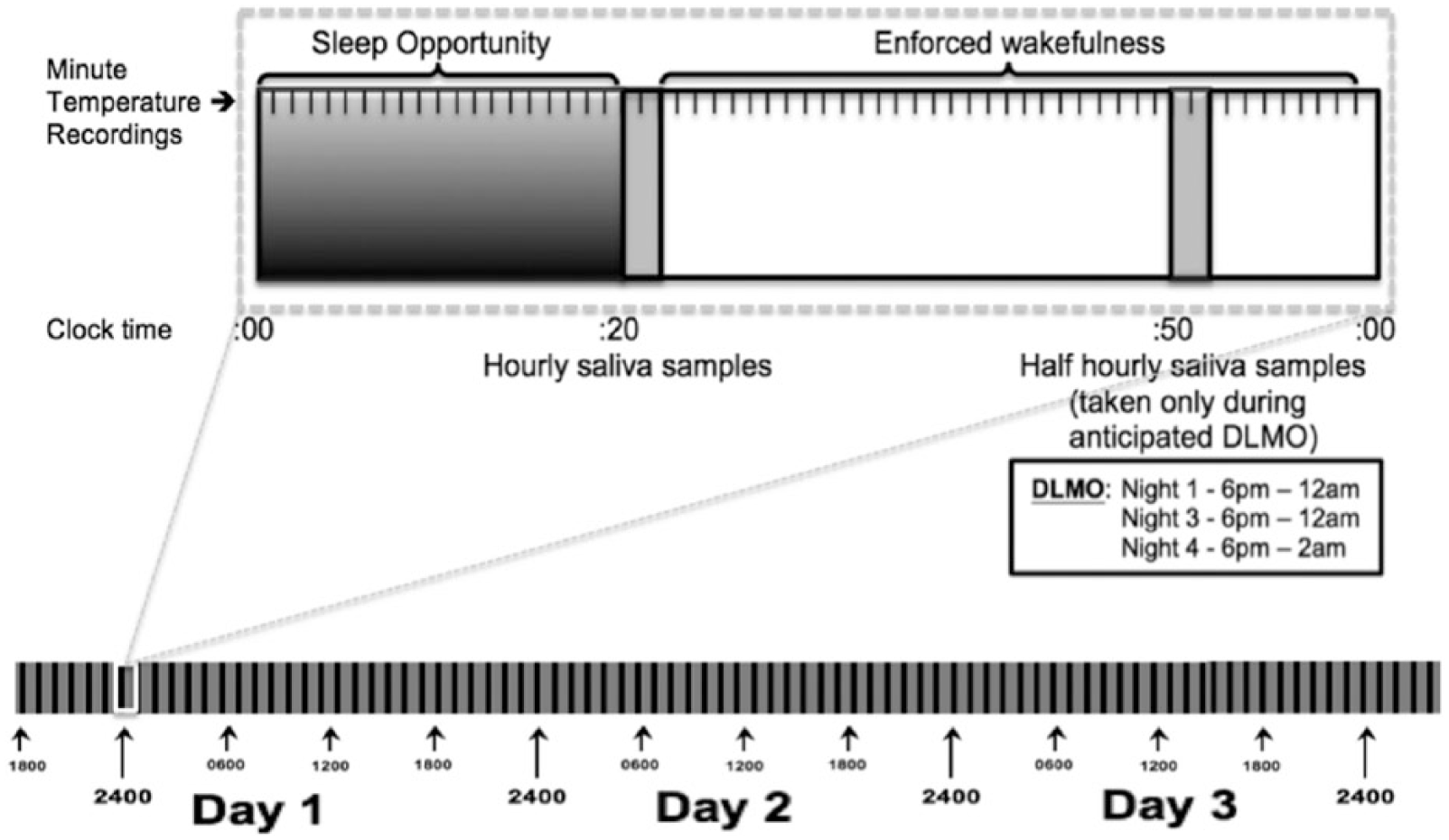
Schematic representation of the “1-h day” modified-constant routine with 20- to 40-min alternating ultradian cycles.
During the enforced wakeful periods, participants could read, watch films, study, listen to music, or talk with research assistants while remaining in bed at a near-supine body position (<45° angle). Care was taken to shield participants from any knowledge of time of day by removing clocks or any electronic device with time information and by training research assistants to avoid revealing time of day directly or inadvertently through their behavior or disposition. Participants were permitted to leave the bed only for toilet breaks and basic hygiene (i.e., brushing teeth but without toothpaste), during which light intensity was maintained at less than 10 lux. They were encouraged to use the commencement of enforced wakefulness periods immediately after the hourly salivary collections for these occasional activities. Participants were offered calorie-controlled snacks (820 kJ; 200 kcal) every 2 h to minimize the thermogenic effects of energy intake. Snacks and accompanying 200 mL of water were served at room temperature to minimize effects on core temperature. The snacks and water were served after awakening and following salivary sampling to avoid contamination of melatonin samples.
Assessment of Circadian Rhythms
Salivary melatonin
Salivary melatonin samples with a sampling quantity of more than 2 mL were collected during the 80-h period using salivettes (Cat. No. 51.1534; Sarstedt Australia Pty. Ltd., Mawson Lakes, South Australia). Hourly samples were taken immediately upon cessation of every sleep opportunity. Half-hourly samples were also taken around the time of expected dim light melatonin onset (DLMO) to obtain a finer grained measure of DLMO. These occurred exactly half an hour following the previous sample on evenings 1 (i.e., Thursday 1850-2350 h), 3 (i.e., Saturday 1850-2350 h), and 4 (i.e., Sunday 1850 h to Monday 0150 h). Participants placed the swab in their mouth and accumulated saliva for 2 min while in a supine position in bed under dim light, to minimize the masking effects of physical movement and light on endogenous melatonin production.
Upon collection, saliva samples were labelled and stored frozen at −20 °C and were later radioimmunoassayed for melatonin concentration (Adelaide Research Assay Facility, Robinson Research Institute, School of Paediatrics and Reproductive Health, University of Adelaide). Samples capturing individuals’ full profiles of melatonin on night 1 and DLMOs on evening 4 were estimated from temperature data and sent for radioimmunoassay analysis. Those whose DLMO was not captured on evening 4 because of late phase timing had samples from night 3 also analyzed. The DLMO on night 3 was used in these participants to calculate melatonin tau. This subsample comprised 7 participants, or 27% of the DSWPD group.
For melatonin analysis, salivettes were thawed and centrifuged for 10 min at 2500 rpm, and the swabs were removed from the casing, retaining the supernatant. To measure melatonin in the saliva, the minimum detectable dose of melatonin (i.e., analytical sensitivity) was 4.3 pM direct radioimmunoassay (RIA), using reagents from Buhlmann Laboratories (AG, Allschwil, Switzerland) (Voultsios et al., 1997). The low % was 9.2, and the high QC %CV was 15.3.
Core body temperature
Participants were instructed to swallow an ingestible telemetric temperature sensor, the Jonah core temperature capsule, that transmitted to a VitalSense wireless physiological telemetric monitor (Philips Respironics, Mini Mitter Co., Inc., Bend, OR, USA). This occurred immediately upon entry to the sleep laboratory (1500-1600 h) to maximize stabilization of core temperature measurement from the start of the protocol at 1800 h. Temperature was recorded continuously for 80 h in 60-sec epochs. The VitalSense consists of a data logger with a resolution of 0.01 °C, which receives radio signals every 15 sec from the temperature-sensitive ingestible capsule. VitalSense is a valid index of core temperature, and between temperatures of 32 and 42 °C, the Jonah capsules show an accuracy deviation of ±0.1 °C from the actual body temperature (Byrne and Lim, 2007; McKenzie and Osgood, 2004).
Participants were instructed to maintain the VitalSense device within 0.5 m of their body since the capsules cannot transmit data at a distance more than 1 m. The capsules pass through the gastrointestinal tract, with variable times of 12 to 48 h depending on individual gastrointestinal motility. The absence of data transmission on the VitalSense was used as an indication of an excreted capsule from the gastrointestinal tract, and the capsule was replaced by ingesting a new capsule. During the 80-h laboratory session, 1 to 3 capsules were typically ingested per participant.
Data Analysis and Tau Calculation
The phase of the melatonin rhythm was calculated from the time of DLMO by linear interpolation of the melatonin concentrations. Melatonin concentrations from the protocol capturing the first and final DLMO evenings were plotted on KaleidaGraph version 4.1.3
Recorded core temperature data from the VitalSense were downloaded and saved via the VitalSense software to a Microsoft Excel 2007 software program at the end of each protocol. To obtain mean hourly core temperature values, the transmitted temperatures were summed and divided by the number of recorded values per hour. During the first 5 h of capsule transit, temperature-decreasing artifacts sometimes occurred following snacks and water (Byrne and Lim, 2007). This was occasionally evident in 29% of the sample (control n = 6; DSWPD n = 7; N24SWD n = 1); in such cases, these data were excluded from the hourly mean calculations. The total data loss across the 80 h for all participants equated to 5%. Hourly mean temperature values were analyzed by a KaleidaGraph (Synergy Software, Reading, PA, USA) least squares curve-fitting option. The curve-fitting option consisted by a cosine function comprising the fundamental near-24-h oscillation and its first near-12-h harmonic (Brown and Czeisler, 1992; Duffy et al., 2001). The mathematical formula enables the curve to stretch or contract somewhat around a 24-h period until the greatest variance explained was reached and the temperature tau (Tτ) value was generated. The program was not constrained within particular ranges in the search for periods (i.e., taus) since temperature data showed robust, clear trends. Curve fit solutions always had high face validity with variance explained by the fitted curves ranging from r = 0.79 to 0.98 with a mean value of r = 0.90. From these best-fit curves, individual time of core temperature minimum (Tmin) and the period length (Tτ) were determined.
Statistical Analysis
DLMO, Mτ, Tmin, Tτ, and demographic variables were entered and analyzed using SPSS software version 22 (SPSS, Chicago, IL, USA) for Macintosh. Independent-samples t-tests with 2-tailed probability were used to compare group means and distributions on demographic variables and circadian phase positions. Cohen’s d was also used to calculate effect sizes between groups. For inferential statistics, a repeated-measures analysis of variance (ANOVA) was used to test the interaction and main effects of group on taus, and effect sizes were measured using partial eta square (η2). An analysis of covariance (ANCOVA) was used to test tau differences between groups by controlling for gender. Simultaneous multiple regressions and standardized beta values were further used to assess sleep delay variances, as explained by circadian parameters (see supplementary data for sample circadian curves).
Results
Baseline Assessment
Sleep and lifestyle characteristics between DSWPD patients and control sleepers are shown in Tables 1 and 2, respectively. These data are not presented for N24SWD patients, whose sleep characteristics were free-running and could not be captured by central tendency measures. Their sleep characteristics are thus illustrated in Figure 3. DSWPD patients had symptoms consistent with the ICSD-3 diagnosis of DSWPD, as shown in Table 1. Table 2 indicates that DSWPD and N24SWD patients had significant disturbance to their lifestyle compared with controls. DSWPD presented with evening-type scores (i.e., MEQ and MCTQ) compared with the intermediate-type controls. No significant differences were found between controls and DSWPD patients on lifestyle factors such as age, BMI, and caffeine and alcohol intake. The gender ratio is representative of the clinical population, with higher prevalence of DSWPD estimated in males than females (Paine et al., 2014). All 3 groups obtained similar amounts of sleep during the 80-h laboratory protocol; thus, this factor would not have affected the period estimates. Throughout the 80-h protocol, DSWPD patients obtained 722 ± 123 min of sleep, N24SWD patients 776 ± 229 min, and controls 744 ± 111 min [F(2,45) = 0.384, p = 0.68].
Summary of delayed sleep-wake phase disorder diagnosis in patients based on ICSD-3 criteria.
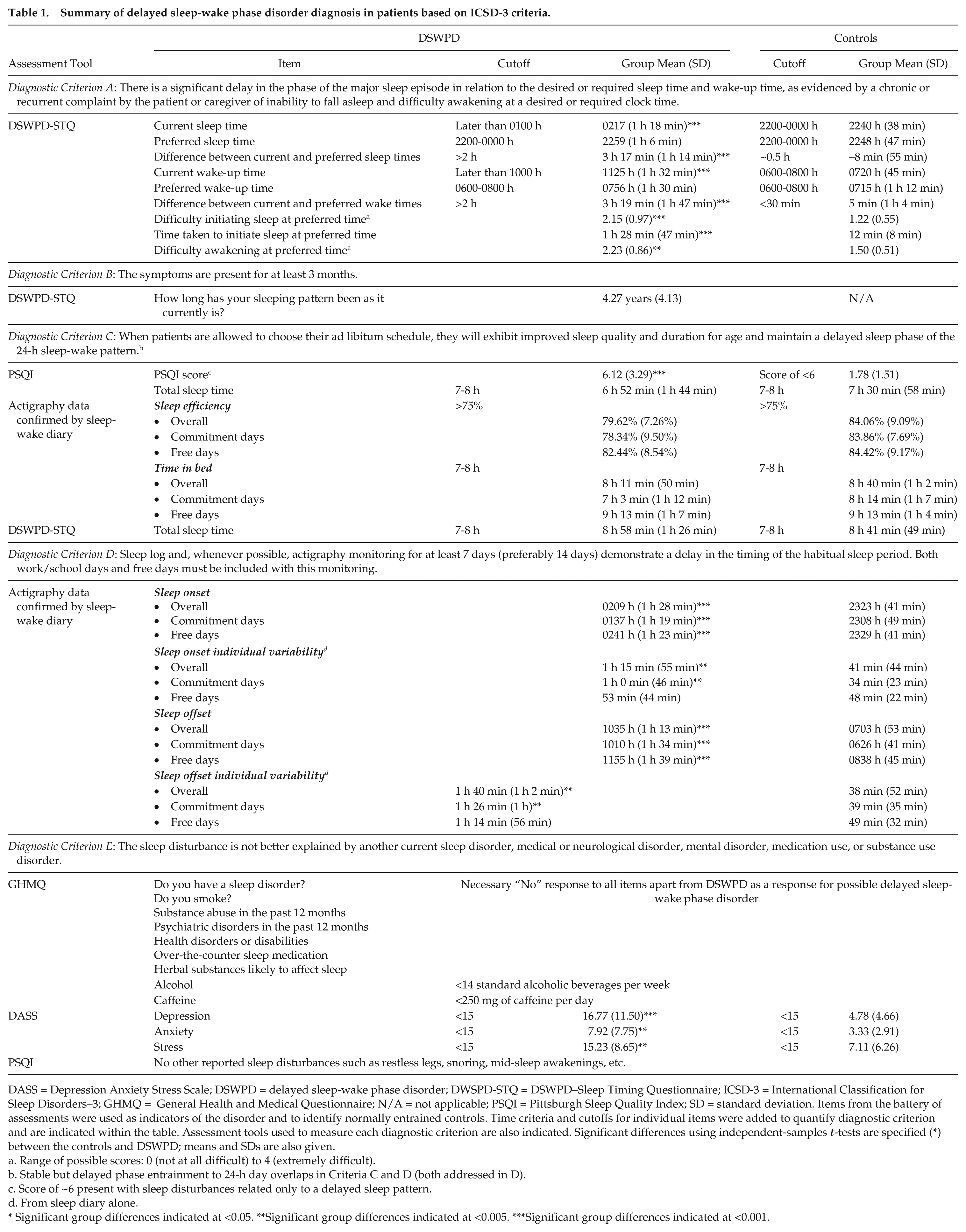
DASS = Depression Anxiety Stress Scale; DSWPD = delayed sleep-wake phase disorder; DWSPD-STQ = DSWPD–Sleep Timing Questionnaire; ICSD-3 = International Classification for Sleep Disorders–3; GHMQ = General Health and Medical Questionnaire; N/A = not applicable; PSQI = Pittsburgh Sleep Quality Index; SD = standard deviation. Items from the battery of assessments were used as indicators of the disorder and to identify normally entrained controls. Time criteria and cutoffs for individual items were added to quantify diagnostic criterion and are indicated within the table. Assessment tools used to measure each diagnostic criterion are also indicated. Significant differences using independent-samples t-tests are specified (*) between the controls and DSWPD; means and SDs are also given.
Range of possible scores: 0 (not at all difficult) to 4 (extremely difficult).
Stable but delayed phase entrainment to 24-h day overlaps in Criteria C and D (both addressed in D).
Score of ~6 present with sleep disturbances related only to a delayed sleep pattern.
From sleep diary alone.
Significant group differences indicated at <0.05. **Significant group differences indicated at <0.005. ***Significant group differences indicated at <0.001.
Group differences (mean and SD) in lifestyle factors, current estimated sleep timing, sleep quality, and associated impairments derived from prescreening measures.
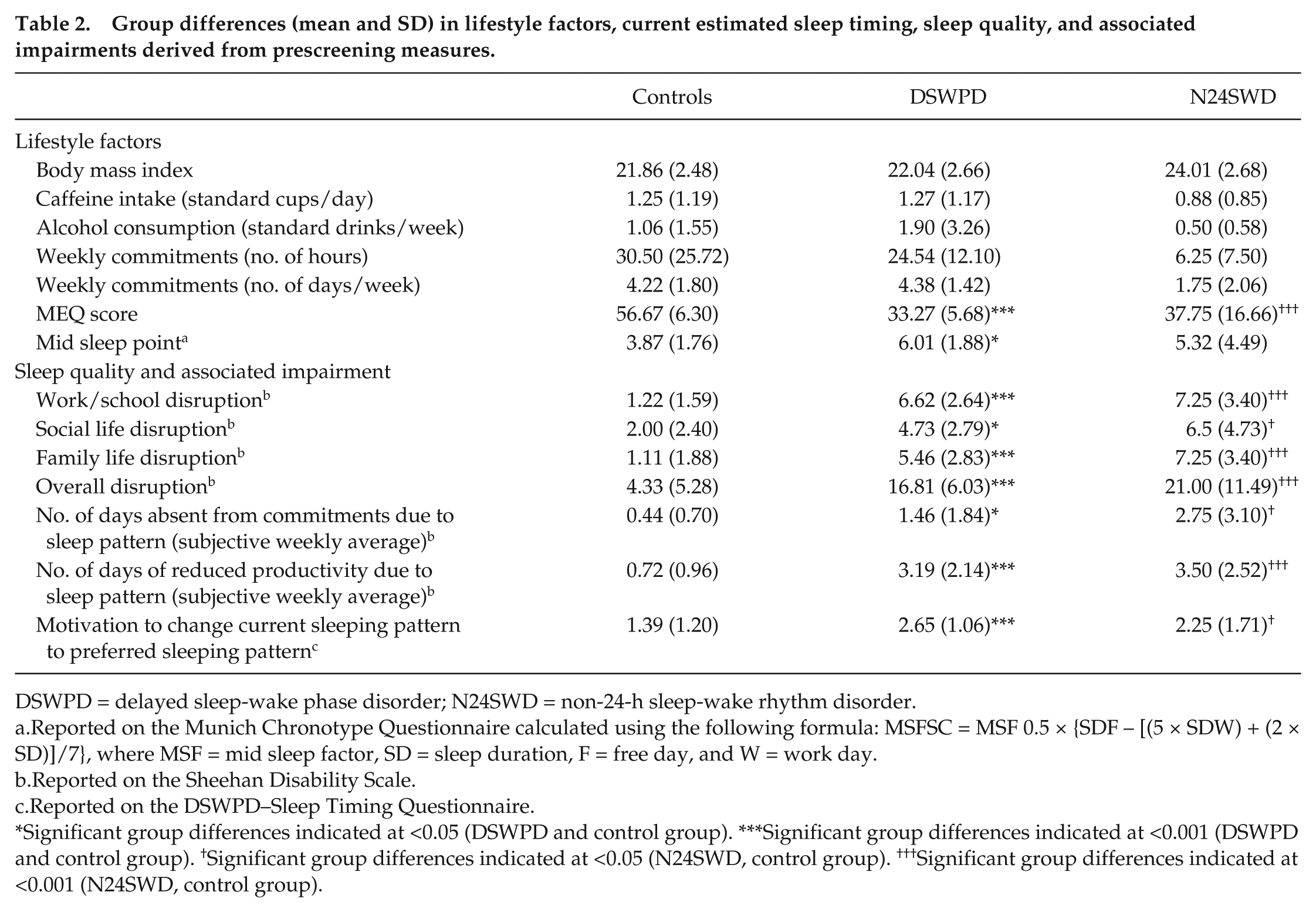
DSWPD = delayed sleep-wake phase disorder; N24SWD = non-24-h sleep-wake rhythm disorder.
Reported on the Munich Chronotype Questionnaire calculated using the following formula: MSFSC = MSF 0.5 × {SDF – [(5 × SDW) + (2 × SD)]/7}, where MSF = mid sleep factor, SD = sleep duration, F = free day, and W = work day.
Reported on the Sheehan Disability Scale.
Reported on the DSWPD–Sleep Timing Questionnaire.
Significant group differences indicated at <0.05 (DSWPD and control group). ***Significant group differences indicated at <0.001 (DSWPD and control group). †Significant group differences indicated at <0.05 (N24SWD, control group). †††Significant group differences indicated at <0.001 (N24SWD, control group).
Actigraphy of 4 N24SWD patients prior to commencement of the 80-h experimental laboratory procedure.
Circadian Phase Assessment
The timing of DLMO and Tmin on the first night of the ultradian routine was compared between controls and DSWPD using independent-samples t-tests. These results confirmed significantly later timed phases of both markers, with DLMO occurring at 2312 h ± 2 h in DSWPD patients and at 2056 h ± 1 h 26 min in controls [t(42) = 4.04, p < 0.001, d = 1.27]. Likewise, Tmin occurred later in DSWPD patients (0711 h ± 2 h 10 min) than controls (0420 h ± 1 h 29 min), with a significant mean difference of 2 h 51 min [t(42) = 5.19, p < 0.001, d = 1.49]. Furthermore, these initial phase markers on the first night were highly correlated (r = 0.80, p < 0.001) with participants having later salivary melatonin onsets and also having later Tmin, with an average of 7 h 39 min ± 1 h 26 min later (see Fig. 4). Similar associations between phase markers were found when groups were analyzed separately. In DSWPD, DLMO preceded Tmin by 7 h 54 min ± 1 h 32 min (r = 0.73, p < 0.001), and in controls DLMO preceded Tmin by 7 h 17 min ± 1 h 13 min (r = 0.64, p = 0.006; see Fig. 4).
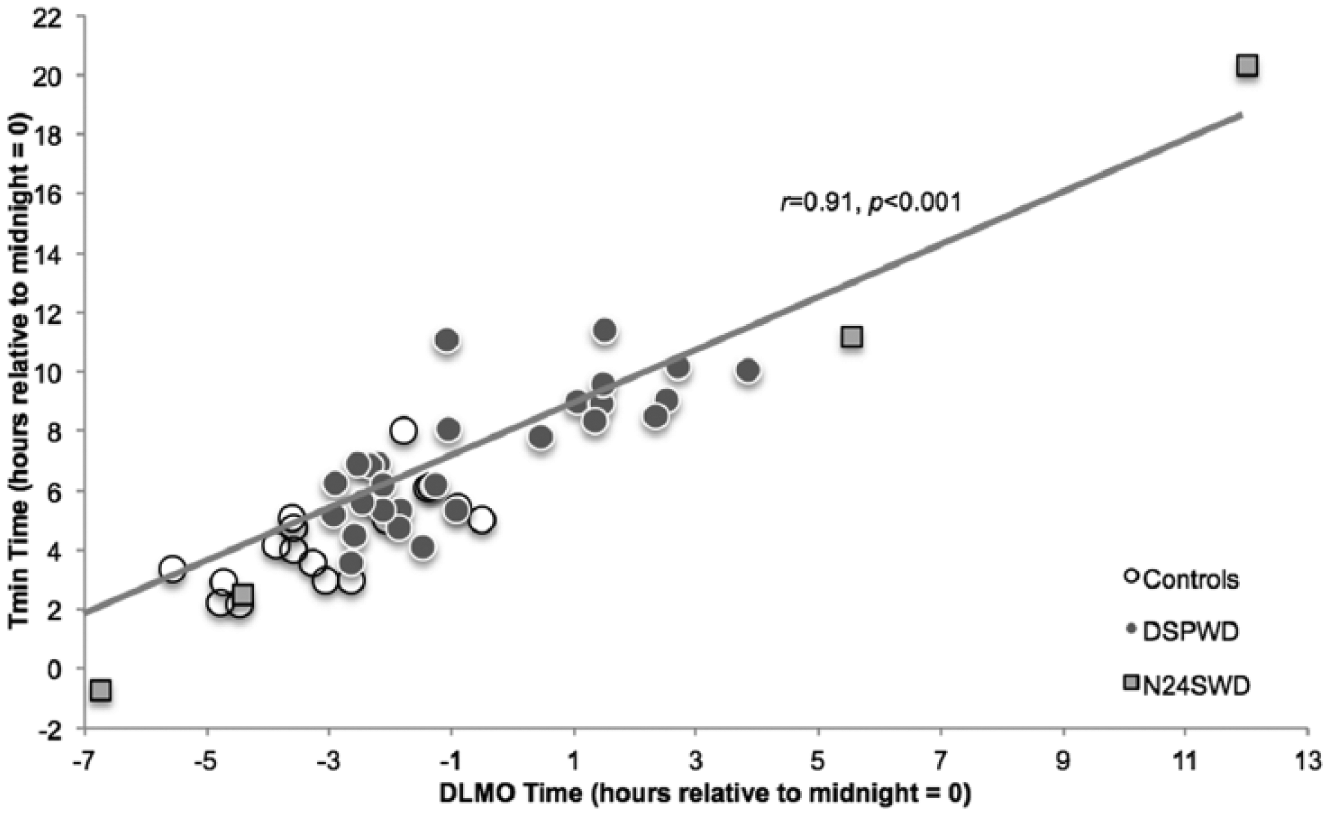
The timing of dim light melatonin onsets (DLMO) versus core temperature minimum (Tmin) of DSWPD patients, N24SWD patients, and controls. Data were sampled from the first evening of the 80-h constant routine. The timing of phase markers showed a strong, positive correlation with (r = 0.91, p < 0.001) and without (r = 0.80, p < 0.001) the data of N24SWD patients. Phase marker associations between time of Tmin and DLMO for DSWPD patients and controls combined are represented with the line of best fit.
This pattern was also consistent in N24SWD patients, who displayed highly correlated phase markers (r = 0.99, p = 0.006); however, this is largely due to their very disparate circadian phases on day 1 since they span the entire 24-h period. Although the interval between DLMO and Tmin was somewhat shorter in N24SWD patients (6 h 27 min ± 59 min) compared with DSWPD patients and controls, these differences in phase marker timing were not statistically significant between the 3 groups.
Circadian Tau Assessment
Table 3 presents individual tau estimates and Figure 5 presents the mean taus of controls, DSWPD patients, and N24SWD patients. A 2 (tau marker: temperature, melatonin) × 3 (group: DSWPD, N24SWD, control) repeated-measures ANOVA revealed no overall interaction or main effect of tau marker but a significant main effect of group [F(45,2) = 10.25, p < 0.001, η2 = 0.32]. Post hoc tests with a Bonferroni adjustment further revealed overall significant mean differences between taus of DSWPD patients and controls (mean difference [MD] = 15 min, standard error [SE] = 5 min, p = 0.014), between taus of N24SWD patients and DSWPD patients (MD = 23 min, SE = 9 min, p = 0.038), and between taus of N24SWD patients and controls (MD = 38 min, SE = 9 min, p < 0.001).
Core body temperature taus (Tτ) and melatonin taus (Mτ) of each participant.
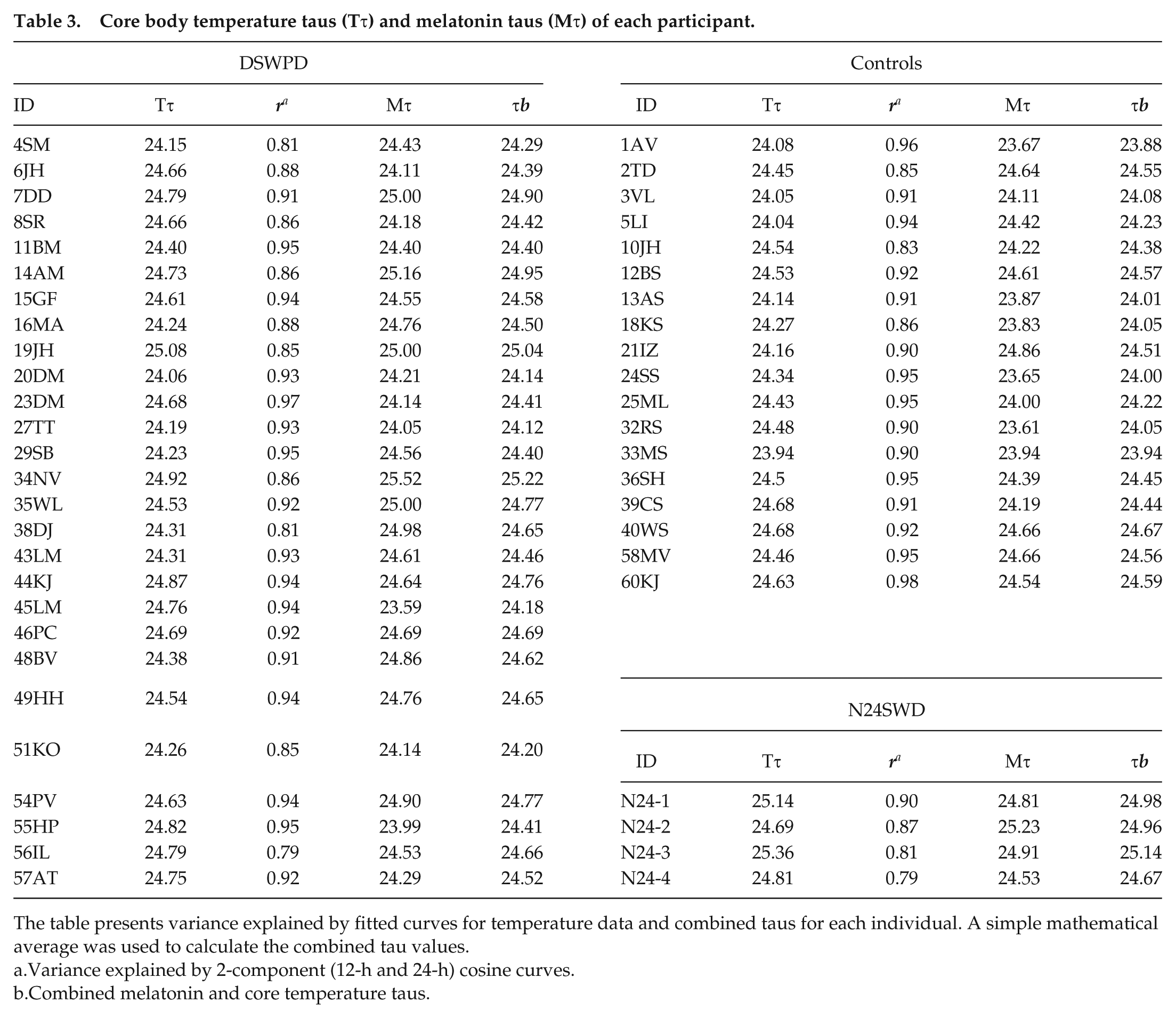
The table presents variance explained by fitted curves for temperature data and combined taus for each individual. A simple mathematical average was used to calculate the combined tau values.
Variance explained by 2-component (12-h and 24-h) cosine curves.
Combined melatonin and core temperature taus.
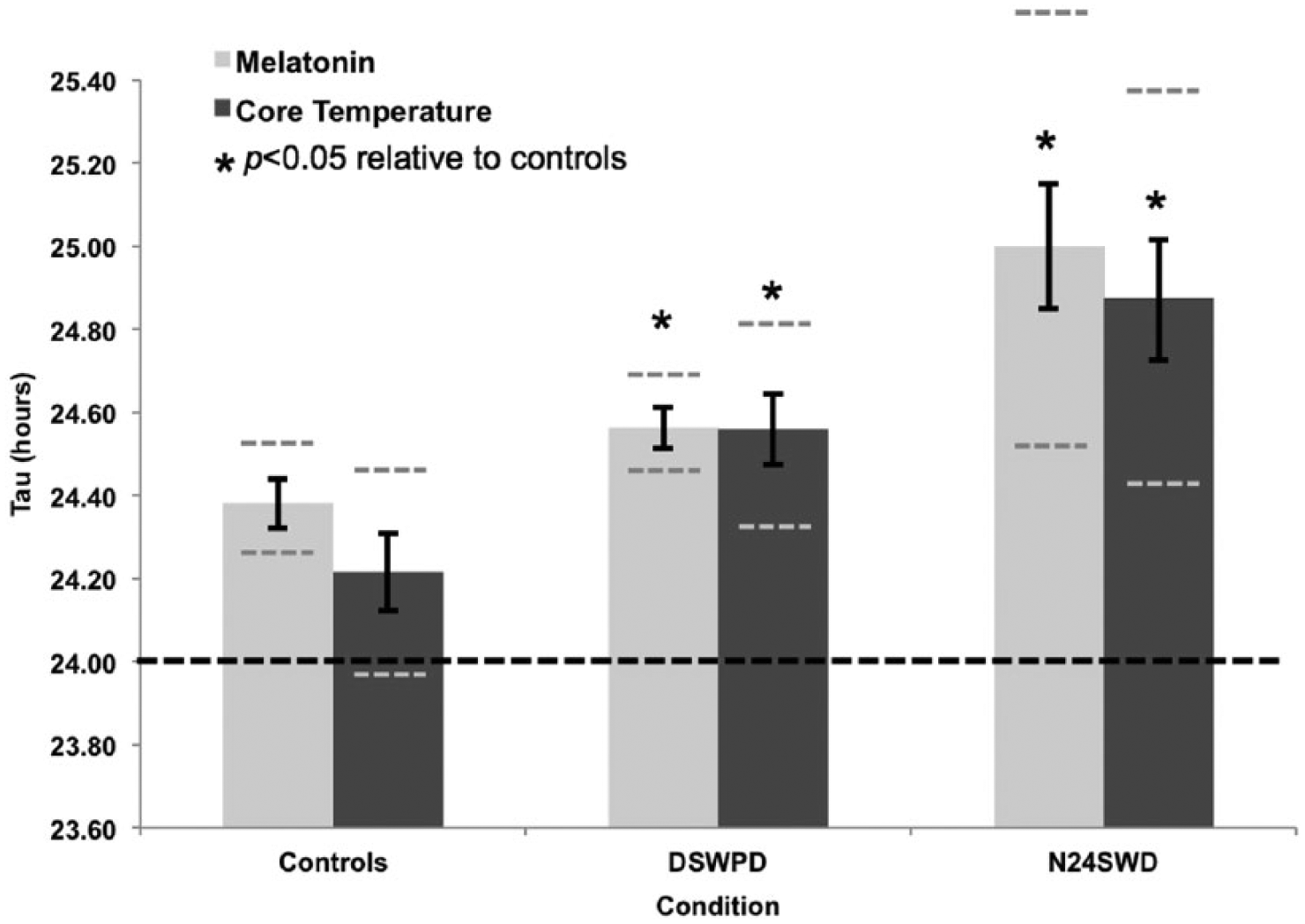
Mean circadian period lengths (taus) of core body temperature (Tτ: dark-colored bars) and salivary melatonin (Mτ: light-colored bars) in normally entrained controls, DSWPD patients, and N24SWD patients measured during an 80-h modified-constant routine with 20- to 40-min alternating ultradian cycles. DSWPD patients had significantly longer melatonin rhythm taus than controls (p = 0.03, d = 0.70) and longer temperature rhythm taus relative to controls (p = 0.01, d = 0.80). Melatonin and temperature (24 h 52 min ± 17 min) taus were longer in N24SWD patients than in DSWPD patients (p = 0.007, p = 0.06) and controls (p < 0.001, p = 0.02, respectively). Black vertical lines indicate standard error of means, gray dashed horizontal lines indicate 95% confidence intervals, and the black dashed horizontal line represents the 24-h timing of the earth’s daily rotations.
The main aim was to compare circadian rhythms of DSWPD patients and controls; therefore, a 2 (tau marker: temperature, melatonin) × 2 (group: DSWPD, control) repeated-measures ANOVA was used to compare taus in DSWPD patients and controls. A significant effect of group was found [F(41,1) = 8.61, p = 0.006, η2 = 0.18], demonstrating that the taus of core temperature and salivary melatonin in DSWPD patients (M = 24 h 34 min, SE = 3 min) were significantly longer than those of controls (M = 24 h 17 min, SE = 4 min). Independent-samples t-tests were further used to verify the simple effects, which showed significantly longer melatonin (p = 0.03, d = 0.70) and core temperature (p = 0.01, d = 0.80) taus in DSWPD patients compared with controls. Neither a significant interaction nor main effects of tau marker were found. There was also a significant correlation between core body temperature and melatonin taus (r = 0.32, p = 0.04), suggesting that those patients with longer temperature tau tended to have longer melatonin rhythm tau (see Fig. 6). Furthermore, N24SWD patients had even longer melatonin (p = 0.007) and temperature taus (p = 0.06) than both DSWPD and control participants (Mτ p < 0.001; Tτ p = 0.02).
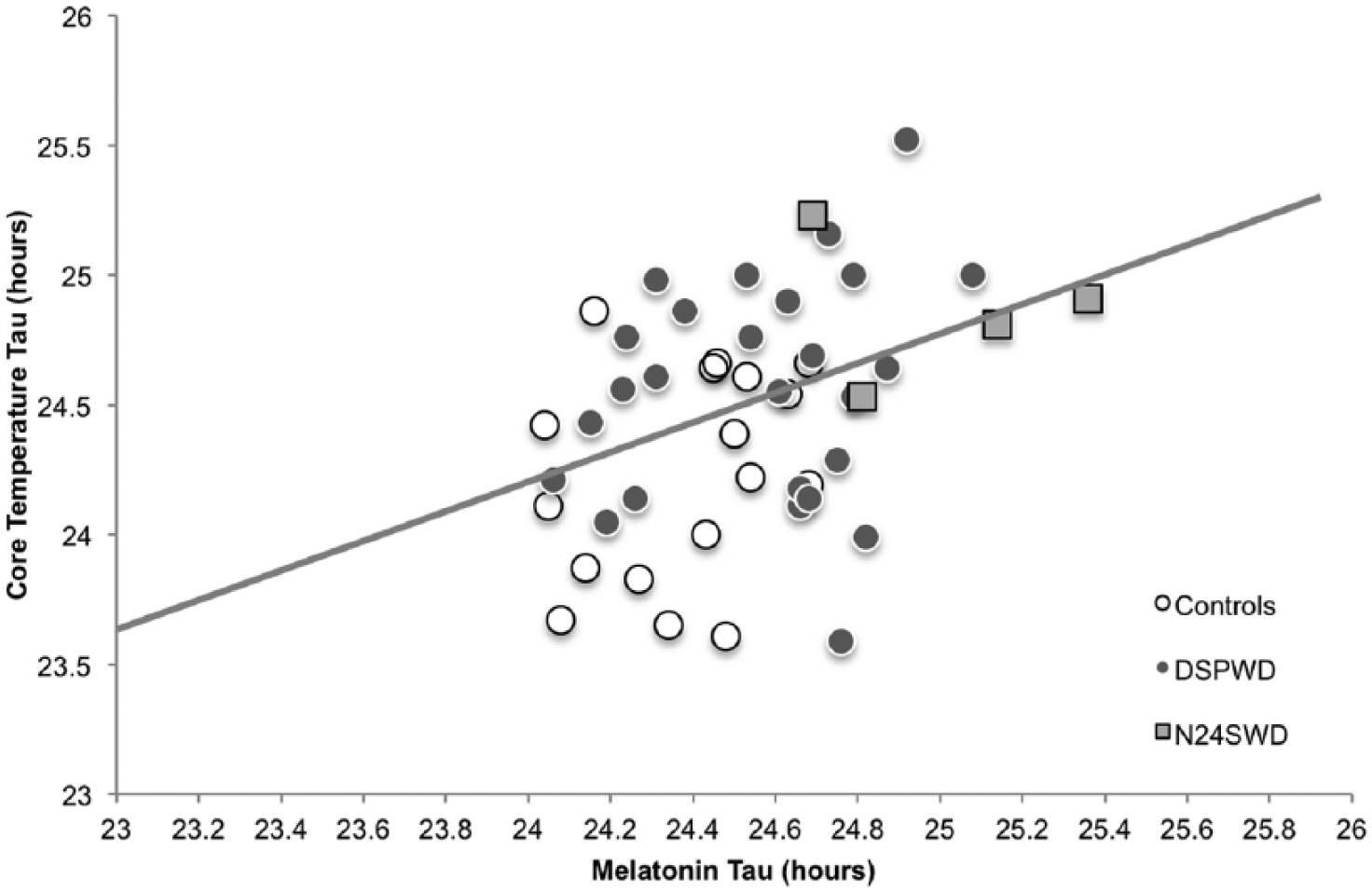
Period lengths (taus) of circadian rhythms as measured by salivary melatonin (x-axis) versus core body temperature (y-axis) of DSWPD patients, N24SWD patients, and controls. Circadian taus for the overall sample were positively correlated (r = 0.38, p = 0.009) and remained correlated when N24SWD patients (r = 0.32, p = 0.04) were removed from the analysis.
Our samples included more male than female participants, particularly in the DSWPD group (65% min) compared with controls (56% min). Given the report by Duffy et al. (2011) that females have shorter taus compared with males, we tested the imbalance in our groups. An independent-samples t-test revealed that males had overall longer melatonin rhythm taus (Mτ 24.59 h ± 0.30 h) compared with females (Mτ 24.42 h ± 0.29 h [1-tailed t(46) = 1.88, p = 0.035]) but did not have longer temperature taus (Tτ 24.47 ± 0.46 h vs. Tτ 24.44 ± 0.46 h [t(46) = 0.14, p = 0.898]).
Hence, two 1-way between-subjects ANCOVAs were carried out to assess gender bias on taus between groups. These were conducted separately for Mτ and Tτ. Checks were carried out to confirm homogeneity of regression and linear relationship between covariate and dependent variable. The between-subjects factor comprised 3 groups: DSWPD patients, N24SWD patients, and controls. The covariate comprised gender, and this was not significantly related to the differences in Mτ [F(1,48) = 2.51, p = 0.121, η2 = .05]. Adjustment for this covariate continued to result in a statistically significant effect of the between-subjects factor of condition [F(2,48) = 8.81, p = 0.001, η2 = 0.30]. The adjusted Mτ mean for DSWPD patients was 24.55 h ± 0.05 h, N24SWD patients 24.99 h ± 0.13 h, and controls 24.39 h ± 0.06 h.
Similarly, gender was not significantly related to the differences in Tτ [F(2,48) = 0.08, p = 0.78, η2 = .002]. Adjustment for this covariate continued to result in a statistically significant effect of the between-subjects factor of condition [F(2,48) = 5.81, p = 0.006, η2 = 0.21]. Again, the adjusted Tτ mean for DSWPD patients was Tτ 24.56 h ± 0.08 h, N24SWD patients 24.87 h ± 0.21 h, and controls Tτ 24.21 h ± 0.10 h.
Circadian Rhythms and Sleep Timing
Results suggest that significant tau differences could contribute to delays in sleep timing. To test the contribution of a longer circadian tau on delayed sleep timing, a multiple regression analysis was used. Sleep timing is essentially the major concern for DSWPD patients; therefore, bedtime and get-up times from objective actigraphy data were used as outcome variables. Again, due to a lack of a meaningful central tendency in N24SWD measures, these patients were excluded from the analyses. Specifically, 2 simultaneous multiple regressions were conducted to investigate whether Mτ and Tτ could predict bedtimes and get-up times in the DSWPD group. The results of the simultaneous multiple regressions are presented in Table 4, indicating that taus explained 12% of the variance in sleep onset [R2 = 0.117, F(2,42) = 2.59, p = 0.09] and 19% of the variance in sleep offset times [R2 = 0.189, F(2,42) = 4.442, p = 0.02]. In particular, Mτ was the significant predictor of sleep delays but not Tτ.
Prognostic power of salivary melatonin and core body temperature period lengths (taus) to estimate sleep delays as measured by bedtime and get up time from actigraphy.
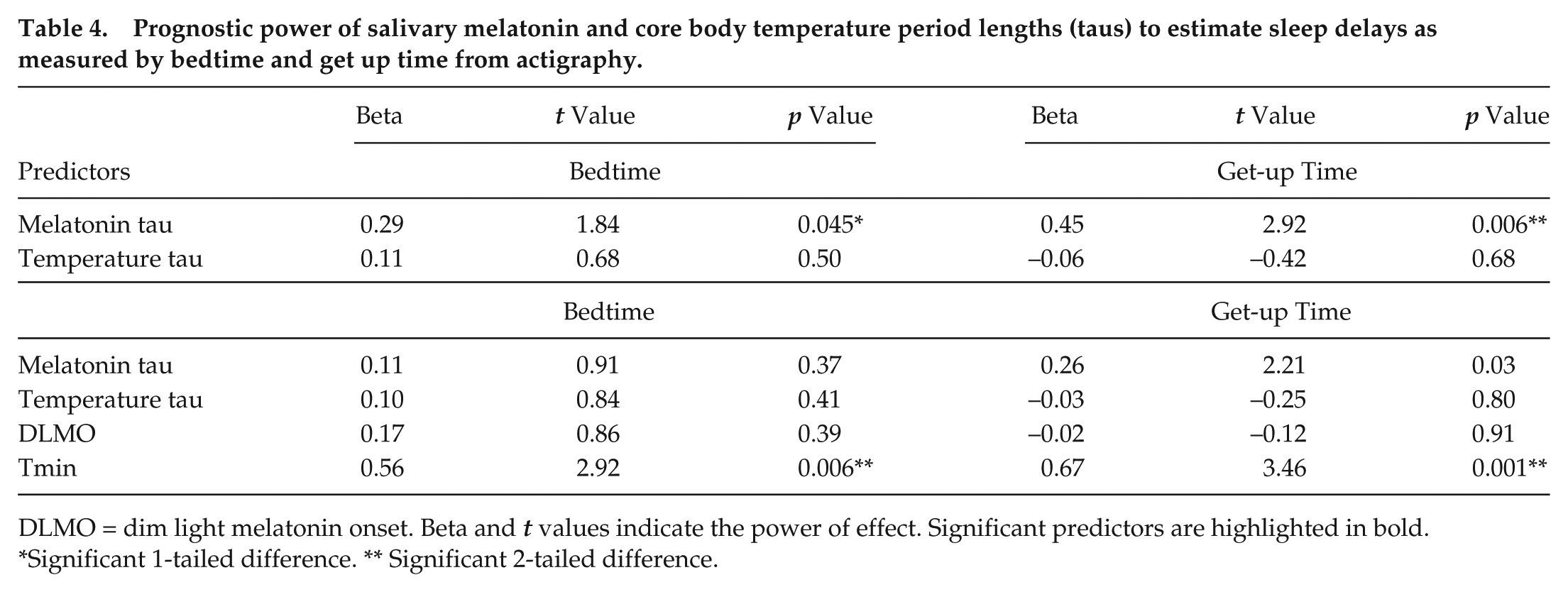
DLMO = dim light melatonin onset. Beta and t values indicate the power of effect. Significant predictors are highlighted in bold.
Significant 1-tailed difference. ** Significant 2-tailed difference.
Given that DSWPD is caused by a delay in circadian phase, we wanted to further investigate the predictive power of both phase delays and tau elongation on sleep delays. The same simultaneous regressions were conducted with the addition of DLMO and Tmin as predictor variables. The results of these analyses indicate that together, longer taus and phase delays explain 58% of the variance in sleep onset [R2 = 0.579, F(2,42) = 12.71, p < 0.001] and 66% of the variance in sleep offset times [R2 = 0.656, F(2,42) = 17.62, p < 0.001] (Table 4). This implies that although longer taus affect sleep timing, circadian phase delays (in particular Tmin timing) were better predictors of sleep timing, at least for the week prior to the laboratory determination of circadian phase delay.
Discussion
Using an 80-h ultradian constant routine, the aim of the present study was to investigate salivary melatonin and core body temperature circadian rhythms in DSWPD patients compared with controls. Only 2 previous studies have investigated core temperature taus of DSWPD patients and controls. Campbell and Murphy (2007) studied a single DSWPD patient (Tτ 25.38 h) relative to 3 controls in a sleep-wake self-selected free-running protocol (Tτ 24.44 h), while our group (Micic et al., 2013) studied 6 DSWPD patients (Tτ 24.90 h) and 7 controls (Tτ 24.48 h) in an ultradian routine. Both studies provide preliminary empirical evidence of longer taus in DSWPD patients. The present investigation was the first to replicate these circadian rhythm findings, in the largest sample to date, of DSWPD patients compared with controls and the first to include the melatonin circadian rhythm. We found that the timing of circadian rhythms on the first night of in-laboratory measurement occurred significantly later in DSWPD patients relative to normal sleepers. Moreover, taus of DSWPD patients were significantly longer relative to controls. Therefore, outcomes are consistent with previous extensive reports of circadian delays of DLMO and Tmin as well as longer circadian taus in DSWPD patients (Campbell and Murphy, 2007; Micic et al., 2013).
In addition, this is the first study to investigate taus of DSWPD patients with those diagnosed with N24SWD rhythms (“free running”), which is another CRSWD. This group was also expected to exhibit significantly longer endogenous taus relative to normal sleepers. During screening, the serendipitous group of 4 N24SWD patients identified exhibited taus that were longer still compared with both the control and DSWPD groups. The measured DLMO of these 4 patients was delayed by an average of 1 h per 24 h during the ultradian routine. Likewise, core body temperature exceeded the solar cycle by 52 min each day. These results are consistent with recent findings of Kitamura et al. (2013), who were the first to investigate melatonin rhythm taus in 6 full-sighted N24SWD rhythm patients. In a 7-day forced desynchrony protocol, the investigators found that the endogenous DLMO taus of N24SWD patients were significantly longer than their intermediate chronotypes, who were comparable to our controls as intermediate chronotypes. Thus, both studies have shown longer than normal melatonin rhythm taus in the N24SWD patients to comparable significance levels (p < 0.002).
The present results suggest that longer taus exhibited by DSWPD and N24SWD rhythm patients under nonentrained conditions may contribute significantly to the phase-delay of their major sleep periods. In the absence of 24-h zeitgeber exposure in our laboratory protocol, patients have a greater tendency to delay their circadian rhythms compared with normal sleeping individuals. Slower oscillations of the endogenous body clock, or suprachiasmatic nucleus (SCN), can potentially delay patients’ peak alertness (Duffy et al., 1998) and sleepiest phases to a later time compared with normal sleepers and conventional sleep times, despite patients’ desires and occasional attempts to sleep at more conventional earlier times. Not only were the circadian taus longer in the patients than controls, but there were significant relationships within the DSWPD group between tau length and typical wake times.
Once delayed, the DSWPD patients’ sleepiest phase will occur significantly later (e.g., Tmin = 0711 h ± 2 h 10 min according to Table 1), hence posing extreme physiological difficulty for DSWPD patients to get out of bed when socially necessary at this time. Similarly, a later peak alertness period in DSWPD (e.g., 2100-0100 h) would coincide with conventional bedtimes and inhibit sleep onsets, thus postponing patients’ sleep onset time to approximately 0200 h (Table 1). This circadian shift can explain DSWPD patients’ recurrent inability to fall asleep earlier, even when sleep debt has built up over consecutive nights of insufficient sleep.
Our findings also explain patients’ tendency to sleep later on the weekends to repay their sleep debt (Taylor et al., 2008; Yang and Spielman, 2001). Patients who have repeated nights of late bedtimes and early rise times to meet commitments will likely compensate for sleep loss on their free days. On average, our DSWPD patients were falling asleep at 0137 h on commitment days and 0241 h on free days while controls slept at 2308 h on commitment days and only ~21 min later on their free days (Table 1). Although both groups spent more time in bed on free days relative to commitment days, on their free mornings, DSWPD patients slept 2.5 h longer than controls. This extra sleep-in will avoid the morning bright light that entrains the circadian system, thus enabling the endogenous circadian system tau to be unmasked and uninhibited by morning light. Several studies indicate that in healthy sleepers, delaying wake-up times just on the weekend results in subsequent circadian delays comparable to their estimated average ~15 min longer than 24-h taus (Taylor et al., 2008; Yang et al., 2001). Therefore, patients exhibiting a significantly longer tau would be expected to yield comparably greater circadian delays, thus perpetuating a vicious cycle and further contributing to circadian and sleep delays.
Similarly, the near-25-h pace at which N24SWD patients’ SCNs tend to oscillate would make it nearly impossible for rhythms to synchronize to the 24-h day. A daily pressure to phase-delay by 1 h would require the N24SWD patient to effectively phase advance by that same amount each day to remain entrained. Phase response curves to singe pulses of light stimulation would suggest that a phase advance of an hour would require a long (4-7 h) and intense (~10,000 lux) light pulse (Khalsa et al., 2003; Dawson et al., 1993). Therefore, only the most arduous and strict light therapy regime is likely to have any success in synchronizing the circadian system of N24SWD patients to the 24-h clock. Such lighting conditions are unlikely to be encountered in the typical postawakening environment of an N24SWD patient.
The findings of desynchrony are particularly prominent in totally blind patients (Sack et al., 2000) whose blindness arises from ocular or retinohypothalamic track pathology. These patients cannot receive photic stimulation necessary to entrain to the natural 24-h light-dark cycle (Saper et al., 2005). In essence, these blind patients’ sleep patterns passively and temporally “free-run” because circadian taus are unperturbed by the exogenous light-dark cycle that entrains healthy humans. In a crossover treatment study of 7 N24SWD blind patients, oral melatonin effectively entrained 6 patients whose taus ranged between 24.2 and 24.6 h; however, the patient with a 24.9-h tau did not achieve the same entrainment (Sack et al., 2000). Hence, endogenous pressure from the SCN would be a prominent factor making it almost impossible for patients with significantly longer taus to maintain synchrony with the 24-h light-dark cycle.
Other Etiological Contributors
It seems that the endogenous SCN output is only a single element among others that contributes to sleep misalignments. There is evidence from our findings and particularly those of Kitamura et al. (2013) that not all patients with N24SWD rhythms exhibit significantly longer taus. Our results indicate overlap in the length of taus among all 3 groups, and approximately half of DSWPD patients exhibited taus that were within the “normal” range in the present study of less than 24.6 h (Fig. 6). This suggests that longer taus can explain about 12% to 19% of the variance in DSWPD patients’ sleep timing and that other factors can also contribute to misalignments in DSWPD and N24SWD disorders. Previous research suggests that both groups of patients may also exhibit abnormal functioning of entraining mechanisms. DSWPD patients may have abnormal hypersensitivity of evening melatonin suppression, and thus be prone to greater phase delay, when exposed to light in the evening (Aoki et al., 2001; Cain et al., 2013). However, a reversed abnormality in the sense of hyposensitivity to morning light would reduce its phase-advancing effect and the braking effect of morning light on the stronger than normal tendency to phase delay in these patient groups. This implicates the functioning of the retinohypothalamic tract. A dysfunction particularly in the ganglion cells that contain melanopsin has been linked to seasonal depression (Berson, 2003) and could also explain the association between depression and circadian misalignment (Roecklein et al., 2013), also apparent in our group of patients showing higher depression scores (see DASS scores in Table 1). Further studies are important to specifically investigate the contribution of phase change light sensitivity in DSWPD and N24SWD patients, both in the evening and in the morning.
A potential abnormality in pineal activation has been recently suggested, arising from the melatonin secretion profiles of DSWPD patients in the present study. Compared with the normally sleeping control participants, the DSWPD patients had a reduced secretion of melatonin between the DLMO and melatonin acrophase (Micic et al., 2015). The cause of this “blunted” initial surge of melatonin needs to be investigated. Is it a deficient signal from the SCN to the pineal or an underresponsive pineal gland? Its effects may be to feed back less circadian advancing effect to the SCN compared with normal controls and less ability to counteract the stronger tendency to delay with their longer taus.
There also appears to be a clear split in the circadian phases of DSWPD patients (Fig. 4). Fourteen were imbedded in the cluster of “late” controls and may be classed as “non–circadian delayed” DSWPD, and 12 had much later phase markers and could be classed as “circadian delayed” DSWPD. This adds weight to the emerging literature that approximately half of those clinically diagnosed with DSWPD do not portray abnormal intrinsic delays of core temperature and melatonin rhythms (Rahman et al., 2009). It also reinforces the recommendation for objective phase assessment as part of DSWPD diagnosis (Keijzer et al., 2014; Rahman et al., 2009). Clinicians may be better informed about diagnosis and efficacy of treatment by separating patients who display circadian misalignment and those who show sleep delays but with normal circadian phase. For example, the former may respond more effectively to chronobiologic therapies (Wirz-Justice et al., 2004), while the latter may require regimes that focus predominantly on cognitive behavioral therapy (CBT; Gradisar et al., 2011).
Clinical Recommendations
The clinical implications of these findings are to investigate treatment options that can specifically target prolonged taus. While these treatment options may not yet be viable, genetic studies and further identification of polymorphisms could point to future treatment (Kripke et al., 2014). With our current tools for chronobiologic intervention, the results support the rigorous and persistent use of morning bright-light therapy and evening melatonin administration to palliate the conditions. This method of treatment has been successful for treating DSWPD as well as N24SWD disorder. Combination treatments of both bright-light therapy and melatonin administration show greatest efficacy in advancing patients’ sleep timing (Keijzer et al., 2014; Wirz-Justice et al., 2004). It is crucial to recognize that significantly longer taus can explain the frequently reported long-term relapse following the current forms of therapy (Regestein and Monk, 1995; Weitzman et al., 1981; Wilhelmsen-Langeland et al., 2013; Saxvig et al., 2013).
While this treatment may allow patients to wake up at a regular and conventional time, the persistent strong tendency to delay coming from an abnormally long tau could make them susceptible to relapse. Indeed, previous results of a randomized controlled trial with morning bright-light therapy and evening melatonin administration indicated that gradually advancing bed and rise times significantly advanced DLMO timing and improved subjective sleepiness, fatigue, and cognitive performance in both treatment conditions (Saxvig et al., 2014; Wirz-Justice et al., 2004; Weitzman et al., 1981). However, only patients who received combination melatonin and morning light therapy showed favorable outcomes in the 3-month follow up. Therefore, the combination of early evening low-dose melatonin administration and morning bright-light stimulation would certainly be indicated for the circadian-delayed and longer-tau DSWPD and N24SWD patients.
Nevertheless, it would seem that that delayed sleep in DSWPD patients without circadian delays and tau elongation is not due to circadian misalignment and, therefore, chronobiological treatments are unlikely to be effective. This group of patients may suffer adjustment difficulties or maladaptive personality traits (Wilhelmsen-Langeland et al., 2013; Micic et al., Forthcoming), rather than an abnormal circadian rhythm characteristic (Dagan and Eisenstein, 1999; Regestein and Monk, 1995). Hence, for this clinical subsample, CBT aimed at increasing behavioral control over sleep-wake schedules, teaching good sleep hygiene and circadian education, and mitigating maladaptive beliefs that contribute to inappropriate sleep-wake schedules would be most indicated (Gradisar et al., 2011). In any case, without a measure of circadian phase in the clinical diagnosis of DSWPD, all tools in the armamentarium including chronobiological and CBT would be indicated.
Limitations and Conclusions
In the past, resultant measures of circadian rhythms, particularly taus, have varied slightly between studies associated with methodological differences. From forced-desynchrony studies over at least 1 week duration, the average tau of healthy individuals is estimated to be 24.15 h (Czeisler et al., 1999; Duffy et al., 2001; Klerman et al., 2002). In the present study using the relatively short duration (80-h) ultradian-modified constant routine method, we were able to replicate several previous findings (Duffy et al., 2011; Campbell and Murphy, 2007; Kripke et al., 2005; Micic et al., 2013) and found taus in our healthy participants of 24.22 to 24.37 h. While this method may yield slightly longer estimates of tau than the forced desynchrony method, we found comparably longer taus in the DSWPD and N24SWD patients.
Several methodological considerations and limitations should be noted when interpreting the findings of the current study. Empirical evidence shows that the period estimates are different between free-running and forced-desynchrony studies (Klerman et al., 2002). However, the method used in the present study has not been compared with the other protocols. Hence, explanations do not exist as to why the ultradian method finds taus that typically range between the other 2 methods of assessing period lengths. Although we have established differences of taus between groups, a possible perceived limitation is that we cannot vouch for the absolute accuracy of this method in measuring taus. Future work is important to compare taus derived from both normal and DSWPD patients in both methods.
Additionally while body temperature exhibits a circadian rhythm, this rhythm can be masked by factors such as posture, activity, and sleep. In this highly controlled study, sleep duration during the 20-min napping opportunities is the only factor not controlled. We know that sleep can affect core body temperature (Lack and Lushington, 1996), but these sleep opportunities do not affect individuals’ Tmin timing when measured by an ultradian routine compared with a constant routine; therefore, the period estimate would not be affected either. Significant correspondence between temperature and melatonin taus in the present study further strengthens this conclusion since melatonin is independent of sleep-wake in the dim light laboratory environment (Benloucif et al., 2005; Klerman et al., 2002).
The present outcomes suggest that overall, DSWPD patients have significantly longer taus of salivary melatonin and core temperature rhythms than controls, and the taus of N24SWD patients were longer still. As such, taus longer than normal are likely to contribute to sleep delays in patients with DSWPD and N24SWD. At least in the DSWPD sample, further research is necessary to untangle various other etiological factors that pose difficulties in implementing successful therapies and ensuring long-term efficacy of treatment interventions.
Footnotes
Acknowledgements
This study was funded by a grant from the Australian Research Council (Discovery Project DP120101401).
Conflict of Interest Statement
The author(s) declared the following potential conflicts of interest with respect to the research, authorship, and/or publication of this article: L.L. is a shareholder in Re-time Pty Ltd. M.G. has performed paid speaking engagements for the Australian Psychological Society and is a shareholder in ResMed. H.J.B. was paid speaker for SLEEP meeting CME course (2010, 2012), Society of Behavioural Sleep Medicine CME (2011), Ohio State University CME (2011), and University of Chicago CME (2012). S.F. was supported to give a keynote at Working Time Society, Denmark (June 2015).
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.