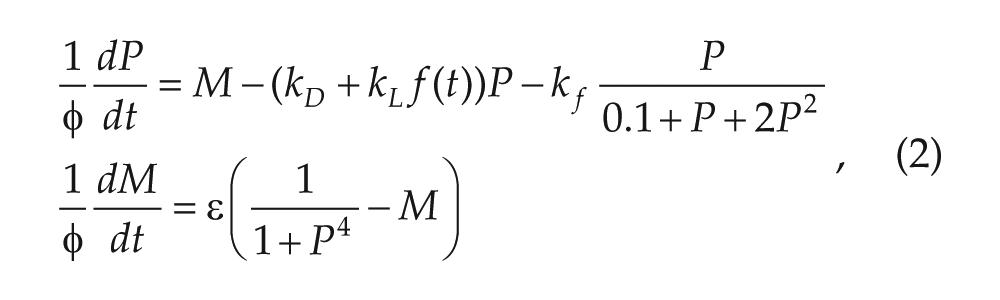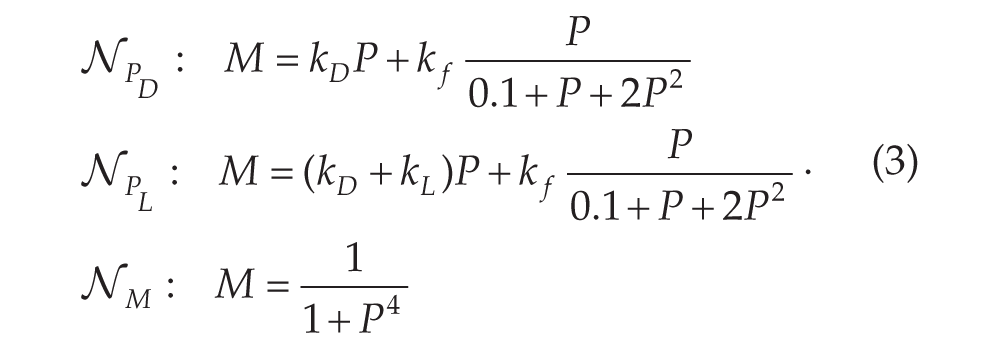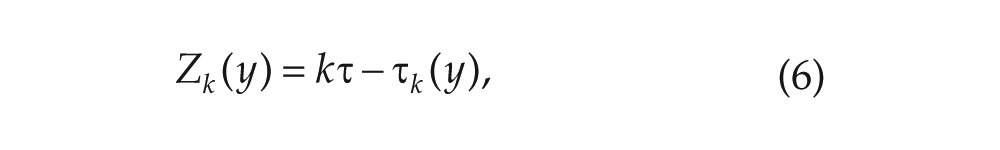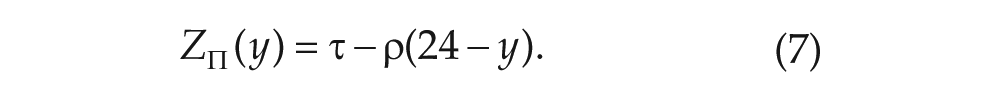Circadian oscillators found across a variety of species are subject to periodic external
light-dark forcing. Entrainment to light-dark cycles enables the circadian system to align
biological functions with appropriate times of day or night. Phase response curves (PRCs)
have been used for decades to gain valuable insights into entrainment; however, PRCs may
not accurately describe entrainment to photoperiods with substantial amounts of both light
and dark due to their reliance on a single limit cycle attractor. We have developed a new
tool, called an entrainment map, that overcomes this limitation of PRCs and can assess
whether, and at what phase, a circadian oscillator entrains to external forcing with any
photoperiod. This is a 1-dimensional map that we construct for 3 different mathematical
models of circadian clocks. Using the map, we are able to determine conditions for
existence and stability of phase-locked solutions. In addition, we consider the dependence
on various parameters such as the photoperiod and intensity of the external light as well
as the mismatch in intrinsic oscillator frequency with the light-dark cycle. We show that
the entrainment map yields more accurate predictions for phase locking than methods based
on the PRC. The map is also ideally suited to calculate the amount of time required to
achieve entrainment as a function of initial conditions and the bifurcations of stable and
unstable periodic solutions that lead to loss of entrainment.
The ability of circadian oscillators to entrain to daily cycles of light and temperature has
been called both the most important and the most misunderstood property of circadian
rhythmicity (Johnson et al., 2003).
Entrainment implies that an endogenous oscillator has matched its period to that of an
external periodic forcing and has established a stable phase relationship with the forcing
signal. The process of circadian entrainment has been studied extensively using tools from
dynamical systems and oscillator theory (Ananthasubramaniam et al., 2014; Bagheri et al., 2008; Bordyugov
et al., 2013a; Gu et al.,
2012, 2013; Indic et al., 2006; Johnson et al., 2003; Leise and Siegelmann, 2006; Leloup and Goldbeter, 2013; Ramkisoensing et al., 2014; Rand et al., 2004; Roenneberg et al., 2003; Serkh and Forger, 2014; Woller et al., 2014). Perhaps the most
commonly used method relies on phase response curves (PRCs) that measure the change in the
phase of an endogenous limit cycle oscillation (typically in constant darkness or DD) induced
by a perturbation (typically a light pulse) as a function of the phase at which the
perturbation is applied (Johnson,
1999). Alternatively, one could perturb an oscillator in constant light (LL) with
dark pulses. Such PRCs can be constructed for light or dark pulses of arbitrary strength and
duration. However, for a PRC to accurately predict properties of entrainment to periodic light
or dark pulses, the perturbations must be weak or brief enough that the oscillator relaxes
back to the DD or LL limit cycle attractor before the next pulse arrives. Since circadian
oscillators are naturally subject to forcing in which the light duration is several hours
followed by several hours of darkness, the PRC may not be ideally suited to accurately
determine entrainment in this context. In fact, we show that PRCs do not accurately predict
the phase of entrainment in 3 different mathematical models of circadian clocks that are
subjected to photoperiods with substantial amounts of both light and dark, such as 12-h:12-h
light-dark (12:12 LD) cycles. For these LD cycles, the entrained solution is best thought of
as a combination of 2 limit cycle attractors (i.e., the DD and LL limit cycles), rather than
as a perturbation of a single limit cycle attractor (Peterson, 1980).
In this paper, we introduce a new tool, a 1-dimensional entrainment map,
which is not based on perturbing the DD or LL oscillator. Instead, the map uses information on
how both the 12-h light period and the 12-h dark period affect cycle duration as a function of
the time elapsed since the lights last turned on. We show that the stable fixed point of the
map corresponds to a stable LD-entrained solution. Properties of the entrained solution, such
as the range of entrainment as a function of parameters or the speed with which entrainment
occurs, are then inferred from properties of the map. We demonstrate that the map accurately
predicts the phase of entrainment independent of the photoperiod across a variety of different
models. Our findings are consistent with previous results in the literature (Bordyugov et al., 2015; Granada et al., 2013) and provide a new
method for studying high-dimensional circadian models that are difficult to analyze
mathematically.
The entrainment map is an example of an iterated map, a class of dynamical systems in which
time is treated as discrete rather than continuous (Strogatz, 1994; Wu and Rul’kov, 1993). One-dimensional iterated maps
have the form
where is a function that maps values from an interval back into that interval.
Maps can be used as models of natural phenomena where time is inherently discrete or as tools
for analyzing continuous-time models consisting of ordinary differential equations (ODEs). The
entrainment map we develop is an example of the latter and, in principle, can be constructed
for any of the myriad ODE models of circadian clocks in the literature, from detailed models
of molecular clocks in a variety of species to phenomenological models of human circadian
rhythms (Bordyugov et al., 2013b;
Forger et al., 2007; Gonze, 2011; Roenneberg et al., 2008). The maps live in lower
dimensional spaces than the original systems, which facilitates geometric visualization and
the understanding of complicated dynamics. For example, many circadian ODE models exhibit
limit cycle oscillations. These periodic solutions correspond to fixed points of the maps.
Determining properties of these steady-state solutions, such as stability, is much easier for
fixed points than for limit cycles. Furthermore, solution trajectories of maps are found by
iterating the function to obtain a sequence of values that dictate the behavior of solutions of the map and, as a result,
the corresponding ODE (Hirsch et al.,
2013).
We construct entrainment maps for circadian oscillators under forcing by a periodic
light-dark cycle. For this map, the variable tracks the number of hours that have passed in the th light-dark cycle between the lights turning on (defined as ) and the oscillator reaching a certain location in phase space. Both
and take values between 0 and 24, where and are equivalent. Thus, maps a circle onto itself and is referred to as a circle
map. The theory of circle maps is well-developed and has been applied extensively
to study cardiac rhythms and other types of biological oscillators (Glass, 1991; Keener and Glass, 1984; Winfree, 2001). More generally, the search for an
entrained solution is an example of a classic problem in which an oscillator is forced by an
external periodic input. Much prior theoretical work has been done in the context of circadian
oscillators (Bordyugov et al., 2015;
Roenneberg et al., 2003, 2010a). What is common to all of these
studies, as well as ours, is that there exists a range of parameters over which the period of
the oscillator becomes equal to that of the forcing, and stable one-to-one entrainment
occurs.
In this paper, we focus on 3 distinct circadian oscillator models of different complexity.
The first is the Novak-Tyson (NT) model of the Drosophila molecular clock
(Tyson et al., 1999). This is a
2-dimensional model for which we provide a detailed derivation of the entrainment map. The
other two models we consider are the 3-dimensional Gonze model (Gonze et al., 2005) and the 180-dimensional Kim-Forger
model (Kim and Forger, 2012) of the
mammalian molecular clock. For all of these models, we demonstrate that the map accurately
predicts the stable entrained phase relative to the underlying light-dark forcing.
Additionally, we show that the map enables us to characterize, across different models, how
properties such as the range of entrainment depend on parameters associated with the intrinsic
oscillator and the external forcing. In this sense, the map reveals certain universal
properties about how different circadian models respond to changes in parameters that are
common to all models, such as the speed of the intrinsic oscillator, light intensity, and
photoperiod. In turn, this provides insights and the ability to make predictions about higher
dimensional models that are not so easily gained through direct simulations.
Model and Methods
In this section we introduce the circadian clock models that we shall use in our study and
the 1-dimensional map that we have developed to analyze entrainment properties of such
models.
The Novak-Tyson Model
The NT model (Novak and Tyson,
2008; Tyson et al.,
1999) for the molecular circadian clock in the fruit fly
Drosophila contains 2 state variables representing mRNA concentration
() and protein () concentration:
where all parameters are positive. The variable represents 2 proteins (PER and TIM) that form a heterodimer and
create a negative feedback loop by inhibiting transcription of their own genes. The
parameter is taken to be small, which creates a separation of time scales between
the and variables. The parameter governs the rate of flow of trajectories in the phase plane and will
directly affect the period of any solutions we find. The parameter is the baseline protein degradation rate during darkness. In
Drosophila, light is known to increase degradation of TIM;
is the added degradation rate due to light. The parameter
captures the extent of positive feedback due to stabilization of the
protein after dimerization. The function describes the light stimulus. In complete darkness, . In complete light, . When the model is subjected to a periodic photoperiod, is chosen to be a smooth cut-off version of a periodic step function
that takes on the value during darkness and during light.
Consider first the situation of either constant darkness , or constant light, . In each case, the nullclines consist of the set of points where the
right-hand sides of the equations in Equation (2) equal 0. In particular, we
define -nullclines for each case together with a common -nullcline.
Both and form cubic shaped curves in the phase plane. We sometimes use the notation to refer to the -nullcline when it does not matter whether we are referring to dark or
light. The curve is qualitatively sigmoidal shaped. Depending on where the
and nullclines intersect, the ensuing fixed point(s) may be stable or
unstable. Any intersection that occurs on the left or right branches of -nullclines are stable, while those that occur on the middle branch will
be unstable provided that and are not too large. The standard set of parameters that we use for a
single cell produces an unstable fixed point along the middle branch. In either constant
darkness or constant light, it is straightforward to use dynamical systems methods to
prove the existence of a stable limit cycle (periodic orbit) solution, as discussed in the
Results section.
The Gonze Model and the Kim-Forger Model
We also consider 2 different models of the molecular circadian clock in mammals. The
Gonze model (Gonze et al.,
2005) is a modified version of the Goodwin oscillator (Goodwin, 1965) describing the core negative
feedback loop of the circadian cock. There are 3 state variables that represent
concentrations of mRNA (), protein (), and protein in its active form () of a clock gene such as Per or
Cry:
Specifically, is the nuclear form of PER/CRY that inhibits transcription of the clock
gene. In mammals, light is known to increase the transcription rate of
Per1 and Per2 (Golombek and Rosenstein, 2010); thus, we have
included the LD forcing in the first term of the equation. The following parameter values were used: , , , , , ,, , , , and .
The Kim-Forger model contains the core negative feedback loop as well as additional
secondary feedback loops. The model has 180 state variables representing nuclear and
cytoplasmic concentrations of mRNA and protein products for several clock genes including
Per1, Per2, Cry1, Cry2, Bmal, Npas2, and Rev-erb, as
well as other regulatory elements. For a detailed description of the full model, see Forger and Peskin (2003) and Kim and Forger (2012). Here we only
show the differential equations for the 2 state variables that receive LD forcing, which
are Per1 and Per2 mRNA in the nucleus ( and ), and the state variable that we will use to define the entrainment map
for this model, which is BMAL protein in the cytoplasm ():
where , , , and are state variables representing the probability of the E-box promoter
being activated, Bmal mRNA and CLOCK/NPAS2 protein concentrations in the
cytoplasm, and the concentration of unphosphorylated BMAL-CLOCK/NPAS2. The parameters
and are basal transcription rate constants in darkness, is additional transcription due to light, and are degradation rate constants, and is the rate constant for folding and nuclear export of mRNA. Finally,
the parameters , , , and are rate constants for BMAL translation, BMAL degradation, and binding
and unbinding of BMAL to CLOCK/NPAS2. We used the parameter values given in Kim and Forger (2012), with
, , and .
For all 3 of these models, we will show via simulation that the 1-dimensional entrainment
map yields accurate predictions for the phase of entrainment.
Effects of Sensory Input: Light and Darkness
We shall be interested in the response to 3 different scenarios: constant darkness,
constant light, and periodic switching between the two with a prescribed photoperiod. We
will show that over a wide range of parameters, the models produce stable periodic
solutions in constant darkness, referred to as DD, or in constant light, denoted LL. We
denote the “free-running” period of the DD oscillator as . In the case of periodic forcing, we follow a protocol that is often
used in experiments. Namely, we will study the entrainment of oscillators to a periodic
signal that models a photoperiod of a 24-h cycle consisting of an hour “light” interval (L) followed by a 24- hour “dark” interval (D). When an oscillator is subjected to a
light-dark photoperiod (LD), depending on parameters, it may entrain to the 24-h forcing.
By entrain we mean that the solution is periodic and that it has a period
of 24 h. If these conditions are met, we call the ensuing periodic solution an
LD-entrained solution.
To consider an oscillator subjected to periodic forcing of hours of light followed by hours of darkness, we allow to vary periodically between 0 and 1. In our simulations,
is a rectangular wave with instantaneous transitions. When analyzing the
periodically forced system, we consider to be a smooth approximation of a rectangular wave.
The Entrainment Map
The entrainment map is the primary tool we will use to understand the properties and
consequences of phase-locking. To define , we choose a Poincaré section as an dimensional hyperplane that intersects a point on the LD cycle. To
define a hyperplane, we only need to specify the value of 1 of the variables and the
direction of the flow as it crosses the hyperplane. For example, in the 2-dimensional NT
model, is a 1-dimensional line segment that intersects the LD-entrained
solution chosen along the left branch of at where . This is a natural location to place since the flow forces trajectories to enter a neighborhood of that
branch. We note that the method still works if we choose to lie along a different portion of the LD cycle, for example, away from
the left branch, as discussed in the Supplementary Material, Section R2. In the Gonze and
Kim-Forger models, there is no natural location to choose for the Poincaré section. We
choose the section in these cases by specifying a variable that does not receive the
light-dark forcing. For the 3-dimensional Gonze model, is a 2-dimensional hyperplane chosen at where . For the 180-dimensional Kim-Forger model, is a 179-dimensional hyperplane chosen at where . Assume that the oscillator has an initial condition lying on the
Poincaré section. We define to be the amount of time that has passed since the beginning of the most
recent LD cycle. When the trajectory is again on , we define the map to be the amount of time that has passed since the onset of the most
recent LD cycle. The domain and range of are the set . The domain is considered to be periodic, where and are equivalent. The Poincaré map relates the value of at one cycle to its value at the next; for . Unless otherwise noted, we limit our discussion of the properties of
the map to parameter sets where both the LL and DD limit cycles exist. Details on how we
conducted the numerical simulations to construct the map are provided in the Supplementary
Material, Section M1.
The map can be decomposed in the following way. Define a return map
that measures the time a trajectory starting on , hours into an LD cycle, takes to return to . Then . Because of the mod operation, it is not readily clear whether the
trajectory returns to in the same LD cycle in which it started or in a subsequent one. To
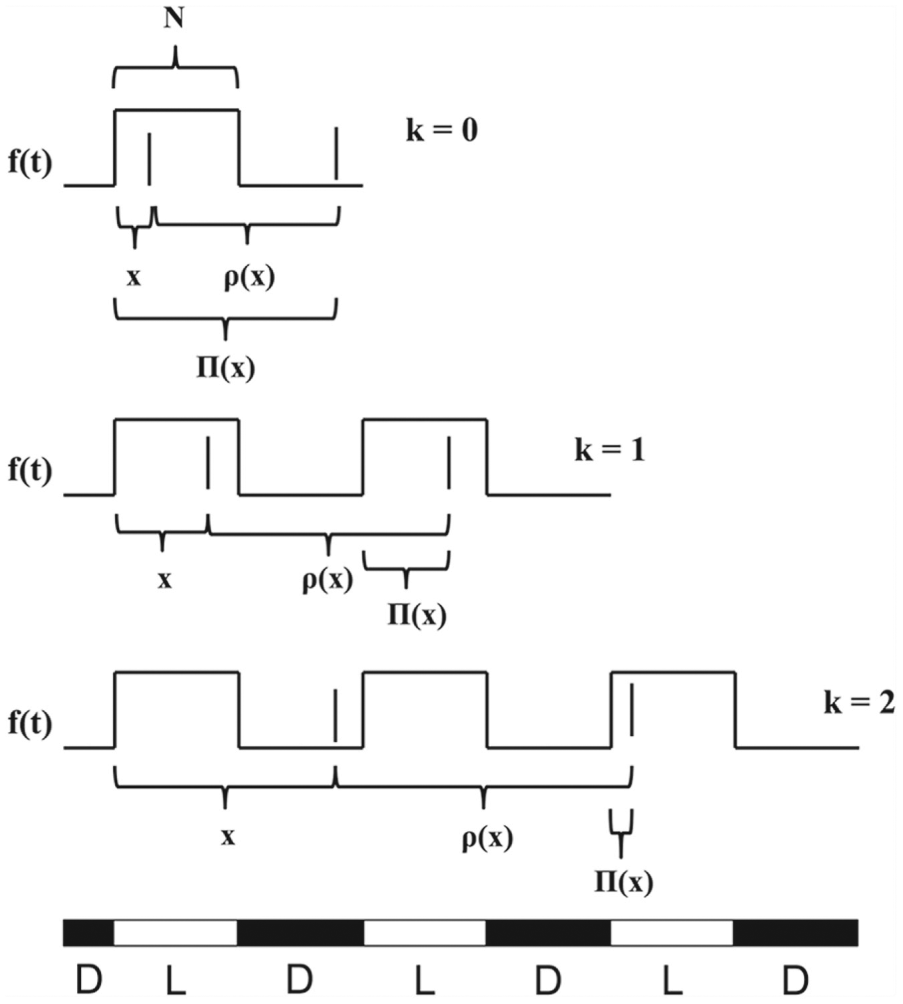
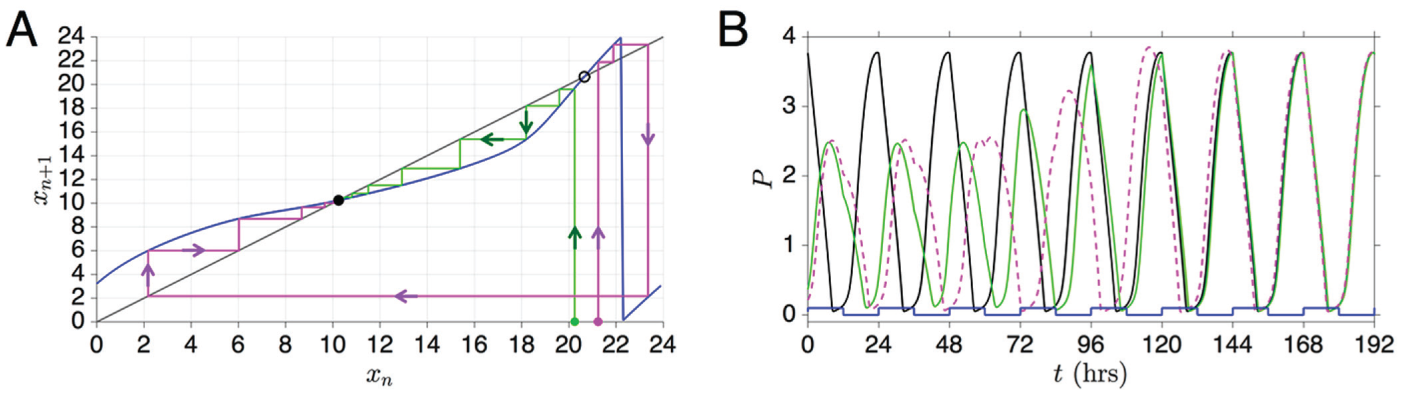
clarify this point, note that if , the trajectory will return back to within the same LD cycle in which it started. Thus, . If , then the trajectory returns in the next LD cycle and . In general, we can write , for and . The case corresponds to the trajectory returning to in the same LD cycle it started in, in the next cycle, in the subsequent cycle, and so on. See Figure 1 for an illustration.
Schematic that defines , , and in various cases. In each panel, the solid vertical lines denote 2
consecutive moments in time when the oscillator crosses the Poincaré section
. The square wave denotes , which varies between 0 and 1, and the LD photoperiod is shown at
the bottom of the figure. The time is measured as the distance in time from the start of the first
light pulse (when increases to 1) to the first crossing of , and is the distance in time between crossings of . The value measures the distance in time from the start of the most recent
onset of lights to the next crossing of . The 3 panels shown correspond to the 3 possible scenarios that
arise in defining the entrainment map.
The map measures the time it takes a trajectory leaving to return to it. It has a periodic domain , and its range lies in the set of positive numbers. It has several
generic properties: is continuous (since the forced vector field of Equation (2) is
smooth), is periodic at its endpoints (since the domain is periodic), and is decreasing on a subinterval of its domain (as shown in the Results
section).
A stable fixed point of the map , denoted , corresponds to a one-to-one entrained solution and determines the phase
of locking. A stable fixed point satisfies 2 conditions, and . Both conditions can be checked by plotting the map and determining
whether and with what slope the graph intersects the diagonal. When a stable, phase-locked
solution exists, the map is ideally suited to calculate the time to approach the stable solution
starting from any initial condition. If is a stable fixed point of the map and is a sequence of iterates of the map, we say that the solution is
entrained if there exists , such that for all , . Given an initial condition , we use the map to determine and thereby determine the time to entrainment. We will show by
comparison to direct simulations of the full system that this method is quite
accurate.
There is a considerable amount of flexibility in how the Poincaré section is chosen. This
is because of a basic theoretical result of smooth differential equations: namely, that
solution trajectories depend continuously on initial conditions. To build the map
, for each , we start with an initial condition that lies on the intersection of
and the LD-entrained solution (e.g., in the Gonze model at the value
, , and ). Because the value of is not necessarily the fixed point , the ensuing trajectory is not the entrained solution. But the theory of
smooth dynamical systems guarantees that the solution trajectory will lie close to the
LD-entrained solution for a finite amount of time. The terms close and finite can be made
mathematically precise, but for our purposes it is enough to guarantee that the trajectory
again crosses in a neighborhood of the crossing point of the entrained solution. This
basic theoretical fact is what allows the entrainment map to be constructed for
high-dimensional models for which the LD-entrained solution has first been numerically
computed. Different choices of the Poincaré section will lead to different entrainment
maps. But for any choice, the ensuing LD-entrained solutions will all have the same phase
of entrainment, as discussed in the Supplementary Material, Section R2.
The entrainment map is defined by setting as the reference phase with respect to the onset of lights in a 24-h
light-dark cycle. This is in contrast to PRC-based methods, where the reference phase is
typically assigned to a point along the DD oscillator. Although it may not seem that
assigning the reference phase to a point on the underlying light-dark forcing instead of
DD or LL makes much of a difference, we will show that doing so allows us to use
information from the DD, LL, and LD-entrained solutions to construct the map.
Additionally, the entrainment map allows the light pulse (or the dark pulse) to be split
up so that it appropriately represents the relationship of the oscillator to the ongoing
light-dark cycle and not just as a single pulse at a predetermined phase as in a PRC. As a
result, we will show that for all the models we tested, the map ends up yielding a more
accurate prediction about phase locking than the PRC.
Results
We begin by analyzing the NT model. Many of the details of the phase plane analysis can be
found in the Supplementary Material. We then turn to the Gonze and Kim-Forger models and
show that the entrainment map also works well in these higher-dimensional settings.
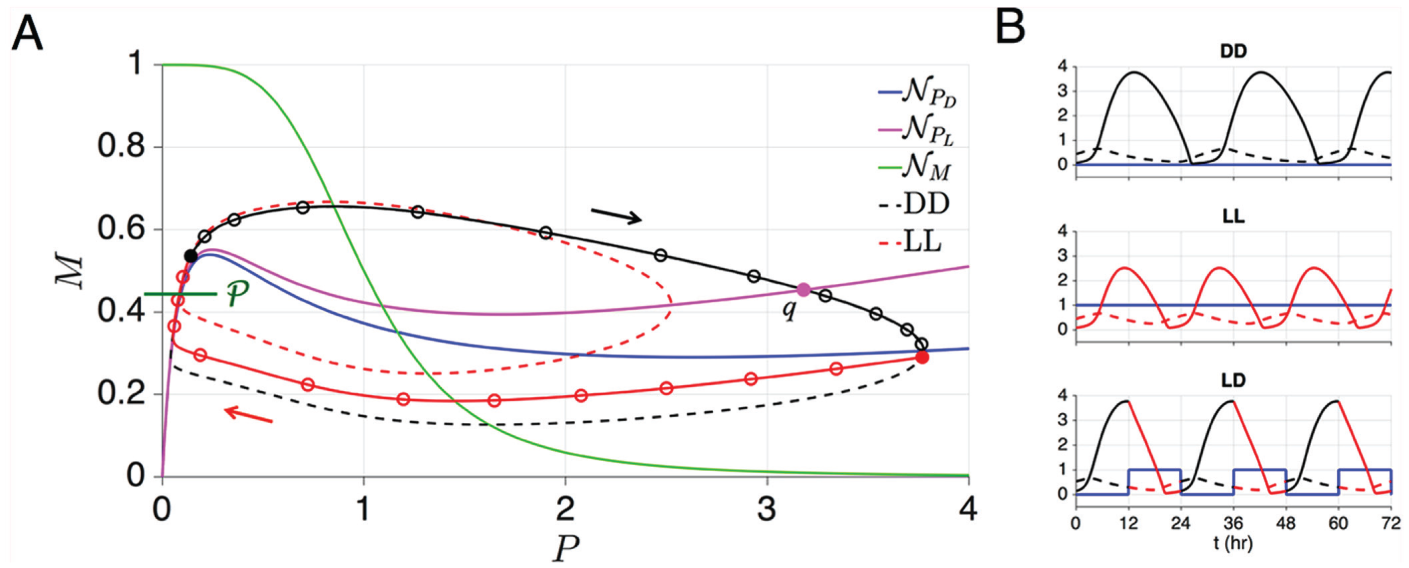
LD-Entrained Oscillations in the Novak-Tyson Model
When , Equation (2) produces a limit cycle corresponding to a DD oscillator (black
dashed curve in Fig. 2A). This
stable limit cycle exists due a simple application of the Poincaré-Bendixson theorem.
Namely, one can build a bounded, positively invariant region that surrounds the single
fixed point that lies on the middle branch of . Since this fixed point is unique and unstable, the Poincaré-Bendixson
ensures the existence of a stable limit cycle. When , the -nullcline shifts up in phase space, but the fixed point remains on the
middle branch. Again the Poincaré-Bendixson theorem applies, proving the existence of the
LL limit cycle (dashed red curve in Fig.
2A). Note that the DD limit cycle has a larger amplitude in the direction. This is a consequence of the difference in the
-nullcline between the DD and LL cases. Also note that the 2 limit cycles
share a common fast-slow structure that allows them to largely overlap near the left
branch of . Indeed, as , Equation (2) becomes singularly perturbed and portions of the LL and DD
trajectories would lie on the left and right branches of . For any , the period of the LL limit cycle is less than that of the DD limit
cycle (the opposite is true for the Gonze and Kim-Forger models, as discussed in the
section on higher dimensional models). This is a result of the faster dynamics in the LL
case due to being higher in the phase space than .
(A) The DD (dashed black), LL (dashed red) and LD (solid red-black) solutions of the
NT model are shown in the phase plane. The LD-entrained solution overlaps the DD
oscillator’s trajectory during darkness (black) and then deviates from it near
when the lights turn on (red). Trajectories move clockwise in the
plane. Open circles denote hourly intervals. The green horizontal line segment is
. The point is the intersection of the LD-entrained trajectory with
and is referred to in the Supplementary Material, Section R1. (B)
Time courses of the (solid) and (dashed) variables. The free-running period of the DD oscillator
( [blue line], top trace) is longer than that of the LL oscillator
(, middle trace). Entrainment to a 24-h periodic 12:12 forcing
(bottom trace) results in an LD-entrained oscillation.
In the presence of light-dark forcing, the ensuing trajectory spends its time in phase
space approaching the DD cycle during darkness and the LL cycle during light. This idea
was noted in earlier work by Peterson
(1980) (Johnson et al.,
2003; Pittendrigh,
1981). In some sense, the trajectory uses the stable structure of the DD and LL
limit cycles to transiently approach the appropriate one during relevant time intervals.
For this planar model, this back and forth serves to bound where in phase space the
trajectory can lie at any moment in time. The LD-entrained solution is shown in Figure 2A. Note that the entrained
solution tracks the DD limit cycle for a portion of its trajectory and then deviates from
it near . From that point, the trajectory is attracted toward the LL limit cycle,
although it reaches the left branch of the -nullcline prior to entering a neighborhood of the LL cycle. This is
because the rate of attraction to the LL limit cycle is not overly large. Open circles
along the LD-entrained solution denote hourly intervals. Figure 2B shows time traces of the DD, LL, and
LD-entrained solutions. The latter additionally shows the relationship of the entrained
solution to the light-dark pulse. For the parameters used in the simulation, the peak
value of happens to occur near the beginning of the dark period. The entrainment
map will help to explain why this is the case.
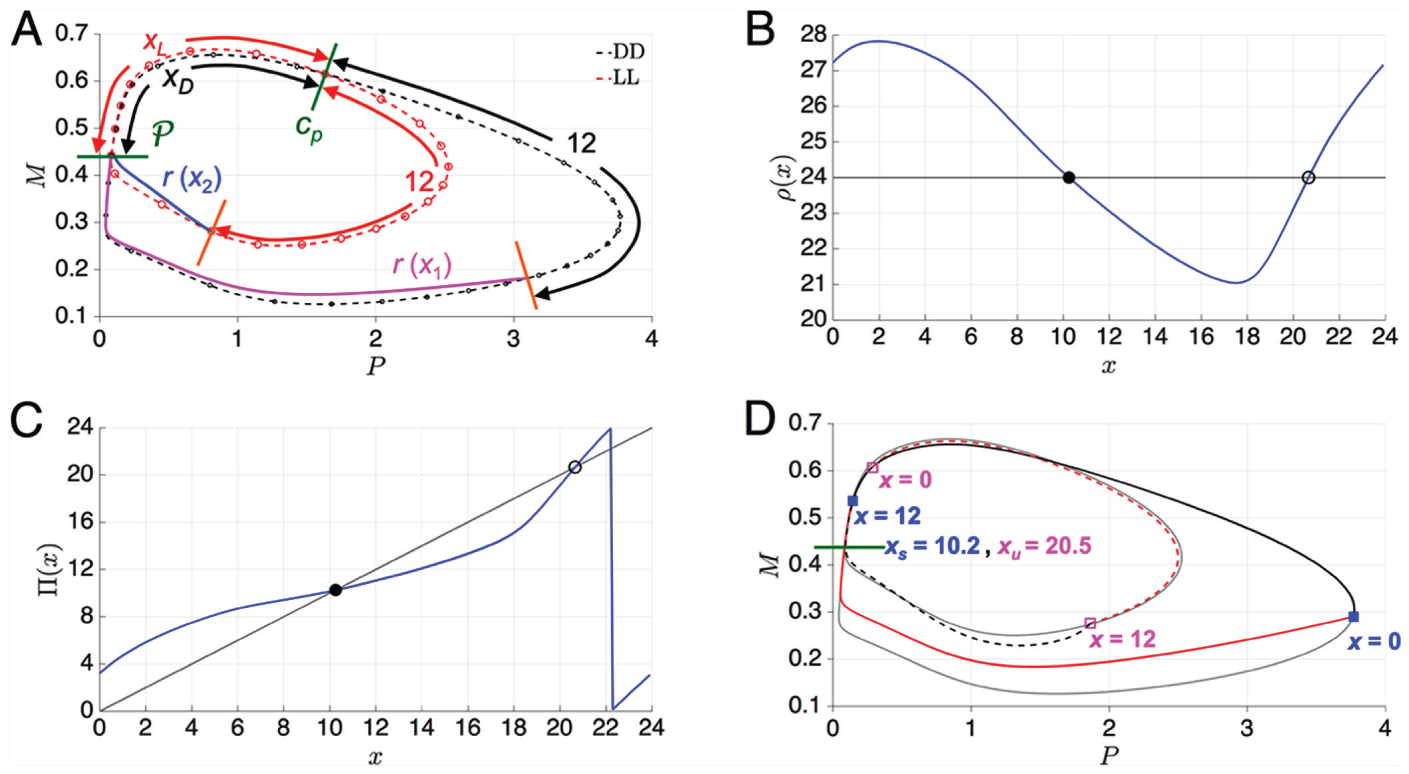
Let us consider a specific set of parameters referred to as the canonical set for the NT
model ( and a 12:12 photoperiod). Figure 3A shows an annotated phase plane of the LL
and DD cycles for this set of parameters. On the respective cycles, solid dots depict
hourly intervals. The total length of time of the LL cycle is 21.64 h, and that of the DD
cycle is 28.9 h. The 2 cycles effectively overlap along the left branch of . They also intersect near , . Call this point To facilitate the construction of , let us first consider and explain why it has an interval of points on which it is decreasing.
Let be the time it takes a trajectory to evolve along the LL cycle from
to and the time along between these 2 objects. For the case of , and . Define and . Let us consider the trajectory that has initial condition
x1. This trajectory evolves along the LL cycle for until it reaches . At this point, the lights switch off and the trajectory now must follow
the DD cycle for 12 h. The length of time of the DD cycle between and is 22.6 h. Thus, when the lights turn back on, the trajectory is very
far from . In contrast, the trajectory with initial condition travels for hours on DD to . From here it travels 12 h on the LL cycle. The length of time on LL
from to is 14.6 h. Thus, after 12 h on LL, the trajectory is very close to
If denotes the remaining time for each trajectory to reach , it is clear that . Given that is close to , it follows that The continuity of then implies there must be a subinterval of on which is decreasing. Note that the actual interval on which is decreasing may in fact contain rather than be contained in it. Because of periodicity, must also be increasing over a different interval(s). Figure 3B shows for the case . Note that the curve is qualitatively a cubic with a single interval
over which it is decreasing. This interval contains . Since is continuous on a closed interval, it attains its maximum and minimum
values. It is fairly easy to get a bound on these values. The upper bound is attained near
and is obtained by taking the LL cycle from to and the DD cycle from back to . This yields a time of 29.6 h. The lower bound occurs near
and is obtained by taking DD from to , then LL back to , yielding a time of 20.9 h. This yields a bound on on as . Note from Figure
3B that along the decreasing portion, there exists a unique value for which . There is also another value along the increasing portion of for which . Although it is obvious from our simulation results that and exist, analytically proving this fact is beyond the scope of this
paper.
(A) DD (dashed black) and LL (dashed red) limit cycles with open circles indicating
hourly intervals starting at . is an intersection point of these 2 limit cycles. Times along the DD
and LL limit cycles between and are indicated as xD and
xL. The orange lines are located 12 h in time along the
DD and LL limit cycles from the point . The times h and h are numerically computed as the remaining time it takes the
respective trajectories (magenta and blue) to return to . (B) Numerically computed . Solid and open dots depict and . (C) Numerically computed . Solid and open dots depict crossings with the diagonal and
correspond to the stable and unstable phase locked solutions and . (D) Stable (solid) and unstable (dashed) LD limit cycles
corresponding to and . The LL (light gray, smaller amplitude) and DD (light gray, larger
amplitude) solutions are shown for comparison.
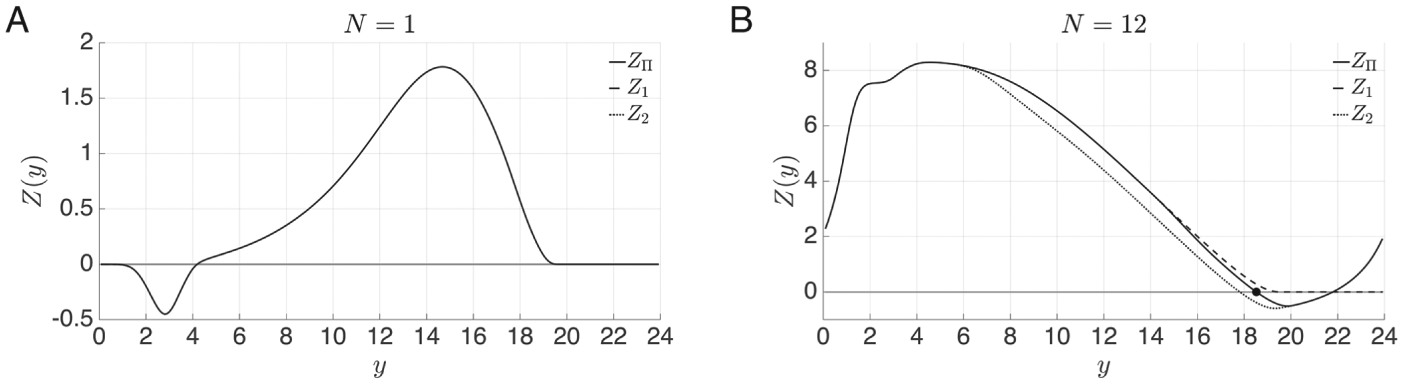
In Figure 3C, we show the graph
of the map . Note that and are fixed points of this map. Further, since , implying that the graph of is increasing wherever it is continuous. It has a discontinuity whenever
passes through a multiple of 24. Finally, note that , implying that is a stable fixed point, whereas , implying that is an unstable fixed point. The stable fixed point corresponds to a stable LD-entrained solution and is shown in Figure 3D. The stable fixed point
occurs at , which means that when the LD-entrained solution starts on
, it initially experiences 1.8 h of light. This is followed by 12 h of
darkness, where the trajectory tracks the DD limit cycle. After this time, the trajectory
is again subject to conditions of light, and it can be seen that the trajectory deviates
from the DD limit cycle. Similarly, corresponds to an unstable LD-entrained solution, which is also shown in
Figure 3D. The value
, so it initially experiences about 3.5 h of darkness when starting on
before then experiencing 12 h of light and then subsequent darkness. In
some sense, for this set of parameters, its trajectory experiences light and darkness at
opposite locations in phase space relative to the solution associated with . Further consequences of the existence of and are explored below when we discuss the dynamics of entrainment.
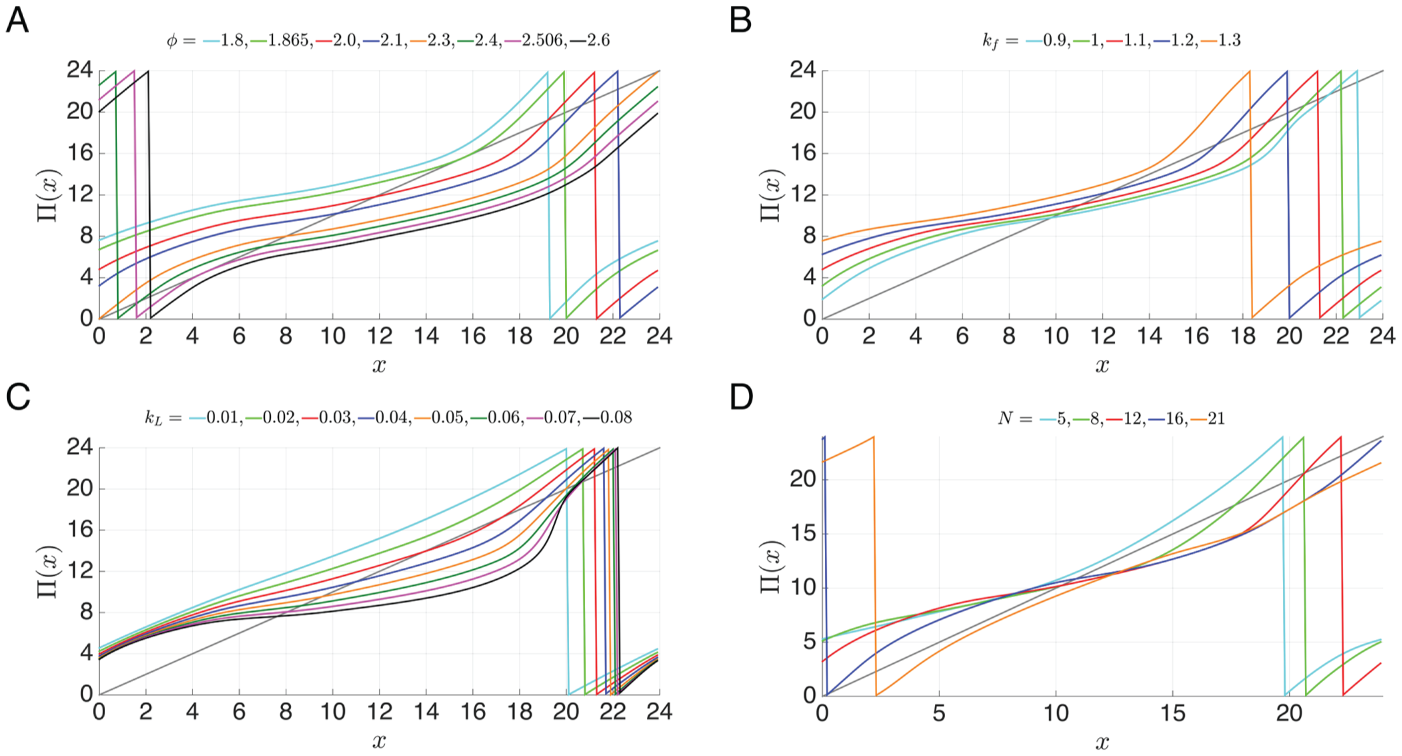
Dependence of on Parameters
To study how the map depends on parameters, we start with the canonical set of parameters
and systematically vary one at a time. We begin with the parameter , which determines the intrinsic period of the oscillator by controlling
the rate of evolution of both the and variables. Larger implies a faster oscillator. If is too large, however, then the intrinsic period of the circadian clock
is too small (much less than 24) and phase-locking with a 24-h forcing will not be
possible. The same conclusion occurs when is too small and the circadian clock is too slow. Thus, there exists an
interval such that stable one-to-one entrainment occurs for lying in that interval. This is a standard result from the forced
oscillator literature; see Glass
(1991) for an example.
Supplementary Figure S1 shows that shifts down when is increased. This makes sense as faster dynamics imply smaller transit
times and so should be smaller. In addition, also shifts to the right, as explained in the Supplementary Material,
Section R1. Figure 4A shows the
map for several different choices of φ. As φ increases, the curves shift down and to the right, the
discontinuity moves to the right, and the stable fixed point moves down and to the left. Biologically, this means that faster
intrinsic oscillators encounter light earlier in the subjective day when entrained than do
slower intrinsic oscillators. In other words, fast intrinsic oscillators have advanced
phases of entrainment (morning larks) and slow intrinsic oscillators have delayed phases
of entrainment (night owls) (Bordyugov
et al., 2015). Note that as increases, the value of increases from around 5 h at to 15 h at . The value corresponds to the saddle node bifurcation where the map lies above and
tangent to the diagonal; corresponds to a second saddle node bifurcation that occurs when the map
lies below and tangent to the diagonal. The interval is called the -range of entrainment. To understand the path through which entrainment
is lost, note that as approaches from above or from below, the fixed points and merge in a saddle-node bifurcation. This merging corresponds to a
saddle-node bifurcation of periodic orbits in phase space, as described in Supplementary
Material, Section R1.
The entrainment map changes systematically as intrinsic (A, B) and extrinsic (C, D)
model parameters are varied. (A, B) Increasing the intrinsic speed of oscillators by
(A) increasing or (B) decreasing moves the stable fixed point of the map to the left and leads to
entrained oscillators that encounter light earlier in the subjective day. Loss of
entrainment occurs when the curves do not intersect the diagonal. (C) Increasing
also moves the stable fixed point of the map to the left,
corresponding to increased light intensity advancing the phase of entrainment. The
increased concavity of as increases suggests that brighter light decreases the amount of time
required to reach stable entrainment. (D) Changes in light duration () have only a minor effect on the location of the stable fixed point
of the map, suggesting that the photoperiod does not significantly affect the phase of
entrainment for these parameter values. However, the fixed points do undergo
saddle-node bifurcation if the light duration is too short or too long, corresponding
to loss of entrainment in extreme photoperiods.
The parameter is associated with positive feedback in the accumulation of protein due
to inhibition of protein degradation (Novak and Tyson, 2008; Tyson
et al., 1999). Increasing moves the -nullcline up in the phase space. However, this effect is much more
pronounced on the left branch of the -nullcline. As a result, the dynamics slow down near the left branch,
more than they speed up away from it, resulting in a net slower speed. This causes
to shift up and to the left and the stable fixed point to move down and to the right (Fig. 4B). Thus, increasing the amount of positive
feedback slows down the intrinsic oscillations (consistent with recent findings in other
circadian models; Ananthasubramaniam
and Herzel, 2014) and leads to entrained clocks encountering light later in the
subjective day. When increases to 1.27, the fixed points and merge at a saddle node bifurcation, indicating that too much positive
feedback can lead to loss of entrainment. As decreases, the fixed point along the middle branch of eventually becomes stable, causing the disappearance of the LL
oscillation at and DD at . Note, however, that for , even though we no longer have self-sustained LL and DD oscillations, it
is still possible for the system to be driven by the forcing to exhibit 24-h rhythms.
Mathematically, this occurs because in this range of , the model reduces to driving a stable fixed point with the 24-h
forcing. The ensuing limit cycle is relatively small in amplitude and lies in a
neighborhood of the intersection of the and nullclines. From a circadian point of view, this is an example of
“masking” rather than entrainment (Mrosovsky, 1999).
Understanding the effect of changes in and is somewhat more complicated than the changes due to and . The reason is that changes in dramatically affect the right branch of , while changes in affect where in phase space the LD-entrained oscillator experiences
conditions of light or dark. As a result, their respective effects on the speed of the
LD-entrained oscillator are less uniform than with the intrinsic parameters.
Increases in light intensity are modeled by increasing . For small values of , entrainment is not possible, since the forced oscillator will still be
too slow. Thus, there is a minimum strength of needed to entrain. Over a range of values, increasing speeds up the oscillations, since in general, the dynamics under lights
are faster than during dark. Figure
4C shows the effect of changing on the map . Because increases in speed up the dynamics, the map shifts down and to the right (although
there is small region near where this is not true for the larger values of ) and the stable fixed point moves to the left. This corresponds to
increased light intensity advancing the phase of entrainment. With our canonical parameter
set, the intrinsic oscillator has a long DD period () and a delayed phase of entrainment (i.e., a night owl). Thus, this
result is consistent with the use of bright light therapy to treat patients with delayed
sleep phase syndrome (Dodson and Zee,
2010). For this figure, we had to lower the Poincaré section to to guarantee that for any initial condition of the map, the ensuing
trajectory crossed in the first cycle of its oscillation. Note that this results in
and for the canonical case. Increased values of cause the linear portion of the equation that determines to dominate. In this case, can become monotone increasing and nearly linear, which produces a
stable fixed point at the intersection of for . Thus, the LL limit cycle will fail to exist. Observe that for
although the LL oscillation fails to exist, the LD-entrained solution
will continue to exist. For this case, during the light portion, the trajectory tracks the
left branch of toward the stable fixed point. This fixed point prohibits the trajectory
from leaving and, as a result, puts an upper bound on how fast the oscillator can
evolve. When the lights turn off, the nullcline switches to , which is cubic shaped, and the trajectory circles the unstable
intersection point of and before returning the left branch of . Thus, for arbitrarily large values of , oscillations persist, such that increasing the intensity of light can
speed up the oscillations only to a certain extent and never too fast to disrupt
entrainment.
Figure 4D shows how the map Π(x)
depends on the photoperiod (LD). Increasing means that the oscillator is exposed to more light. Generally, this does
speed up the dynamics. This fact is reflected by the discontinuity moving to the right as increases, similar to how it moves for all the other parameter
variations that speed up the dynamics. What differs with changes in from other cases is that the map as a whole does not shift down and to
the right. Entrainment continues to disappear through saddle node bifurcations if the LD
forcing is too heavily skewed toward either the light or dark intervals. When
is large, the photoperiod contains too long an interval of light. The
oscillator becomes too fast and loses the ability to entrain in the same way that
entrainment is lost when increases. In particular, the left part of the map shifts down and to the right with increases in just as it does with increases in . The right part of the map shifts up when is decreased and the dark interval dominates. In that case, the
oscillator eventually becomes too slow to entrain, similar to when is decreased. Further details explaining why the map behaves in this way
are provided in the Supplementary Material, Section R1.
Dynamics of Entrainment
Having understood the situations that lead to the existence of the LD-entrained stable
and unstable solutions, let us now turn to understanding some aspects of the transient
dynamics of trajectories that start with initial conditions that lie off of these limit
cycles. The most straightforward way to do this is to use the map to track the transient dynamics associated with an initial condition
. This will allow us to make comparisons to a direct simulation in which
we start an oscillator on the Poincaré section , hours into an LD cycle.
For the map , a cobweb diagram from any initial condition
x0 ≠ xu shows that the trajectory converges to the stable fixed point
. We can readily determine 2 characteristics of entrainment: the time to
entrainment and the direction of entrainment. In the Methods section, we defined a
solution to be entrained if there exists , such that for all , . The integer depends on and refers to the number of iterates needed for the condition to hold
from any given initial condition . We can calculate the time to entrainment in the following way. In the
cobweb diagram, at each iterate, the trajectory hits either the upper or lower branch of
. Let denote the value of the discontinuity of the map. Depending on the
location of the discontinuity , the time associated with that iterate will differ (see the Supplementary
Material, Section R2, for details). The direction of entrainment refers to whether the
cobwebbed iterate moves to the left or the right. When , then the iterate moves to the right, and the phase of the oscillator
with respect to the onset of the lights is larger. Thus, we call this a phase
delay. Alternatively, we obtain a phase advance when the
iterate moves to the left when . Note, however, that transitions between branches follow a different
rule. If , then a transition from the lower branch at one iterate to the upper
branch at the next iterate is considered a phase delay, even though . This is because this type of iterate is equivalent to more than 24 h
passing before the return to . If , then the transition from upper branch to lower branch is considered a
phase advance even though , since this type of iterate implies 2 crossings within one 24-h cycle.
Finally, observe that the unstable fixed point demarcates which initial conditions lie on trajectories that entrain
through phase advancing or phase delaying.
To study a specific example, we return to the map associated with the canonical set of
parameters. Here , , and . Let us choose an initial condition . This point lies to the left of , so under the cobweb dynamics, it will move to the left and converge to
from above, thereby phase advancing (green trajectory of Fig. 5A). Comparing to direct
simulations, we find that the entrainment time is (6.6 days), which matches very well with the predictions of the map
(157.247 h). As another example, choose , which lies to the right of . Now the iterates of the map move to the right, hit the lower branch of
, and then are cobwebbed back to a smaller value of , from where they continue moving right (magenta trajectory in Fig. 5A). In this case, the oscillator
entrains through phase delaying. From the map, we find the entrainment time to be 180.775
h (7.5 days) and from direct simulations to be 180.767 h. In Figure 5B, we show the time traces for solutions with
each of these 2 initial conditions. Note that the green trajectory phase advances, while
the magenta trajectory phase delays, consistent with the predictions of the map. Explicit
values calculated from iterates of the map and direct simulations are shown in Tables S1
and S2 in the Supplementary Material, Section R2.
Entrainment maps systematically reveal entrainment dynamics such as whether
entrainment will occur through phase advances or phase delays. (A) Cobweb diagram
showing convergence to starting from 2 different initial conditions (green trajectory) and (magenta trajectory). The former entrains by phase advancing, the
latter by phase delaying. (B) The approach to the stable cycle shown in the
versus plane. Green and magenta traces correspond to the same values as in
part A.
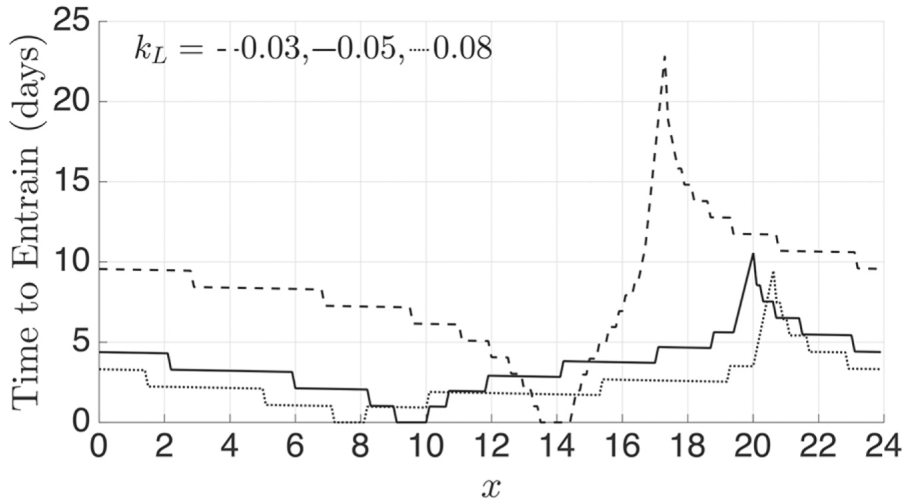
The map allows us to easily calculate entrainment time for any initial condition, for a
range of parameters. For example, to see how light intensity affects entrainment time, we
varied the initial conditions between 0 to 24 for 3 different values of ; see Figure 6
(also see the Supplementary Material, Section R2, which shows how choosing a different
Poincaré section affects entrainment time). The solid curve denotes the canonical case
, while the dashed and dotted curves are for and , respectively. As expected, for each curve, initial conditions that
start in a neighborhood of (local minima of curve) take no time to entrain, while those in a
neighborhood of take the longest. Note that and change as a function of , as can be seen from Figure 4C. Further note that stronger light leads, on average, to the shortest entrainment times, as can be observed
by noting that the dotted curve lies on average below the others. This result can be
predicted based on how the shape of changes with . Observe from Figure
4C that as increases, the map has a region where the slope becomes smaller (roughly
between and ) and a region where the slope and concavity increase (roughly between
and ). It is in the region to that entrainment time dramatically decreases for . The reason is that iterates of the map use this sharp change in slope
to make bigger advances toward the fixed point. This suggests that the amount of time
required for a circadian oscillator to reentrain following a phase shift in the light-dark
cycle, for example, after rapid transmeridian travel, will depend on light intensity with
brighter light resulting in less jetlag.
Time to entrain depends on light intensity and the proximity of initial phase to the
stable and unstable fixed points. Larger values of , on average, lead to faster entrainment. For each case, the minimum
entrainment time of 0 occurs for initial conditions in a neighborhood of
and the maximum in a neighborhood of ( and for , respectively).
Relating to a PRC
The phase response curve of an oscillator measures the change in asymptotic phase
experienced when a perturbation is given to an oscillator along different parts of its
limit cycle. In the context of our problem, we shall construct the PRC using a light pulse
of length hours with . Specifically, start with a DD oscillator on the Poincaré section
. Allow it to evolve for hours along the DD limit cycle and then perturb it by introducing a
light pulse of hours. We define
where is the free running period of the DD oscillator and is the time when the perturbed oscillator crosses for the th time. At each cycle, the quantity measures the deviation of the transit time of the perturbed oscillator
from the DD oscillator. is often called the th-order PRC. Here, phase 0 is chosen as a point on the DD oscillator and
thus the perturbed phase is defined relative to the DD oscillator itself. In general, such
PRCs are typically used when 2 conditions are met: The duration of the light pulse is small, and the magnitude is not too large. These conditions imply that the effect of the light
pulse is felt within the current or subsequent cycle to when it is applied and that the
oscillator returns quickly to its intrinsic limit cycle thereafter, albeit with a
potentially different phase. Both in theory and, as we will show, in practice, these
conditions limit the applicability of using PRCs to determine phase-locking.
The map can be directly compared to provided that both functions have the same domain. First, note that the
domain of is the interval , while the domain for is . Thus, the 2 functions can be compared only when or by truncating the domain of (if ) or (if ). To consider a case of , we set and . In turn, this implies that and , as shown in Figure S3 of the Supplementary Material, Section R3.
Substitute into Equation (6) for to obtain
The subscript on the PRC in this case indicates that this function is obtained from our
map rather than directly through simulation. Additionally, it is convenient
to leave expressed in terms of rather than in terms of so that we can avoid the mod operation.
In Figure 7A, we show the results
of directly computing the PRCs and for and with the map-based PRC . Note that because the effect of a single light pulse wears off
relatively quickly, for . A stable phase-locked solution will occur where the PRC has a root with
negative slope. All 3 PRCs quantitatively agree with one another, although it is difficult
to discern that the PRC is non-zero for small and large values of . In Figure 7B,
we show the results for a case where the duration of the light pulse is not small,
specifically . Note here that , , and differ quantitatively for larger values of . In particular, all 3 have different stable roots. By comparison to
direct simulations, the roots predicted by and are incorrect, whereas the root predicted by the map-based
is correct.
The PRC fails to accurately predict the phase of entrainment for long light pulses,
whereas the entrainment map is accurate for light pulses of any length. (A) For a
short light pulse (), , , and all agree with one another. (B) For a longer light pulse
(), only correctly predicts the value of the root computed by direct
simulation (solid dot at ).
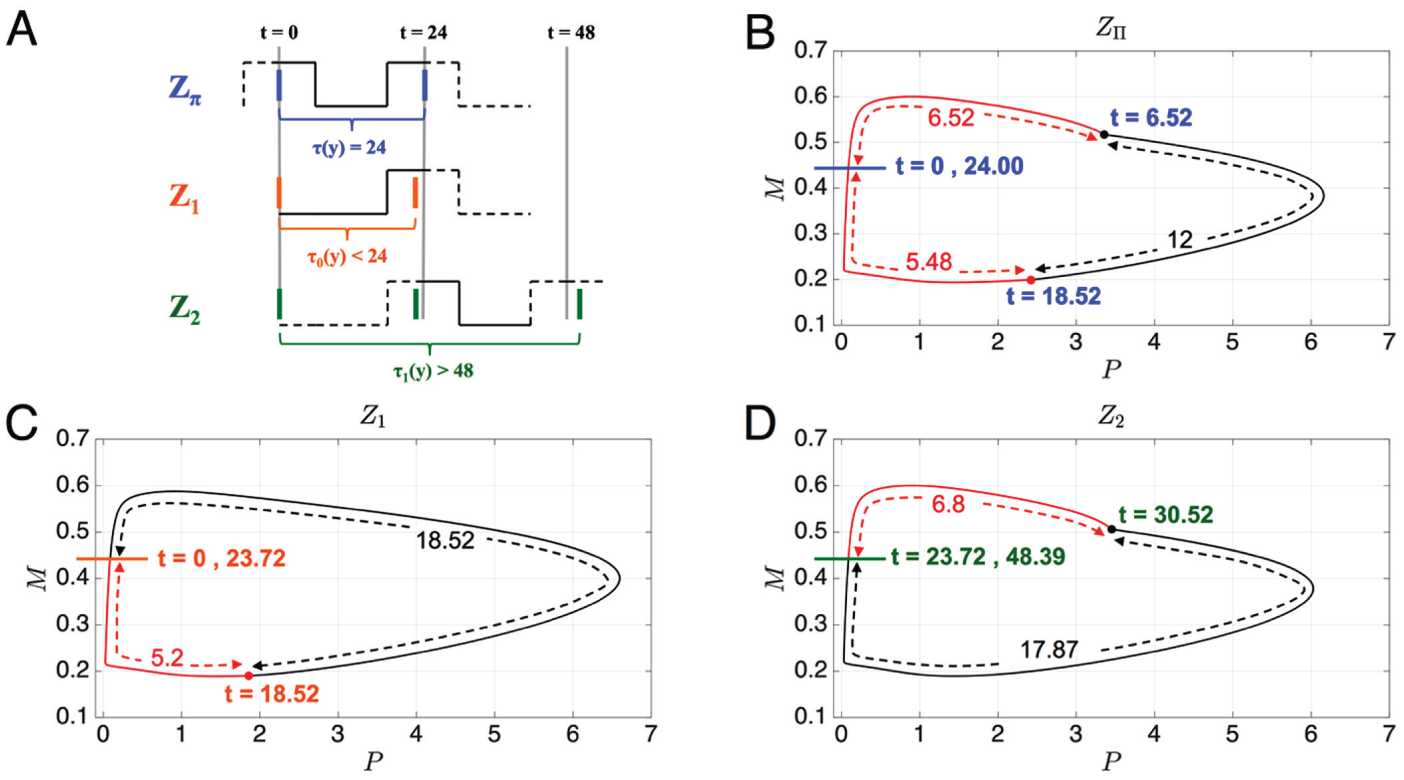
The preceding example of allows us to illustrate the primary difference between the entrainment
map and the PRC, as well as the limitations of the latter. In Figure 8A, we show schematic diagrams
of the case where is chosen as the root of (), which depicts where in time the oscillator is subjected to light.
Figures 8B, 8C, and 8D show trajectories in the phase plane. In Figure 8A, the first solid line of
each schematic denotes and represents the trajectory starting on the Poincaré section
. For the map , is small (since is large), and thus the trajectory starts out with the lights on. In the
phase plane, the trajectory will follow the LL cycle for hours, before then following the DD dynamics for 12 h. After that time,
the trajectory will again be subjected to light during the remaining time it takes to return back to . Thus, when constructing the entrainment map , the light pulse is allowed to be split up over different portions of
the cell’s trajectory, as shown in Figure
8B. In contrast, when constructing the PRCs and , the light pulse is never split up. In the phase space, the trajectory
now traverses along DD for hours before the lights turn on. Thus, in comparison to the
-based trajectory, it is in a very different location in phase space when
the LL dynamics come into play. The first crossing of yields . As can be seen from Figure 8C, the oscillator has received only a small portion of the light pulse
to this point, but it receives the light in a region of phase space that speeds up its
oscillation. The second solid orange line in Figure 8A shows where the oscillator is relative to
the light cycle at the moment it again crosses . The light gray lines show 24-h intervals and represent where the
oscillator would have been in the absence of light input. Thus, . After this point, the trajectory receives the remaining portion of
light shown in Figure 8D but is
actually in a region of phase space where the added light slows it down (lower schematic
of Fig. 8A). In fact, it is slowed
down so much that at the second crossing of , .
Comparison of how the light pulse affects the calculation made by the various PRCs.
(A) Schematic showing where in time the oscillator crosses (solid blue, orange, and green) and where the reference oscillator
does for and (solid gray). The portion of the light pulse that is used in the
calculation is shown in solid in each case. (B-D) Each panel shows where in phase
space the light pulse affects the oscillator. (B) Effect of light pulse is split up in
time, to , and then again from to . (C) The light pulse is only felt at the end of the oscillation from
to . (D) The light pulse is only felt during to 30.52.
When the intrinsic period of the DD oscillator is different than 24 h, then we must
restrict the domain of or to be able to directly compare them. Alternatively, we can simply
compare the predictions regarding the phase of locking made by the map versus those made by either or . We did so over a set of values (from to 8.25, with ) that span a range of intrinsic periods from 23.5 to 35.2. In all cases, when compared to direct simulations,
the predictions made by the map were more accurate than those made via the PRCs; for details, see Table
S3 in the Supplementary Material, Section R3. We also checked the accuracy of the
predicted phase of entrainment for an alternative PRC construction protocol that was based
on detecting peaks of the P variable rather than crossings of a Poincaré
section. Specifically, we perturbed the canonical DD oscillator () with light pulses () and then measured the phase shift of the peak time in the sixth cycle
after the perturbation. The PRC predicted the entrained phase correctly (within 2-min
accuracy) for but was off by more than 30 min for . In contrast, the entrainment map predicted the entrained phase
correctly (within 1 second accuracy) in both cases.
Higher Dimensional Models
We now show that the entrainment map can be defined in higher dimensional settings by
constructing maps for the Gonze and Kim-Forger models. In each of these cases, the
Poincaré section is chosen at a fixed value of a variable that does not receive the
light-dark forcing as described in the Methods section. Note that in addition to being of
different dimensionality, these models also differ in the manner in which light enters the
equations. In the fly model (NT), light increases the rate of protein degradation, whereas
in the mammalian models (Gonze and Kim-Forger), light increases the rate of mRNA
transcription. These distinct effects of light lead to contrasting relationships between
DD and LL dynamics across the models. Specifically, the LL limit cycle has a shorter
period than the corresponding DD limit cycle in the NT model but a longer period than the
corresponding DD limit cycle in the Gonze and Kim-Forger models. Despite these differences
in the behavior of these models under constant dark and constant light conditions, we find
that the entrainment maps for these models share some common features and are able to
accurately capture the effects of light-dark forcing in all 3 cases.
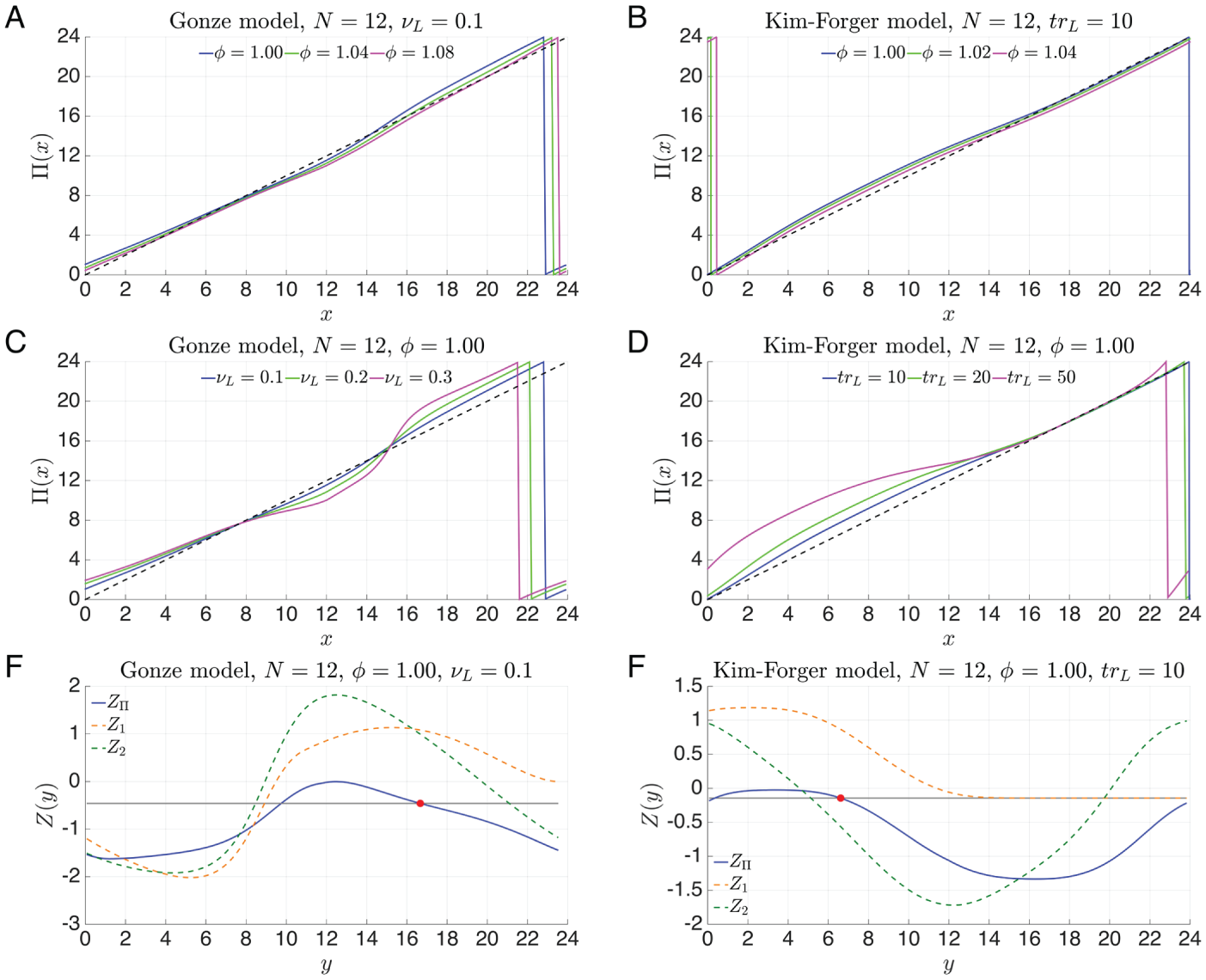
Figures 9A and 9B show the entrainment maps for 3
different values of the intrinsic periods of the respective models. Although it is
difficult to see, note that as the intrinsic oscillator speeds up (increasing
), the maps move the same way as they did in the NT model. Namely, the
entrained phase shifts to an earlier part of the light-dark cycle. The maps for both
models also share with the NT maps the property of having 1 point of discontinuity. This
commonality across the 3 models, together with those discussed below, suggests that the
entrainment map provides a robust method for finding universal properties of circadian
oscillators.
Entrainment maps and PRCs for the Gonze model (left) and Kim-Forger model (right). (A
and B) Variations in the maps due to changes in the intrinsic period of the
oscillators. As with the NT model Figure 4A, the map shifts down and to the right for faster intrinsic
oscillators. (C and D) Stronger light affects entrainment. There is not much
difference in the phase of entrainment but a pronounced difference in the concavity of
the map, which will lead to faster entrainment. (E and F) Comparisons of the PRC
generated from our map, , versus those generated from perturbations of the DD limit cycle.
The red dot is the entrained phase calculated via direct simulation.
In Figures 9C and 9D, we show how the maps behave in
response to increases in light intensity ( in the Gonze model and in the Kim-Forger model). In both models, as light is increased, the
phase of entrainment does not change much. Instead, at various places, the concavity of
the map increases. As these properties were also found in the NT model (Fig. 6), we would predict that
stronger light leads to faster entrainment in both of these models just as it did in the
NT model.
In Figures 9E and 9F, the PRCs , , and are shown for a 12:12 photoperiod. What is interesting about these
graphs is how poorly the conventional PRCs Z1 and Z2 perform in
predicting the entrained phase (red dots). fails to produce an entrained phase for the Gonze model while producing
a set of nonunique solutions for the Kim-Forger model. is quantitatively incorrect for both models. The reason why the
conventional PRC fails is because it is calculated using a perturbation of the DD limit
cycle. In contrast, the entrainment map based PRC, , perturbs off of the LD-entrained solution and produces an accurate
prediction. For the Gonze model, it is possible to see that the DD and LD-entrained
solutions lie in very different regions of phase space (see Suppl. Fig. S5). So the fact
that the PRC and map-based predictions are different is not surprising. For the Kim-Forger
model, it is nearly impossible to tell where in phase space the DD and LD-entrained
solutions lie. But we would infer that they are not near one another.
Discussion
Circadian rhythms enable organisms to appropriately align physiological and behavioral
processes with the 24-h environmental cycles conferred by the earth’s daily rotation.
Rhythms persist when the organism is isolated from external time cues; however, in constant
conditions the period of circadian oscillators is typically not exactly 24 h. For example,
the average human circadian rhythm is 24.2 h while that of mice is 23.5 h (Ripperger et al., 2011). These
oscillators are subject to periodic external forcing such as the light-dark cycle that
adjusts the phase of the rhythm and entrains them to a period of 24 h. The entrained rhythm
must have the proper phasing with respect to external events in order for the circadian
system to confer a selective advantage to the organism (Ben-Shlomo and Kyriacou, 2002). Two important
questions arise: Under what conditions do circadian oscillators phase-lock to external
forcing, and when they do what determines the phase of entrainment?
In this paper, we have developed a new method to determine the phase-locking properties of
a circadian oscillator relative to a 24-h light-dark cycle. We derived an entrainment map
for a 2-dimensional NT model of the Drosophila molecular
circadian clock and then showed that the map can be similarly derived for the 3-dimensional
Gonze and the 180-dimensional Kim-Forger models of the mammalian molecular circadian clock.
In particular, if the oscillator lies on a Poincaré section and is the time from the onset of the most recent light-dark cycle,
is measured when the oscillator first returns to and predicts the time from the beginning of the most recent light-dark
cycle. A fixed point of the map corresponds to a phase-locked LD-entrained solution. In all
models, for canonical sets of parameters, we showed that the map typically has 2 fixed
points, , which is stable, and , which is unstable. Further, since corresponds to where the lights turn on (nominally 0700 h), knowing the
value of allows us to determine where along the trajectory in phase space the
lights turn on and off. For example, in the NT model with the set of parameters that
corresponds to an intrinsic period (canonical parameter set with ), the LD-entrained solution has the lights turn on 14 h before
reaches its maximum value. This is consistent with biological findings
that the highest mRNA levels for Per, an important gene in the
Drosophila circadian clock, are found 2 h after lights off in a 12-h:12-h
light-dark cycle (Young,
1989).
Using the entrainment map allowed us to understand and predict the effects of parameter
changes in the model. Indeed, parameters that have similar effects influence the entrainment
map in the same way across all 3 models. For example, increasing decreases the intrinsic period of the oscillators. In all cases, the
entrainment map shifts down and to the right, moving the stable fixed point
to smaller values. The analysis also revealed several interesting roles
for the unstable fixed point . First, separates the set of initial conditions that entrain via phase advance or phase delay. Second, initial conditions
that are in a neighborhood of took the longest to entrain, largely independent of the direction of
entrainment. Third, loss of entrainment could occur due to changes in parameters through a
stereotypical saddle-node bifurcation when and merge. Some changes in parameters, however, never lead to a loss of
entrainment: in particular, increases in light intensity ( in NT, in Gonze, and in Kim-Forger). Fourth, the existence of implies the existence of an unstable entrained periodic solution. As shown
in Figure 3D, this unstable limit
cycle is smaller in amplitude than its stable counterpart. It is also shifted by a distance
of roughly in terms of when it experiences light relative to the stable limit cycle.
For the canonical set of parameters, this implies that when the unstable limit cycle is
experiencing light, the stable limit cycle is experiencing dark, and vice versa. Thus, the
solutions are effectively 180° out of phase. As changes and the system nears a saddle-node bifurcation, the difference in
phases between the stable and unstable solutions shrinks (eventually going to 0) and
entrainment occurs mostly through phase advance if or phase delay if .
The basic idea behind constructing an entrainment map for any particular model is quite
general. Namely, in the phase space of the model, choose a Poincaré section along a point of
the LD-entrained solution. Then compute the map by taking initial conditions that lie at the intersection of the map and
the LD-entrained solution and measure the return time to the Poincaré section.
is then the amount of time that has passed since the onset of lights in
the most recent light-dark cycle. As such, the entrainment map will always be 1-dimensional.
What will vary is the dimensionality of the Poincaré section . As our simulations have shown, even when the Poincaré section is high
dimensional, as in the Kim-Forger model, the entrainment map very accurately predicts the
stable phase of entrainment. In these high-dimensional cases, the map has the added
advantage that effects of changes in parameters are more easily predicted because these
changes are stereotypical across models.
Comparison to Prior Work
There are 2 major paradigms for entrainment in the circadian field: discrete (or
nonparametric) entrainment and continuous (or parametric) entrainment (Roenneberg et al., 2010a). The
former is based on the idea of a free-running oscillator, DD for example, experiencing
discrete phase shifts in response to perturbative inputs along its limit cycle and in
particular light-dark transitions. This approach has led to methods based on the PRC and
does not explicitly take into account the periodic nature of the forcing signal. The
second paradigm assumes that entrainment occurs through continuous adjustments to the
speed of the oscillator and involves a velocity response curve (VRC) (Beersma et al., 1999; Rand et al., 2004; Taylor et al., 2010, 2014). The VRC characterizes
regions in the circadian phase space where the speed of the oscillator increases or
decreases as a function of a change of parameter—for example, light intensity. It can be
thought of as the variational derivative of the phase with respect to a parameter of
interest. This approach is categorized as being continuous, rather than discrete, because
it assumes that light affects the oscillator throughout the circadian cycle and not just
at light-dark transitions. Both PRC- and VRC-based methods are fundamentally based on
using the isochron structure of the underlying DD oscillator to determine the change in
phase. The phase of these oscillators is defined relative to a reference point on the DD
oscillator itself.
Our approach to studying entrainment is different than these methods in that we view the
periodic 24-h light-dark forcing as being the underlying rhythm that sets the reference
point for determining phase. Namely we assign the time at which lights turn on as the
reference and do not separately define a phase for the DD or LL oscillators. We
determine how a circadian oscillator with a prescribed initial condition relative to
lights-on adjusts its dynamics over a periodic 24-h light-dark cycle. By considering the
24-h range of possible initial conditions, we obtain the entrainment map . Depending on the initial condition, is calculated by splitting up either the light or dark pulse over the
course of 1 or more 24-h cycles. For example, with 12:12 LD forcing, since corresponds to lights-on, the initial condition leads to a computation that starts with the oscillator at
and imposes 8 h of light, followed by 12 h of dark and then 12 h of
light and so on. The value is determined when the trajectory first returns to independent of whether this time took less than, more than, or the same
as 24 h. If, for example, , then initially 5 h of dark would be imposed followed by 12 h of light,
then 12 h of dark, and so on. In the former case, the light pulse would be split up; in
the latter, the dark pulse would be split up. In contrast, PRC- and VRC-based methods only
use a single light pulse, and often after imposing too long a dark duration (because they
perturb off of DD conditions), which can lead to only a portion of the light pulse being
taken into account by the time the oscillator returns to its reference phase.
The entrainment map is similar in spirit to the circadian integrated response
characteristic (CIRC), proposed by Roenneberg et al. (2010b), in that both approaches attempt to take into account
the periodic nature of the light-dark forcing. Common to all approaches is the need for a
well-defined oscillatory solution that can be described as a limit cycle in an appropriate
phase space. In most cases, this is the DD limit cycle. The entrainment map, in addition,
uses the structure of the LL limit cycle to help define its properties. This is closely
related to the suggestion by Peterson
(1980), who noted that an LD-entrained solution at some moments in its cycle
tries to approach the stable DD cycle and at other moments approaches the LL cycle. Our
analysis involving does not explicitly incorporate a VRC. However, it does use the fact
that changes in parameters differentially affect the speed with which a trajectory evolves
in different regions of phase space. For example, we showed how changes to intrinsic
parameters and of the NT model had a more uniform effect on speed compared with those
associated with the intensity and duration of the light pulse.
These points of difference and commonality aside, the entrainment map is simply a tool to
make predictions about the phase-locking properties of a circadian oscillator. As a
result, the findings obtained through its use should qualitatively match prior findings
that use other methods that themselves match to empirical data. In Bordyugov et al. (2015), the authors derive
conditions on entrainment and its dependence on parameters in an abstract setting of the
Kuramoto phase oscillator model (Kuramoto, 1984). They then test the predictions made by the phase model on a
modified Goodwin model of gene regulation with solely negative feedback (i.e., the Gonze
model; Gonze et al., 2005) and
on a 19-dimensional molecular clock model (Relogio et al., 2011). They consider a
2-dimensional parameter space consisting of the mismatch between the free-running period
of the intrinsic oscillator and the period of the forcing and the relative strength of
zeitgeber input to the oscillator amplitude. Their primary finding is that the one-to-one
phase-locked solutions form a wedge-like structure (Arnold tongue) in this space and that
the range of stable phases that exist within this wedge can span as much as 12 h. In
particular, for weak zeitgebers, the 12-h span is achieved over a relatively small span of
mismatches in the periods. As relative zeitgeber strength is increased, a larger range of
period mismatch is needed to achieve the 12-h span of stable phases. This result is based
on some earlier work of Granada and Herzel (Granada et al., 2013) that established the
so-called 180° rule, which is another way to say that the range of stable phases spans 12
out of 24 h. Our results are very similar. For example, in Figure 4A, different values of correspond to different free-running periods of the DD oscillator. Since
we held the forcing period constant at 24, changes in correspond to changing the period mismatch, or equivalently to
considering a horizontal slice of the Arnold tongue in Figure 1A of Bordyugov et al. (2015). We see from Figure 4A that the range of stable
phases spans roughly an 11-h range (xs≈15.8 when
to xs≈4.8 when ). Using the entrainment map, we could also create a 2-parameter Arnold
tongue representation, say for and . To do so, we would combine results from Figures 4A and 4C. The Arnold tongue would also be wedged-shaped in
the space. For fixed , a horizontal slice is obtained by varying between and , as discussed above. For fixed , a vertical slice of the Arnold tongue could be obtained from Figure 4C, which shows that
entrainment is lost when decreases (creating a lower edge of the Arnold tongue) but persists as
increases.
Granada and Herzel (2009)
studied how the time to entrainment is affected by various parameters. One of their main
findings, derived for an abstract Poincaré oscillator and then applied to a Goodwin model,
is that larger relative zeitgeber strengths leads to faster average entrainment. Their
study was specifically designed to minimize the impact of different initial conditions of
the oscillators. In our work, we can use the entrainment map to quickly calculate the
entrainment time for any initial condition for any fixed set of parameters. Figure 6 shows results for different
values of , which is our equivalent to zeitgeber strength. As can be seen from that
figure, increases in do result in an average faster entrainment. This can be concluded by
visual inspection that the area under the curves decreases as increases. Our work additionally suggests that initial conditions that
lie in a neighborhood of the unstable fixed point can take arbitrarily long times to
entrain.
From a more mathematical point view, since is a circle map, our findings are consistent with the vast literature on
forced oscillator models. In the general theory, when a specific parameter is varied, the
forced system transitions through a series of different phase-locked (called
solutions) and quasi-periodic (often called dense orbits) states. The
solutions are actual periodic solutions of the map. These types of
solutions occur when the mismatch between the intrinsic oscillator period and the forcing
period leads to a rational rotation number, roughly defined as the ratio of oscillator
period to forcing period. Dense orbits arise when the rotation number is irrational. These
results are a consequence of Denjoy’s theorem on circle maps and subsequent work of Arnold
and many others (Arnold, 1965;
Glass, 1991; Keener and Glass, 1984). Prior work
shows that under certain assumptions, the rotation number varies continuously with a
parameter of interest and that there exist parameter intervals over which different
solutions can be found. In our case, we focused on one-to-one phase
locking, showing that there is an interval for stable entrainment. We did not exhaustively explore the existence of
dense orbits or other solutions with rational rotation numbers, but we expect those to
exist. Circle maps are also widely used in other contexts such as cardiac dynamics and,
more generally, any process that involves clocking (Winfree, 2001). In such cases, a phase transition
curve (PTC) is used to build a map to predict the effect of periodic forcing. The PTC
measures the normalized phase of resetting as a function of the normalized stimulus phase.
By normalized we mean that phase values lie between 0 and 1. The
associated map at each cycle then measures the new phase as a function of the old. The
PTC, therefore, is constructed using the PRC. Thus, PTCs face the same limitations in
their use that PRCs do and can make predictions that are not borne out by a direct
simulation or experiment; see Glass et
al. (2002).
Future Directions
In this paper we have focused on light-dark forcing since it is the dominant zeitgeber
for circadian clocks. However, the entrainment map can readily be extended to nonphotic
entraining signals, such as temperature cycles, and perhaps give insight into how the
circadian system integrates information from multiple environmental signals occurring
simultaneously. The ease with which time to reentrainment can be calculated and visualized
for different initial conditions by cobwebbing makes the map a useful tool for
investigating jetlag scenarios. The map may also prove to be an effective tool for
studying entrainment and synchronization in networks of coupled oscillators.
Entrainment maps of detailed molecular clock models such as Kim-Forger can be used to
generate hypotheses about how specific mutations or other manipulations of clock
components affect entrainment and to make predictions regarding entrainment mechanisms
that can then be tested experimentally. One interesting question to explore using
entrainment maps is the effect that different proposed mechanisms of transcriptional
regulation (such as protein sequestration and Hill-type repression) have on entrainment
properties (Kim et al., 2014;
Kim, 2016).
Beyond circadian oscillator models, we believe that entrainment maps can also be
successfully constructed based on data from real organisms or experimental preparations.
To illustrate our suggested protocol, we consider the cyanobacterial circadian clock, for
which the phosphorylation status of the protein KaiC is a convenient phase marker both in
vivo and in vitro (Kim et al.,
2015). To construct phase response curves for in vivo cultures, experimentalists
maintain the cultures in constant light and then expose them to 4-h dark pulses at
different phases and measure the resulting phase shift (Kim et al., 2012). To construct an entrainment map,
we would instead maintain the cultures under a 12:12 LD cycle. The minimum of the KaiC
phosphorylation waveform will play a role analogous to the Poincaré section
. Once the culture is entrained, there will be a stable phase
relationship between the onset of lights and the minimum of the KaiC phosphorylation
rhythm. This gives us one data point for the map, namely the stable fixed point
. To obtain other data points, we will measure how the entrained
oscillator responds to phase shifts in the LD cycle. Specifically, in one cycle we will
extend the light or dark duration by a certain number of hours and then record the time of
2 subsequent minima of the KaiC phosphorylation waveform. This will give us an ordered
pair . In fact, each KaiC phosphorylation minimum observed until the culture
reentrains will give us one additional data point for the map. Once the culture has
reentrained, we can repeat the experiment with a different phase shift of the LD cycle to
sample other areas of the map. Alternatively, the experiment can be done in parallel
rather than serially by dividing the cyanobacterial culture into a population of
subcultures and applying different LD phase shifts to each subculture.
Footnotes
Acknowledgements
We thank Horacio Rotstein for helpful discussions regarding forced oscillators and Yong-Ick
Kim for discussions about the cyanobacterial circadian clock. This work was supported in
part by the National Science Foundation DMS-1412877 (C.D.), DMS-1555237 (C.D.) and
DMS-1122291 (A.B.).
Conflict of Interest Statement
The author(s) have no potential conflicts of interest with respect to the research,
authorship, and/or publication of this article.
AnanthasubramaniamBHerzogEHerzelH (2014) Timing of neuropeptide coupling
determines synchrony and entrainment in the mammalian circadian clock.
PLoS Comput Biol10:e1003565.
3.
ArnoldV (1965) Small denominators, I: mappings of the
circumference onto itself. Am Math Soc Transl46:213-284.
4.
BagheriNTaylorSMeekerMPetzoldLDoyleF (2008) Synchrony and entrainment properties of
robust circadian oscillators. J R Soc Interface5:S17-S28.
5.
BeersmaDDaanSHutR (1999) Accuracy of circadian entrainment under
fluctuating light conditions: contributions of phase and period
responses. J Biol Rhythms14:320-329.
6.
Ben-ShlomoRKyriacouB (2002) Circadian rhythm entrainment in flies
and mammals. Cell Biochem Biophys37:141-156.
7.
BordyugovGGranadaAHerzelH (2013a) How coupling determines the
entrainment of circadian clocks. Eur Phys J B82:227-234.
8.
BordyugovGWestermarkPKorencicABernardSHerzelH (2013b) Mathematical modeling in
chronobiology. Handbook Exp Pharmacol217:335-357.
9.
BordyugovGAbrahamUGranadaARosePImkellerKKramerAHerzelH (2015) Tuning the phase of circadian
entrainment. J R Soc Interface12:20150282.
10.
DodsonEZeeP (2010) Therapeutics for circadian rhythm sleep
disorders. Sleep Med Clin5:701-715.
11.
ForgerDPeskinC (2003) A detailed predictive model of the
mammalian circadian clock. Proc Natl Acad Sci U S A100:14806-14811.
12.
ForgerDGonzeDVirshupDWelshD (2007) Beyond intuitive modeling: combining
biophysical models with innovative experiments to move the circadian clock field
forward. J Biol Rhythms22:200-210.
13.
GlassL (1991) Cardiac rhythms and circle-maps—a
classical problem. Chaos1:13-19.
14.
GlassLNagaiYHallKTalajicMNattelS (2002) Predicting the entrainment of reentrant
cardiac waves using phase resetting curves. Phys Rev E65:021908.
15.
GolombekDRosensteinR (2010) Physiology of circadian
entrainment. Physiol Rev90:1063-1102.
16.
GonzeD (2011) Modeling circadian clocks: from
equations to oscillations. Cent Eur J Biol6:699-711.
17.
GonzeDBernardSWaltermannCKramerAHerzelH (2005) Spontaneous synchronization of coupled
circadian oscillators. Biophys J89:120-129.
18.
GoodwinB (1965) Oscillatory behavior in enzymatic
control processes. Adv Enzyme Regul3:425-428.
19.
GranadaAHerzelH (2009) How to achieve fast entrainment? The
timescale to synchronization. PLoS One4:e7057.
20.
GranadaABordyugovGKramerAHerzelH (2013) Human chronotypes from a theoretical
perspective. PLoS One8:e59464.
21.
GuCLiuZSchwartzWIndicP (2012) Photic desynchronization of two
subgroups of circadian oscillators in a network model of the suprachiasmatic nucleus
with dispersed coupling strengths. PLoS One7:e36900.
22.
GuCXuJLiuZRohlingJ (2013) Entrainment range of nonidentical
circadian oscillators by a light-dark cycle. Phys Rev E Stat
Nonlin Soft Matter Phys88:022702.
23.
HirschMSmaleSDevaneyR (2013) Differential Equations, Dynamical Systems, and
an Introduction to Chaos. Oxford, UK:
Elsevier.
24.
IndicPGurdzielKKronauerRKlermanE (2006) Development of a two-dimension manifold
to represent high dimension mathematical models of the intracellular mammalian circadian
clock. J Biol Rhythms21:222-232.
25.
JohnsonC (1999) Forty years of PRCs—what have we
learned?Chronobiol Int16:711-743.
26.
JohnsonCElliotJFosterR (2003) Entrainment of circadian
programs. Chronobiol Int20:741-774.
27.
KeenerJGlassL (1984) Global bifurcation of a periodically
forced nonlinear oscillator. J Math Biol21:175-190.
28.
KimJ (2016) Protein sequestration versus Hill-type
repression in circadian clock models. IET Syst Biol10:125-135.
29.
KimJForgerD (2012) A mechanism for robust circadian
timekeeping via stoichiometric balance. Mol Syst Biol8:630.
30.
KimJKilpatrickZBennettMJosicK (2014) Molecular mechanisms that regulation
the coupled period of the mammalian circadian clock. Biophys
J106:2071-2081.
31.
KimYVinyardDAnanyevGDismukesGGoldenS (2012) Oxidized quinones signal onset of
darkness directly to the cyanobacterial circadian oscillator.
Proc Natl Acad Sci U S A109:17765-17769.
32.
KimYBoydJEspinosaJGoldenS (2015) Detecting KaiC phosphorylation rhythms
of the cyanobacterial circadian oscillator in vitro and in vivo.
Methods Enzymol551:153-173.
33.
KuramotoY (1984) Chemical Oscillations, Waves, and
Turbulence. Heidelberg, Germany:
Springer.
34.
LeiseTSiegelmannH (2006) Dynamics of a multistage circadian
system. J Biol Rhythms21:314-323.
35.
LeloupJGoldbeterA (2013) Critical phase shifts slow down
circadian clock recovery: implications for jet lag. J Theor
Biol333:47-57.
36.
MrosovskyN (1999) Masking: history, definitions, and
measurement. Chronobiol Int16:415-429.
RelogioAWestermarkPWallachTSchellenbergKKramerAHerzelH (2011) Tuning the mammalian circadian clock:
robust synergy of two loops. PLoS Comput Biol7:e1002309.
43.
RippergerJJuddCAlbrechtU (2011) The daily rhythm of
mice. FEBS Lett585:1384-1392.
44.
RoennebergTDaanSMerrowM (2003) The art of entrainment.
J Biol Rhythms18:183-194.
RoennebergTRemiJMerrowM (2010b) Modeling a circadian
surface. J Biol Rhythms25:340-349.
48.
SerkhKForgerD (2014) Optimal schedules of light exposure for
rapidly correcting circadian misalignment. PLoS Comput
Biol10:e1003523.
49.
StrogatzS (1994) Nonlinear Dynamics and Chaos: With
Applications to Physics, Biology, Chemistry, and Engineering.
Reading, MA:
Perseus.
50.
TaylorSWebbASmithKPetzoldLDoyleF (2010) Velocity response curves support the
role of continuous entrainment in circadian clocks. J Biol
Rhythms25:138-149.
51.
TaylorSCheeverAHarmonS (2014) Velocity response curves demonstrate
the complexity of modeling entrainable clocks. J Theor
Biol363:307-317.
52.
TysonJHongCThronCNovakB (1999) A simple model of circadian rhythms
based on dimerization and proteolysis of per and tim. Biophys
J77:2411-2417.
53.
WinfreeA (2001) The Geometry of Biological Time.
New York: Springer Science & Business
Media.
54.
WollerAGonzeDErneuxT (2014) The Goodwin model revisited: Hopf
bifurcation, limit-cycle, and periodic entrainment. Phys
Biol11:045002.
55.
WuC.Rul’kovN (1993) Studying chaos via 1-d maps: a
tutorial. IEEE Trans Circuits Syst I Fundam Theory Appl40:707-721.
56.
YoungM (1989) The molecular control of circadian
behavioral rhythms and their entrainment in drosophila. Annu Rev
Biochem67:135-152.
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.