
Abstract
Temporally coordinated resorption and synthesis is the key to maintaining healthy bones. Articular cartilage is a highly specialized connective tissue within the joints that lines the surface of a long bone. Emerging evidence has suggested a critical role of the circadian system in controlling cartilage and bone biology. Articular cartilage is sparsely populated with chondrocytes, surrounded by abundant extracellular matrices that are synthesized and maintained solely by chondrocytes. Once damaged, the articular cartilage tissue has poor capacity for endogenous repair, leaving the joints prone to osteoarthritis, an age-related painful condition that affects millions of individuals worldwide. An important question is how articular cartilage has evolved its remarkable capacity to maintain homeostasis and withstand daily biomechanical challenges associated with resting/activity cycles. Equally important is how this avascular and aneural tissue senses time and uses this information to coordinate daily phases of metabolic activity and tissue remodeling/repair. Bone tissue derived from cartilage has similarly sparse populations of resident cells living in dense and largely mineralized matrices. We discuss recent progress on circadian clocks in these matrix-rich skeletal tissues and highlight avenues for future research.
Cartilage and Bone
Articular cartilage is a highly specialized connective tissue. It consists of a dense extracellular matrix (ECM) with sparsely populated chondrocytes, the only cell type in this tissue (Hardingham et al., 2002). Articular cartilage provides a smooth and lubricated surface for joint movement, and its function is dependent on proteoglycans (e.g., aggrecan) and collagens (mainly collagen type II) in the ECM (Asanbaeva et al., 2008). The biomechanical environment surrounding a chondrocyte in the articular cartilage is particularly harsh: high osmolarity, low oxygen, no innervation, no blood vessels, and repeated compressive loading and strain. Chondrocytes live in isolation and rarely divide throughout adult life. Once damaged, this tissue is very difficult to repair (Poole et al., 1987). Excessive degradation of the ECM in articular cartilage is a common feature of 2 common musculoskeletal morbidities, osteoarthritis (OA, associated with old age) and rheumatoid arthritis (RA, an autoimmune condition) (Sophia Fox et al., 2009; Kumar et al., 2016). The role of circadian rhythms and RA has been extensively reviewed (Spies et al., 2014; Buttgereit et al., 2015). Therefore, we focus on links between circadian clocks and OA.
Long bones derived from cartilage are essential for weight bearing, supporting skeletal muscles, and regulating mineral balance (Feng and McDonald, 2011). Longitudinal long bone growth depends on the expansion and endochondral ossification of the growth plate. Of note, another type of bone formation, such as calvarial bone or bone fracture, is through intramembranous ossification, which does not involve cartilage. The growth plate is a cartilaginous tissue found in children and adolescents and replaced by bone in adult humans (Melrose et al., 2016). Bone homoeostasis depends on 2 opposing processes, resorption and synthesis. Normal bone remodeling requires strict temporal control of these 2 processes, regulated by osteoclasts and osteoblasts, to guarantee maintenance of the bone mass and structural quality (Feng and McDonald, 2011). The third cell type (and the most abundant in mature bone, >95% of all bone cells) is the highly mechano-sensitive osteocytes, which play critical signaling roles by communicating with osteoblasts and osteoclasts. In general, osteoclasts rapidly absorb the old mineralized bone, and then osteoblasts are able to slowly form the new bone (Sommerfeldt and Rubin, 2001; Downey and Siegel, 2006). One common feature of chondrocytes and bone cells is scarce cell numbers and abundant ECM. Therefore, it is critical to understand how these highly specialized cells integrate temporal, spatial, and biomechanical cues to regulate their tissue homeostasis. One such cue is the circadian timing mechanism.
The Circadian System
Daily 24-h rhythms in physiology and behavior are generated by biochemical time pieces, known as circadian clocks (Reppert and Weaver, 2002). The circadian system (Figure 1) is orchestrated by a master clock in the suprachiasmatic nucleus (SCN) of the hypothalamus, which in turn controls secondary self-sustained autonomous circadian clocks present in most peripheral tissues (Yoo et al., 2004). The master clock is entrained by the light/dark cycle. The SCN uses this information to convey rhythmic time cues to other brain regions influencing locomotor activity, body temperature, hormone secretion, and feeding behavior (Cermakian and Sassone-Corsi, 2000; Gamble et al., 2014), as well as to different body organs. Peripheral clocks contain identical clock genes/proteins as the SCN and can be synchronized by external time cues other than the light/dark cycle (e.g., feeding/fasting, temperature cycles, and physical exercise). Individual organs integrate intrinsic metabolic and biomechanical cues, as well as extrinsic light/dark cycle signals, to generate and maintain robust cellular rhythms (Reppert and Weaver, 2002; Roenneberg and Merrow, 2005; Takahashi et al., 2008; Dibner et al., 2010).
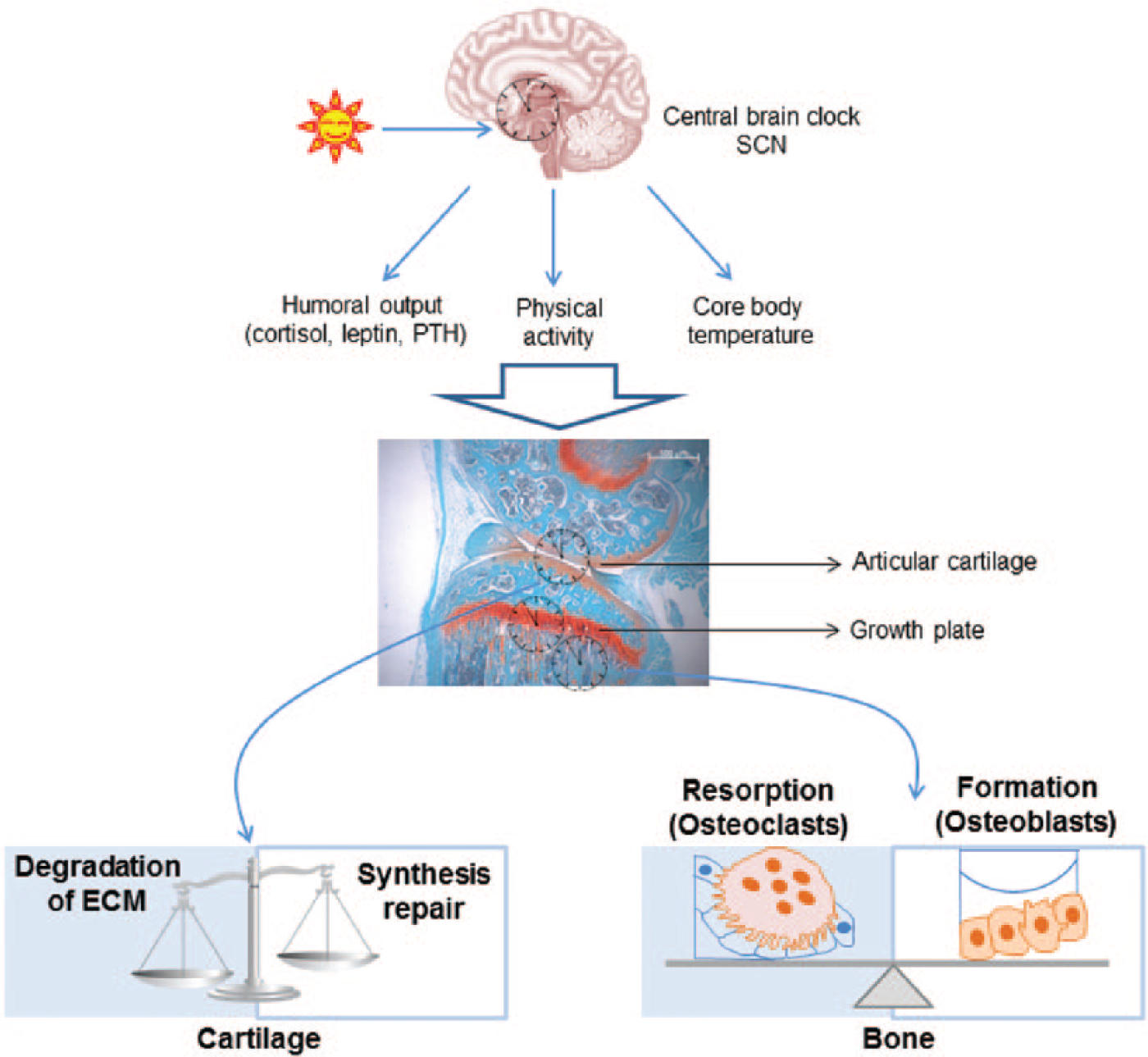
The circadian timing system temporally controls tissue homeostasis in cartilage and bone. The master clock in the suprachiasmatic nucleus (SCN) receives light and other environmental zeitgebers (time cues) to entrain the internal timing system. The SCN then generates rhythmic signals (e.g., daily surges of hormones, physical activity/rest cycles, and body temperature fluctuations) to synchronize peripheral oscillators, including those in the skeletal system (such as cartilage and bone). Through rhythmic control of various biological pathways, the circadian clocks temporally coordinate tissue metabolism, homeostasis, and integrity. During aging or in diseases, loss of temporal segregation of opposing events in skeletal tissues, such as extracellular matrix (ECM) degradation/repair in cartilage or bone resorption/synthesis, would disrupt the fine balance between catabolism and anabolism, predisposing the tissues to pathological changes. PTH, parathyroid hormone.
At the molecular level, the mammalian circadian oscillator comprises a transcriptional-translational feedback loop. There are nearly a dozen core clock genes identified in mammals: Period (PER) family, Cryptochrome (CRY) family, BMAL1 (brain and muscle ARNT-like 1/Arntl), Clock (Circadian locomotor output cycles kaput), NPAS2, DEC1/DEC2 (Bhlhb2/Bhlhb3), REV-ERBα/β (NR1D1/ NR1D2; nuclear receptor subfamily), RORα/RORγ (retinoid acid receptor [RAR]–related orphan receptor), DBP/TEF/HLF, and E4BP4 (NFIL3) (Reppert and Weaver, 2001; Schroeder and Colwell, 2013). Most of these clock factors are transcription factors with relatively short half-lives, allowing periodic rise and fall of their levels within a 24-h day (Roenneberg and Merrow, 2005; Takahashi et al., 2008). In the morning, transcription factors Clock and BMAL1 (the “positive arm”) stimulate the expression of Per and Cry by binding to the cis-regulatory element E-box within their promoters (Ko and Takahashi, 2006). Subsequently, the PER and CRY proteins (known as the “negative arm”) bind to Clock and BMAL1 and inhibit their transcriptional activation. This eventually reduces the expression of Per and Cry, forming a negative feedback loop. Furthermore, there is an auxiliary stabilizing loop, which consists of several modulatory nuclear hormone receptors, including REV-ERBs and RORs (Schroeder and Colwell, 2013). In addition, clock transcription factors also regulate the expression of output genes, known as clock-controlled genes (CCGs). These CCGs play key roles in driving ~24-hr rhythms in diverse biological processes (e.g., cell signaling, kinase, and electrical activity), as well as adenosine triphosphate (ATP) production (Reppert and Weaver, 2002; Schibler, 2007).
Tissue-Specific Clock Functions
Interestingly, although the basic mechanism of the molecular clock is very similar among tissues, there is only a small overlap of rhythmic genes between any 2 given tissues. Comparing the circadian transcriptomes in different tissues, it was found that out of 9995 genes that showed circadian oscillations in one tissue or another, only 41 genes were rhythmic in at least 8 of 14 mouse tissues analyzed (Yan et al., 2008). This concept was reinforced by the fact that even within the skeletal system (e.g., cartilage, tendon, and skeletal muscle), the rhythmic transcriptomes differ vastly (Yeung et al., 2014). Indeed, many of the CCGs are tissue-specific transcription factors (such as MyoD in muscle and KLF15 in heart) and characteristic structural proteins, or fundamental metabolic pathways in a particular tissue (Zvonic et al., 2006; McCarthy et al., 2007; Zvonic et al., 2007; Andrews et al., 2010; Jeyaraj et al., 2012; Gossan et al., 2013). In view of this, it is probably not surprising that diverse pathologies have been reported in mice carrying mutations of clock genes. For example, global Bmal1 knockout (KO) mice show a premature aging phenotype in multiple tissues, including cataract, sarcopenia, arthropathy, and so on. Similarly, homozygous Clock Δ19 mutant mice are hyperphagic and obese with attenuated energy balance, and they develop metabolic syndrome over time (Turek et al., 2005). Tendon is an essential component of the musculoskeletal system that provides transmission of forces and an attachment point of muscle to bone. The Clock Δ19 mice and the Clock/Npas2 double KO mice also show spontaneously calcifying tendons (Yu, 2011; Yeung et al., 2014). The concept of tissue-specific clock function is further supported by conditional ablation of the core clock genes in peripheral tissues. Mice with conditional deletion of Bmal1 in pancreatic islets developed a diabetes mellitus–like disorder, with impaired insulin release and glucose tolerance (Marcheva et al., 2010). Disruption of the circadian clock in the liver, adipocytes, skin, and heart leads to hypoglycemia and increased glucose clearance, obesity, epidermal aging, and increased sensitivity to hypertrophy, highlighting the importance of local clocks in tissue physiology and diseases (Lamia et al., 2008; Durgan et al., 2011; Janich et al., 2011; Paschos et al., 2012).
Clocks in Cartilage
Evidence of Circadian Rhythms in Cartilage
Circadian rhythms in cartilage metabolism have been known for decades. Since the 1980s, there have been several rat models using growth plate cartilage to study the hypothesis of day/night changes in cartilage homeostasis. Measuring chondrocyte proliferation markers identified that the most active phase of proliferation was the early morning, which induced the growth plate expansion that peaked at noon (Stevenson et al., 1990). Interestingly, collagen matrix is also mainly synthesized at noon in growing rat cartilage, as revealed by incorporation of [3H] proline (Igarashi et al., 2013). Serum levels of several biomarkers related to cartilage metabolism show day/night variations, including aggrecan, type II collagen, cartilage oligomeric matrix protein (COMP), hyaluronic acid, keratin sulfate, and transforming growth factor (TGF)–β (Kong et al., 2006; Andersson et al., 2006).
The circadian rhythms in cartilage in vivo could be either system driven, as has been shown in other tissues (Kornmann et al., 2007), or driven by local molecular clocks within chondrocytes. In contrast to growth plate, studies of circadian clocks in articular cartilage have only started in the past few years. This has been greatly facilitated by the development of new techniques, such as the real-time bioluminescence photon counting/imaging system (Yamazaki and Takahashi, 2005) and the generation of transgenic mouse models that harbor a short half-life luciferase (Yoo et al., 2004). By imaging the femoral bone of a PER2::Luciferase (PER2::Luc) gene reporter mouse, strong circadian oscillations of PER2::Luc activities were observed from both the growth plate and femoral head cartilage, which maintained for several months (Okubo et al., 2013). Similar approaches have been used to monitor clock gene activities from isolated mouse femoral head, which revealed robust circadian oscillations in the articular cartilage of this tissue (Dudek et al., 2016). Importantly, these in vitro approaches highlight tissue-autonomous rhythms in cartilage. Interestingly, targeted genetic deletion of the core clock gene Bmal1 from chondrocytes led to a complete abolition of the cartilage circadian rhythm, supporting a key role of the molecular clock machinery in driving this local clock (Dudek et al., 2016). Time course quantitative polymerase chain reaction (qPCR) studies have confirmed rhythmic expression of most core clock genes in different types of cartilages, including femoral head articular cartilage, xiphoid cartilage, rib cartilage, and growth plate (Dudek et al., 2016; Gossan et al., 2013; Honda et al., 2013). These studies have also been expanded to human articular cartilage. For instance, by immunohistochemistry, the presence of clock proteins in the articular cartilage of knee joints was demonstrated. Furthermore, using lentiviral delivery of clock reporters, cell-autonomous circadian rhythms in gene expression have been demonstrated in a human chondrocyte-derived cell line (Gossan et al., 2013; Dudek et al., 2016).
Clock Targets in Cartilage
Cartilage homeostasis is regulated by many key transcription factors and signaling pathways. Using cartilage samples collected from mice kept in constant darkness, time-series microarrays were undertaken to reveal the extent of circadian control in this tissue. This approach demonstrated that the expression of 4% of genes was circadian regulated in mouse xiphoid cartilage (Gossan et al., 2013). This number is likely an underestimate in articular cartilage in mice (Yang et al., unpublished RNA sequencing results in femoral head cartilage). Many of these rhythmic genes are involved in metabolic pathways, ECM remodeling (e.g., Timp4), apoptosis (e.g., X-linked inhibitor of apoptosis, Xiap), and catabolic extracellular proteases (e.g., ADAMTSs and matrix metalloproteinases [MMPs]) (Gossan et al., 2013; Dudek and Meng, 2014). ADAMTS4 is one of the main aggrecanases responsible for aggrecan degradation in articular cartilage (Verma and Dalal, 2011). MMP14 (aka MT1-MMP) is a membrane-bound collagenase that regulates the pericellular ECM. MMP14 is also involved in the regulation of chondrogenic differentiation and the activation of other MMP proteases, including MMP13, the main collagenase involved in OA (Knauper et al., 1996). MMP14 KO mice demonstrate widespread skeletal pathologies, including articular cartilage destruction (Holmbeck et al., 1999; Knauper et al., 2002; Szabova et al., 2009; Chellaiah and Ma, 2013). Interestingly, the rhythmic pattern of Timp4, an inhibitor of MMPs, is in complete antiphase to MMP14, indicating temporal separation of the activities of these 2 opposing enzymes. Honda et al. (2013) also performed time-series partial-genome arrays in both permanent and growth plate cartilage from rats. Although a different statistical model was adopted to identify circadian regulated genes, significant numbers of ECM-related rhythmic genes were found to be common in rat and mouse cartilage (Table 1). Importantly, anabolic ECM genes such as fibrillins, laminins, and netrin peaked in the early morning. Together, these studies demonstrate a circadian control mechanism for the anabolic and catabolic pathways in cartilage tissues. These data strongly support a hypothesis that the circadian rhythm in cartilage temporally segregates the activities of different ECM-related molecules to different (optimal) times of the day, and thus may provide an efficient mechanism by which cartilage can recuperate following bouts of nocturnal activity (Figure 1).
Rhythmic ECM-Related Genes in Cartilage and Bone.
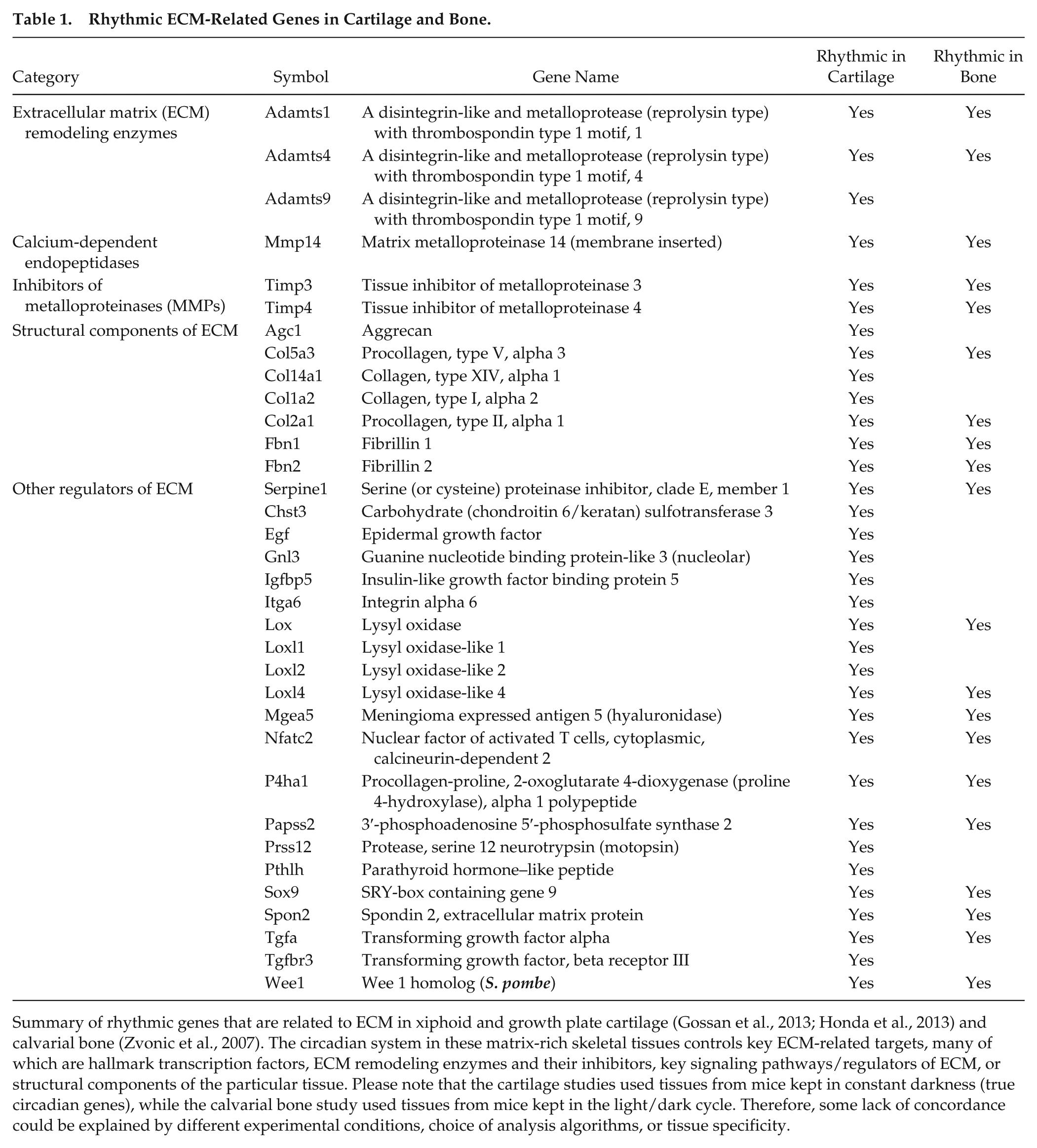
Summary of rhythmic genes that are related to ECM in xiphoid and growth plate cartilage (Gossan et al., 2013; Honda et al., 2013) and calvarial bone (Zvonic et al., 2007). The circadian system in these matrix-rich skeletal tissues controls key ECM-related targets, many of which are hallmark transcription factors, ECM remodeling enzymes and their inhibitors, key signaling pathways/regulators of ECM, or structural components of the particular tissue. Please note that the cartilage studies used tissues from mice kept in constant darkness (true circadian genes), while the calvarial bone study used tissues from mice kept in the light/dark cycle. Therefore, some lack of concordance could be explained by different experimental conditions, choice of analysis algorithms, or tissue specificity.
Interestingly, the chondrocyte-selective deletion of Bmal1 in mice led to constant expression levels of most rhythmic genes identified in wild-type (WT) mice, indicating locally expressed BMAL1 as a crucial regulator of circadian gene expression in vivo (Dudek et al., 2016). The global changes of gene expression patterns in the Bmal1 KO cartilage indicate an overall shift toward a more catabolic state of the chondrocytes. Two key pathways relevant to cartilage homeostasis, TGF-β and nuclear factor of activated T cells, cytoplasmic 2 (NFATc2), were shown to be dysregulated (Dudek et al., 2016). TGF-β signaling is essential for the maintenance of cartilage metabolism and structural integrity, and its dysregulation triggers a catabolic response in arthritic and aged chondrocytes (Finnson et al., 2008; van der Kraan et al., 2009; Bauge et al., 2013). Meanwhile, NFATc2 is a key chondrocyte transcription factor, and its deletion results in OA-like cartilage degeneration, typified by decreased anabolic signaling and increased inflammatory and catabolic pathways (Wang et al., 2009; Rodova et al., 2011; Greenblatt et al., 2013). Loss of BMAL1 could affect cartilage health in multiple ways, given the changes of diverse pathways in Bmal1 KO cartilage. Therefore, further studies are clearly needed to explore additional pathways that link circadian disruptions to cartilage metabolism.
Entrainment Factors for the Cartilage Clock
One key feature of the molecular circadian clock is that it can be entrained by various time cues. Unlike the central clock in the SCN, which receives light input from the retina through the retinohypothalamic tract, most peripheral clocks are regulated through rhythmic signals that are under the control of the SCN, such as temperature and hormones (e.g., dexamethasone) (Gossan et al., 2013; Dudek and Meng, 2014). As a way of adaptation, different body organs have evolved to be differentially sensitive to various time cues. For instance, circadian clocks in the lung and fibroblasts can be regulated by temperature cycles that approximate body temperature changes. In contrast, the SCN seems to be protected from daily body temperature variations (Buhr et al., 2010). Daily feeding signals strongly entrain peripheral clocks in certain tissues, such as the liver and colon (Damiola et al., 2000; Sládek et al., 2007). Cartilage tissues are avascular and not innervated; therefore, it is of particular interest to understand how this local clock communicates with the rest of the body to maintain coherent circadian rhythms. One candidate pathway is glucocorticoid signaling, which shows a daily rhythm controlled by the SCN and has been shown to reset cartilage clocks (Gossan et al., 2013; Okubo et al., 2013). Studies using cartilage explant cultured under 12-h/12-h temperature cycles suggest that the core body temperature rhythm could also serve as a potential synchronizing factor for cartilage clocks. It is worth noting that the activity of the hypothalamic-pituitary-thyroid axis, including the expression of parathyroid hormone (PTH), has long been known to show daily variations (Greenspan et al., 1997; Dudek and Meng, 2014). PTH regulates cAMP/protein kinase A (PKA)/CREB signaling and stimulates Per1 expression in both osteoblasts and growth plate chondrocytes, although its role in articular cartilage remains unknown (Hinoi et al., 2006; Hanyu et al., 2011). In addition, the daily rhythm of activity/rest could also be an endogenous entrainment factor for the cartilage clock. This hypothesis is supported by results showing that the expression of the Clock gene was mechanosensitive (Kanbe et al., 2006). Further studies are clearly needed to test how mechanical loading may affect circadian pacemaking, at least in the skeletal system.
Dysregulation–Diseases
Disruptions to circadian rhythms (e.g., during aging or in shift workers) have been linked to various human diseases, including obesity, diabetes, cardiovascular diseases, and even cancer (Evans and Davidson, 2013; Henriksson and Lamia, 2015; Martino and Young, 2015). In cartilage from aged mice, the amplitude of circadian rhythm has been shown to decrease by ~40%, which could contribute to age-related disease onset in this tissue. Over the past few years, evidence is emerging that environmental or genetic clock disruptions may also predispose cartilage to pathologies such as OA, one of the most prevalent joint diseases with no effective disease-modifying treatment available. There are multiple risk factors for OA, including aging, obesity, trauma to the joint tissue, and chronic inflammation in the joint cavity. Indeed, altered expression of several key clock genes has been found in a mechanical injury model of OA in mouse (by destabilization of the medial meniscus; Gossan et al., 2013). Interleukin 1 (IL-1), a critical cytokine for MMP-dependent cartilage degradation, disrupts circadian rhythms in mouse cartilage explants and chondrocytes in a nuclear factor κB (NFκB)–dependent manner (Guo et al., 2015). It was reported that long-term (22 weeks) environmental disruption to light/dark cycles in mice, a situation that mimics many years of rotating shift work or chronic jet lag, predisposed the mouse knees to OA-like damages (Kc et al., 2015). Moreover, a progressive arthropathy phenotype has been reported in global Bmal1 KO mice (Bunger et al., 2005). This phenotype is age dependent and highly localized (in calcaneal tendon and costosternal cartilage) and requires absence of Bmal1 during development (Yang et al., 2016). Therefore, whether this pathology is due to circadian dysfunction is still unclear. Moreover, one caveat of such systemic rhythm disruption paradigms (either genetic or environmental) is the global dysregulation of circadian rhythms in locomotion, body temperature, or hormones, which may confound the interpretation of any phenotype observed (Yu and Weaver, 2011). One way to overcome these confounding factors is to use a conditional KO model by removing the local clock selectively in chondrocytes. Using this gene targeting approach, severe age-related degeneration and lesions were observed in the articular cartilage of the knee joints (Dudek et al., 2016). These studies prove an essential role of the core clock factor BMAL1 in cartilage homeostasis. At this stage, we cannot exclude the possibility that BMAL1 may also be involved in the regulation of nonrhythmic genes. However, the fact that loss of chondrocyte-BMAL1 leads to global dysregulation of the patterns of rhythmic genes strongly supports a role of circadian rhythms in the homeostasis of articular cartilage (Dudek et al., 2016). It has been reported that the levels of BMAL1 and the number of BMAL1-positive chondrocytes in human osteoarthritic cartilage are significantly lower than nonarthritic cartilage, indicating a role of clock mechanisms in human OA (Dudek et al., 2016).
Clocks in Bone
Evidence of Clocks in Bone Metabolism and Physiology
Bone homoeostasis depends on 2 opposing processes, resorption and synthesis, which requires strict temporal control to maintain the bone mass and structural quality (Feng and McDonald, 2011). A circadian rhythm in bone formation was demonstrated by measuring radiolabeled proline and galactose. Synthesis and secretion of ECM components by hypertrophic chondrocytes and osteoblasts exhibited a ~24-h rhythm in growing rats, starting in the early morning (resting phase) and peaking at around 1 p.m. (Igarashi et al., 2013). By measuring calcium and phosphorus fractions, it was shown that cartilage mineralization in rat tibial growth plate occured at night during the activity phase (Russell et al., 1984). Daily variations of serum markers linked to bone metabolism, such as calcitonin, calcium, C-telopeptide, osteocalcin, parathyroid hormone, skeletal alkaline phosphatase (ALP), and tartrate-resistant acid phosphatase, have been reported (Greenspan et al., 1997; Srivastava et al., 2001; Shao et al., 2003). Moreover, during ossteointegration (the process by which bone implants become integrated with the surrounding tissue), the expression of clock genes was associated with expression of cartilage ECM markers (Col2a1, Col10a1, and link protein) (Mengatto et al., 2011). It was suggested that Clock/Npas2 may act directly on E-box elements in the promoters of Col2a1, Col10a1, and Acan (aggrecan). Indeed, subsequent studies by Hinoi et al. (2006) demonstrated the presence of circadian gene expression in femurs and a functional E-box in the first intron of Col2a1. A recent study also demonstrated robust circadian rhythms in the fracture healing sites within femurs (Kunimoto et al., 2016).
In recent years, the increasing appreciation of peripheral clocks has prompted studies of the molecular clocks in resident bone cells. Among different cell types, the circadian clock in osteoblasts is most extensively studied. It has been shown that the clock affects bone formation potentially through regulating osteoblast differentiation from bone marrow–derived stem cells (Goldstrohm and Wickens, 2008). The core clock complex BMAL1/Clock directly regulates the expression of nocturnin (Noc), a deadenylase posttranscriptionally modulating gene expression (Guntur et al., 2011). Differentiation of osteoblasts inhibits Noc expression, while overexpression of Noc blocks osteoblastogenesis (Goldstrohm and Wickens, 2008; Li et al., 2008). The daily expression of Per1, Per2, and several osteoclast markers (NFATc1 and Cathepsin K, Ctsk) was shown in cancellous bone in mice, as well as in cultured osteoclasts (Fujihara et al., 2014). To our knowledge, no studies so far have characterized molecular clocks in osteocytes. However, Samsa et al. (2016) used histomorphometry to demonstrate a significantly reduced number of osteocytes (and osteoblasts) in bones of global Bmal1 KO mice, indicating a possible role of BMAL1 in the terminal differentiation of osteoblasts.
Rhythmic Targets in Bone
As a highly calcified tissue, genome-wide transcriptome studies in long bone tissue have been technically challenging. Neither has this type of study been done in cultured bone cells (osteoblasts, osteoclasts, or osteocytes). However, the analysis of the daily transcriptome in calvarial bone (ossified by intramembranous ossification) has provided some clues about the scale of clock targets within bone tissues. This approach revealed that the messenger RNA (mRNA) expression of 26% of identified genes was rhythmic with a 24-h oscillation, including bone morphogenetic protein (BMP), fibroblast growth factor (FGF), WNT signaling pathways, and genes that encode multiple ADAMTS proteases and various procollagen isoforms (Zvonic et al., 2007). Among these clock targets, a significant number of genes are related to ECM regulation (Table 1). These matrix-related CCGs strongly support a role of the circadian rhythm in maintaining homeostasis of this matrix-rich and mineralized skeletal tissue.
Entrainment Factors for the Bone Clock
The circadian clock in bone can be entrained by both external stimuli and internal signals, to maintain the daily rhythms in bone metabolism (formation and resorption) and bone mass (Bjarnason et al., 2002; Karsdal et al., 2008; Komoto et al., 2012). SCN output time cues may be transmitted to the bone through changes in leptin levels, glucocorticoid signaling, and feeding/fasting cycles (Le Minh et al., 2001; Komoto et al., 2012). Leptin was proposed as one of the entrainment signals for osteoblasts, and it may regulate the cell intrinsic clocks through activation of Beta2-adrenergic receptor (Adrb2) on the cell surface. Upon receptor activation, the level of cAMP-responsive element binding protein 1 (Creb1) was increased, leading to elevated expression of Bmal1 and Clock (Downey and Siegel, 2006). Dexamethasone stimulated the expression of clock genes, as well as induced rhythmic bone resorption in cultured osteoclasts in vitro (Fujihara et al., 2014). The rhythmic expression of NFATc1 and Ctsk was abolished in cancellous bone from adrenalectomized mice, where the amplitude of rhythmic Per1 and Per2 genes was dramatically reduced. Remarkably, glucocorticoid injection restored the circadian expression of these osteoclast-specific genes in vivo (Fujihara et al., 2014). The daily variations in bone metabolism seem to be governed by timing of food intake and fasting, providing an additional entrainment mechanism for bone (Bjarnason et al., 2002; Karsdal et al., 2008).
Dysregulation–Bone Defects
Recent studies using circadian gene KO mice have demonstrated an essential role of the bone clock in regulating bone metabolism. A high bone mass (HBM) phenotype was reported in mice lacking either the Per1 gene or the Per/Arnt/Sim (PAS) domain of Per2 from 6 weeks of age and progressed with aging, associated with an increased number of osteoblasts in bones as identified by both biochemical and histomorphometric analyses. This phenotype is consistent with the upregulated rates of mineral apposition and bone formation (Fu et al., 2005; Maronde et al., 2010). A similar HBM phenotype was also observed in mice with double KO of Per1/Per2 or Cry1/Cry2 (Fu et al., 2005). In addition, Cry2 was shown to influence osteoclast activity, whereas Per2 was reported to act on osteoblasts in mice (Maronde et al., 2010). In contrast, global deletion of Bmal1, the positive regulator within the clock, led to a low bone mass phenotype, which at least partially involves compromised capacity of mesenchymal stem cells to differentiate into osteoblasts (Samsa et al., 2016). Bmal1 KO mice are generally shorter (with significantly shorter bones), and the overall skeleton weight is lower compared with WT mice (Bunger et al., 2005; Fu et al., 2005; Kondratov et al., 2006; Takarada et al., 2012). This shorter bone phenotype was proposed to be caused by the altered circadian expression of Ihh, which is a key regulator of endochondral ossification within the growth plate (Takarada et al., 2012). Although not proven yet, these opposing phenotypes in different clock mutant animals strongly point to a mechanism in which different clock factors regulate bone formation by coordinated actions on the activities of osteoblasts and osteoclasts (Figure 1). It is therefore likely that the circadian clock in bone cells has evolved to temporally orchestrate the 2 opposing events to maintain bone homeostasis.
Therapeutic Targeting Potential
Understanding the circadian regulation of homeostasis and metabolism in cartilage and bone provides a novel avenue to treat chronic (and age-related) diseases, such as OA and osteoporosis. For instance, in cartilage from mice (a nocturnal species), it has been discovered that anabolic genes mainly peaked at early night (before activity), while catabolic genes peaked early in the day (after activity) (Dudek and Meng, 2014). Therefore, drugs targeting these pathways should be used at times of the day their targets are most active, so as to attain maximum efficacy while minimizing side effects (Kaur et al., 2013; Zhang et al., 2014). Indeed, chronotherapy, which is to deliver drugs based on the body clock time, has been trialed in OA pain management since 1980s. Remarkably, a chronotherapeutic dosing regimen showed doubled analgesic effectiveness and 4 times tolerance of indomethacin treatment to reduce OA pain (Levi et al., 1985). Furthermore, the development of compounds directly targeting molecular clocks has been described, including CK1 enzymes, CRY proteins, REV-ERB nuclear receptors, SIRT1 compounds, and lithium. Although these compounds are still at an early stage of development and mainly toward inflammation or metabolic disorders, they are also potentially useful to treat other diseases in the future (Meng et al., 2008, 2010; Hirota et al., 2012; Solt et al., 2012). Given the well-established role of cortisol in entraining peripheral clocks, local delivery of synthetic glucocorticoids to the diseased joint may have potential therapeutic value to boost the intrinsic cartilage rhythm, in addition to their anti-inflammatory actions. Moreover, as the rhythmic genes in cartilage and bone are controlled not only by the local clocks but also by the time cues that are under the control of the central clock, it is possible to systemically target the circadian system to slow down disease progression. Mechanical stimulation through exercises is well known to improve cartilage and bone function. Interestingly, the mammalian and Drosophila circadian clocks have previously been identified as mechanosensitive (Kanbe et al., 2006; Simoni et al., 2014). If mechanical stimulation can reentrain the clocks in cartilage and bone, perhaps a regular schedule of gentle exercise would directly benefit the local skeletal clocks in addition to having feedback effects on the central clock in the brain. From the circadian clock perspective, it is probably worth considering not only the duration of the physical exercise but also the timing. Although speculative at this stage, sticking to a regular exercise scheme on a daily basis is likely to be more beneficial than exercising at random times. Perhaps a personalized schedule, according to an individual’s chronotype (preference to be alert early or late), would work most efficiently (Facer-Childs and Brandstaetter, 2015; Roenneberg and Merrow, 2016). Additional systemic time cues (e.g., regular mealtimes) may also be used to enhance the impaired skeletal clock during diseases or even aging (Feillet et al., 2006; Kanbe et al., 2006; Hughes and Piggins, 2012; Simoni et al., 2014).
Concluding Remarks and Future Directions
The identification of functional circadian clocks in matrix-rich cartilage and bone tissues has opened up an exciting new avenue for skeletal research. This temporal control mechanism provides a critical time cue for chondrocytes and resident bone cells to coordinate ECM homeostasis with the daily rhythmic cycles of rest/activity, body temperature fluctuations, and surges of hormones. During aging or in shift workers, disruptions to circadian rhythms may compromise this protective mechanism, leading to increased risk of tissue pathologies. Future work should be directed toward understanding the major clock-controlled pathways in articular cartilage and long bone, as well as establishing cause-effect relationships using genetic studies in animal models (such as cell type–specific ablation of individual clock components in chondrocytes or different bone cells). The musculoskeletal system is highly relevant to daily rest/activity cycles, and there may be a strong component of mechanical control for skeletal clocks. Moreover, it is important to demonstrate changes of the circadian clock in patients with skeletal diseases that affect cartilage and bone. So far, detailed studies of clocks in different types of cartilage, different joints, and different bone cell types are still lacking. Finally, effects of clock-acting interventions (systemic approaches, pharmacological compounds, or chronotherapeutic dosing of drugs) on cartilage/bone metabolism and matrix homeostasis should be tested in vivo in experimental models.
Footnotes
Acknowledgements
This work has been funded by the following grants: a Medical Research Council (MRC, UK) project grant (MR/K019392/1), a Wellcome Trust (UK) Core funding (088785/Z/09/Z) to the Wellcome Trust Centre for Cell-Matrix Research, and an Arthritis Research UK Senior Research Fellowship (20875).
Conflict of Interest Statement
The author(s) have no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.