
Abstract
Our understanding of the molecular underpinnings of the mammalian circadian oscillator and its tight connection to physiology has progressed tremendously during the past decades. The liver is considered the prototypic experimental model tissue for circadian research in peripheral organs. Studies on liver clocks have been highly productive and yielded information about widely different aspects of circadian biology. The liver, as one of the largest organs in the body, has often been used for the identification of core clock and auxiliary clock components, for example, by biochemical purifications. Because the liver is also a major metabolic hub, studies addressing the interplay between circadian clocks and metabolism have been insightful. In addition, the use of liver-specific loss-of-function models for clock components highlighted not only specific physiological roles of the hepatic clock but also its interplay with systemic cues and oscillators in other organs. Recently, technological advances in omics approaches have been successfully applied on the liver, providing a comprehensive depiction of pervasive circadian control of gene expression and protein and metabolite accumulation. In this review, we chose to illuminate specific examples that demonstrate how different experimental approaches—namely, biochemical, metabolic, genetic, and omics methodologies—have advanced our knowledge regarding circadian liver biology and chronobiology in general.
The mammalian circadian clock is structured in a hierarchical manner. A master clock in the suprachiasmatic nuclei (SCN) in the brain is entrained by light-dark cycles and synchronizes peripheral clocks present in all cells of the body through various redundant pathways (e.g., neuronal, endocrine, and blood-borne signals) (Balsalobre et al., 2000; Reddy et al., 2007; Welsh et al., 2010; Gerber et al., 2013). The SCN and peripheral clocks are assembled from cell-autonomous oscillators that function on the basis of transcription-translation feedback loops. The transcription factors BMAL1 and CLOCK drive the expression of the negative feedback loop members—namely, the Per and Cry genes. In turn, once PER and CRY proteins accumulate, they bind BMAL1/CLOCK and inhibit their own transcription. A reinforcing feedback loop composed of ROR and REV-ERB family members controls Bmal1 gene expression. In addition, diverse posttranscriptional processes fine-tune circadian oscillations in a tissue-specific manner (Partch et al., 2014; Zhao et al., 2014; Asher and Sassone-Corsi, 2015; Dibner and Schibler, 2015; Fang and Lazar, 2015).
Since the identification of the first mammalian core clock gene about 20 years ago by Takahashi and coworkers (King et al., 1997), enormous progress has been made in unraveling the molecular framework of the core clock circuitry and its various physiological and metabolic traits. Remarkably, chronobiology studies done on liver have turned out to be a fertile ground, advancing our knowledge regarding almost every aspect of circadian biology. The liver has been used for the identification of core clock and auxiliary clock components through various biochemical approaches. Liver-specific genetic models for clock genes have been instrumental in identifying specific physiological roles for the hepatic clock and provided an integrated whole-organism view of clock functions. The application of recently developed omics approaches has provided a comprehensive depiction of the pervasive circadian control of gene expression and protein and metabolite accumulation. In this review, we wish to highlight specific examples that illustrate how the liver as a model organ has been used in combination with different experimental methodologies (Figure 1)—namely, biochemical, metabolic, genetic, and system approaches—to address eminent and open questions related to circadian liver biology and chronobiology in general. We feel that it is beyond the scope of this review to provide a comprehensive overview of all aspects of circadian liver biology. In this regard, we would like to refer the reader to a number of reviews that were published in recent years (Kornmann et al., 2007b; Maywood et al., 2007; Schmutz et al., 2012; Everett and Lazar, 2014; Hong et al., 2014; Adamovich et al., 2015; Li and Lin, 2015; Reinke and Asher, 2015).
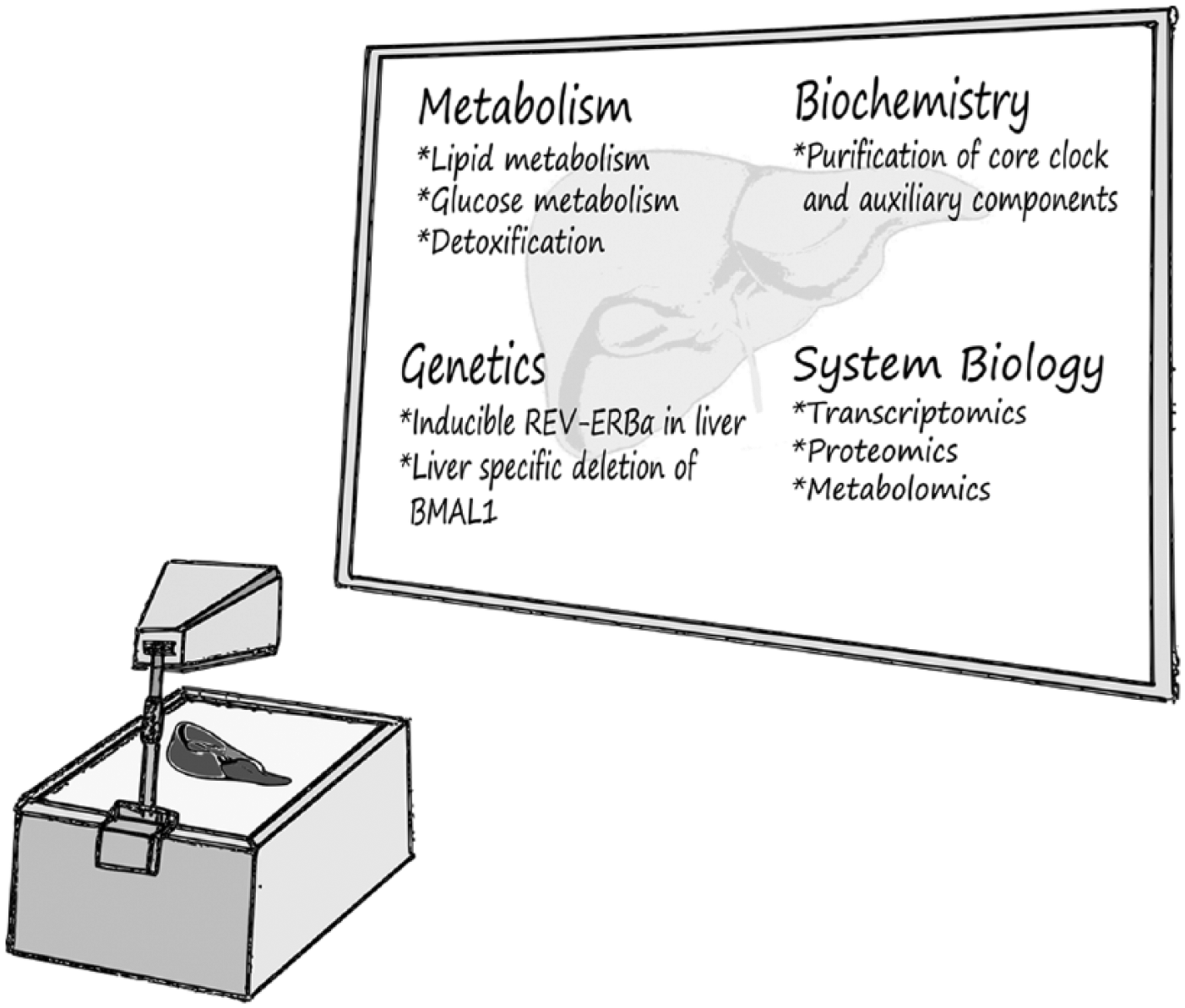
The liver as a prototype for studying circadian biology. Various experimental approaches, ranging from biochemical through genetic to metabolic approaches, have been applied on liver samples to study different aspects of circadian biology. These approaches have advanced our knowledge regarding the molecular architecture of the core clock circuitry, the physiological significance of hepatic clocks on liver metabolism, and whole-body physiology.
Biochemical Approaches on Liver Tissue Reveal Novel Clock Components and Regulatory Mechanisms
The liver is the second largest organ in our body after the skin. It consists of parenchymal hepatocytes and various resident nonparenchymal cells, including Kupffer cell macrophages, fat-storing stellate cells, and endothelial cells (Baratta et al., 2009). Most cells are hepatocytes, which makes the liver cell population rather homogeneous compared to most other organs. Anatomically, the liver is relatively easy to access and harvest, and the tissue is readily homogenized due to its parenchymal nature. Established protocols for the separation of liver cells into their organelle constituents (e.g., nuclei, cytoplasm, mitochondria) exist, providing researchers with ideal starting material for biochemical experiments. Particularly attractive is the possibility to examine the size and composition of large protein complexes that are associated with clock components. Temporal and spatial analyses of dynamic changes in these protein complexes are highly informative and can be used to identify and study core clock components and clock output proteins as detailed below.
The Weitz group has extensively used the potential of this approach during the past couple of years. Their findings revealed that the majority of BMAL1 is present in complexes of 200 to 400 kD, but a fraction of BMAL1 seemed to be present in even larger protein complexes. A subsequent purification of BMAL1-containing complexes from Bmal1-deficient cells expressing a FLAG-hemagglutinin tandem epitope tagged BMAL1 revealed RACK1 and PKCα as nuclear components of BMAL1-complexes during the repression phase of the molecular oscillation at nighttime. RACK1 and PKCα were shown in cultured cells to be essential for the function of the core clock (Robles et al., 2010). In a recent study, CLOCK/BMAL1-containing complexes were purified from liver by an alternative method that relies on the affinity of CLOCK/BMAL1 toward E-box–containing oligonucleotides. In this assay, the adaptor protein WDR76 interacted specifically during the daytime activation phase with CLOCK/BMAL1 in mouse liver. WDR76, in turn, recruited the Ddb1–Cullin-4 ubiquitin ligase complex to Per gene promoters and other CLOCK/BMAL1 target genes and marked them for repression by PER-containing protein complexes (Tamayo et al., 2015).
The purification of core clock protein-containing complexes from mouse organs was further extended to the PER proteins. A previous report found PER1 and PER2 to exist in very large complexes that change in size during the day (Brown et al., 2005). For the purpose of purification from mouse liver, the Weitz group generated 2 transgenic lines that express either PER1 or PER2 as FLAG-hemagglutinin fusion proteins. This approach has the additional advantage that complexes can be purified with 2 different components as bait, thus providing enhanced specificity. Both variants were functional as assayed in circadian running wheel analyses, and their protein expression levels oscillated throughout the day similarly to their wild-type counterparts. The purification of protein complexes from the livers of these mice resulted in novel and unexpected mechanistic insights into the repressive functions of PER proteins. It is noteworthy that most of the interactions that were identified might be rather labile because a reversible protein-protein crosslinker was present during the affinity purification step of the procedure. Additionally, most interacting proteins were purified in substochiometric amounts. Remarkably, nucleic acids were also copurified, and ribonuclease treatment shifted PER-complexes toward a smaller size indicating a stabilizing function of associated RNAs. Reassuringly, many known components of PER-complexes were found, among them PER1-3, CRY1-2, CLOCK, BMAL1, CSNK1D1, CSNK1E1, and NONO. Novel interacting proteins that were characterized further were the RNA-binding proteins PSF, DDX5, and DHX9. PSF, a pre-mRNA splicing factor related to NONO (Brown et al., 2005), recruits repressive histone deacetylase complexes to target gene promoters (Duong et al., 2011). Additionally, the RNA helicases DDX5 and DHX9 were identified as part of a complex mechanism that guarantees proper elongation and termination by RNA polymerase as well as processing of the resulting mRNAs (Padmanabhan et al., 2012). A variation of the purification method comprising high salt extraction of proteins from chromatin resulted in a 1.2-MD Period protein complex, which was significantly bigger than the 800-kD complex that was identified with the previous extraction methods and which might represent a subcomplex. This purification method also revealed NuRD, a chromatin-associated corepressor complex, to be split into one subcomplex that acts as a coactivating mechanism for CLOCK/BMAL1 and another subcomplex that supports the repressive function of Per proteins (Kim et al., 2014). In summary, protein purification from mouse liver turned out to be an effective and fruitful approach due to the availability of large amounts of homogeneous tissue. It offers a unique opportunity to study the dynamics of large protein complexes and allows the detection of weak interactions, substochiometric complex members, nucleic acid components, and splitting of complexes that are likely to play a role in the core clock circuitry. Although these studies were performed with liver tissue, it is conceivable that their findings are relevant for the clock machinery in general and not specifically for the liver clock.
The Liver and Circadian Metabolism
The liver is a metabolic organ that performs essential functions in detoxification, anabolic and catabolic processes, synthesis of serum proteins, and many other homeostatic processes. Like most other tissues in the body, it has to adapt to periodic changes in its systemic milieu since mammals exhibit rest-activity and feeding-fasting cycles throughout the day (Asher and Schibler, 2011). Therefore, essentially all physiological and metabolic processes in the liver are regulated in a daily manner (Figure 2) (see also relevant sections in the omics part detailed below). Rhythmic regulation of liver metabolism can be driven by the circadian clock or by systemic cues, or in many instances by a combination of both. In particular, feeding is a dominant circadian timing signal (zeitgeber), which is relayed to the circadian clock by nutrients and metabolites, for example, NAD (Nakahata et al., 2009; Ramsey et al., 2009), glucose (Stephan and Davidson, 1998), and polyamines (Zwighaft et al., 2015). Interestingly, some metabolic pathways are organized in the form of distinct feedback loops that are coupled to the core circadian oscillator. They can function predominantly intracellularly (e.g., NAD+/NADH) or exert systemic effects on clocks within different organs (e.g., glucose). Energy and metabolite levels are signaled to the molecular oscillator through metabolic sensors such as SIRT proteins (Asher et al., 2008; Nakahata et al., 2008; Chang and Guarente, 2013; Masri et al., 2014), PARP-1 (Asher et al., 2010), AMPK (Lamia et al., 2009), and PGC-1α (Liu et al., 2007). Redox responsiveness of the circadian system, for which proof of principle has been demonstrated with purified core clock transcription factors (Rutter et al., 2001), has only been analyzed in red blood cells but may also have implications for the adaptation of the liver clock to nutrient availability and metabolic state (O’Neill and Reddy, 2011; O’Neill et al., 2011), Additionally, temperature sensors (Reinke et al., 2008; Buhr et al., 2010; Saini et al., 2012) are hardwired to cellular clocks. Importantly, entrainment of the liver oscillator by feeding-fasting cycles is dominant over all other known zeitgebers in peripheral organs and can fully uncouple the liver clock from the central pacemaker in the SCN (Damiola et al., 2000).
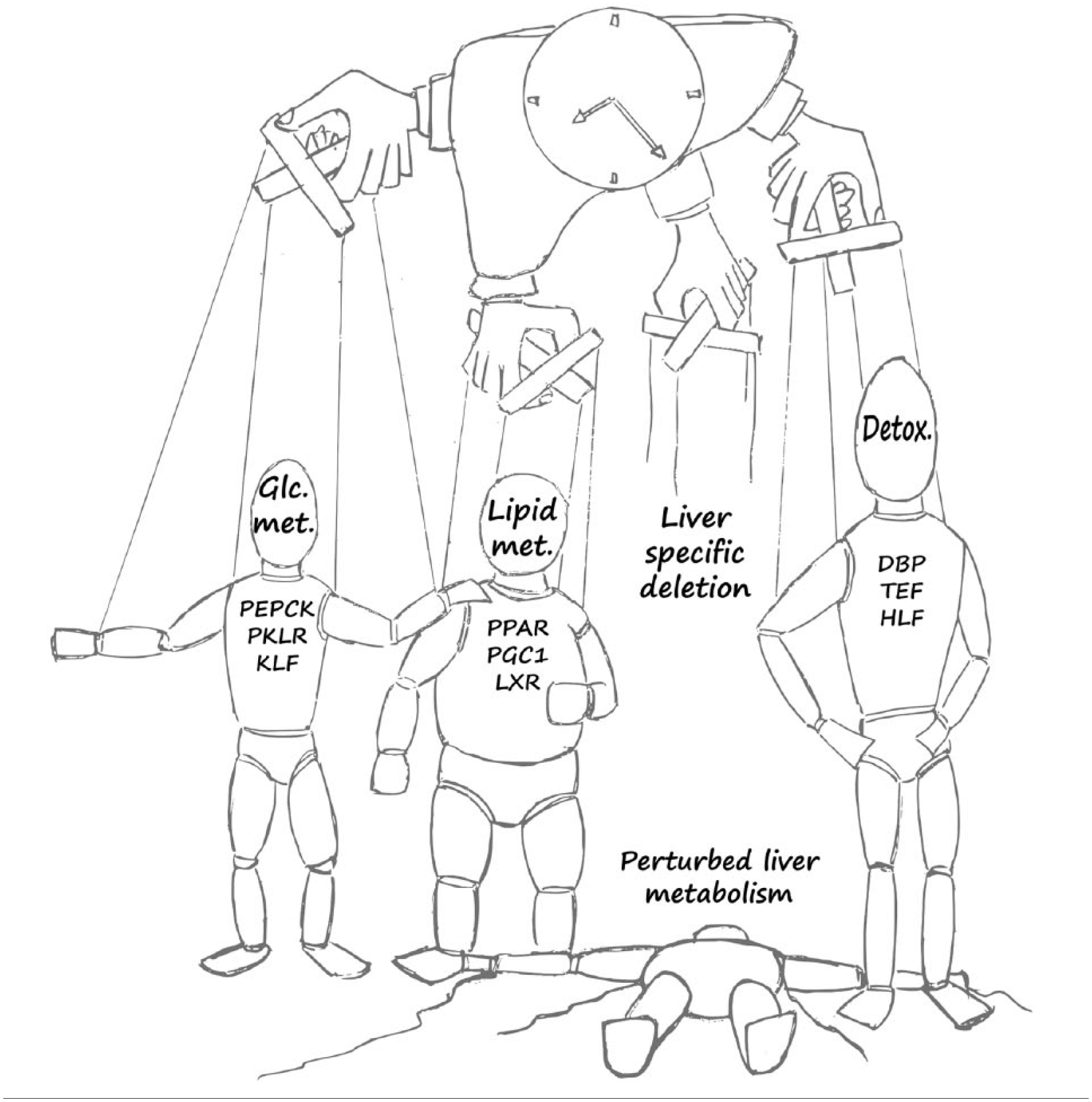
The liver as a metabolic hub and its regulation by the circadian clock. Daily expression of master regulators and rate-limiting enzymes of key hepatic metabolic outputs related to glucose metabolism (Glc. Metab.) (e.g., phosphoenolpyruvate carboxykinase [PEPCK], pyruvate kinase isozymes R [PKLR], Krüppel-like factor [KLF]), lipid metabolism (Lipid Met.) (e.g., peroxisome proliferator-activated receptors [PPARs], peroxisome proliferator-activated receptor gamma coactivator 1-alpha [PGC1], liver X receptor [LXR]), and detoxification (Detox.) (e.g., D site of albumin promoter [DBP], thyrotroph embryonic factor [TEF], hepatic leukemia factor [HLF]).
Analyses of global mRNA expression levels around the day initially revealed a comprehensive picture of the daily regulation of liver metabolism (see also relevant sections in the omics part detailed below) (Akhtar et al., 2002; Panda et al., 2002; Storch et al., 2002; Zhang et al., 2014). Virtually all hepatic processes appear to be under the control of the circadian clock, including detoxification, cholesterol and bile acid metabolism, glucose and lipid homeostasis, production of thyroxine hormone, and many more. Interestingly, global peak expression levels were observed at the end of the light and dark phases, which indicates different physiological requirements mediated by arousal and the start of the resting period.
The liver is a central organ for the control of blood glucose levels, which serves as a good example for the tight interplay between circadian and metabolic regulation. The combined results of several studies revealed complex clock-dependent regulation by positive and negative regulators of the core clock mechanism with multiple clock-output pathways converging on rate-limiting enzymes of glucose metabolism. Several CLOCK/BMAL1-driven transcription factors have been shown to regulate the expression of glycolytic and gluconeogenic genes. CRY proteins use alternative mechanisms by interacting either with G protein–coupled receptors or glucocorticoid receptor to control gluconeogenesis in the liver (So et al., 2009; Lamia et al., 2011). Krüppel-like transcription factors connect the circadian clock to the expression of glycolytic and gluconeogenic genes, in addition to bile acid metabolism and nitrogen homeostasis (Gray et al., 2007; Guillaumond et al., 2010; Jeyaraj et al., 2012; Han et al., 2015). Importantly, specific loss of clock function in the liver leads to large fluctuations in blood glucose levels (Lamia et al., 2008).
Recent studies indicate that mitochondrial functions are also under circadian control (Peek et al., 2013; Jacobi et al., 2015). Mitochondrial nutrient utilization—namely, the ability of mitochondria to metabolize carbohydrates versus fatty acids—is under circadian control, in part through the temporal accumulation of the respective rate-limiting enzymes (Neufeld-Cohen et al., 2016). These examples demonstrate that metabolic and circadian regulation is tightly intertwined in the liver, which therefore serves as an organ of choice for studies of physiological and pathological processes involving metabolic regulation and core clock control.
Genetic Models of Liver Specific Knockouts of Clock Components
The hierarchical structure of the mammalian circadian timing system raises the question to which extent the physiological and metabolic regulation in peripheral organs such as the liver is dominated by the central clock in the SCN, tissue-specific mechanisms, or both. Hence, determining the specific role of hepatic clocks in vivo and distinguishing them from systemic effects driven by the master clock in the brain and/or other peripheral organs is of great interest. To this aim, mouse models harboring a liver-specific nonfunctional clock have been generated.
Schibler and colleagues carried out the first endeavor and generated a transgenic mouse line that overexpresses REV-ERBα exclusively in hepatocytes (Kornmann et al., 2007a). Overexpression of hepatic REV-ERBα is under the control of a Tetracyclin (Tet)–off regulatory system and thus can be switched on and off at will. Once doxycycline is omitted from the diet, REV-ERBα is overexpressed in hepatocytes, resulting in strong repression of Bmal1 expression and consequently a nonfunctional clock in these cells. With this mouse model, they conducted a genome-wide comparison between the circadian liver transcriptome of mice with and without a functional clock (i.e., doxycycline-treated and untreated mice). They found that although most of the genes that cycle in liver cease to oscillate when the hepatic clock is abrogated, some genes, including the clock’s negative limb members Per1, Per2, Cry1, and Cry2 and a handful of other genes, including heat-shock genes, retain their rhythmic expression. They concluded that the master clock in the SCN generates, on a daily basis, systemic cues that merely support the peripheral clock, which is the main driving force underlying circadian rhythmicity in liver.
Weitz and coworkers took a different approach to ablate the clock in mouse liver. They employed a Cre-loxP recombination strategy to generate a liver-specific deletion of Bmal1 (L-Bmal1−/−) or to inactivate Bmal1 in all body cells (Bmal1−/−) (Lamia et al., 2008). L-Bmal1−/− mice suffer from hypoglycemia, specifically during the inactivity phase. By contrast, mice deficient for BMAL1 in the whole body do not display overt perturbations of their resting blood glucose levels. These findings suggest that in liver, BMAL1 contributes to glucose homeostasis by controlling a daily rhythm of hepatic glucose export that counterbalances the daily cycle of glucose ingestion resulting from the SCN-driven fasting-feeding cycle. Hence, the liver clock is important for buffering circulating glucose in a time-of-day–dependent manner.
In these 2 mouse models, clock function was disrupted in hepatocytes through direct or indirect interference with Bmal1 expression. Hence, the observed phenotypes might be simply related to missing functions of BMAL1-controlled output genes. Future studies with different liver-specific clock knockouts—for example, by loss-of-function of Per/Cry genes, RORE (ROR response element), or D-Box binding genes (Fang et al., 2014)—are bound to unveil additional physiological phenotypes. The intersection of several knockout model systems should reveal bona fide processes that are more likely related to circadian oscillation of the liver than to specific functions of individual core clock components. Furthermore, these liver-specific knockout models are highly valuable in dissecting the specific role of the liver clock and its interplay with systemic cues emerging from the SCN and other peripheral organs. The use of various omics approaches, which are the subject of the next section, that emerged as powerful tools during recent years is expected to shed more light on these issues.
The Circadian Omics Era and The Liver
The liver has turned out to be a fertile ground for a wide variety of omics studies as well, ranging from genome-wide analysis of core clock transcription factor binding, through liver global transcriptomics and recently ribosome profiling, to proteomics and finally metabolomics. Comprehensive examination of the transcriptional architecture of the circadian transcriptional regulatory loop on a genome-wide scale in mouse liver identified time-dependent patterns of transcription factor binding at promoter and enhancer regions and of chromatin states (Rey et al., 2011; Koike et al., 2012; Fang et al., 2014). Additional studies measuring nascent RNA transcription and regulatory RNAs around the clock in mouse liver characterized circadian enhancers and other circadian regulatory mechanisms (Vollmers et al., 2012; Fang and Lazar, 2015).
Extensive transcriptome profiling performed throughout the day in the liver has demonstrated the pervasive circadian control of physiology and metabolism (Akhtar et al., 2002; Panda et al., 2002; Storch et al., 2002; Vollmers et al., 2012). As mentioned above, these studies highlighted various metabolic pathways (e.g., glucose homeostasis, lipid metabolism, detoxification) that are rhythmic and potentially under circadian clock control. A recent comprehensive analysis found that the liver had more circadian genes (~3000, ~16%) than any other organ studied so far (i.e., aorta, adrenal gland, brain, adipose tissue, kidney, lung, and skeletal muscle) (Zhang et al., 2014). Furthermore, in agreement with previous genome-wide analyses, it emphasized the tissue specificity of these transcriptional rhythms, although the molecular clock is active in the entire body. More recent studies monitored simultaneously mRNA transcription, accumulation, and translation using total RNA sequencing upon ribosome profiling and uncovered a subset of genes that are translated in a rhythmic fashion (Atger et al., 2015; Janich et al., 2015).
In addition, liver proteomics analyses identified circadian oscillations in protein abundance. Remarkably, comparison of circadian proteome and transcriptome data demonstrated that for some genes, the daily patterns of mRNA and protein accumulation were not correlated. For instance, the levels of many proteins encoded by rhythmically expressed mRNAs do not oscillate at significant levels, while some proteins produced by constantly expressed transcripts do cycle in abundance (Reddy et al., 2006; Mauvoisin et al., 2014; Robles et al., 2014). Furthermore, recent proteomics analysis of isolated mitochondria from mouse liver throughout the day identified extensive oscillations in the mitochondrial proteome. The majority of cycling mitochondrial proteins peaked during the early light phase. Of note, there was very low correlation between the phase of cycling mitochondrial proteins and their respective transcripts, suggesting that posttranscriptional mechanisms play a central role in mitochondrial protein accumulation (Neufeld-Cohen et al., 2016).
In view of the expansion of advanced technologies, several labs commenced to employ high-throughput metabolomics approaches to study circadian rhythms. Studies conducted with mouse liver samples from wild-type and clock mutant mice identified clock-controlled circadian oscillations of various metabolite groups, including amino acids, carbohydrates, lipids, and, more specifically, fatty acids (Eckel-Mahan et al., 2012). An integrative database was launched (CircadiOmics database) that combines circadian genomics, transcriptomics, proteomics, and metabolomics (Patel et al., 2012), thereby facilitating the use of the currently available data to decipher circadian control mechanisms of various metabolic pathways. A subsequent circadian lipidomics analysis on mouse liver revealed ~17% of all analyzed lipids to display circadian rhythmicity (Adamovich et al., 2014).
In summary, different omics approaches provided tremendous insight into the intricate mechanisms implicated in rhythmic transcription/translation and protein and metabolite accumulation. Yet the integration of these different data sets is challenging and still in its infancy. Here again, many of the findings might be applicable to any other peripheral organ and even to the master clock in the brain. Future studies are expected to shed more light on the different mechanisms and integrate them into a conclusive and coherent picture.
Summary and Open Questions
The liver has been a rewarding model system for circadian research. Most of our knowledge about peripheral clocks and their integration into mammalian physiology and metabolism stems from genetic or biochemical studies of liver tissue. In recent years, these findings have been complemented by global analyses of central molecules such as mRNAs, proteins, and small metabolites, yet these data sets should be better integrated into coherent models. Examples for the current barriers that need to be overcome are the use of different genetic backgrounds in animal studies, varying experimental paradigms and nonuniform statistical evaluation and data presentation in omics experiments. Guidelines regarding sampling frequencies, number of replicates, and uniform criteria for statistical analyses (e.g., false discovery rate) are expected to provide us with better and more reliable data sets that can be integrated into a coherent and comprehensive picture and thus will better serve the scientific community.
As discussed above, in addition to hepatocytes, which comprise 60% of the total cells and 80% of the volume of liver (Malarkey et al., 2005), the liver contains many other different cell types. Hence, it would be interesting to examine the contribution of these different cell types to the circadian rhythmicity of the various liver functions. Furthermore, the unique liver architecture, which consists of liver lobules of hexagonal structure, is composed of hepatocytes radiating from a central vein that carries blood out from the liver and portal triads (i.e., hepatic artery, hepatic portal vein, and bile duct) that deliver blood and nutrients into the liver and are present along each of the lobule’s corners. This intricate structure generates a gradient of nutrient availability from the portal triads in the periphery to the central vein at the center and therefore might play a role in the “metabolic information flow” that participates in synchronization of circadian clocks in liver. Future studies are likely to face these questions and address them in depth.
One eminent question in the field and obviously a major challenge we are still facing is related to the specific function and the importance of the liver clock. Several studies commenced addressing this issue, for example, by pinpointing the role of the liver clock in the systemic control of blood glucose levels. Nevertheless, these studies represent only the tip of the iceberg, and assessing the role of the liver clock in the complex context of whole-body physiology and metabolism is extremely important. Deregulated liver physiology is a critical parameter in the development of metabolic syndrome and fatty liver disease in humans. The circadian clock regulates all essential liver functions and is therefore an obvious target for intervention strategies aimed at the prevention and treatment of these and related medical conditions. The implementation and generation of additional liver-specific knockout models in combination with biochemical purification methods and systems biology approaches will undoubtedly be critical in these endeavors.
Footnotes
Acknowledgements
We apologize to all colleagues whose work could not be cited due to space limitations. The authors gratefully acknowledge funding by the German-Israeli Foundation for Scientific Research and Development (G-1199-230.9/2012). H.R. is supported by the Deutsche Forschungsgemeinschaft (RE 3046/2-1). G.A. is supported by the Israel Science Foundation (ISF 138/12), the HFSP Career Development Award (HFSP CDA00014/2012), and the European Research Council (ERC-2011 METACYCLES 310320). G.A. is recipient of the EMBO young investigator award and incumbent of the Pauline Recanati career development chair.
Conflict of Interest Statement
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.