
Abstract
Visible light on Earth largely comes from the sun, including light reflected from the moon. Predation risk is strongly determined by light conditions, and some animals are nocturnal to reduce predation. Artificial lights and its consequent light pollution may disrupt this natural behavior. Here, we used 13 years of attendance data to study the effects of sun, moon, and artificial light on the attendance pattern of a nocturnal seabird, the little penguin Eudyptula minor at Phillip Island, Australia. The little penguin is the smallest and the only penguin species whose activity on land is strictly nocturnal. Automated monitoring systems recorded individually marked penguins every time they arrived (after sunset) at or departed (before sunrise) from 2 colonies under different lighting conditions: natural night skylight and artificial lights (around 3 lux) used to enhance penguin viewing for ecotourism around sunset. Sunlight had a strong effect on attendance as penguins arrived on average around 81 min after sunset and departed around 92 min before sunrise. The effect of moonlight was also strong, varying according to moon phase. Fewer penguins came ashore during full moon nights. Moon phase effect was stronger on departure than arrival times. Thus, during nights between full moon and last quarter, arrival times (after sunset) were delayed, even though moonlight levels were low, while departure times (before sunrise) were earlier, coinciding with high moonlight levels. Cyclic patterns of moon effect were slightly out of phase but significantly between 2 colonies, which could be due to site-specific differences or presence/absence of artificial lights. Moonlight could be overridden by artificial light at our artificially lit colony, but the similar amplitude of attendance patterns between colonies suggests that artificial light did not mask the moonlight effect. Further research is indeed necessary to understand how seabirds respond to the increasing artificial night light levels.
Sunlight is the main source of energy for biological systems on the surface of the Earth (Bradshaw and Holzapfel, 2010). Rotation, orbital motion, and tilt of the Earth cause marked daily and seasonal variation on sunlight levels. Even at night, the reflected sunlight from the moon (i.e., moonlight) causes high variation of light levels because of the moon’s orbit around the Earth. Organisms on the surface of the Earth have evolved under this regime of light-dark cycles, which have remained practically invariant for long time periods, although recent proliferation of artificial lighting systems could disrupt these natural cycles (Gaston et al., 2014).
Animals are commonly classified as diurnal or nocturnal on the basis of the effect of sunlight on their activity patterns. For example, humans are largely diurnal as they are mainly active during daylight hours, although usage of artificial light during the past decades has extended our activity into night hours (Navara and Nelson, 2007; de la Iglesia et al., 2015). Many other diurnal species have also benefited by extending their foraging times at artificially illuminated areas (Negro et al., 2000; Santos et al., 2010; Dominoni et al., 2013). Among nocturnal animals, moonlight effects have been widely studied in the context of reproduction (e.g., Harrison et al., 1984), communication (e.g., Penteriani et al., 2010), predator-prey relationships (e.g., Daly et al., 1992; Penteriani et al., 2013; Saldaña-Vázquez and Munguía-Rosas, 2013), and anthropogenic-induced mortality (Gundersen and Andreassen, 1998; Rodríguez and Rodríguez, 2009; Mizuta, 2014). Artificial lights can be brighter than the moon, and thus they have the potential to mask the effect of moonlight on the physiology and behavior of nocturnal and diurnal animals (reviewed in Kronfeld-Schor et al., 2013).
In marine ecosystems, the effect of light on seabirds and marine mammals is more difficult to assess due to their life cycle. Marine-adapted terrestrial breeding species have to cope with a trade-off as they live in contrasting marine and terrestrial environments. If they are well adapted to the marine environment, they will, by comparison, underperform on land (Griffin and Kram, 2000). Many, particularly the smaller, of them show nocturnal behavior on land (visit the breeding colonies at night) in an attempt to avoid predation. Seabirds, such as petrels and murrelets, are typical cases. Commuting behavior (i.e., regular travels between breeding colonies and foraging locations) is strongly influenced by moon phase in petrels (e.g., Riou and Hamer, 2008; Bourgeois et al., 2008; Taylor et al., 2012; Rubolini et al., 2014) often by reducing visits to colonies and calling during moonlit nights (Mougeot and Bretagnolle, 2000; Miles et al., 2013). Artificially increased light levels may allow visual predators to be more efficient. For example, higher predation rates have been estimated in storm-petrel colonies under the influence of artificial lights (Oro et al., 2005).
Among seabirds, the little penguin Eudyptula minor is a good model for studying the effect of light on the annual pattern of colony attendance. Even outside the breeding season, this resident species returns ashore on a regular basis, and thus, adults are present at colonies throughout the year. Little penguins are diurnal foragers at sea but nocturnal on land (Stahel and Gales, 1987). They arrive at the colony after sunset and depart before sunrise (Klomp and Wooller, 1991; Davis and Renner, 2003). During breeding, the little penguin is a diurnal central-place forager with normal foraging trips of 1 to 10 days before returning ashore at night for courtship, to incubate eggs or feed chicks (Chiaradia and Nisbet, 2006; Saraux et al., 2011). During daylight hours on land, they stay in burrows, underground, or under vegetation but never venture out in the open, showing a typical nocturnal behavior on land. In this study, we assessed how sunlight and moonlight affected attendance patterns ashore of little penguins at Phillip Island, Australia, by automatically recording arrival and departure times at 2 colonies over 13 years. First, we evaluated the effect of sunlight on the attendance behavior of penguins annually to examine if arrivals and departures from colonies varied seasonally with respect to sunset or sunrise, respectively. Second, we studied the effect of moonlight on the arrival and departure times. Third, we assessed the effect of moonlight on the number of penguins coming ashore at sunset and going to the sea before sunrise. Last, we evaluated if artificial illumination at penguin arrival time after sunset can affect the natural attendance behavior of penguins. We compared 2 distinct colonies: one colony under a natural night sky and another with artificial illumination at an ecotourism penguin viewing attraction, the Penguin Parade, on Phillip Island, Australia. We hypothesized that arrival and departure times are influenced by moonlight with penguins arriving later in the night or departing earlier in the morning during the brighter nights around the full moon. Similarly, we expected a lower number of penguins at colonies (arriving or departing) during the brighter moonlit nights. Finally, we expected penguin arrivals were independent of moon cycle at the colony lit with artificial light, as artificial light intensity can override moonlight (Kronfeld-Schor et al., 2013).
Methods
Study Sites
Phillip Island in southeastern Australia (S 38°29′, E 145°15′) sustains more than 1% of the global population of little penguins E. minor (BirdLife International, 2015). Approximately 26,000 to 28,000 adult penguins breed on the island (Sutherland and Dann, 2012). We examined the arrival and departure patterns of little penguins at the Penguin Parade and Cullen Cove on Summerlands Peninsula. The study period spanned from 1 June 2001 to 31 August 2014 (13 years) at Penguin Parade (S 38°30′37′′, E 145°09′01′′), and from 17 July 2003 to 16 January 2014 (10 years) at Cullen Cove (S 38°31′04′′, E 145°07′16′′). The Penguin Parade colony is a popular ecotourism destination, receiving over half million tourists each year to watch little penguins crossing the beach in the evening (Dann and Chambers, 2013). Some form of artificial lighting has been used at the Penguin Parade from the 1960s, but since 1987, orange halogen lights (Linear halogen, 240 V, 2600 lm; Osram Sylvania, Danvers, MA, USA) of around 3 lux have been turned on from sunset to 1.5 h after the arrival of the first penguins to enhance the viewing of penguins. At Penguin Parade, penguins emerge at a white sand beach, crossing a gentle slope of 15 to 110 m of the beach depending on the tide height that oscillates by up to 1.5 m (Laaksonen, 2011). Cullen Cove is closed to the public at night, and it is under a natural light regime as no artificial lights occur in the vicinity (Suppl. Figure S1). At Cullen Cove, penguins cross a shorter stretch of basaltic rock beach between 20 and 35 m at the same 1.5-m tide oscillation.
Arrival and Departure Patterns
Individual penguins were marked with unique passive integrated transponders (PITs). Arrival and departure patterns were continuously recorded by an automated penguin monitoring system located in Penguin Parade colony (see Chiaradia and Kerry, 1999), developed by the Australian Antarctic Division (Kerry et al., 1993). A simple data-logger system consisting of a transponder reader platform was set up at Cullen Cove. Both systems record time and date of attendance of PIT-marked penguins, and they were located less than 15 m above the high tide mark and less than 10 m from the edge of nesting sites (Suppl. Figure S1).
Variables
Repeated readings from a same individual on the same night and direction (i.e., arrival or departure) were removed, only keeping the earliest arrival or the latest departure reading. To build statistical models (see below), we also removed outlier readings of birds arriving or departing after or before 5 min of sunset or sunrise, respectively (less than 0.1% of the data set). Outliers were kept in Figure 1 to provide a complete view of attendance pattern, but they were not included in the statistical analyses.
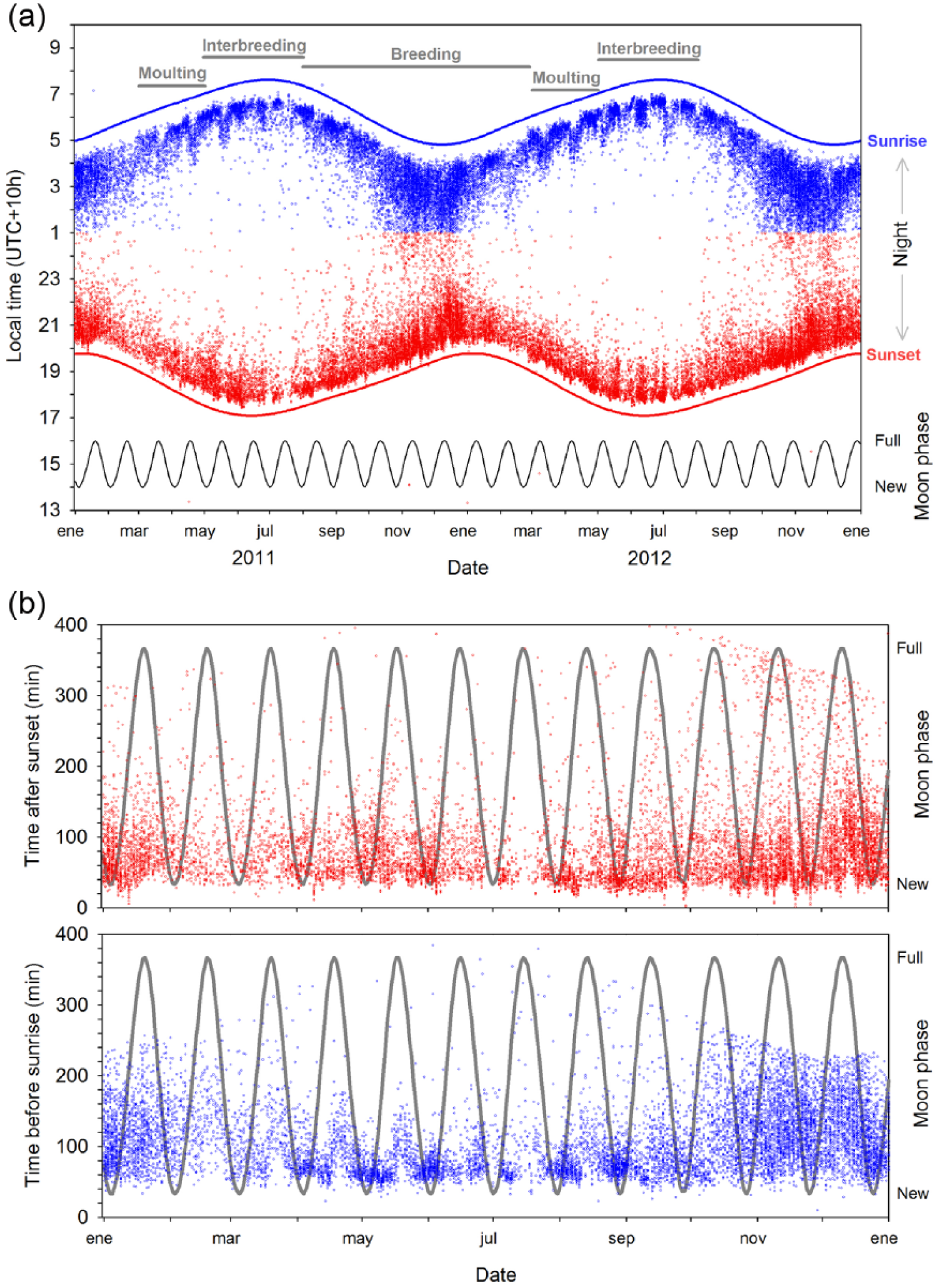
Times of arrival (after sunset) and departures (before sunrise) of little penguins at Penguin Parade, Phillip Island, Australia. (a) Seasonal sunlight effects on the arrival and departure of penguins. Breeding phenology (moulting, interbreeding and breeding seasons) and events of sunset and sunrise are shown during a two-year period (2011–2012). (b) Effect of the moon phase (grey lines) on the attendance time relative to sunset (arrivals) or sunrise (departures) during a year (2011).
Apparent sunset and sunrise times were obtained from the Earth Research System Lab, NOAA (available at http://www.esrl.noaa.gov/gmd/grad/solcalc/sunrise.html). The fraction of moon disc illuminated per night was obtained from the U.S. Naval Observatory website (http://aa.usno.navy.mil/data/docs/MoonFr action.php) and used to describe the lunar phase. Fraction of the moon disc was converted into radians (θ) assuming a lunar cycle of 29.5 days (0 and 2π radians correspond to a new moon, and π radians corresponds to a full moon). Trigonometric functions (cosine and sine) were included in the statistical models as explanatory variables to investigate possible effects of lunar phase on the departure and arrival of penguins to the colony (see deBruyn and Meeuwig, 2001, for details).
Statistical Analysis
To test whether time of arrival after sunset or departure before sunrise was influenced by moon phase, we used general linear mixed models. Following the periodic regression approach suggested by deBruyn and Meeuwig (2001), cosine(θ), sine(θ), cosine(2*θ), and sine(2*θ) transformations were included as fixed terms to model potential semilunar cycles (i.e., 2 peaks substantially unequal in amplitude per lunar month). A 3-level factor describing the breeding phenology of penguins (breeding: August-February; molting: March-April; and interbreeding: May-July; Reilly and Cullen, 1981, 1983; Salton et al., 2015) and a 2-level factor for the colony (Cullen Cove or Penguin Parade) were included as fixed terms. In addition, the interaction of the trigonometric functions and the colony factor was included to test the potential different effect of moon phase between colonies. As random factors, we included transponder identification (ID) of penguins and date because observations from the same individuals or nights are not independent. Response variables (time of arrival after sunset and time of departure before sunrise) were log transformed. We visually checked whether the assumptions of normally distributed and homogeneous residuals were fulfilled by inspection of histograms of residuals and plots of residuals against fitted values and covariates (both indicated no obvious deviations from these assumptions).
To investigate the potential effects of moon phase on the number of penguins crossing the beach, we ran generalized linear mixed models with negative binomial error distribution and log link function. Cosine(θ), sine(θ), cosine(2*θ), and sine(2*θ) transformations were included as fixed terms to model potential semilunar cycles on the number of penguins. A 2-level factor for the colony (Cullen Cove or Penguin Parade) and its interactions with trigonometric functions were included as fixed terms. We included year and month as random factors because of the huge interannual and seasonal variability in the number of penguins visiting the breeding colonies (see Suppl. Figure S2). Overdispersion was assessed by calculating the sum of squared Pearson residuals and dividing by the sample size minus number of parameters (i.e., 13 corresponding to 10 fixed terms, 2 random terms, and 1 negative binomial dispersion parameter; Zuur et al., 2013). The overdispersion parameters of the full models ranged between 0.90 and 0.94. Model validation included visual inspection of plots. We plotted residuals against fitted values and fixed and random terms (Zuur et al., 2013).
All statistical analyses were conducted in R version 3.0.3 (R Foundation for Statistical Computing, Vienna, Austria). Models were fitted using the lmer and glmer.nb functions of the R-package lmer4 (Bates et al., 2014). We obtained p values by using the R-package lmerTest (Kuznetsova et al., 2014). To achieve more reliable p values, we fitted the models using maximum likelihood (rather than restricted maximum likelihood). The significance of the full models compared to the null model comprising only the breeding factor and the random terms was established using likelihood ratio tests.
Results
The automated penguin monitoring system at Penguin Parade identified 885 individual penguins with 295,696 readings (159,222 arrivals and 136,474 departures) over 4830 nights. At Cullen Cove, the reading platform identified 1849 individuals and recorded 298,027 readings (158,875 arrivals and 139,152 departures) over 2899 nights.
On average, penguins arrived around 81 min after sunset and departed around 92 min before sunrise throughout the year. Negligible numbers of penguins were recorded during daylight hours (Figure 1a). Penguin arrivals and departures varied seasonally according to sunset and sunrise times, respectively. Thus, penguin real-time attendance was more than 2 h earlier around winter solstice (i.e., when night duration is longer) than around summer solstice (i.e., when night duration is shorter) (Figure 1a).
Overall, the full models explaining timing of arrival or departure as a function of moon cycle were highly significant compared to the null models (likelihood ratio tests: arrivals χ2 = 299.32, df = 8, p < 0.001; departures χ2 = 810.37, df = 8, p < 0.001).
Arrival and departure times showed semilunar cycles (i.e., terms including sine(2*θ) and cosine(2*θ) were significant), but semilunar cycles were described by different terms in each colony (see Suppl. Table S1). Interactions between colony and trigonometric functions reached high significance, indicating that moon phase has a different effect on both colonies (Suppl. Table S1). Despite these significant differences between colonies, the modeled attendance patterns were similar at the artificially illuminated Penguin Parade and Cullen Cove, with natural light conditions. Thus, penguins arrived later and departed earlier around full moon and last quarter nights than around first quarter nights at both colonies (Figure 2). The effect of moonlight, measured as the amplitude of the cyclic pattern (i.e., maximum minus minimum value), was stronger at departure than at arrival at both colonies. The average amplitude for arrivals was 12 min in comparison with the average 26 min for departures (Figure 2). In addition, the modeled cyclic pattern for arrivals showed similar amplitude at both colonies, despite Penguin Parade being artificially illuminated at arrivals (12.2 min at the artificially lit Penguin Parade and 11.8 min at the naturally lit Cullen Cove).
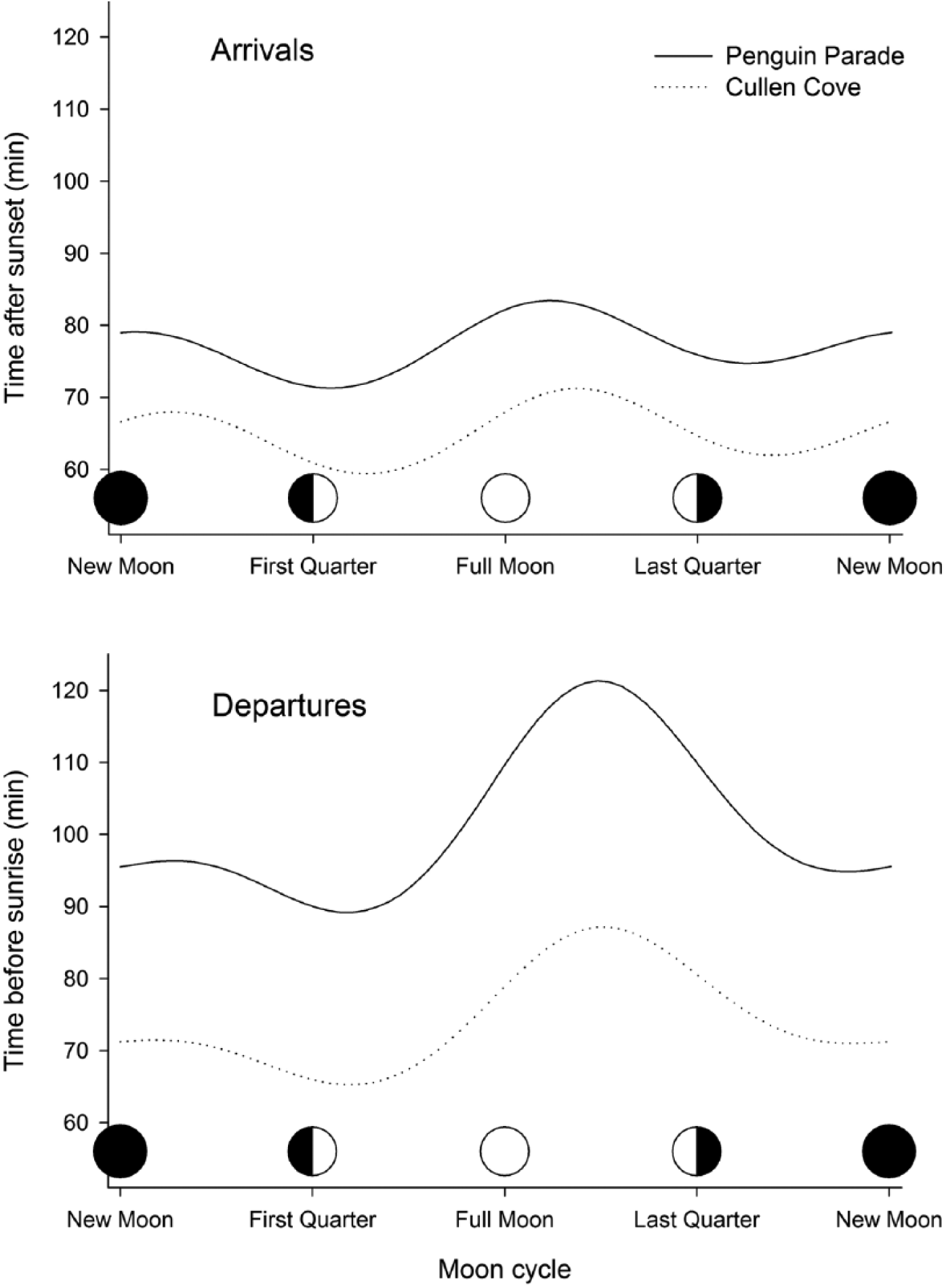
Arrival and departure timings of little penguins Eudyptula minor in relation to moon phase at 2 colonies (Penguin Parade and Cullen Cove) on Phillip Island, Australia. Cyclic effects were estimated by the inclusion of trigonometric functions of a circular variable (moon phase) in general linear mixed models (Suppl. Table S1). For better visualization, only the effects for the level “breeding” are displayed.
Overall, the full models explaining the number of penguins arriving at or departing from the colonies as a function of moon cycle were highly significant compared to the null models (likelihood ratio tests: arrivals χ2 = 36.44, df = 8, p < 0.001; departures χ2 = 44.79, df = 8, p < 0.001). The number of penguins arriving at or departing from the colonies showed clear lunar cycles described, with cosine(θ) being the only transformation reaching significance in both models (Suppl. Table S2). The minimum number of penguins was recorded during the full moon nights (Suppl. Table S2 and Figure 3).
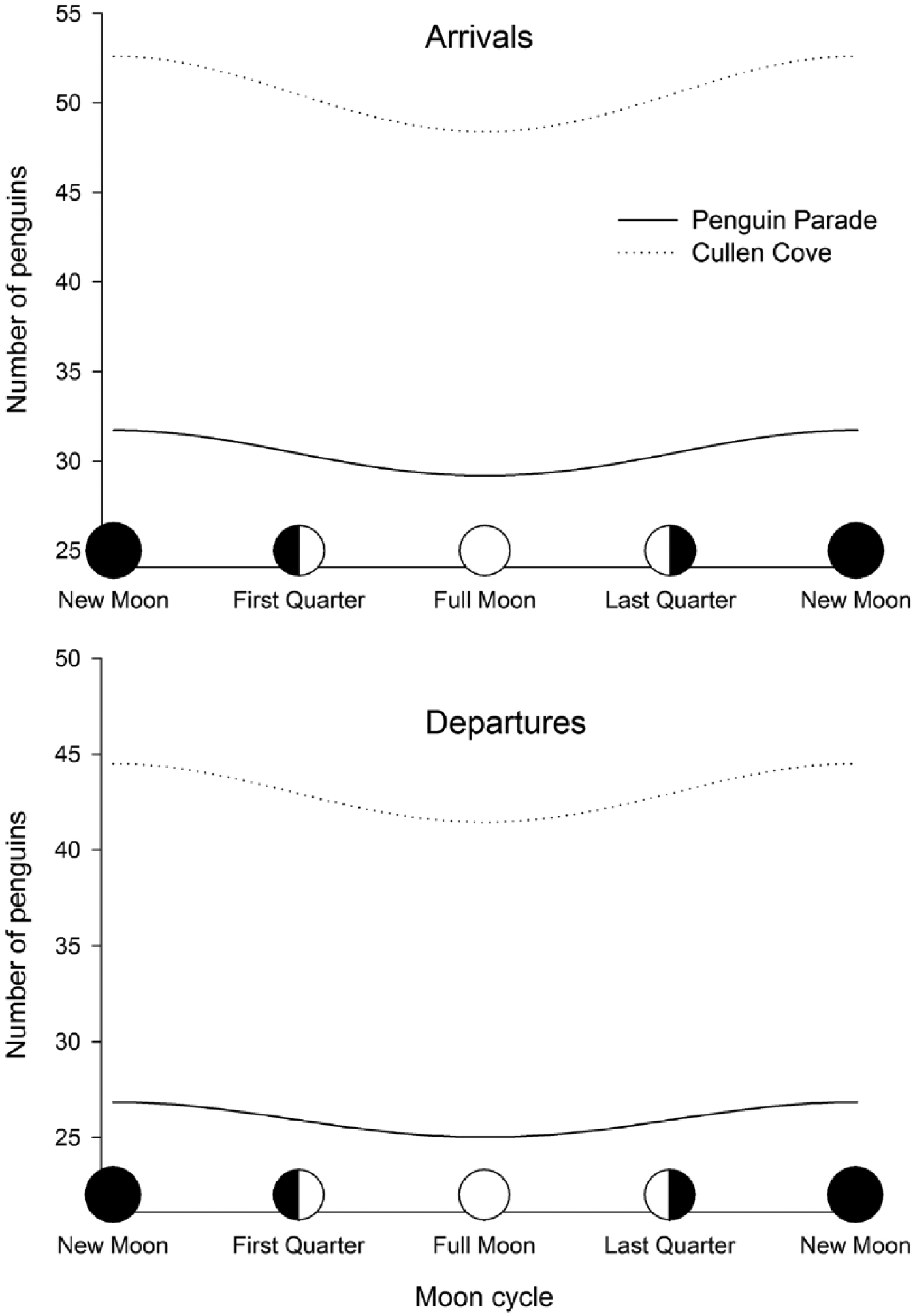
Mean number of little penguins Eudyptula minor coming ashore or going to the sea in relation to moon phase at 2 colonies (Penguin Parade and Cullen Cove) on Phillip Island, Australia. Cyclic effects were estimated by the inclusion of trigonometric functions of a circular variable (moon phase) in generalized linear mixed models (Suppl. Table S2).
Discussion
Despite a substantial part of the world’s biodiversity being nocturnal (around 30% of vertebrates and 60% of invertebrates; Hölker et al., 2010), chronobiologists have centered their attention on the effects of solar cycles (daily and annual) on biological rhythms, while the effects of lunar cycles mediated by moonlight have received much less attention (Kronfeld-Schor et al., 2013). In addition, our knowledge of moonlight effects is highly skewed between terrestrial and marine ecosystems. While terrestrial ecologists have centered on predator-prey relationships, marine ecologists have focused on behavior and reproduction (Kronfeld-Schor et al., 2013; but see, for example, Cruz et al., 2013). The increase in artificial lighting worldwide is an additional and growing challenge to the emerging field of night ecology. Here, we studied the effect of sunlight and moonlight on the attendance behavior of a nocturnal seabird on land, the little penguin, over 10 years in 2 colonies. We examined the attendance pattern of penguins in response to the natural light of the sun and moon. Penguin arrivals and departures depended primarily on the sunlight, as they covaried with sunset and sunrise times, respectively, throughout the annual cycle (Figure 1a; Klomp and Wooller, 1991). After removing the seasonal effects by standardizing arrival or departure times in relation to sunset and sunrise, respectively, we found a significant effect of moon phase on the attendance timing, which varied between colonies, although with similar cyclic patterns (Suppl. Table S1). Thus, penguins arrived later and departed earlier around full moon and last quarter nights than around first quarter nights at both colonies (Figure 2). The number of penguins attending both colonies was influenced by moonlight, with lower numbers around full moon nights.
Moonlight levels show a high variation due to phase and position of the moon in the sky (Austin et al., 1976; Suppl. Figure S3). As expected, the effect of the moonlight on departures was stronger than on arrivals (Figure 2). It is still daylight when penguins form groups in the water before coming ashore after sunset, so the sunlight has stronger influence on penguin behavior. During full moon and last quarter nights, moonlight levels are high in the 2 h before sunrise (departure). However, for the same period, moonlight levels are low in the 2 h after sunset (arrival). Although the range or amplitude of arrival times was less than half of departure times, the question on why penguins arrived later during full moon and last quarter nights (i.e., when nights were dark) deserves further studies.
Selective pressures of penguins on arrivals and departures from colonies are different and variable throughout the annual cycle. Thus, arriving birds come back to the land after spending daylight hours foraging at sea. Penguins arriving earlier do so probably for nest-burrow defense or maintenance, courtship, or thermoregulatory purposes (Davis and Renner, 2003). However, departing penguins could be more interested in starting a new foraging trip as soon as practicable, especially during breeding, when they must forage for nestlings and themselves. Indeed, our findings showed that penguins leave the colony earlier during breeding (Suppl. Table S1).
But why do penguins go to sea earlier during bright nights? Light level may well determine the foraging efficiency of visual predators, especially for pursuit-diving seabirds as coefficient extinction of light is higher in the water than in the air. Depth and frequency of diving are reduced during poor light conditions in other seabirds (Wanless et al., 1999; Regular et al., 2011). Little penguins are visual predators, which forage during the day, and have never been recorded diving and foraging at night (Preston et al., 2008; Pelletier et al., 2014), suggesting that light levels at night are too low for fishing (Cannell and Cullen, 1998). Thus, penguins could go to the sea earlier during bright nights aiming to improve their foraging efficiency by arriving earlier at their preferred foraging areas.
Unlike migratory animals, little penguins are present throughout the year at their colonies, although the numbers are higher during the summer months, coinciding with the breeding season (Reilly and Cullen, 1981; Salton et al., 2015), and lower during the interbreeding season or in bad years, when they have to make longer foraging trips (Chiaradia and Nisbet, 2006; Kato et al., 2008; Saraux et al., 2011; Zimmer et al., 2011). Although we did not include these sources of variation, we detected significant effects of moon phase on the number of penguins coming ashore. Penguins show a more predictable time attendance pattern than other nocturnally active seabirds on land (e.g., petrels or murrelets; Nelson and Peck, 1995; Warham, 1996). While petrels adjust their colony arrivals at times when the moon is not in the night sky by rafting nearby (Keitt et al., 2004; Bourgeois et al., 2008; Rubolini et al., 2014), penguins come ashore almost at the same time every night (although they also congregate offshore before sunset, probably waiting for lower light levels). In this sense, the attendance behavior of penguins is more similar to the commuting behavior of bats, although with differences. Bats depart from colonies after sunset and arrive before sunrise.
It is known that weather conditions, especially fog, can delay the arrival of little penguins (Chiaradia et al., 2007; but see Klomp and Wooller, 1991), as well as in other nocturnal seabirds (e.g., murrelets; Nelson and Peck, 1995). In our study, we did not take into account the weather conditions because our long-term data set includes different weather conditions throughout the moon phases, and it is difficult to obtain accurate weather information at the precise arrival and departure times. Tides could also affect the time of arrivals detected by the automated monitoring penguin system as during low tides penguins must traverse longer distances between the water’s edge and the monitoring system (Laaksonen, 2011). Thus, during nights between full moon and last quarter when the tide is low after sunset, we would expect to find delays in the arrival times (Laaksonen, 2011). However, tide is also low during nights between new moon and first quarter after sunset, and delays in arrival times were not recorded. Therefore, the pattern found in our study is independent of tide levels or it is masked by other variables.
Artificial Illumination
Moonlight had a different effect on the arrival and departure timings at both colonies, which was supported by significant interactions between trigonometric functions and colony (Suppl. Table S1), although timing of attendance of both colonies showed similar oscillatory patterns. The maximum and minimum values of time after sunset or before sunrise were reached at the same moon phases, that is, shortly after full moon and after the first quarter, respectively (Figure 2).
Artificial lights could override the maximum luminance of the full moon, and thus high levels of light pollution could mask some of the behaviors modulated by moonlight (Kronfeld-Schor et al., 2013). In our study, artificial lights at Penguin Parade produce an illuminance of 3 lux on the beach, which is higher than the maximum moonlight illuminance (around 0.215 lux; Austin et al., 1976). Consequently, we predicted that the moonlight effect should be weaker in the artificially lit colony since it would be overridden by artificial light. Contrary to our predictions, we found a similar amplitude in the modeled cyclic arrival pattern at both colonies, which may indicate that artificial illumination, at least at the levels employed in our study, does not mask the moon effect on the arrival attendance behavior.
Artificial illumination has been used at the Penguin Parade colony since the 1980s. Given the high philopatry of penguins to breeding colonies, many of the birds have been crossing a lit environment since fledging. Thus, habituation of penguins to artificial lights could explain lack of response in the predicted direction. In this sense, results of a preliminary study conducted at the Penguin Parade indicated that attendance behavior of penguins, measured as number of penguins coming ashore, velocity, group size, and timing, was not affected when artificial illumination increased up to 15 lux using the same type of lights (A.R., unpublished data).
Given that 1) site-specific differences and presence/absence of artificial illumination at both colonies are inseparable, 2) the potential complexity of the response to the interaction between artificial light and moonlight, and 3) the probable habituation of penguin to artificial lights, we cannot completely rule out an effect of artificial illumination on penguin behavior. However, we believe that the different effect of moon on both colonies could be site specific. The contrasting beach substrate and length of beach to be walked by penguins at the 2 sites could explain the different response to moonlight on both colonies. If there was an effect of the artificial light, we would expect a much stronger signal as reported in other seabirds that are fatally attracted to artificial lights (Rodríguez et al., 2014). The effect of artificial lights on wildlife is an emergent field, and further research is indeed necessary to understand how seabirds and other nocturnal species respond to ever increasing artificial night light levels.
Footnotes
Acknowledgements
We are grateful to Phillip Island Nature Parks: the rangers for their assistance in observational data collection and the research department for their continued support, particularly the late Mike Cullen, for his pioneering thinking on penguin attendance patterns at Phillip Island. We thank Ruth Martínez for her help in handling the database and the Australian Antarctic Division for kindly donating the automated penguin monitoring system (APMS), particularly Knowles Kerry and Kym Newbery. The APMS data collection and maintenance over the years were only possible thanks to many field assistants and electronic engineers (in chronological order since 1994): Brad Philips, Judy Clarke, Sarah Robinson-Laverick, Julija Yorke, Tom Daniel, Peter Knerborne, Marcus Salton, Julie McInnes, Rohan Long, Sandi Laaksonen, Hugh Davies, Darren Steirt, Matt Simpson, Claire Saraux, Laure Pelletier, Kean Maizels, and Danielle Hedger. Two anonymous reviewers and the editor William J. Schwartz provided constructive comments on early drafts. This work received grants and donations from BHP-Billiton, AAT Kings, Penguin Foundation, and Australian Academy of Science. A.R. was supported by a Marie Curie International Outgoing Fellowship within the 7th European Community Framework Programme (330655 FP7-PEOPLE-2012-IOF).
Conflict of Interest Statement
The author(s) have no potential conflicts of interest with respect to the research, authorship, or publication of this article.
Notes
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.