
Abstract
Most organisms in temperate or tropic regions employ the light-dark (LD) cycle as the primary Zeitgeber to synchronize circadian rhythms. At higher latitudes (>66°33′), continuous illumination during the summer presents a significant time-keeping dilemma for polar-adapted species. Lapland longspurs (Calcarius lapponicus), arctic-breeding migratory songbirds, are one of the few recorded species maintaining an intact diel rhythm in activity and plasma melatonin titers during polar summer. However, it is unknown whether rhythms are endogenous and entrain to low-amplitude polar Zeitgeber signals, such as daily variations in light intensity and the spectral composition of the sun (as measured by color temperature). Wild-caught male and female longspurs were brought into captivity, and locomotor activity was assessed using infrared detection. To examine if rhythms were endogenous, birds were exposed to constant bright light (LL; 1300 lux) or constant darkness (DD; 0.1 lux). All birds exhibited free-running activity rhythms in LL and DD, suggesting the presence of a functional circadian clock. Mean periods in LL (22.86 h) were significantly shorter than those in DD (23.5 h), in accordance with Aschoff’s rule. No birds entrained to diel changes in light intensity, color temperature, or both. To examine endogenous molecular clock function, the Per2 gene was partially cloned in longspurs (llPer2) and transcripts were measured in hypothalamic tissue punches, eye, and liver using competitive polymerase chain reaction. Ocular llPer2 gene expression was periodic in LL and elevated at ZT24 (CT24) for LD or constant conditions (LL and DD), but llPer2 rhythmicity was not detected in hypothalamus or liver. Plasma melatonin was significantly lower in LL compared with LD or DD. In conclusion, rhythmic ocular Per2 expression and melatonin secretion may maintain the circadian activity rhythm across the polar day.
The primary environmental cue, or Zeitgeber, that synchronizes daily rhythms of most tropical- and temperate-dwelling organisms is the solar light-dark (LD) cycle (Pittendrigh, 1981; Aschoff, 1989). In Antarctica or north of the Arctic Circle (>66°33′ latitude), this Zeitgeber is noticeably lacking for much of the summer and winter months, where continuous light (polar day) or darkness (polar night), respectively, prevails (Pielou, 1994). The seasonal absence of the LD cycle poses a unique dilemma to the regulation of circadian rhythms among polar-adapted organisms.
Several adaptations of the biological clock have evolved to cope with continuous polar day or night. Some species, such as reindeer (Rangifer tarandus) and ptarmigan (Lagopus mutus), completely abandon circadian organization during winter and summer and resort to ultradian rhythms (Stokkan et al., 1994; Eloranta et al., 1995; Reierth and Stokkan, 1998; van Oort et al., 2005; Bloch et al., 2013). Among high-arctic shorebirds, there is a diversity of time-keeping approaches depending on species, sex, and breeding stage. During the preincubation stage, birds are arrhythmic but switch at incubation to either a free-running rhythm that is synchronized among pairs (semipalmated sandpiper [Calidris pusilla]), a robust 24-h rhythm (caregiving sex of uniparental shorebirds), or remain arrhythmic (competitive sex of polygamous shorebirds; Steiger et al., 2013). Lastly, several taxa, such as migratory songbirds (Hau et al., 2002; Silverin et al., 2009; Ashley et al., 2013; Steiger et al., 2013), arctic ground squirrels (Uroceitelllus parryii; Swade and Pittendrigh, 1967; Long et al., 2005; Williams et al., 2012), and bumblebees (Bombus spp.; Stelzer and Chittka, 2010), exhibit consistent intact rhythms in behavior and physiology throughout the polar summer. These observations suggest entrainment to alternative environment cues that may include subtle daily variations in light intensity, spectral composition of light, or ambient temperature, but experimental evidence is lacking (Swade and Pittendrigh, 1967; Krüll, 1985; Pohl, 1999). Presumably, persistence of diel rhythms is necessary in these particular species to capitalize upon diel fluctuations in food supply, predators, and/or thermoregulatory requirements.
In mammals, the hypothalamic suprachiasmatic nucleus (SCN) serves as the major pacemaker that regulates daily cycles in physiology and behavior (Buhr and Takahashi, 2013). However, in birds, the pineal gland, the retinae of the eyes, and hypothalamus act as independent oscillators that are mutually coupled (Gwinner and Brandstätter, 2001; Cassone, 2014). Within the hypothalamus, 2 nuclei are involved with circadian organization in birds: the medial suprachiasmatic nucleus (mSCN) and visual suprachiasmatic nucleus (vSCN; Cassone and Moore, 1987; Yoshimura et al., 2001). On a molecular level, these oscillators are controlled by a highly conserved mechanism that involves rhythmic transcription and translation of clock genes. The major components of the clock machinery involve the interplay between positive elements clock and bmal1 and negative elements Period 1 (Per1), Per2, Per3, and cryptochrome 1 (cry1) and cry2 (Tei et al., 1997; Zheng et al., 1999a; Zheng et al., 1999b; Yoshimura et al., 2000; Yasuo et al., 2002; Yasuo et al., 2004a; Yasuo et al., 2004b; Helfer et al., 2006). Per1 expression is absent in birds (Yoshimura et al., 2000; Yasuo et al., 2002). Avian homologues of Per2 have been cloned in a variety of bird species (e.g., Japanese Quail [Coturnix coturnix japonica], chicken [Gallus domesticus], house sparrow [Passer domesticus], blue tit [Cyanistes caeruleus], and redheaded bunting [Emberiza bruniceps]; Yoshimura et al., 2000; Brandstätter et al., 2001; Doi et al., 2001; Steinmeyer et al., 2012; Singh et al., 2013). Circadian expression of Per2 has been demonstrated in the eye and pineal gland (Yoshimura et al., 2000; Doi et al., 2001). In addition, several clock genes are expressed rhythmically in the mSCN of quail (Yasuo et al., 2002), whereas house sparrows generate independent pPer2 rhythms in the mSCN and vSCN (Abraham et al., 2002; Abraham et al., 2003).
Lapland longspurs (Calcarius lapponicus), songbirds that migrate to Alaska and northern Canada to breed every year from wintering areas in temperate regions, maintain diel rhythms in activity during polar day with a quiescence period of only 4 to 5 h (Ashley et al., 2013; Steiger et al., 2013). Plasma melatonin is also rhythmic (Hau et al., 2002; Ashley et al., 2013) and negatively correlated with daily fluctuations in light intensity and ambient temperature (Ashley et al., 2013). The spectral composition of sunlight varies over the polar day (measured by color temperature [K]; Pohl, 1999), and birds possess photoreceptors that are sensitive to differing wavelengths of light (Hart, 2001). Thus, it is possible that diel fluctuations in the spectral qualities of sunlight could synchronize rhythms. At this juncture, it is unclear whether these low-amplitude photic and thermal cues act as alternative zeitgeber signals (Swade and Pittendrigh, 1967; Krüll, 1976b, 1976c; Pohl, 1999) or simply mask overt activity rhythms (Aschoff and von Goetz, 1989).
The aims of this study were 3-fold. We sought to determine (1) whether longspurs maintain an endogenous circadian rhythm when exposed to constant light (LL) or darkness (DD) compared with LD, (2) whether manipulations of light intensity (quantity) and color temperature (quality) can induce entrainment, and (3) whether circadian clock gene expression, specifically Per2, exhibits rhythmicity similar to patterns observed in house sparrows (Brandstätter et al., 2001; Abraham et al., 2002; Helfer et al., 2006) and other avian species (Yoshimura et al., 2000; Doi et al., 2001; Singh et al., 2013). Per2 mRNA levels peak early in the day and are light sensitive (Abraham et al., 2002; Yasuo et al., 2003; Helfer et al., 2006; Singh et al., 2013). We hypothesized that longspurs would possess a functioning circadian clock and therefore would display free-running rhythms in LL and DD compared with LD. We further predicted that experimental alterations in light intensity and color temperature would entrain behavioral rhythms and that Per2 expression in hypothalamus, eye, and liver would exhibit rhythmicity and specifically increase during subjective day in LD and constant conditions.
Materials and Methods
Animals
Twenty-eight male and 20 female Lapland longspurs were captured in Barrow, Alaska (71°N, 156°W) in the summer of 2009 (for study site details, see Ashley et al., 2013) and held in temporary indoor aviaries before transport by plane to the University of Alaska Anchorage (UAA; 61°N, 150°W) in modified pet carriers on July 1. Birds were then housed indoors within the UAA vivarium in 2 indoor flight aviaries on a light cycle representative of short day lengths (8L:16D) at a temperature of 20 °C ± 1 °C. Birds were provided with food (bird seed mix) and water ad libitum and given weekly protein supplements of moistened egg food (Quiko) and live mealworms. Procedures adhered to the National Institutes of Health Guide for the Care and Use of Laboratory Animals and were approved by the UAA Institutional Animal Care and Use Committee, as well as municipal, state, tribal, and federal authorities.
Experimental Settings
After birds finished their prebasic molt (October 3), subjects were randomly assigned to 1 of 3 rooms of identical dimensions that contained individual bird cages (36.2 × 41.3 × 43.8 cm) on a stainless-steel rack (4 cages per row). After random assignment, we attempted to balance sex ratio across each group: room 1 (9 males, 7 females), room 2 (9 males, 7 females), room 3 (10 males, 6 females). To provide a measure of general activity, each cage was outfitted with infrared motion detectors (model PK IS215T; Honeywell, Morristown, NJ) to measure beam breaks. Each detector was placed about 10 cm above the upper perch of the cage. The detector produced a downward, cone-shaped beam that permitted detection of movement within all areas of the cage. To eliminate interference with other detectors, cardboard dividers were placed between adjacent cages. This manipulation also visually isolated birds from each other. Each detector was wired to an interface box that could accommodate up to 7 different channels; each interface box (7 total) was connected to a 56-channel hub directly plugged into a PC computer in an outer room running ClockLab (Actimetrics, Wilmette, IL). The number of beam breaks was recorded for each channel and binned into 5-min intervals.
Lighting
To ensure that birds were exposed to similar lighting conditions and to manipulate light intensity and color temperature, each room was outfitted with custom-built light sources. On a separate metal rack, 224-cm (8-ft) fluorescent bulbs (32 W, 6500 K color temperature; Sylvania, Danvers, MA) were mounted in quadruplet vertically. In total, there were 3 quadruplets of bulbs spaced evenly. Within each quadruplet, only 2 identical bulbs were turned on at once. This arrangement allowed for instantaneous switching between different light sources that differed in light intensity, color temperature, or both. We quantified light intensity (lux) using a light meter at each bird’s cage, and the distance from the light source to the cages was standardized such that each cage received about 1300 lux (subjective “day” light setting). This value approximates the lower threshold of light intensities recorded in polar summer (Arendt, 2012; Ashley et al., 2013). Light schedules (timing of on/off) were controlled by computer using Clocklab’s integrated light control module. A separate “night” light that emitted only blue light was continuously on (plugged into an outlet on the opposing wall relative to birds); dim blue light is often employed in avian studies to prevent photostimulation because its shorter wavelengths fail to penetrate the avian skull to activate deep encephalic photoreceptors compared with the longer wavelengths of red light (Oishi and Lauber, 1973). When fluorescent lights were switched off (referred to as “dark”), the light intensity was about 0.1 lux.
Experiments
Experiment 1
From October 3 to 16, birds were exposed to a 16L:8D photoperiod with lights-on at 0700 h. This photoperiod was chosen to acclimate birds to long days before continuous exposure to LL. All birds entrained to 16L:8D. To examine activity rhythms of birds to constant conditions (LL and DD) relative to LD, birds were exposed to the same order of light treatments—(1) LL (October 16-22), (2) 12L:12D (October 22-27), (3) DD (October 27-29)—instead of counterbalancing treatments across rooms. This ensured that birds from separate rooms were not differentially photostimulated, which could have an effect on rhythms when the experiment was repeated (see below). Activity rhythms were recorded throughout these periods. The LD treatment (lights-on at 0700 h) was deliberately placed after the LL treatment to reentrain birds that displayed free-running rhythms. The DD treatment had to be shortened to 3 d because some birds were losing body mass because little feeding occurred in darkness. Activity onsets and offsets were estimated using the automated detection function in Clocklab. Free-running periods were calculated by periodogram analysis using Clocklab software for ≥3 d of light treatment.
Experiment 2
From October 27 to November 10, a separate set of experiments was conducted to examine whether alterations in either light intensity or color temperature were sufficient to entrain these birds. For these studies, birds were placed in 3 d of DD to induce free-running rhythms and then exposed to various light exposures. In room 1, we tested whether a 55% reduction in light intensity could lead to entrainment. To manipulate light intensity without altering color temperature, a 2-stop 0.5 neutral density filter was used (Rosco, Stamford, CT) to reduce light intensity by 55%. In this group, birds received 12 h of 780 lux (reduced from 1300 lux by disconnecting 1 of 2 lights per quadruplet) followed by 12 h of 350 lux (same bulbs with filter). This latter value represents lower light intensity that falls just below minimum light intensities recorded in the field in Barrow, Alaska, during polar summer (Ashley et al., 2013). Color temperature remained the same (6500 K) and was verified by a precision colormeter (Cal-COLOR 400; Cooke Corp., Romulus, MI). In room 2, color temperature was manipulated without altering light intensity by using bulbs with the same wattage. In this group, birds received 12 h of 6500 K at 780 lux (blue light) and then 12 h of 3500 K at 780 lux (red light). Again, this alteration in spectral composition is similar to the range of natural light spectra encountered in northern Alaska (Ashley et al., 2013). In room 3, birds were exposed to 12 h L (780 lux, 6500 K) and then 12 h D. After 6 d (November 5), all birds were exposed to DD for 3 d.
To investigate whether simultaneous reductions in light intensity and color temperature entrained activity rhythms, birds in rooms 2 and 3 were exposed to 12 h of 780 lux at 6500 K and then 12 h of 350 lux at 3500 K for 3 d (November 7-10). Birds in room 1 received the control treatment of 12 h of L (1300 lux, 6500 K) followed by 12 h of darkness for 3 d (Fig. 1).
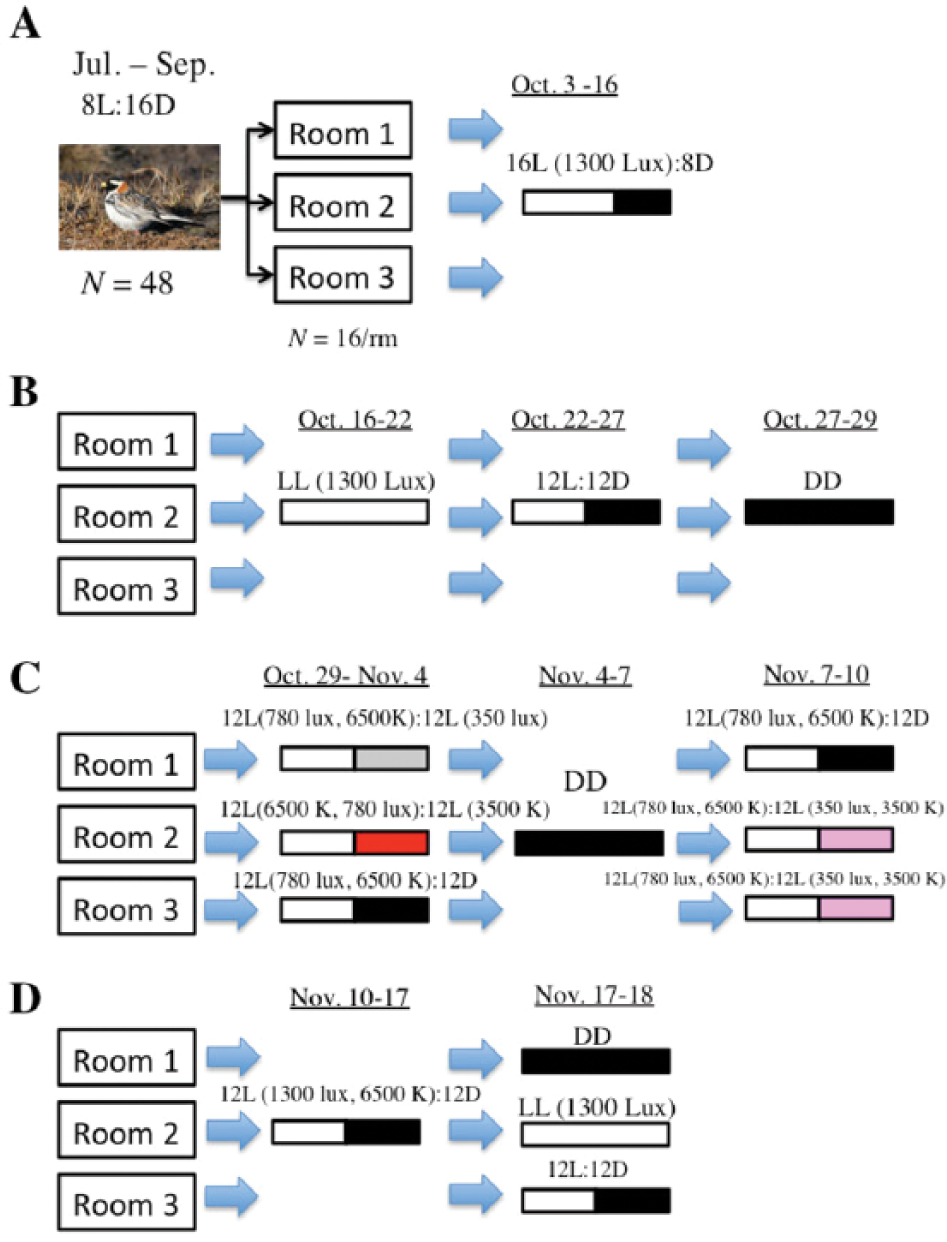
Experimental time line. (A) Male and female longspurs were maintained on a nonbreeding photoperiod of 8L:16D for approximately 3 mo and then transferred to individual cages. Birds were exposed to 16L:8D for 2 wk. (B) Longspurs were sequentially exposed to LL (1300 lux, 6500 K), 12L (1300 lux, 6500 K):12D, and DD treatments and activity rhythms were measured. (C) Light intensity (lux) or color temperature (K) were reduced by 55% and 46%, respectively, for the second 12 h of the daily 24-h cycle, and entrainment of activity rhythms was assessed: group 1, 12L (780 lux, 6500 K, white):12L (350 lux, 6500 K; gray); group 2, 12L (780 lux, 6500 K):12L (780 lux, 3500 K; red). These treatments followed exposure to DD (0.1 lux) for 3 d, which typically induced free-running rhythms. An additional manipulation that involved simultaneous reductions in light intensity and color temperature was conducted on 2 groups of longspurs to assess entrainment of rhythms: groups 2 and 3, 12L (780 lux, 6500 K):12L (350 lux, 3500 K; pink). Throughout these experiments, a third group was exposed to 12L:12D to serve as a control. (D) Birds were entrained to 12L:12D; exposed to either LD, DD, or LL; and then euthanized at 4 different time points to assess Per2 expression in hypothalamus, eye, and liver.
Experiment 3
To examine patterns of Per2 gene expression in the brain, eye, and liver in relation to LD, LL, and LD groupings, birds were first exposed to 12L (1300 lux, 6500 K):12D for 6 d (November 11-18; Fig. 1). Phasing of entrainment was staggered across rooms such that lights turned on in rooms 1, 2, and 3 at 1000, 0900, and 1100 h, respectively, and all birds synchronized activity rhythms. A staggered design prevented tissue collection from occurring at the same time in all rooms. On November 17, rooms 1, 2, and 3 were switched to DD, LL, and LD at 1000, 0900, and 1100, respectively. We followed a similar sampling protocol used to examine pper2 expression in house sparrows that involved sampling of tissue over a 48-h period (Abraham et al., 2002). Starting at circadian time (CT) 6 or zeitgeber time (ZT) 6 (midday), 2 Lapland longspurs were taken from their cage and blood samples were obtained (see specific methods below). Birds were then immediately exposed to isoflurane vapors using a vaporizer (5% induction) for <1 min and then rapidly decapitated. Brain, eyes, and liver were extracted and immediately placed on dry ice and stored at −80 °C for further analysis. This sampling protocol continued at CT12 (ZT12, just after lights-off), CT18 (ZT18), and CT24 (ZT24, just after lights-on). A second series consisting of 2 longspurs were killed from each room at the same time points during the next 24-h cycle. Although it was anticipated that DD and LL birds would free-run slightly from 24 h as reported, this deviation is minor (see the Results section; at least for the second 24 h), and thus time points from both series were combined to yield a sample size of 4 at each time point (6 h, 12 h, 18, 24/0 h; Abraham et al., 2002). Sampling during dark periods occurred using only dim, blue light (Oishi and Lauber, 1973). One hypothalamus sample was lost in the LD, ZT24 group.
Blood Collection
Blood samples (about 200-300 µL) were obtained from the alar wing vein using a 26-gauge needle and collected into heparinized microcapillary tubes. These samples were stored for <30 min on ice. Blood was separated by centrifugation for 5 min, and the plasma fraction was collected with a Hamilton syringe and stored at −20 °C for later determination of melatonin concentration (see below).
Micropunch Dissection
Brains were cut into 20-µm coronal sections with a cryostat at −20 °C. Using anatomical markers from the canary brain atlas (Stokes et al., 1974), tissue punches of the hypothalamic region (starting at the tractus septomesencephalicus) of alternate sections were collected using a 4-mm-diameter punch and then stored in Trizol (Life Technologies, Carlsbad, CA).
Cloning of Per2 in Lapland Longspurs
Eyes from Lapland longspurs collected in 2008 were used for cloning a partial Per2 gene sequence in this species. Primers were designed based on the pPer2 sequence of the house sparrow (Passer domesticus; GenBank accession number AY007259). Subcloning of polymerase chain reaction (PCR) products was performed using pGEM-T Easy Vectors (Promega, Madison, WI) according to the manufacturer’s instructions, and DNA inserts of the positive clones were amplified with PCR using M13 primers. Sequencing of Per2 occurred at the UC Berkeley DNA Sequencing Facility (Berkeley, CA) using an Applied Biosystems 3730 DNA sequencer and is reported in GenBank (336 bp; accession number FJ695136.1). The sequence is 98% homologous to the house sparrow pPer2 sequence.
Competitive PCR Analysis
To quantify mRNA encoding the per2 polypeptide, a competitive PCR analysis was performed according to methods outlined in Ubuka et al. (2003, 2005). Briefly, total RNA from liver, spleen, and brain punches was isolated using phenol/chloroform extraction and then quantified using Nanodrop. Total RNA was reverse transcribed using Moloney murine leukemia virus transcriptase (Life Technologies). Oligonucleotides used for product amplification of longspur Per2 precursor polypeptide were as follows: forward primer, 5′-ATTCCGGATGACTCCCTACC-3′, and reverse primer, 5′-AAAAGGCTGCCCTCCATACT-3′, to yield a 316-bp product. Competitor DNA was synthesized by PCR using cDNA generated from the eye using the same forward primer as above, but a different reverse primer was used: reverse primer: 5′-
Radioimmunoassay
Plasma concentrations of melatonin were determined by direct radioimmunoassay after extraction following Goymann et al. (2008). A 4-parameter logistic curve was derived using Immunofit 3.0 (Beckman, Inc., Fullerton, CA) to calculate sample concentration. Samples were run in 2 separate assays, and the lower detection limit was determined as the first value outside the 95% confidence intervals for the zero standard (Bmax) and was 8.2 and 7.2 pg/mL, respectively. The intra-assay coefficients of variation (CVs) of melatonin standards were 8.8% and 1.5%. The intraextraction CVs of plasma pool samples ran in both assays were 3.3% and 4.4%. The interassay (standard melatonin) and interextraction CVs were 8.6% and 0.9%, respectively. Melatonin concentrations were adjusted for individual extraction recoveries (mean ± SD, 80.0% ± 0.44%).
Statistics
For experiment 1, circadian periods (τ) of LL and DD longspurs were compared using repeated-measures analysis of variance (ANOVA) with lighting treatment (LL to DD) as the repeated measure and sex and room as main factors. Because neither sex nor room was a significant factor in the above analysis (see the Results section), they were not assessed further. For experiment 2, Fisher exact tests were used to assess the percentage of longspurs that entrained to different light treatments compared with LD. For experiment 3, temporal variation of Per2 expression in eye, liver, and hypothalamus and melatonin concentration in plasma were examined, and values were logarithmically transformed to improve normality. Two-way ANOVAs were conducted with light treatment (LL, DD, and LD) and sampling time (6, 12, 16, 24/0 h) as main factors to examine differences among time points (Refinetti et al., 2007). For post hoc assessment, Fisher PLSD test was employed to assess within- and between-treatment comparisons. To assess underlying patterns of rhythmicity, harmonic regression using CircWave (v.1.4 software; R. Hut: http://webpage2.woelmuis.nl/downloads.htm) was employed. This method fits data collected from different individuals to a fundamental sinusoidal wave, which is then modified by harmonic input. Significance was designated at p < 0.05.
Results
Experiment 1
All longspurs entrained to 12L:12D. The mean circadian period (τ) of male and female longspurs exposed to either LL or DD was less than 24 h (LL: males, 22.93 ± 0.20 h; females, 22.76 ± 0.15; DD: males, 23.74 ± 0.37 h; females, 23.17 ± 0.37 h; Fig. 2). When shifted to DD, 31 of 48 longspurs exhibited an increased τ relative to previous LL exposure, and, on average, this increase was significant in the overall repeated-measures ANOVA (F1,42 = 6.54, p = 0.01; Fig. 3A, B). No other significant effects or interactions were detected, including those involving sex or room (all p’s > 0.23).
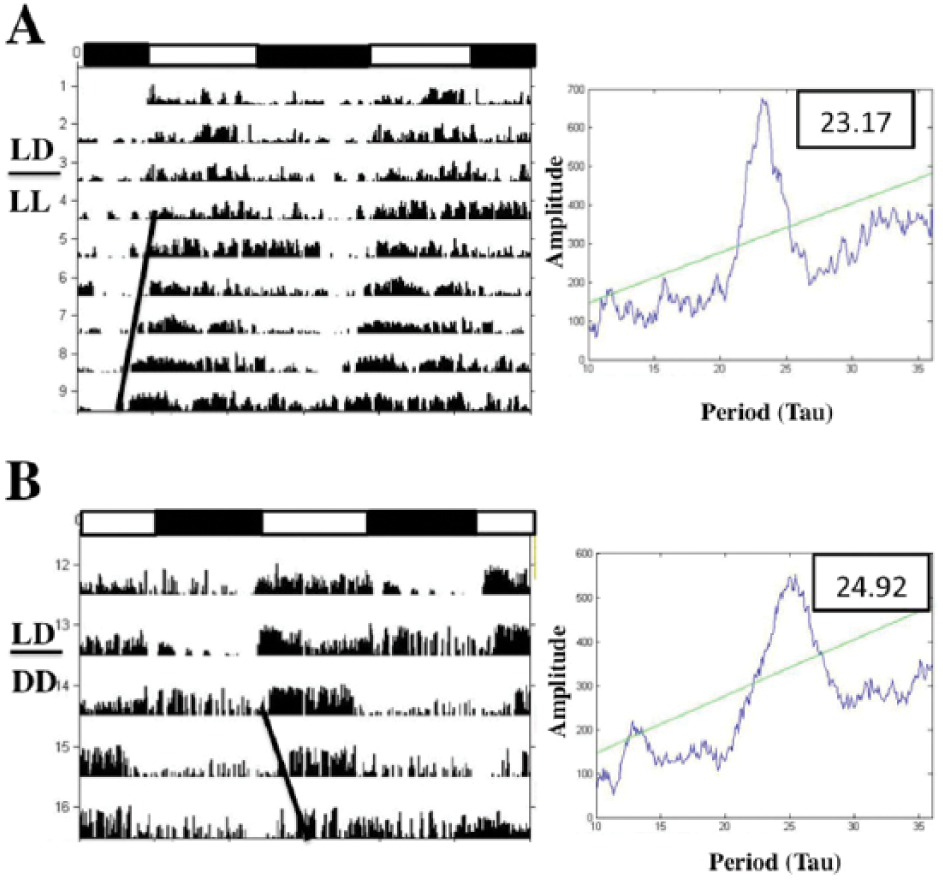
Representative actograms (double-plotted) and chi-squared periodograms of activity rhythms of longspurs exhibiting (A) entrainment to 12L:12D or transition to a free-running rhythm upon exposure to LL (B) and DD (C). Horizontal bars represent light-dark schedule (black depicts dark phase). Slope of bolded black line indicates free-running period (τ).
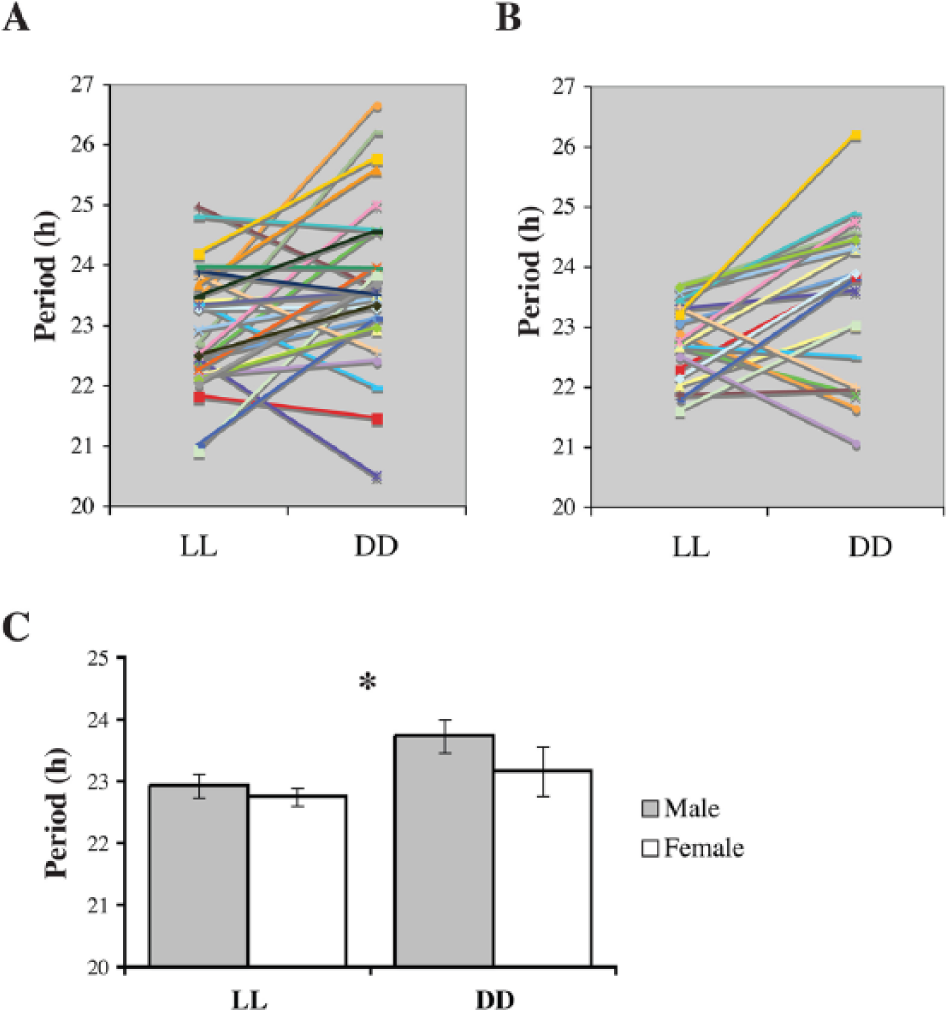
Free-running periods of locomotor activity for (A) male (n = 28) and (B) female (n = 20) longspurs exposed to LL and then DD. Different colors and symbols indicate individual longspurs. (C) Mean ± SEM values are depicted. Asterisk denotes a significant effect (p < 0.05) of lighting treatment (LL vs. DD) upon τ (irrespective of sex).
Experiment 2
Longspurs exposed to 12L (780 lux, 6500 K):12D following DD treatment exhibited entrainment of activity rhythms (Fig. 4A). Changes in light intensity, color temperature, or a combination of both failed to induce entrainment in longspurs. Specifically, all of 16 longspurs exhibited a free-running rhythm or had no discernable rhythm when exposed to daily variations in light intensity (12L [780 lux, 6500 K]:12L [350 lux, 6500 K]) or color temperature (12L [780 lux, 6500 K]:12L [780 lux, 3500 K]; Fig 4B, C). More specifically, in response to the light intensity alterations, 7 longspurs exhibited free-running rhythms (τ: 23.01 ± 0.45), whereas periodogram analysis failed to detect a rhythm in 9 birds. A similar pattern was observed in relation to the color temperature treatment; 5 birds had free-running rhythms (τ: 22.74 ± 0.39) whereas 11 displayed no detectable rhythm. In addition, none of the 32 birds entrained to simultaneous shifts in light intensity and color temperature (12L [780 lux, 6500 K]:12L [350 lux, 3500 K]; 10 birds free-ran [τ: 23.45 ± 0.21] whereas 22 birds were arrhythmic). Free-running rhythms in experiment 2 were not significantly different from those rhythms measured in experiment 1 (paired t-tests, all p’s > 0.17). Given the results, the response of longspurs to light intensity or color temperature treatments (free-run or arrhythmia) differed significantly from those exposed to LD (entrainment; Fisher exact tests, all p < 0.0001).
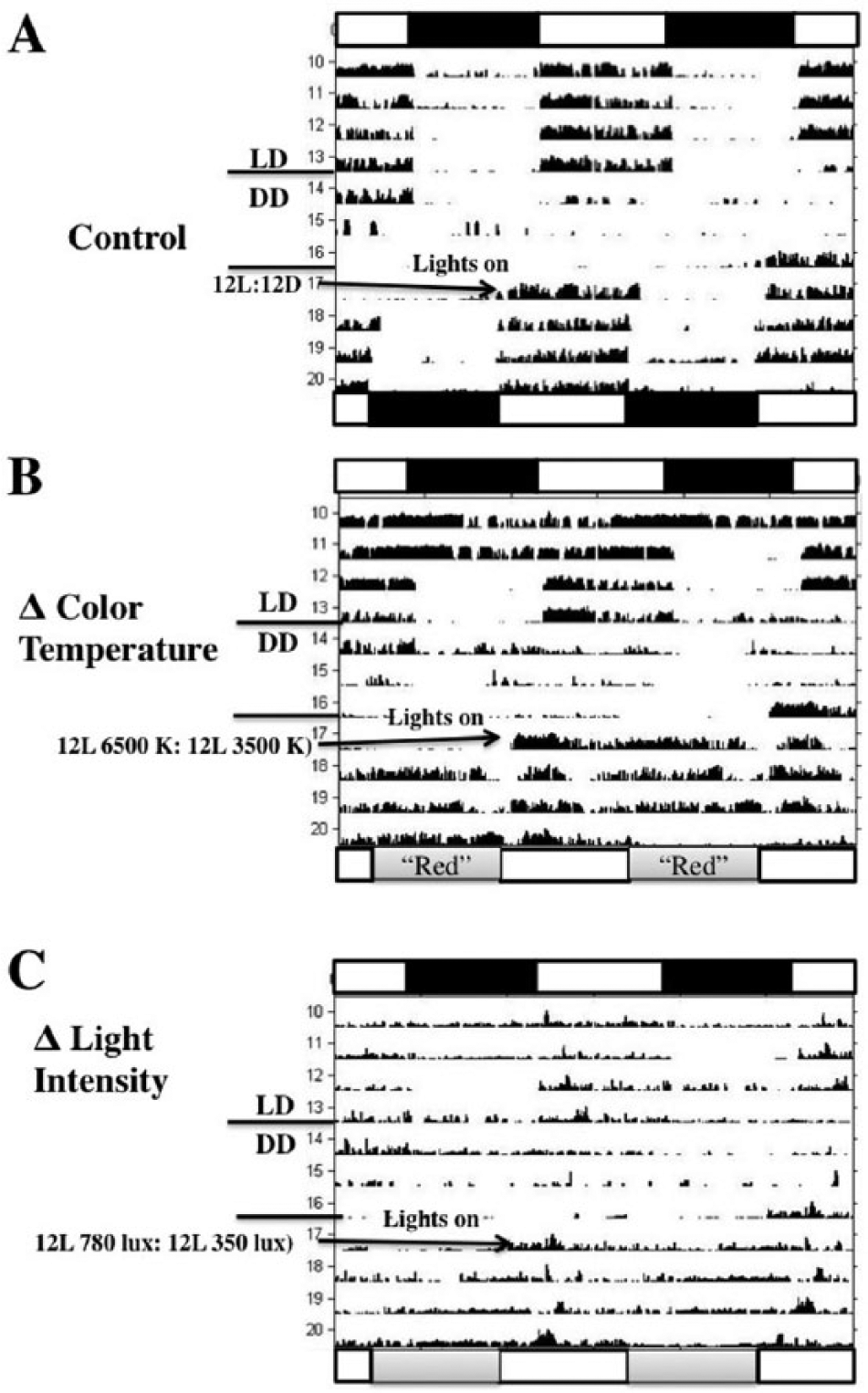
Representative actograms (double-plotted) of longspur activity rhythms subjected to 12L:12D, 3 d of DD, and then (A) 12L (780 lux, 6500 K; horizontal white bar):12 DD (black bar; control), (B) 12 L (780 lux, 6500 K):12L (780 lux, 3500 K; lower gray bar designated as “red”), or (C) 12 L (780 lux, 6500 K):12L (380 lux, 6500 K, gray bar). Horizontal bars on the top of graphs represent light-dark schedule (black depicts dark phase) of initial 12L:12D treatment. Horizontal bars on the bottom of graphs represent experimental light regimes. B shows a tendency toward a free-running rhythm, whereas C displays arrhythmia.
Experiment 3
All birds entrained to the 12L:12D photoperiod before starting the experiment. For llPer2 expression of hypothalamic punches, there were no significant effects of light treatment (2-way ANOVA; F2,35 = 2.29, p = 0.12), sampling time (F3,35 = 0.84, p = 0.48), or their interaction (F6,35 = 1.12, p = 0.37; Fig. 5A). Moreover, hypothalamic llPer2 expression did not display a periodic relationship with time of day (harmonic regression, all p’s > 0.05). However, there was a significant effect of sampling time on llPer2 expression in the eye (2-way ANOVA; F3,36 = 5.17, p = 0.005) but no significant effects of light treatment (F2,36 = 0.98, p = 0.38) or its interaction with sampling time (F6,36 = 1.20, p = 0.33). More specifically, ocular llPer2 expression was higher at 24 h compared with 6, 12, and 18 h (Fisher PLSD, all p < 0.048) for LD and LL birds (Fig. 5B). These findings were further corroborated by significant circadian periodicity identified in ocular llPer2 expression of LL birds (harmonic regression, F2,13 = 4.48, p = 0.03, R2 = 0.408) but not LD birds. For llPer2 expression in liver, there was a significant effect of lighting treatment (2-way ANOVA, F2,36 = 7.39, p = 0.002). Neither sampling time (F3,36 = 1.70, p = 0.19) nor its interaction with lighting treatment (F6,36 = 1.48, p = 0.22) was statistically significant. On average, llPer2 expression in liver was significantly higher in LD birds compared with LL and DD longspurs (Fisher PLSD, all p < 0.004; Fig. 5C). Liver llPer2 expression was not associated with time of day (harmonic regression, all p’s > 0.05).
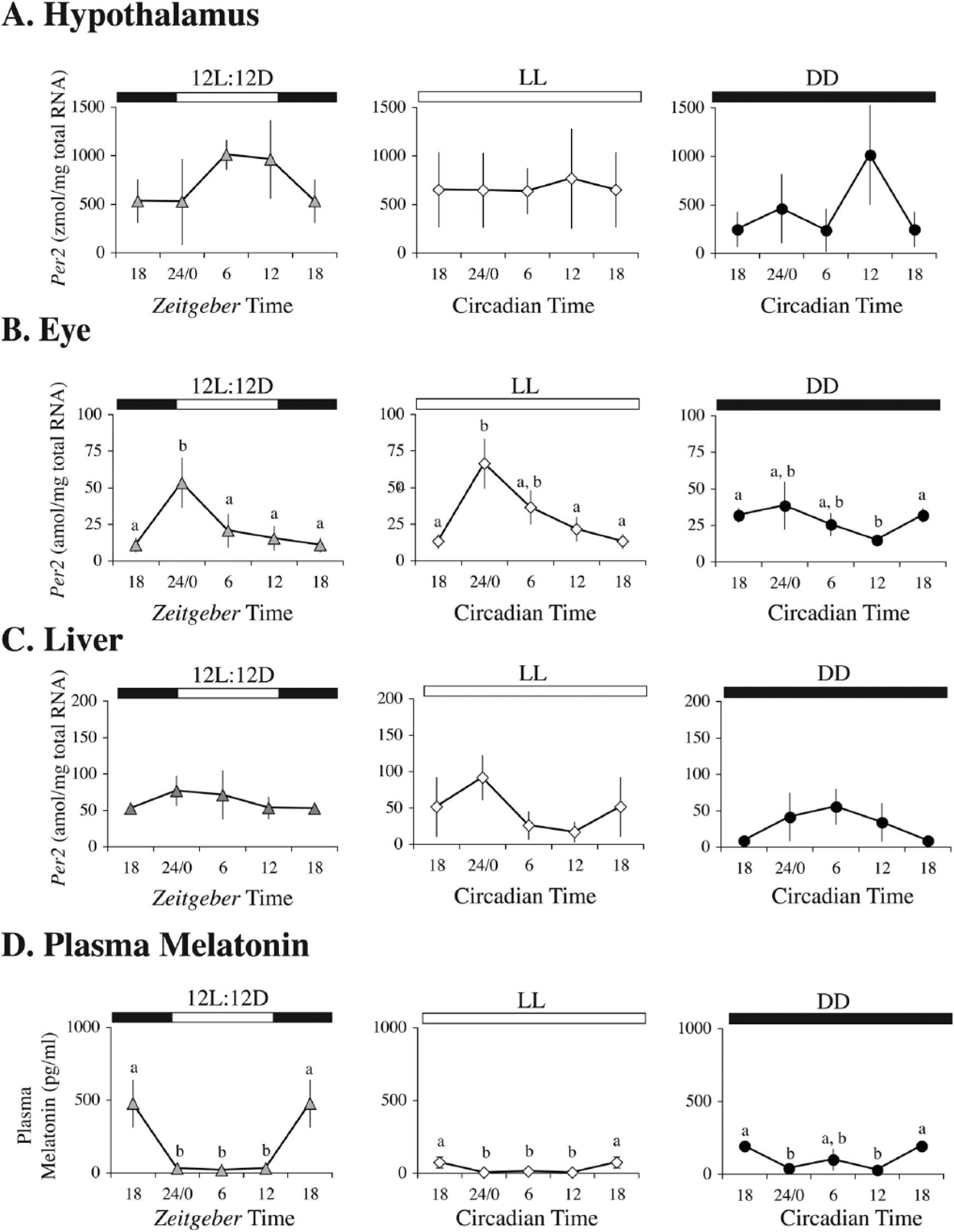
Per2 mRNA concentration in (A) hypothalamus (zmol/mg total RNA), (B) eye (amol/mg total RNA), and (C) liver (amol/mg total RNA) and (D) plasma melatonin concentration (pg/mL) at 4 different time points in LD, LL, and DD. Bars represent mean ± SEM. Horizontal bars on top of graphs represent light-dark schedule (black depicts dark phase). ZT18 and CT18 are double-plotted for better visualization of rhythms. Within-group comparisons that share letters indicate no significant differences between time points. n = 4 per group, except for the hypothalamus, ZT24/0 group (n = 3).
Lighting treatment (F2,36 = 6.97, p = 0.003) and sampling time (F3,36 = 17.06, p < 0.0001) had significant effects on plasma melatonin concentration, but there was no significant interaction effect between the 2 factors (F6,36 = 0.82, p = 0.56). Irrespective of time, average melatonin concentration was suppressed in LL longspurs compared with LD and DD birds (Fig. 5D; Fisher PLSD, all p < 0.014). Irrespective of treatment, melatonin concentration was significantly elevated at 18 h relative to 6-, 12-, and 24-h sampling periods (Fisher PLSD, all p < 0.001). Furthermore, a significant melatonin rhythm in relation to the 24-h cycle was identified in LD birds (harmonic regression; F2.13 = 5.33, p = 0.02, R2 = 0.45) but not in LL or DD birds.
Discussion
Free-Running Rhythms in Constant LL and DD
Male and female longspurs exhibited period lengths that were significantly less than 24 h when exposed to LL or DD, indicating the presence of an endogenous clock(s) that regulates locomotor activity. These findings also support the hypothesis that free-living longspurs are likely synchronizing rhythms to low-amplitude polar Zeitgeber cues because our data indicate that an environment lacking these potential entraining agents, namely, constant captive conditions, induces free-running activity rhythms. Although exposure to bright LL (2000 lux) can induce rapid arrhythmia in some species (e.g., pigeons, Columba livia; Yamada et al., 1988), house sparrows will initially exhibit a short free-running period for >7 d when exposed to LL (100 lux), but rhythms eventually become arrhythmic (Cassone et al., 2008). Even though our study used lighting of higher intensity (1300 lux) to attempt to mimic the lower threshold of polar-day light intensities, longspurs still expressed free-running rhythms. Because activity rhythms were assessed for a short period of time (<7 d), we can only speculate that continual LL in this species leads to arrhythmia. In additional experiments (experiment 2), some longspurs became arrhythmic, suggesting that prolonged exposure to nonentrainable light regimens leads to abandonment of circadian rhythms.
Jürgen Aschoff (1960, 1981, 1989) first recognized that free-running rhythms are affected by light intensity. This relationship, termed Aschoff’s rule, states that free-running periods measured in DD will shorten for diurnal animals and lengthen for nocturnal animals when exposed to LL (Aschoff, 1979). Although there are a number of exceptions, especially among diurnal species (Refinetti, 2006), free-running periods were significantly shorter on LL compared with DD, which supports Aschoff’s rule. Despite free-living longspurs being exposed to continuous bright light on their breeding grounds, previous studies have rejected the possibility that longspurs are exhibiting shortened free-running periods; instead, an intact, 24-h rhythm is maintained (Hau et al., 2002; Ashley et al., 2013; Steiger et al., 2013).
Accumulating evidence indicates that circadian rhythms are influenced not only by natural selection but also by factors relevant to sexual selection, such as mating prospects and parental responsibilities (Helm and Visser, 2010; Steiger et al., 2013). For example, among polygamous shorebird species, the incubating sex is rhythmic during incubation, while the nonincubating sex is arrhythmic and active throughout the polar day (Steiger et al., 2013). These observations suggest that sexual differences in circadian rhythmicity are a consequence of intense sexual selection, which results in skewed mating opportunities between the sexes and the necessity to optimize daily activity rhythms to these demands. In contrast, Lapland longspurs are socially monogamous (cases of polygyny are rare), and males participate in parental duties, such as feeding of nestlings (Hussell and Montgomerie, 2002). Therefore, the lack of sexual differences in free-running rhythms aligns closely with this species’ monogamous mating strategy and life history.
No Entrainment to Diel Shifts of Light Intensity, Color Temperature, or Both
Longspurs failed to synchronize activity rhythms to alternating periodic 12-h fluctuations in light intensity, color temperature, or both treatments combined. Despite constant illumination, the arctic climate is characterized by measurable diurnal fluctuations in light intensity, spectral quality of light, and ambient temperature (e.g., Ashley et al., 2013) that could act as potential entraining agents. It remains to be seen how predictable these variations are, given the vagaries of arctic coastal weather, where cloud cover and fog can unpredictably alter the photic and thermal environment. Nonetheless, previous studies have demonstrated that songbirds can synchronize activity rhythms to artificial changes in light intensity (Krüll, 1976b) and color temperature (Pohl, 1999). Granted that it is difficult to replicate environmental conditions in captivity and that animals often respond differently to captivity versus free-living environments (Calisi and Bentley, 2009), it is not surprising that entrainment of rhythms did not occur. In retrospect, the light intensities used for the experiments, even the reduced setting (350 lux), have been viewed as “bright” light conditions in previous captive studies (e.g., Kumar et al., 2000; Kumar et al., 2007; Cassone et al., 2008; Pandey and Bhardwaj, 2011). Although we were attempting to mimic light intensities of the arctic summer, in reality, the range of values used are far lower than those measured in polar day (exceeds 105 lux). Thus, a future goal would be to identify the range of light intensities that will induce entrainment to better understand this species’ sensitivity to the photic environment.
It has been hypothesized that diurnal changes in the spectral composition of light act as more consistent environmental cues than diel fluctuations in light intensity (Krüll, 1976c; Krüll, 1985). Synchronization of longspur rhythms did not occur under an alternating Zeitgeber cycle of 6500 K (blue light) to 3500 K (red light). However, light intensity was still high for captive standards (760 lux), possibly overstimulating the circadian system. A previous study has demonstrated that chaffinches can entrain rhythms to a cycle of 5600 K to 2900 K at much lower light intensities (2.5 lux; Krüll, 1985). This seemingly indicates that lower light intensities are necessary to induce entrainment to an alternative Zeitgeber. However, if the goal of the study is to replicate polar-day conditions in the wild, then relying on low intensities has little biological relevance for birds living in arctic summer conditions.
llPer2 Rhythmicity in Eye but not Hypothalamus or Liver
Longspurs displayed rhythmicity in ocular llPer2 gene expression when exposed to LL (according to harmonic regression analysis), which provides support for endogenous circadian regulation. Although not direct evidence of periodicity, llPer2 ocular gene expression differed significantly among time points for LL and LD treatments. Expression was highest at ZT24 and CT24 for the LD and LL groups, respectively, whereas differences were dampened in the DD group. In Japanese quail, qPer2 is rhythmically expressed in the eye with peak expression also occurring at ZT0/24 (Yoshimura et al., 2000). In addition, light pulses induce ocular qPer2 expression, which corroborates why Per2 is elevated during the light period (Yoshimura et al., 2000). It is unclear why no detectable rhythms were identified in the hypothalamus or liver, but large standard error bars indicate substantial individual variation. Previous research in house sparrows has demonstrated rhythmic expression in both the mSCN and vSCN (Brandstätter et al., 2001; Abraham et al., 2002), but the methods to assess expression were quite different, relying on immunohistochemical techniques. Our sampling protocol involved using competitive quantitative PCR (qPCR) to assess llPer2 expression directly from tissue punches. Although this method has been used successfully in other avian studies (Perfito et al., 2012; Lopes et al., 2013) and data generated could be used to compare with qPCR results from the liver and eye, it failed to distinguish between gene expression of the 2 hypothalamic structures associated with avian circadian function (or any nearby structures that may have been constitutively expressing llPer2, such as the preoptic nucleus; Abraham et al., 2002). Thus, it is unknown whether lack of rhythmicity occurred because of our inability to detect out-of-phase oscillations occurring independently in the 2 hypothalamic nuclei, a lack of an endogenous rhythm in one or both structures, or a sampling artifact from measuring gene expression from areas in addition to the mSCN or vSCN. Lastly, the 4-h sampling rate could have been too low to adequately detect a rhythm. Thus, these results involving the hypothalamic gene expression are inconclusive and require further investigation.
Rhythms of clock gene expression also exist in peripheral cells and tissue, and these rhythms are self-sustaining in vitro (Yoo et al., 2004) but are synchronized by master pacemaker activity in vivo (e.g., SCN). Although the majority of research involving peripheral clocks has been conducted in mammals (for review, see Mohawk et al., 2012), there is some evidence for rhythmicity of core clock genes in various tissues in birds. For example, in Japanese quail, qPer2 is rhythmic in heart, kidney, and lung but weakly expressed in liver, skeletal muscle, spleen, and testes (Yoshimura et al., 2000). In a night-migratory songbird, the redheaded bunting (Emberiza bruniceps), rhythmic expression of rbPer2 was reported in liver, heart, spleen, gut, and testes, with peak expression phase-aligned to lights-on (Singh et al., 2013). However, in our study, there was no detectable rhythm of llPer2 expression in the liver of birds exposed to either LD or constant conditions. Assuming that peripheral clocks should be phase aligned with central oscillators, we would predict such rhythmicity to be apparent. Nonetheless, LD exposure, on average, elevated liver llPer2 expression compared with LL or LD when all sampling times were combined. Feeding cycles can entrain the liver independently of the SCN and light cycle in rats (Stokkan et al., 2001). Thus, it is conceivable that longspur feeding rhythms were affected by different light conditions, which in turn could have altered liver llPer2 expression, although this possibility is purely speculative.
Suppression of Melatonin in Constant Light
Melatonin is an indole amine hormone that is synthesized and released at night by the pineal gland (Reiter, 1991). Conversely, light suppresses melatonin production, and the light-dark cycle synchronizes circadian oscillators that regulate pineal rhythmicity, which encodes time of day and time of year (Kumar et al., 2000; Bentley, 2001; Brandstätter, 2003). During the polar summer, continuous illumination completely abolishes melatonin rhythms in some species (Barrell and Montgomery, 1989; Miché et al., 1991; Reierth et al., 1999) or substantially dampens them in others, including free-living Lapland longspurs (Cockrem, 1991; Hau et al., 2002; Silverin et al., 2009; Ashley et al., 2013). Upon exposure to LL, captive longspurs also exhibit suppression of plasma melatonin relative to LD and DD, but a rhythm still persists. Considering that constant bright light (2000 lux) completely abolishes melatonin rhythms in pigeons (Yamada et al., 1988), this result was unexpected. We speculate that maintenance of a melatonin rhythm in continuous light is an adaptation to polar-adapted organisms that specifically retain intact behavioral and physiological rhythms during the constant illumination of summer. As evidence, the seasonal dampening of the melatonin amplitude during polar day appears to weaken the self-sustaining circadian pacemaker(s), which enhances entrainability of rhythms to low-amplitude polar zeitgeber signals (Gwinner et al., 1997; Gwinner and Brandstätter, 2001). Yet, in this study, captive longspurs failed to entrain to low-amplitude changes in light intensity and/or color temperature. Given the challenges of mimicking field conditions in captive studies, we hypothesize that longspurs require more accurate light intensity and color temperature cycles that better simulate the photic environment of polar summer to demonstrate entrainment.
Conclusions
This study demonstrates that the circadian rhythm reported in free-living Lapland longspurs during the constant daylight of arctic summer (Hau et al., 2002; Ashley et al., 2013; Steiger et al., 2013) is regulated by an internal circadian clock that cycles endogenously with a period less than 24 h. These findings provide support for the hypothesis that free-living longspurs synchronize diel rhythms to weak environmental cues during the polar day. On a molecular level, there is evidence that llper2 expression and plasma melatonin profiles were rhythmic in the eye and pineal gland, respectively, despite a lack of rhythms in hypothalamic or hepatic llper2 expression. These data suggest that, at the very least, ocular and pineal clocks are important for generating endogenous rhythms in this species. The phasing of these rhythms is similar to those reported in temperate-breeding species (Gwinner et al., 1997; Yoshimura et al., 2000; Gwinner and Brandstätter, 2001). We suspect hypothalamic llPer2 rhythms also play a role and attribute our inability to detect such rhythms to our sampling methods (see above).
As for identifying the polar Zeitgeber that has the potential to entrain longspur rhythms in captivity, we were unable to demonstrate synchronization of longspur activity to artificial diel fluctuations in light intensity, color temperature, or both. Although it is difficult to replicate environmental variables in captive settings, these negative results suggest that longspurs lack the capacity to entrain to some light intensity and color temperature Zeitgeber cycles. However, these results do not necessarily rule out the possibility that longspurs synchronize rhythms to daily light cycles in their natural polar environment; maximum and diel variation of light intensities recorded during polar day can far exceed those used in our captive study by several orders of magnitude (Hau et al., 2002; Ashley et al., 2013). In addition, other abiotic factors, such as UV light, sun azimuth (position), and ambient temperature, exhibit diel variations and could potentially act as a Zeitgeber during arctic summer (Swade and Pittendrigh, 1967; Krüll, 1976a, 1976b, 1976c, 1985; Pohl, 1992, 1999; Ashley et al., 2013). Biotic factors, such as daily activity patterns of predators, prey, competitors, or mates, could also influence rhythmicity, although these are likely synchronized by 1 or more of the abiotic cues discussed above (Hau et al., 2002; Rani et al., 2009). Therefore, the cue(s) that entrain longspur rhythms to polar zeitgeber signals during the arctic summer remains elusive and warrants future investigation.
Footnotes
Acknowledgements
We thank Katie Akpik, Luke George, and Vera Simmonds for field assistance and the staffs at Barrow Arctic Science Consortium and CH2MHill Polar Services for logistical support. We are indebted to the Ukpeagivk Inupiat Corporation for permitting us to conduct fieldwork on their tribal lands. B.M.S. was supported by a University of Alaska Fairbanks Center for Global Change and Arctic Systems Research Award and the University of Alaska Anchorage Fran Ulmer Transformative Research Award. N.T.A. was supported by an Institutional Development Award (IDeA) from the National Institute of General Medical Sciences of the National Institutes of Health under grant 5P20GM103436-13. We thank 2 anonymous reviewers and Dr. William Schwartz for providing helpful feedback that improved the manuscript. This research was funded by the National Science Foundation (grant OPP 0817635).
Conflict of Interest Statement
The author(s) have no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.