
Abstract
The circadian clock plays an important role in adaptation in time and space by synchronizing changes in physiological, developmental, and behavioral traits of organisms with daily and seasonal changes in their environment. We have studied some features of the circadian activity and clock organization in a northern Drosophila species, Drosophila montana, at both the phenotypic and the neuronal levels. In the first part of the study, we monitored the entrained and free-running locomotor activity rhythms of females in different light-dark and temperature regimes. These studies showed that D. montana flies completely lack the morning activity component typical to more southern Drosophila species in an entrained environment and that they are able to maintain their free-running locomotor activity rhythm better in constant light than in constant darkness. In the second part of the study, we traced the expression of the PDF neuropeptide and the CRY protein in the neurons of the brain in D. montana adults and found differences in the number and location of PDF- and CRY-expressing neurons compared with those described in Drosophila melanogaster. These differences could account, at least in part, for the lack of morning activity and the reduced circadian rhythmicity of D. montana flies in constant darkness, both of which are likely to be adaptive features during the long and dark winters occurring in nature.
Adaptation to daily and seasonal changes in environmental conditions is of a crucial importance for organisms living in the northern latitudes, where the adjustment of developmental and behavioral traits with changes in local environmental conditions plays a central role in survival and offspring production. The 2 major internal time-keeping mechanisms underlying such phenotypic rhythmicity are the circadian clock, which regulates daily rhythmicity, and the photoperiodic calendar, which regulates seasonal rhythmicity (Koštál, 2011).
The circadian clock maintains many intrinsic rhythms of organisms with a period that is approximately 24 h. However, since the length of an intrinsic day is not exactly 24 h, the circadian pacemaker has to be resynchronized every day by environmental signals, zeitgebers, such as light or temperature (Dubruille and Emery, 2008). The ability of the circadian clock to entrain circadian rhythms by environmental signals enables organisms to anticipate daily and seasonal changes in their environment and adjust their behavior, development, and reproduction accordingly. The circadian clock is also known to keep the circadian pacemaker oscillating and to maintain the circadian rhythms of the organisms in constant conditions (free-running rhythms) for extended periods.
Under standard laboratory conditions, Drosophila melanogaster fruit flies display a bimodal locomotor activity rhythm with pronounced morning and evening activity peaks and little activity during midday or night (Hamblen-Coyle, 1992; Rieger et al., 2003; Yoshii et al., 2009). Nevertheless, different behavioral features are observed when flies are recorded under natural environmental conditions (Vanin et al., 2012). The master circadian clock of this species is known to be localized at the level of approximately 150 lateral (LN) and dorsal (DN) neurons, which express particular circadian clock genes (reviewed in Taghert and Shafer, 2006; Nitabach and Taghert, 2008). LNs and DNs can further be divided into several subgroups: the small ventrolateral neurons (s-LNvs), the large ventrolateral neurons (l-LNvs), the dorsolateral neurons (LNds), and 3 groups of dorsal neurons (DN1s, DN2s, and DN3s) (reviewed in Helfrich-Förster, 2003), as well as lateral-posterior neurons (LPNs) (Taghert and Shafer, 2006). In D. melanogaster, 4 of 5 s-LNvs and all l-LNvs express pigment-dispersing factor (PDF) neuropeptide (see Yoshii et al., 2009), which plays an important role in synchronization of the oscillations of the circadian clock neurons and in transferring the information from the central circadian pacemaker to downstream neurons (Yoshii et al., 2009). PDF-positive s-LNvs have been implicated in control of the morning locomotor activity in light-dark laboratory (LD) conditions, forming a neuronal basis for the morning (M) oscillator, while the fifth PDF-negative s-LNv and LNds seem to be responsible for the evening activity and are defined as the evening (E) oscillator under LD conditions (Grima et al., 2004; Stoleru et al., 2004; Rieger et al., 2006; Nitabach and Taghert, 2008). In this species, s-LNvs are also the most important clock neurons for maintaining robust circadian locomotor activity rhythms under constant darkness (Renn et al., 1999).
In D. melanogaster, circadian clock genes, including Clock (Clk), cycle (cyc), period (per), and timeless (tim), are expressed in the clock neurons in the fly brain, where they form the molecular basis for circadian oscillation of the clock (reviewed in Peschel and Helfrich-Förster, 2011). In this species, the circadian photoreceptor gene cryptochrome (cry) plays an important role in entraining the flies to day-night cycles by conveying light-induced degradation of TIM protein (e.g., Stanewsky et al., 1998) and contributes also to the adjustment of the evening activity (Emery et al., 2000b). cry has been found to be expressed in s-LNvs, l-LNvs, a subset of LNds, and DN1s (Benito et al., 2008; Yoshii et al., 2008).
At high latitudes (65-69 °N), the drastic changes in day length in late summer and autumn enable organisms to anticipate the forthcoming cold period with a great accuracy, and here the long light and dark periods during mid-summer and mid-winter may have led to a weaker circadian regulation of activity rhythms in constant conditions. For example, reindeer (Rangifer tarandus platyrhynchus) living north of the Arctic Circle do not show rhythmicity in their daily activity during the summer, even though they display a clear circadian locomotor activity rhythm during other seasons (van Oort et al., 2005). Also, latitudinal clines detected in the photoperiodic responses of D. melanogaster suggest that flies from northern populations of the species have reduced light sensitivity (Sandrelli et al., 2007; Tauber et al., 2007).
In plants and animals, a second time-measuring system, the photoperiodic timer, plays an important role in enabling the organisms to cope with seasonal changes in their environment (Koštál, 2011). Even though the genetic regulation of D. melanogaster’s circadian clock is well described (reviewed in Allada and Chung, 2010; Peschel and Helfrich-Förster, 2011), the genetic and physiological mechanisms underlying the photoperiodic calendar are poorly understood, partly due to the fact that D. melanogaster is not adapted to live in the seasonally varying environments at high latitudes (Schiesari et al., 2011). Our study species, Drosophila montana, belongs to the Drosophila virilis species group. The divergence time between D. montana and D. virilis is approximately 10 million years (Spicer and Bell, 2002), while D. virilis (genus Drosophila) and D. melanogaster groups (genus Sophophora) already had diverged about 63 million years ago (Tamura et al., 2004). D. montana has spread at high latitudes around the globe (Throckmorton, 1982) and shows a robust photoperiodic response in female reproductive diapause (Tyukmaeva et al., 2011). Understanding the function of the circadian clock in D. montana will offer great possibilities to trace its role in adaptation to northern environmental conditions both alone and together with the photoperiod timer. In the present study, we have characterized the entrained and free-running locomotor activity rhythms of D. montana females under different LD and temperature conditions. Combining these data with information on PDF and CRY expression in the flies’ neurons enabled us to search for possible explanations for differences in the behavior between the flies of this species and D. melanogaster flies. We aimed to answer the following questions: How do the entrained and free-running locomotor activity rhythms of D. montana flies differ from those of D. melanogaster, and are there differences between these species also at the neuronal level? What kind of environmental factors affect fly rhythmicity in entrained and constant conditions? And does the function of the circadian clock of D. montana show features that could be adaptive to survival in the seasonally varying conditions of high latitudes?
Materials and Methods
Fly Strains and Rearing Conditions
We measured the adult locomotor activity of the females of 3 D. montana isofemale strains (175OJ8, 265OJ8, and 26OL8) that originated from Oulanka (66 °N), Finland, in 2008. The strains have been maintained in constant light of ~300 lux at 19 °C in bottles containing malt medium (Lakovaara, 1969) since their collection. These lighting and temperature conditions roughly resemble the conditions during the flies’ mating season in the wild, even though they lack daily changes in temperature and light intensity.
The fly strains that we chose in this study have been used earlier in studies concerning developmental speed and/or diapausing behavior in D. montana (T. Salminen, unpublished data; Tyukmaeva et al., 2011). While flies of the strains 175OJ8 and 265OJ8 behaved in these studies in a way that is typical of the species, the flies of the strain 26OL8 showed some peculiarities. Thus, it was interesting to see whether we could detect differences between the strains also in the function of the circadian clock.
Recording the Entrained and Free-Running Locomotor Activity Rhythms of Females
To record the locomotor activity rhythmicity, females of the 3 study strains were collected within 1 day of eclosion and transferred into glass tubes (diameter 10 mm and length 70 mm; 1 female per tube). One end of each tube was filled with a 2-cm-thick layer of malt medium and sealed with a plastic film (Parafilm M), while the other end was blocked with a cotton plug. The tubes were inserted into Trikinetics Drosophila Activity Monitors (Waltham, MA) with 32 tubes per monitor, which were then placed in temperature-controlled environmental chambers (Binder FD 115; Binder GmbH, Tuttlingen, Germany) in 90% humidity. The monitors were illuminated with white LEDs (ProLED; MBN GmbH, Friedberg, Germany) with a light intensity of ~150 lux. The entrained locomotor activity rhythm of the flies was studied by maintaining the monitors under a 22:2, 20:4, or 16:8 LD cycle at 16 °C or 20 °C for 8 days. After the entrainment, the monitors were placed either into constant darkness (DD) or constant light (LL) at the same temperature for 12 days to trace the existence and period of free-running activity rhythms.
After the locomotor activity experiments, the females were stored at −20 °C until their reproductive stage was determined on the basis of the size and developmental stage of their ovaries (see Tyukmaeva et al., 2011).
Analysis of Fly Locomotor Activity
The raw locomotor activity data of adult flies were first displayed as double-plotted actograms (48-h plots) for 8 days under different entraining conditions (LD cycles) and for 12 days under constant darkness or light (DD or LL). These data were analyzed with the program ActogramJ (Schmid et al., 2011; available at http://actogramj.neurofly.de). The overall activity level of the flies was traced under different entraining conditions by calculating the mean activity levels of all flies of each strain over the 8 experimental days in 5-min bins. The effects of temperature and photoperiod on the mean activity levels of the flies were analyzed with 2-way analysis of variance (ANOVA) with temperature and photoperiod as fixed factors. Prior to performing these tests, normality assumptions of the distributions of fly activity levels were tested with the Kolmogorov-Smirnov and Shapiro-Wilk tests and homogeneity of variance with the Levene test. Since the expectations for homogeneity of variance for strain 26OL8 were not fulfilled regardless of transformations, the nonparametric Kruskal-Wallis test was used to test the effects of photoperiod on the mean activity level of the flies in different temperatures. The effect of temperature on the mean activity level of the flies of this strain in each photoperiod was tested by using independent samples t test. Because of possible nonnormal distributions of fly activity levels and the small sample size, the results were checked by using Mann-Whitney U tests. All analyses were conducted with PASW Statistics 18.0 (SPSS, Inc., an IBM Company, Chicago, IL).
The actograms were analyzed with the Lomb-Scargle periodogram method to determine whether the flies showed significant rhythmicity in their behavior under entrained and/or constant conditions and to measure the period (τ) of the free-running intrinsic day in constant conditions. A fly was determined to be rhythmic under entraining conditions if the periodogram analysis detected significant periodicity in its activity rhythm measured across several consecutive days (the significance level in the Lomb-Scargle periodogram analysis was adjusted to 0.05). The free-running period (τ)—that is, the length of the intrinsic day—was measured for flies that had first been entrained in 1 of the 3 photoperiods and then kept in constant darkness or light for 12 days (also here the significance level in the Lomb-Scargle periodogram analysis was adjusted to 0.05). The power of the Lomb-Scargle periodogram analysis was defined as the amplitude of the peak only for the rhythmic flies from the Lomb-Scargle periodogram with a significance level of p < 0.05. Flies that did not survive throughout the whole experiment (both entrained and free-running conditions) were excluded from the analysis.
To determine whether the strain, temperature, entraining photoperiod, and/or constant condition treatment affected fly rhythmicity, the locomotor activity data were made binary, with a value of 1 for each rhythmic fly and a value of 2 for each arrhythmic fly. The data were analyzed with hierarchical logit models, and the goodness of fit of the final model, including all significant variables and their interaction terms, was tested by using the likelihood-ratio χ2 test. Since the flies of strains 175OJ8 and 265OJ8 showed no significant differences in their behavior in any model, we conducted a logit model where the data for these strains were combined (group 1), keeping the data for the strain 26OL8 separate (group 2). For more detailed information about hierarchical logit models used here, see Supplemental Figure S1.
Immunohistochemistry
The strain 175OJ8 was used in immunohistochemistry experiments. Prior to PDF and CRY staining, female flies were kept in constant darkness (DD) at 20 °C for 72 h to enhance CRY staining (cf. Yoshii et al., 2008). At zeitgeber time (ZT) 0, the flies were fixed in 4% paraformaldehyde in a phosphate buffer (PB, pH 7.4) with 0.5% Triton X in darkness for 4 h at room temperature and rinsed 3 times for 15 min in PB, after which their brains were dissected in the same buffer. Brains were collected in PB with 0.5% Triton X and subsequently blocked in 5% normal goat serum (NGS) in PB with 0.5% Triton X overnight at +4 °C before treating them with primary and secondary antibodies.
The primary rabbit anti-CRY was developed by Takeshi Todo (Yoshii et al., 2008) and mouse anti-PDF by Justin Blau (obtained from the Developmental Studies Hybridoma Bank [DSHB]). The primary rabbit anti-CRY was diluted by 1:1000 and the primary mouse anti-PDF by 1:2500 in PB containing 5% NGS and 0.5% Triton X-100. Brains were incubated in one of these antibodies for 48 h at 4 °C and rinsed 5 times for 10 min in PB with 0.5% Triton X-100.
The secondary fluorescence-conjugated antibody PDF and CRY stainings were performed for 2 different groups of brains. As secondary antibodies in these stainings, we used mouse anti-PDF Alexa Fluor 546 (goat anti-mouse; Invitrogen, Carlsbad, CA) and anti-CRY Alexa Fluor 488 (goat anti-rabbit; Invitrogen). The secondary antibodies were diluted 1:200 in PB containing 5% NGS and 0.5% Triton X-100 and kept in this buffer overnight at +4 °C. The brains were washed 5 times for 10 min in PB with 0.5% Triton X and 1 time for 10 min in PB with 0.1% Triton X-100 and mounted on Vectashield mounting medium (Vector Laboratories, Burlingame, CA). The fluorescence signals of the whole brains were detected by using a confocal microscope (Olympus FV1000; Olympus, Center Valley, PA). We examined at least 10 brains for PDF and CRY immunostaining. Images were processed using the program ImageJ (available at http://rsbweb.nih.gov/ij/).
Results
Entrained Locomotor Activity Rhythms of D. montana Females
We used only females in this study since the flies of the 2 sexes have been found to show similar locomotor activity profiles (H. Kauranen, unpublished data). The flies of the 175OJ8 and 265OJ8 strains were significantly more active at 20 °C than at 16 °C in all photoperiods (2-way ANOVA for 175OJ8: F1, 365 = 12.6, p < 0.001; for 265OJ8: F1, 356 = 16.9, p < 0.001). In strain 26OL8, the photoperiod and temperature showed interactions in their effects on the flies’ activity, and the flies of this strain were more active at 20 °C than at 16 °C in only 2 of the 3 photoperiods (t test for 26OL8 in 20:4 LD: t = −4.305, df = 113.1, p < 0.001 and in 22:2 LD: t = −7.272, df = 98.6, p < 0.001). Furthermore, the flies of this strain were less active in photoperiod 16:8 LD than in other photoperiods at 20 °C (Kruskal-Wallis: K = 28.56, n = 212, df = 2, p < 0.001) but not at 16 °C (Kruskal-Wallis: K = 4.078, n = 165, df = 2, p = 0.130).
D. montana flies possessed good entrained locomotor activity rhythms with a clear evening activity peak, but they totally lacked morning activity in all studied photoperiods and temperatures (Fig. 1A-D; Fig. 2A-C; Suppl. Fig. S2-S3). In the shortest day length (16:8), the highest activity peaks of the flies overlapped with the lights-off transition, while in the longer day lengths, these peaks usually took place before this (Fig. 2A-C; Suppl. Fig. S4-S5). In all photoperiods, the activity of the flies of all study strains decreased rapidly at the beginning of the dark period and vanished completely toward the end of night.
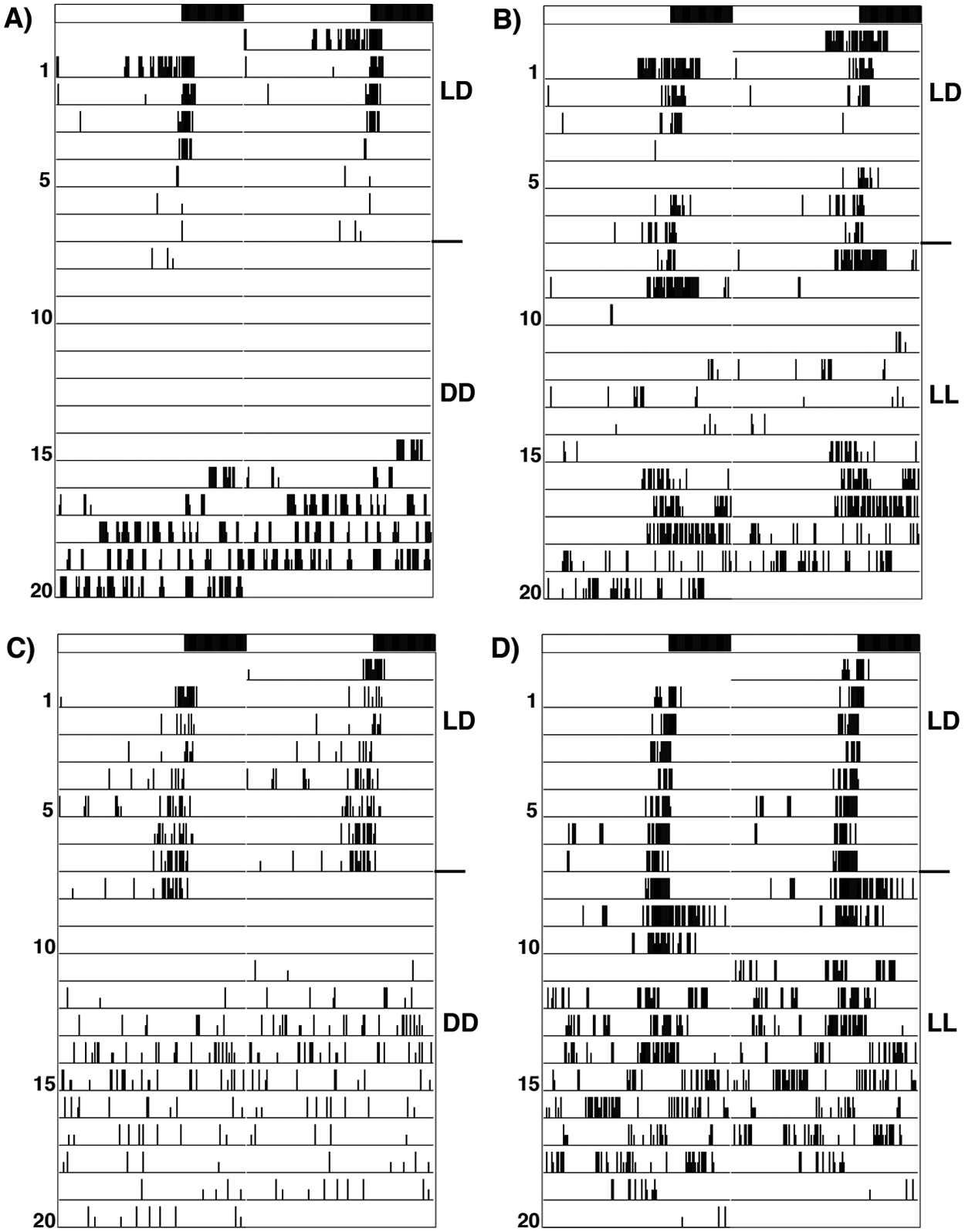
Typical actograms of Drosophila montana females of strain 175OJ8 in different entraining and constant conditions. Each actogram represents an activity profile of 1 fly during 8 days in entraining conditions and 12 days in constant conditions. (A) LD 16:8, 16 °C and release into DD. (B) LD 16:8, 16 °C and release into LL. (C) LD 16:8, 20 °C and release into DD. (D) LD 16:8, 20 °C and release in LL.
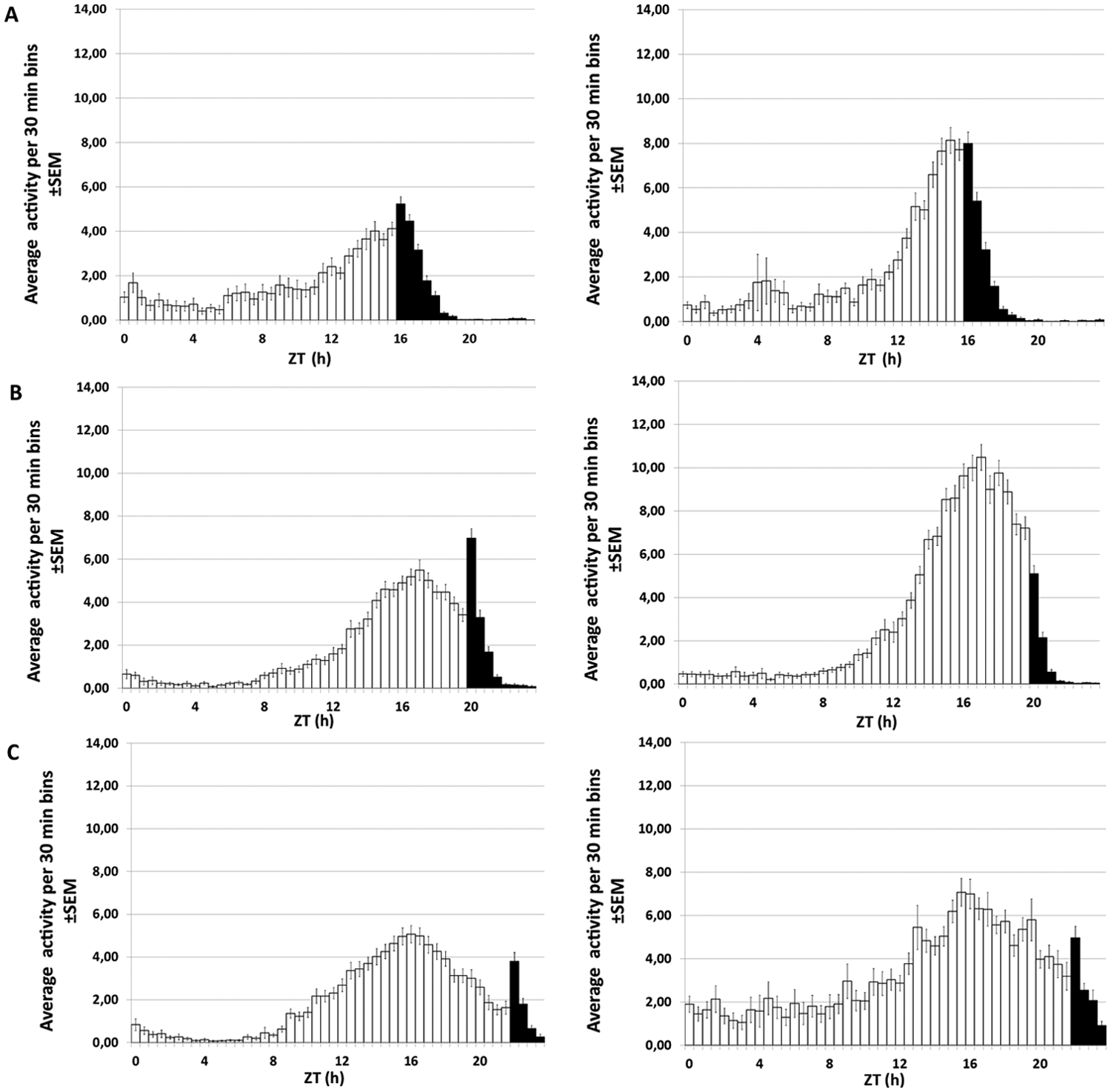
Mean activity profiles of females of strain 175OJ8 under different entraining conditions. (A) LD 16:8, 16 °C (left) and 20 °C (right). (B) LD 20:4, 16 °C (left) and 20 °C (right). (C) LD 22:2, 16 °C (left) and 20 °C (right). The heights of the bars indicate the mean activity levels of the flies during each 30-min bin over 8 days. Light period is indicated by white and dark period by black bars.
When tracing the factors affecting fly rhythmicity in different entraining conditions with logit models, we ended up with the multinomial logit model: entraining rhythmicity × temperature × group + entraining rhythmicity × photoperiod (likelihood ratio [LR] = 11.81, df = 6, p = 0.066). The flies of the strains 175OJ8 and 265OJ8 (group 1) showed stronger rhythmicity at the higher temperature (93.9% of the flies of these strains were rhythmic at 20 °C and 84.5% at 16 °C), and the photoperiod 20:4 increased flies’ rhythmicity in all strains (20:4 LD: 87.6%, n = 338; 16:8 LD: 83.4%, n = 327; and 22:2 LD: 81.6%, n = 271). Overall, the proportion of rhythmic flies was higher in the strains of group 1 (88.7%) than in strain 26OL8 (group 2; 75.9%).
Free-running Locomotor Activity Rhythms of the Females
Most of the females (60%-100%) were totally arrhythmic, showed weak rhythm, or did not move at all on some days during the 12-day monitoring period in constant darkness (Fig. 1A,C; Suppl. Fig. S2A,C; Suppl. Fig. S3A,C; Table 1). Approximately half of the studied flies had an inactive period of about 3 days right after the release in constant darkness, which might have increased their arrhythmicity in DD compared with LL (see Fig. 2B,D; Suppl. Fig. S3B,D; Suppl. Fig. S3B,D). Mann-Whitney U test showed that the flies moved less per 30-min bin in constant darkness (median = 0.93, n = 526, number of bins = 288) than in constant light (median = 2.92, n = 584, number of bins = 288) (U = 2270, Z = −19.630, p < 0.001, r = 0.82).
Locomotor Activity Rhythm Parameters of Drosophila montana in Different Environmental Conditions
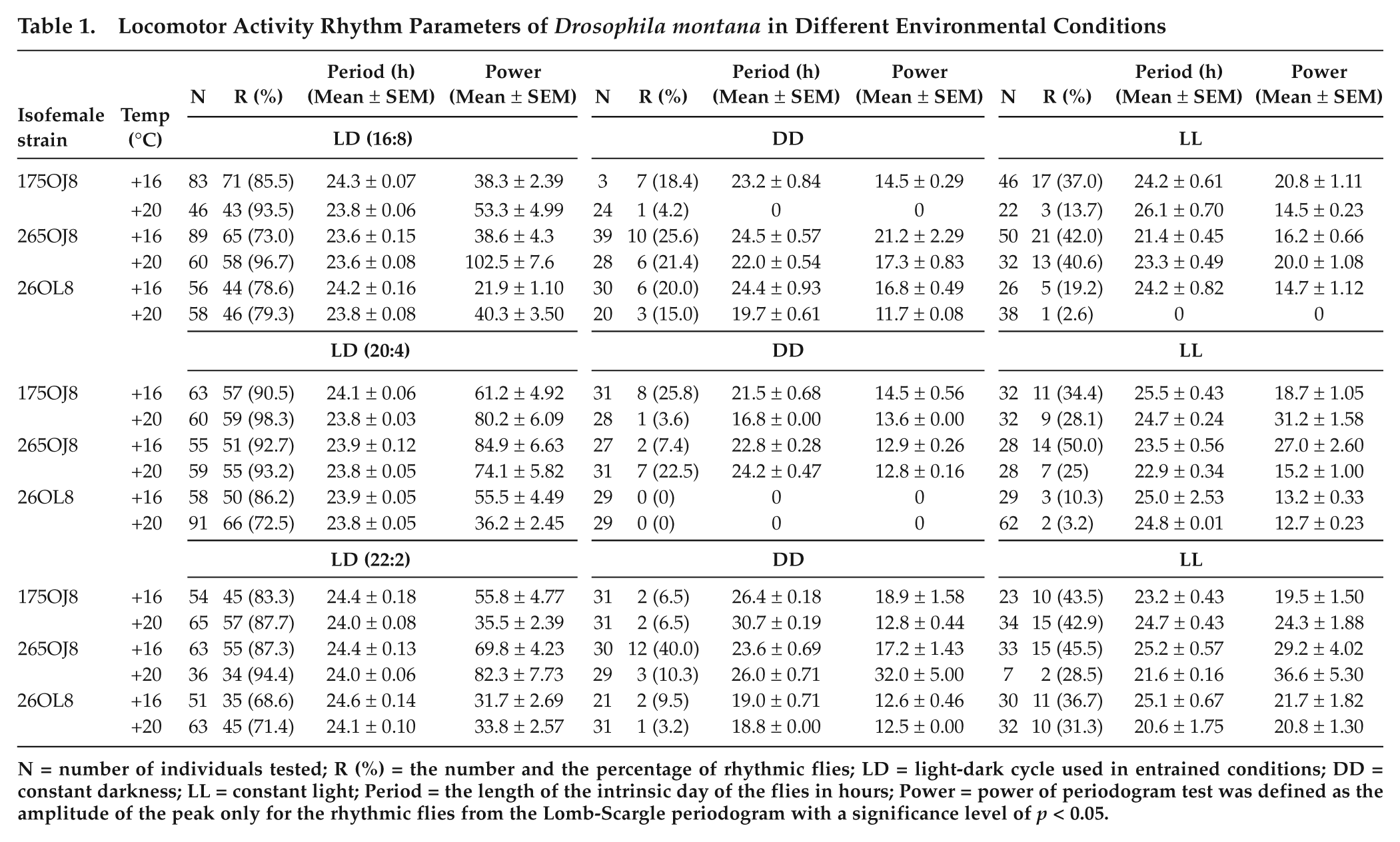
N = number of individuals tested; R (%) = the number and the percentage of rhythmic flies; LD = light-dark cycle used in entrained conditions; DD = constant darkness; LL = constant light; Period = the length of the intrinsic day of the flies in hours; Power = power of periodogram test was defined as the amplitude of the peak only for the rhythmic flies from the Lomb-Scargle periodogram with a significance level of p < 0.05.
When testing factors affecting the free-running rhythmicity in the locomotor activity of the females in constant darkness or light, we ended up with the multinomial logit model: free-running rhythmicity × temperature + free-running rhythmicity × entraining rhythmicity × constant condition treatment + free-running rhythmicity × photoperiod × constant condition treatment × group (LR = 31.6, df = 33, p = 0.54). Flies of both groups were more rhythmic at the lower temperature (27.4% of the flies were rhythmic at 16 °C and 16.0% at 20 °C). The analysis revealed also significant differences between groups 1 and 2 in the ability of the flies to retain their rhythmicity in constant darkness and/or constant light (Table 1, Suppl. Fig. S6). The flies of group 1 retained their rhythmicity better in constant light (proportion of rhythmic flies 37.3%, n = 137) compared with constant darkness (proportion of rhythmic flies 16.7%, n = 61), no matter which photoperiod they confronted before being released into constant light. However, the flies of strain 26OL8 showed significant differences in the proportion of rhythmic flies in constant light and darkness only when they had been entrained in photoperiod 22:2 LD (proportion of rhythmic flies 33.9%, n = 21 in constant light and 5.8%, n = 3 in constant darkness) (Suppl. Fig. S6).
In photoperiods 20:4 LD and 22:2 LD, all females had developed ovaries and were sexually mature at the end of the experiment. However, females that had been kept in photoperiod 16:8 during the entraining conditions developed ovaries only if they had been released in continuous light afterward.
PDF and CRY Immunoreactivity in D. montana
We used PDF and CRY immunostaining to detect PDF- and CRY-positive neurons in the brain of D. montana females (Fig. 3). The study showed PDF to be expressed at circadian time (CT) 0 in l-LNv–like neurons and some neurons in the dorsal brain that seem not to correspond to clock neurons (Hermann et al., 2012). However, the s-LNv–like neurons, which are PDF positive in D. melanogaster (Helfrich-Förster, 1995), showed weak or no expression in D. montana. CRY was expressed at CT 0 in the s-LNv–like neurons, putative LNds, and DNs (see Fig. 3). However, we did not find CRY-expressing l-LNv–like neurons in D. montana.
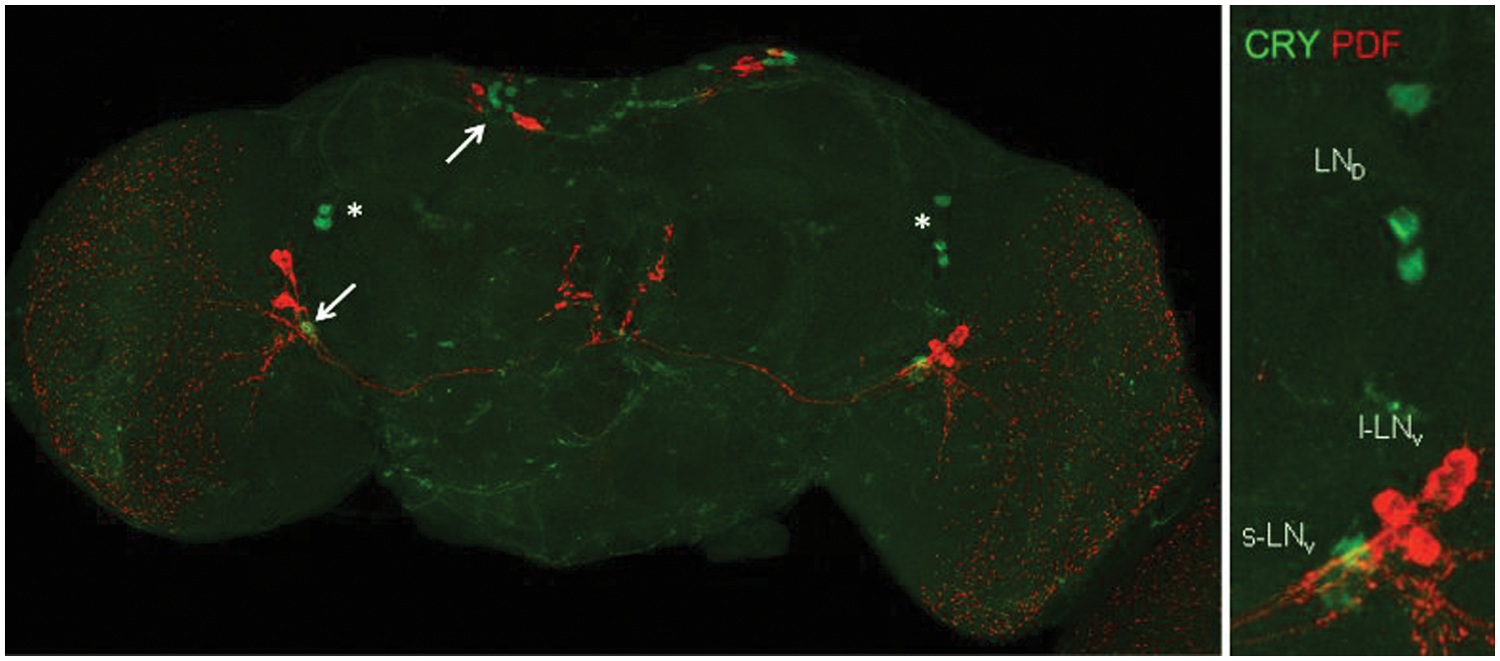
PDF and CRY immunoreactivity pattern in Drosophila montana’s adult central neuronal system. PDF-expressing neurons are colored in red and CRY-expressing neurons in green. PDF-positive large ventrolateral (l-LNv)–like neurons as well as some PDF-positive neurons in the dorsal brain are stained in D. montana. Small ventrolateral (s-LNv)–like neurons are revealed only with CRY staining in D. montana, as well as CRY-expressing putative dorsolateral neurons (LNds) and dorsal neurons (DNs). PDF-positive l-LNvs and additional PDF-positive neurons in the dorsal brain are indicated by arrows; CRY-expressing LNds are marked with an asterisk (*).
Discussion
We have studied the function of the circadian clock in a northern Drosophila species, D. montana, by tracing the entrained and free-running locomotor activity rhythms of the flies in different environmental conditions and identified some brain neurons expressing PDF and CRY. The locomotor activity patterns of these flies appeared to differ from those of D. melanogaster (Allada and Chung, 2010) in several respects, showing more similarity to those of Musca domestica (Helfrich et al., 1985) and D. virilis (Bahn et al., 2009). The study also shows how the interaction between environmental cues and the circadian clock of D. montana may enable the insects to survive in seasonally varying environmental conditions at high latitudes.
Like D. virilis, D. montana flies were active only in the evening and did not display a bimodal locomotor activity pattern in entrained conditions typical, for example, to D. melanogaster flies (Dubruille and Emery, 2008). D. montana resembles D. virilis (Bahn et al., 2009) also in that the flies of this species lose their rhythmicity in constant darkness. In fact, D. montana flies retained their rhythmicity significantly better in constant light than in darkness even with quite high light intensity (~150 lux). This kind of behavior differs completely from that of D. melanogaster flies, which exhibit an evening activity peak in constant darkness (Helfrich-Förster, 2000; Dubruille and Emery, 2008) and become rapidly arrhythmic in constant light probably due to the degradation of TIM protein in response to light mediated by CRY (Konopka et al., 1989; Stanewsky et al., 1998; Emery et al., 2000a). Rhythmicity of D. montana flies in LL is likely an adaptation to the long (even continuous) days during the flies’ mating season in early summer in the northern latitudes. Arrhythmicity, on the other hand, can be adaptive during the long and dark winters at high latitudes, when the flies overwinter under a snow cover.
In entraining photoperiods, the mean activity level of the flies of group 1 (strains 175OJ8 and 265OJ8) and group 2 (strain 26OL8) was affected in different ways by the temperature and photoperiod and their interaction. In free-running conditions, only the flies of group 1 retained their rhythmicity better in constant light than in darkness after entrainment in all photoperiods. These findings are interesting in light of our earlier data, where the flies of strain 26OL8 were found to develop faster than the flies of other D. montana strains and where their propensity to diapause appeared to be lower compared with other strains from the same locality (Salminen et al., unpublished data). The findings suggest that the circadian clock of this strain may have some functional impairments (e.g., in the light signaling input pathway) and that these deficiencies may be reflected also in other life-history traits.
The role of the circadian clock in controlling seasonal changes in insect behavior, alone or together with a photoperiodic timer, is still unclear (Schiesari et al., 2011). Based on the general model proposed by Pittendrigh and Daan (1976), the circadian clock consists of 2 oscillators, which control the morning (M) and evening (E) activity of organisms, and seasonal changes in the phase relationship between M and E oscillators enable the organisms to synchronize their behavior with the seasonal changes in day length. For example, in D. melanogaster, the morning activity peak is advanced and the evening activity peak delayed under long day conditions, while under short day conditions, these activity peaks are closer to each other (Rieger et al., 2003, 2012; Yoshii et al., 2009; etc.). Based on the current view, the neurons of the M-oscillators maintain circadian rhythmicity under constant darkness and short winter-like days (Stoleru et al., 2007; Picot et al., 2007), while the cells belonging to the E-oscillator control the rhythmicity in constant light and under long photoperiods during summer-like days (Stoleru et al., 2004, 2007; Picot et al., 2007). The lack of the morning activity peak in entrained rhythms and the degradation of free-running activity rhythm in constant darkness strongly suggest that the M-oscillators of D. montana do not function in the same way as in D. melanogaster. On the other hand, the E-oscillators may have an important role in maintaining the rhythmicity of these flies in constant light.
Also, the findings on the expression of PDF and CRY in the brain neurons of D. montana flies may refer to a modified function of the M-oscillators. As in D. virilis (Bahn et al., 2009), PDF was not expressed (or showed very low expression) in D. montana in s-LNv–like neurons, which modulate the morning activity in D. melanogaster. The situation resembles that of D. melanogaster Pdf01-mutants, which do not express PDF in their s-LNv–like neurons and also lack morning activity (Renn et al., 1999). Furthermore, ablation of PDF-positive s-LNv neurons in D. melanogaster has been found to result in arrhythmic behavior in constant darkness and the loss of the morning anticipation in laboratory conditions (Shafer and Taghert, 2009). Interestingly, a recent study revealed a complete lack or reduced PDF expression in s-LNv–like neurons also in Drosophila littoralis and Drosophila ezoana (2 species belonging to D. virilis group) (Hermann et al., 2012). The comparison of the amino acid sequence for the PDF antibody region between D. melanogaster, D. virilis, and D. montana showed this region to be identical between the 3 species, and so this finding is not likely to be due to an inability of D. melanogaster’s antibody to detect the same peptide in a different species.
Our study revealed some peculiarities in D. montana also in the expression of CRY, which has been found to be responsible for the arrhythmicity of D. melanogaster flies in LL (Emery et al., 2000a). In the latter species, rhythmic behavior of cryb and cry0 mutants (Emery et al., 2000a; Dolezelova et al., 2007) in LL has been suggested to be due to reduced CRY signaling in the dorsal clock neurons (Dubruille et al., 2009; Zhang et al., 2010a, 2010b). However, the possibility that also the absence of the expression of CRY in the l-LNv plays a part in this phenomenon cannot be excluded as light-arousal and circadian photoreception circuits appear to intersect at the l-LNv (Shang et al., 2008). Furthermore, Emery et al. (2000b) have shown that an overexpression of CRY in the PDF neurons makes flies more sensitive to light pulses, and Fogle et al. (2011) have reported that CRY is necessary for the neuronal firing of the l-LNv upon exposure to bright light. Interestingly, we did not find CRY-expressing l-LNv neurons in D. montana, similar to the situation in D. virilis, D. littoralis, and D. ezoana (Hermann et al., 2012), which suggests that the absence of the expression of CRY in the l-LNv may enable them to retain the rhythmicity in continuous light conditions during northern summers.
Attempts to use antibodies designed against D. melanogaster PERIOD and TIMELESS proteins to stain cells in D. montana (H. Kauranen et al., unpublished data) or D. virilis, D. littoralis, and D. ezoana (Bahn et al., 2009; Hermann et al., 2012) have been unsuccessful. However, staining with D. melanogaster antibodies against vrille (VRI) and PAR-domain protein 1 (PDP1) in D. virilis, D. littoralis, and D. ezoana has shown that all clock neurons known for D. melanogaster are well conserved in their number and location even in species that are not closely related to D. melanogaster (Hermann et al., 2012), and thus the absence of CRY in the l-LNv neurons in these species (and D. montana) is not due to the lack of these neurons. The absence of CRY staining in l-LNvs in D. montana is not likely to be due to a lack of function of D. melanogaster antibody in the above-mentioned species either, as the amino acid sequences for CRY antibody region show high sequence similarities between D. melanogaster and D. virilis (82.9%) and D. melanogaster and D. montana (81.3%). The role of CRY in the rhythmicity in LL also has been brought up in honey bees (Bombus terrestris), which are known to stay rhythmic in constant light (Stelzer et al., 2010) and possess a cryptochrome gene encoding vertebrate-like protein CRY2 that differs from the Drosophila one (see Stelzer et al., 2010). The same kind of protein also has been found in mosquitoes (Anopheles gambiae) and butterflies (Danaus plexippus) (Zhou et al., 2005).
Variation in the function of the circadian clock between the species of Drosophila and the Sophophora genus suggests that the circadian clock has evolved into different directions in different evolutionary lineages and that it may play an important role in adaptation to various kinds of environmental conditions. Interestingly, fly activity rhythms that are under the control of the circadian clock have also been found to show variation within species. Vanlalhriatpuia et al. (2007) have found some strains of Drosophila helvetica to have a unimodal and others a bimodal activity peak in LD, while all strains had unimodal peaks in DD (periods of the free-running rhythms 21.7-26.1 h). The evolution of the “unimodal clock” of the D. virilis group species may have been one factor allowing several species of the group to distribute in high latitudes or altitudes. Understanding the function of the circadian clock of these species will enable detailed studies on the connection between the circadian clock and the photoperiodic timer controlling photoperiodic responses such as female diapause behavior.
Footnotes
Acknowledgements
The work was funded by The Finnish Academy (project 40014) to Anneli Hoikkala. C.H.F. and R.C. acknowledge financial support from the European Community (the 6th Framework Project EUCLOCK no. 018741). We thank M. Karjalainen for the help with the fluorescence microscopy and C. Hermann with immunohistochemistry protocol. J.H. Jennings is acknowledged for reviewing the language and V. Hoikkala and D.J. Parker for help with preparing the figures and checking PDF and CRY antibody sequence similarities. We thank also all persons involved in performing the behavioral experiments. We thank Takeshi Todo (Osaka University, Osaka, Japan) for CRY antibody and Justin Blau for PDF antibody (DSHB, University of Iowa) used in this study.
Conflict of Interest Statement
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Notes
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.