
Abstract
Many organisms change their activity on moonlit nights. Even the fruit fly Drosophila melanogaster responds to moonlight with a shift of activity into the night, at least under laboratory conditions. The compound eyes have been shown to be essential for the perception of moonlight, but it is unknown which of the 5 rhodopsins in the eyes are responsible for the observed moonlight effects. Here, we show that the outer (R1-R6) and inner (R7 and R8) photoreceptor cells in a fly’s ommatidium interact in a complex manner to provoke the moonlight effects on locomotor activity. The shift of the evening activity peak into the night depends on several rhodopsins in the inner and outer photoreceptor cells. The increase in relative nocturnal activity in response to moonlight is mainly mediated by the rhodopsin 6–expressing inner photoreceptor cell R8 together with the rhodopsin 1–expressing outer receptor cells (R1-R6), whereas just rhodopsin 1 of R1 to R6 seems necessary for increasing nocturnal activity in response to increasing daylight intensity.
Light is the most important environmental time cue to synchronize endogenous clocks to the cyclic environment. Consequently, the clocks of most species are very sensitive to light, especially to nocturnal light. The 29.5-day moonlight cycle is used by some animals as a zeitgeber for semilunar and lunar reproductive cycles; others modify their daily pattern of activity, foraging, predation, and communication on moonlit nights (Kronfeld-Schor et al., 2013). Moonlight can increase the activity of nocturnal mammals, as shown for the lemur Eulemur fulvus albifrons and the owl monkey Aotus azarae boliviensis (Erkert and Cramer, 2006), and it can affect the sleep of diurnal mammals as revealed for humans (Cajochen et al., 2013).
Moonlight does also significantly influence the activity pattern of diurnal fruit flies in the laboratory (Bachleitner et al., 2007; Kempinger et al., 2009). Moonlight (0.03 lux) exposure at night causes significant phase shifts of the flies’ morning (M) and evening (E) activity into the night (Bachleitner et al., 2007). In addition, moonlight stimulates nocturnal activity dramatically (Kempinger et al., 2009). The phase shifts of M and E activity under light-moonlight (LM) cycles correlate with phase shifts of the molecular clock (oscillations of the clock proteins PERIOD and TIMELESS) in respective M and E clock neurons, indicating that the observed moonlight effects on activity are at least partially mediated by the circadian clock (Bachleitner et al., 2007). In contrast, the circadian clock turned out to be dispensable for the nocturnal activity increase during moonlit nights because clock mutants respond in the same way (Kempinger et al., 2009). Such direct light effects on activity that bypass the clock are known as masking effects (Mrosovksy, 1999). Masking complements the circadian clock in fine-tuning activity patterns in response to environmental stimuli and therefore represents an important pathway for proper synchronization of the activity rhythms to the day-night cycles (Hut et al., 1999). Although it is doubtful that fruit flies are nocturnal under full moon conditions in nature (Bachleitner et al., 2007; Vanin et al., 2012), Drosophila’s activity patterns in the laboratory can be used as a powerful tool to understand the effects of dim light on activity rhythms.
The moonlight effects on Drosophila’s activity have previously been shown to depend on functional compound eyes and not on the blue-light photopigment cryptochrome (Bachleitner et al., 2007). Yet, it is still unknown which photoreceptor cells and rhodopsins in the eyes are responsible for moonlight detection. The flies’ compound eyes consist of approximately 800 hexagonal ommatidia, containing pigment cells as well as 8 receptor cells (Rister et al., 2013). Receptor cells 1 to 6 (R1-R6) are arranged in the periphery of each ommatidium, span its entire length, and express rhodopsin 1 (Rh1), which has a broad sensitivity to blue-green light (Suppl. Fig. S1). Receptor cells 7 (R7) and 8 (R8) are located in the center of the ommatidium, with R7 being arranged above R8. R7 and R8 define 2 subtypes: In the “pale” cluster R7 expresses the ultraviolet (UV)–sensitive rhodopsin 3 (Rh3) and R8 the blue-sensitive rhodopsin 5 (Rh5). In the “yellow” cluster R7 contains rhodopsin 4 (Rh4) that is sensitive to longer UV wavelengths and R8 the green-sensitive rhodopsin 6 (Rh6). The pale and yellow subtypes are statistically distributed in a 30:70 ratio throughout most of the retina, whereby rhodopsin expression is regulated by sophisticated molecular mechanisms (Rister et al., 2013).
To answer the question of which photoreceptor cells and photopigments mediate the flies’ responses to moonlight, we performed a systematic study monitoring the activity of wild-type (WT) flies and selected photoreceptor mutants under 12:12 LD and LM cycles at 4 different daylight intensities. Our results demonstrate an interaction between inner and outer photoreceptor cells in irradiance detection and WT-like timing of activity.
Materials and Methods
Strains and Rearing
As a control, we used the laboratory strain WTCantonS and 2 different strains caught in the wild. The first strain, named WTALA, was isolated in Val Venosta (Alto-Adige, North Italy, 45°N) and is a mixed culture of 37 isofemale lines (Sandrelli et al., 2007; Rieger et al., 2012). The second one was caught in a small village near Würzburg (Lindelbach, Germany, 50°N) and is referred to as WTLindelbach.
The following photoreceptor mutants were used: clieya mutants that lack the entire compound eyes (Bonini et al., 1993) served as negative controls. rh31 rh41 double mutants lack photopigments in photoreceptor cell 7 (R7) (Vasiliauskas et al., 2011), whereas sevenless, sevLY3 mutants lack the entire R7 cell (Benzer, 1967). sevLY3 mutants are additionally devoid of Rh5 but express more Rh6 than WT flies because pale R7 cells are needed to instruct the underlying R8 to become pale (Rh5-expressing) R8 cells (Chou et al., 1999); consequently, all R8 of sevLY3 mutants are of the yellow type (expressing Rh6). rh52 mutants lack Rh5, rendering 30% of R8 out of function (Yamaguchi et al., 2008), whereby rh61 mutants lack Rh6, leading to a loss of function in 70% of the R8 cells (Cook et al., 2003). The latest studies showed, however, an age-dependent de-repression of Rh5 in the rh61 mutant background, leading to a higher level of R8 containing Rh5 as a pigment (Vasiliauskas et al., 2011). Therefore, less than 70% of R8 should be out of function in rh61. To investigate the effects of a total loss of R8 function, we used rh52;rh61 double mutants (Yamaguchi et al., 2008). Due to the additional knockout of rh5, de-repression is not possible in this mutant. To render both inner receptor cells out of function, a quadruple mutant rh52;rh31 rh41 rh61 was generated. Our starting point was the already recombined rh31 rh41 rh61 triple mutant that we generously received from Nina Vogt (New York University, New York, NY). To knock out the function of photoreceptor cells R1 to R6, we used the rh1-null mutant ninaE17 (neither inactivation nor afterpotential E) (Kumar and Ready, 1995), and to eliminate in addition the function of 70% of the R8 cells, we used ninaE17;rh61 double mutants (Yamaguchi et al., 2010). Because many ninaE17 mutants carry in addition a mutation in the gene for the histamine-gated chloride channel ora transientless ort (CG7411), we made sure by PCR that our used ninaE17 and ninaE17;rh61 double mutant were fine with respect to this (see below).
To minimize differences in light sensitivity caused by the genetic background, all mutants had red eyes, and all were crossed into the ls-timeless background (the L-TIM protein expressed only in ls-tim flies is less light sensitive compared to S-TIM, which is expressed in both ls-tim and s-tim flies) (Sandrelli et al., 2007). Of the WT strains, WTCantonS flies were ls-tim, WTALA flies contained a mixture of both tim forms but were predominantly ls-tim, and WTLindelbach flies were s-tim (as determined by PCR) (Rieger et al., 2012). All flies were raised on a Drosophila medium (0.8% agar, 2.2% sugar beet syrup, 8.0% malt extract, 1.8% yeast, 1.0% soy flour, 8.0% corn flour, and 0.3% hydroxybenzoic acid) at 25 °C in LD 12:12.
PCR
The timeless and ort genes of the different strains were partly sequenced using genomic DNA. To distinguish between s-tim and ls-tim animals, we used the following oligonucleotide primers to amplify (and later to sequence) the genomic DNA: sense TAGGTATCGCCCTCCAAG and antisense TAGGCAGCTCCACAATCA. ort1 is a null mutant carrying a deletion of 569 nucleotides overlapping intron 2 (110 bp) and 459 nucleotides of exons 2 and 3 (Iovchev et al., 2002). The deletions lead to loss of the sequence encoding a substantial portion of the N-terminal extracellular domain and the first 2 membrane-spanning segments and also introduce a frameshift. To exclude the ort1 mutation in ninaE17 mutants, we used the following oligonucleotide primers to amplify the genomic DNA: sense AATATGACACAGGAATACCGCCTGCTC and antisense CGTGTCGCTCAGGACGATGTTTATTAG.
Recording
The locomotor activity of the flies was recorded in a homemade system described first by Helfrich-Förster (1998) and refined by Rieger et al. (2007). Briefly, 2- to 4-day-old single male flies were confined to photometer cuvettes, which were placed with one end in an infrared light beam that recorded the number of walk-throughs in 1-min intervals; on the other end of the cuvette, water and sugar were supplied. Locomotor activity was recorded at 20 °C in a climate-controlled chamber. Illumination was provided by “white” light-emitting diodes (LEDs) (Lumitronix LED-Technik GmbH, Jungingen, Germany) and additional neutral density filters (Lee Filters Worldwide, Andover, UK) for fine adjustment of light intensity. The emission spectrum of the LEDs allowed the excitation of Rh1, Rh5, and Rh6 but not of Rh3 and Rh4 (Suppl. Fig. S1). Thus, photoreceptor cell R7 was not activated by our light conditions.
We applied 2 different light conditions, all consisting of 12 hours of day and 12 hours of night. Locomotor activity was recorded for 7 days either in a rectangular LD cycle or in an LM cycle (moonlight intensity = 0.01 lux) at 4 different daylight intensities (10, 100, 1000, and 10,000 lux).
For every experiment, 32 flies of each genotype were recorded. We only used flies surviving the whole experiment for the calculations, so that the number of analyzed flies varies between 17 and 32.
Data Analysis
The raw data were displayed as actograms using ActogramJ (University of Wuerzburg, Germany; Schmid et al., 2011). In the next step, we analyzed the mean activity profiles of single flies using at least 5 days of recording. To create the average day for 1 experiment, the mean values of at least 17 single activity profiles were calculated and smoothened by a moving mean of 11 values (Helfrich-Förster, 2000; Rieger et al., 2003; Rieger et al., 2012). All average days were normalized and plotted the same size using the program QtiPlot (version 9.8.8, Ion Vasilief, Craiove, Romania) for better visualization. For determination of the E peak phase, we smoothed the activity profiles of single flies over a moving mean of 30. We then determined the times of the E peak manually for each single fly as described by Rieger et al. (2003). Mean diurnal and nocturnal activity levels were calculated out of the unsmoothed average days as described by Rieger et al. (2003).
For statistical analysis, we used a 1- or 2-way ANOVA after testing for normal distribution by the Kolmogorov-Smirnov test. If the data were not normally distributed, p values were adjusted by multiplication with 5 according to Glaser (1978).
Immunohistochemistry
To estimate the de-repression of rh5 in the rh61 mutant background, we performed retina stainings in rh61, ninaE17 rh61, and WTCantonS flies. We also checked the distribution of Rh5 and Rh6 in rh31 rh41 and sevLY3 mutants and Rh1 expression in the outer photoreceptor cells in most mutants. The flies were entrained for 4 (oldest age of the flies at the beginning of the behavior experiments) or 11 days (end of experiments, respectively) in LD 12:12 at a light intensity of 100 lux. On days 4 and 11, the whole flies were fixed in 4% paraformaldehyde (PFA) in phosphate buffered saline (PBS) (pH = 7.4). After washing 5 times for 10 min in PBS, we dissected 15 retinas for each genotype and blocked for 20 min in 5% normal goat serum (NGS) in PBS. In the next step, the retinas were incubated over 2 nights in the primary antibody solution containing rat anti–pigment cell–enriched dehydrogenase (PDH) (1:100; gift of C. Montell, University of California, Santa Barbara, CA, USA) (Wang et al., 2012), mouse anti-Rh5 (1:50; gift of S. Britt, University of Colorado, Aurora, CO, USA) (Salcedo et al., 1999), and rabbit anti-Rh6 (1:1000; gift of C. Desplan, New York University, New York, NY, USA) (Tahayato et al., 2003) or anti-Rh1 (1:30; 4C5; obtained from Developmental Studies Hybridoma Bank, Iowa City, IA) including 5% NGS in PBS with 0.1% Triton X-100 (PBST) (pH = 7.4). After washing the retinas 5 times with PBST for 20 min each, the secondary antibody was applied consisting of Alexa Fluor 488 (goat anti-rat), Alexa Fluor 555 (goat anti-mouse), and Alexa Fluor 635 (goat anti-rabbit) each in a dilution of 1:200 in PBST containing 5% NGS. After incubating overnight, the retinas were washed for up to 4 days in PBST until the red eye pigment disappeared. At the end, the retinas were embedded in Vectashield mounting medium (Vector Laboratories, Burlingame, CA) in a way that the cornea of the eye laid on the glass slide.
In a second set of experiments, we investigated the brains of clieya flies to confirm the presence of Hofbauer-Buchner (H-B) eyelets (Hofbauer and Buchner, 1989). For this staining, the protocol described by Hermann et al. (2013) was used. We applied a primary antibody solution consisting of rabbit anti-Rh6 (1:1000) and mouse anti–pigment-dispersing factor (PDF) (1:1000; obtained from Developmental Studies Hybridoma Bank) containing 5% NGS in PBST. Alexa Fluor 488 (goat anti-rabbit) and Alexa Fluor 635 (goat anti-mouse) were used as secondary antibodies in a dilution of 1:200 and 5% NGS in PBST.
Microscopy and Image Analysis
The brains and retinas were analyzed using laser scanning confocal microscopy (Leica TCS SPE, Leica, Wetzlar, Germany). To excite the fluorophores of the secondary antibodies, we used 3 different laser diodes (488, 532, and 635 nm) and obtained confocal stacks of 2 µm thickness. All images were analyzed using the program Fiji (an ImageJ distribution software). In the case of the retinas, the percentage of ommatidia showing Rh5 or Rh6 staining was analyzed by manually counting the number of Rh5- or Rh6-positive R8 of at least 9 retinas. In clieya brains, we analyzed the presence of the H-B eyelets as well as the course of their tracts towards the accessory medulla (aMe) of 30 flies.
Results
WT Behavior
So far, the response to moonlight was only shown for the laboratory WT strain CantonS (WTCantonS) (Bachleitner et al., 2007; Kempinger et al., 2009). To ensure that the behavioral changes in response to moonlight are also observed in other WT strains, 2 additional strains were tested, WTALA and WTLindelbach, which originally stem from 2 different locations in Europe. Furthermore, for the first time, the absolute activity levels of the flies under LD and LM were measured. The recording system used by Bachleitner et al. (2007) and Kempinger et al. (2009) only allowed us to determine whether a fly was active (value of 1) or inactive (value of 0) during 4-min intervals. Thus, only a qualitative judgment of the nocturnal activity increase in response to moonlight was possible. The quantitative analysis of the present study revealed that moonlight significantly increased the nocturnal activity of all 3 WT strains but that the effects appeared less dramatic as compared to the previous study (Fig. 1). Especially, M activity appeared much lower than in the previous studies. This was because the flies crossed the light beam only once or twice within 4 min in the morning but up to 20 times per minute in the evening. The old system could not distinguish between the different activity levels in the morning and evening: Because the flies crossed the light beam at least once per 4 min at both times, an activity level of “1” was recorded (Bachleitner et al., 2007; Kempinger et al., 2009). Due to the low M activity, the phase of the M peak was hard to determine in the present study. Therefore, we decided to analyze only the timing of the E peak (Fig. 1B).
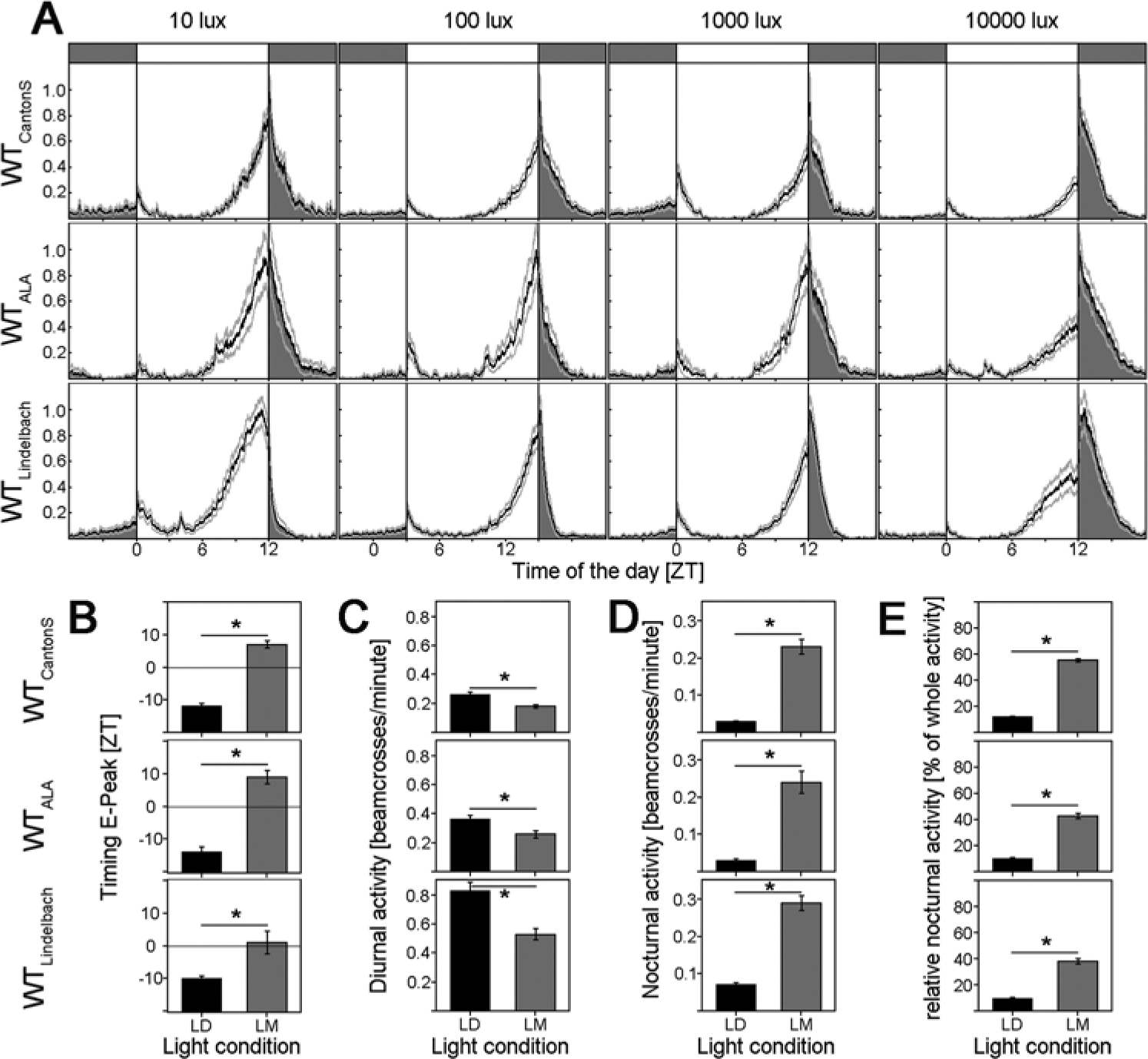
Activity patterns of WTLindelbach, WTALA, and WTCantonS under 12:12 LM cycles of different daylight intensities. (A) Mean activity profiles (±SEM): Activity during the night is indicated in dark gray. From left to right, the light intensity was varied between 10, 100, 1000, and 10,000 lux during the day. All profiles were normalized to 1. (B) Timing of the E peak in LD and LM in minutes after or before lights-off. The value 0 represents the time of lights-off, negative values display the minutes before lights-off, and positive values display the minutes after lights-off, respectively. All WT strains show a significantly delayed E peak in LM compared to LD. The differences range between 15 and 23 min. (C) Mean diurnal activity (±SEM) in LD (black bars) and LM (dark gray bars) calculated as the mean number of beam crosses between ZT0 and ZT12. All genotypes show a significant reduction in diurnal activity upon moonlight simulation. (D) Mean nocturnal activity (±SEM) in LD (black bars) and LM (dark gray bars) calculated as the mean number of beam crosses between ZT12 and ZT0. All genotypes show a significantly higher activity in LM compared to LD. (E) Relative nocturnal activity calculated as the sum of nocturnal activity (exhibited between ZT12 and ZT0) divided by the whole activity of the flies. All 3 WT strains show a significant increase in relative nocturnal activity, which is caused by a decrease in absolute diurnal activity and an increase in absolute nocturnal activity (B, C).
Furthermore, we found that the absolute diurnal and nocturnal activity levels were quite different among the 3 WT strains (Fig. 1C and 1D and Table 1): WTLindelbach was about twice as active as WTCantonS, and WTALA ranged in between the two (Table 1). Similar large differences in the activity level were also revealed for the different photoreceptor mutants (see below). In spite of the differences in overall activity, nocturnal activity increased and diurnal activity decreased in response to moonlight in all 3 WT strains (Fig. 1C and 1D). Therefore, we normalized the activity profiles (maximal activity was set to 1) (Fig. 1A) and calculated the relative nocturnal activity in the percentage of whole daily activity (Fig. 1E) to compensate for differences in absolute activity.
Absolute and relative daily activity levels of the WT strains and the photoreceptor mutants under the LM condition.
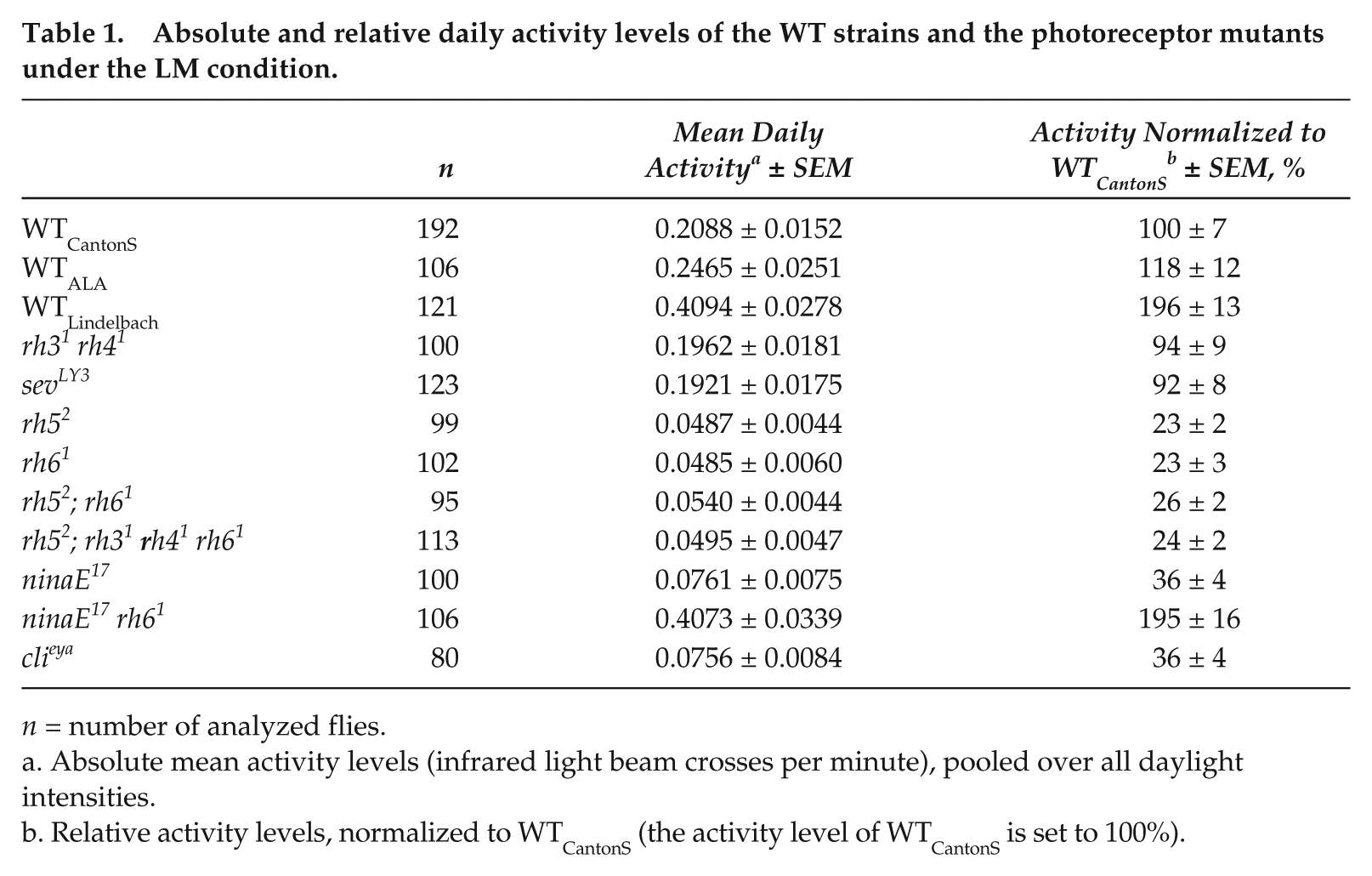
n = number of analyzed flies.
Absolute mean activity levels (infrared light beam crosses per minute), pooled over all daylight intensities.
Relative activity levels, normalized to WTCantonS (the activity level of WTCantonS is set to 100%).
We found the following for all 3 WT strains:
Moonlight simulation (LM) delayed the E peak in comparison to LD. Upon moonlight, the WTCantonS phase delayed the E peak on average by 18.5 ± 1.4 min, WTALA by 22.7 ± 2.7 min, and WTLindelbach by 14.9 ± 3.1 min (Fig. 1B), giving no significant difference between the 3 investigated WT strains of what indicates a minor role of timeless polymorphism on the delay of the E peak on moonlit nights.
Moonlit nights increased nocturnal activity and decreased diurnal activity as compared to dark nights (Fig. 1C and 1D).
The relative nocturnal activity increased with increasing daylight intensity. This was most prominent in WTLindelbach (Fig. 1A) and the least evident but still significant in WTCantonS (Fig. 1A and 4C).
Rhodopsin Expression in the Photoreceptor Mutants
To evaluate rhodopsin expression in the mutants, we performed immunostainings for Rh1, Rh5, and Rh6 in flies that were the same age as our experimental animals (4 days and 11 days). As expected, Rh1 was present in the outer photoreceptor cells of all fly strains except for ninaE17 and ninaE17 rh61 mutants (not shown). The relation between Rh5 and Rh6 expression needed to be determined because de-repression of Rh5 was reported for rh61 mutants (Vasiliauskas et al., 2011), and the absence of pale R7 cells was shown to increase Rh6 expression in R8 (Chou et al., 1999). We found the expected 70%:30% Rh6:Rh5 distribution in WTCantonS flies (Fig. 2A) as well as in rh31 rh41 mutants (Suppl. Fig. S2), whereas sevLY3 expressed Rh6 in approximately 99% of R8 cells (Suppl. Fig. S2). rh61 mutants and ninaE17 rh61 double mutants revealed the expected de-repression of Rh5 that increased slightly with age (Fig. 2A). Nevertheless, the latter appeared less extreme in our flies than was previously reported. We found Rh5 expression in a maximal 50% of R8 cells in 11-day-old flies (Fig. 2A), whereas Vasiliauskas et al. (2011) revealed Rh5 expression in 80% of R8 cells in 14-day-old flies.
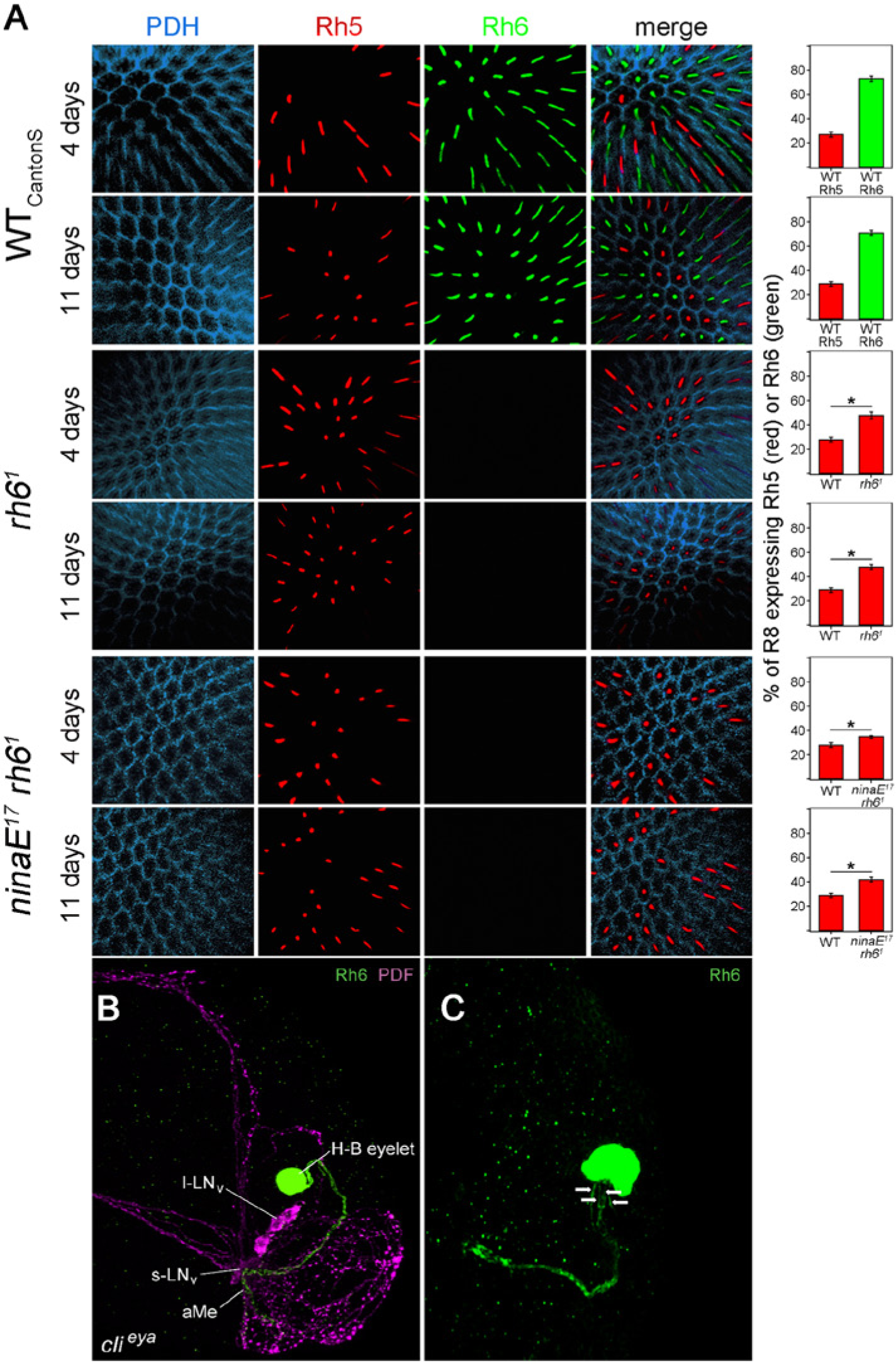
(A) Retinas immunostained with anti-PDH (blue), anti-Rh5 (green), and anti-Rh6 (red) of the genotypes WTCantonS, rh61, and ninaE17 rh61 at the age of 4 and 11 days. Anti-PDH labels the pigment cells surrounding the photoreceptor cells, whereas anti-Rh5 and anti-Rh6 label the rhabdomeres of photoreceptor cell R8 that are located in the centers of the ommatidia. WTCantonS shows Rh5 expression in 30% ± 3% (red bar) and Rh6 expression in about 70% ± 3% (green bar) of R8 as expected from the distribution of pale and yellow ommatidia throughout the retina (Rister et al., 2013), and there was no difference with the age of the flies. In rh61 and ninaE17 rh61 mutants, we could not detect any Rh6 staining, but the number of Rh5-positive R8 cells was significantly increased as compared to WT flies, and this was more pronounced in 11-day-old flies than in 4-day-old flies. (B) The H-B eyelet and PDF-positive lateral neurons (s-LNv and l-LNv) in the right brain hemisphere of a clieya fly. The neurites of the H-B eyelet (green, marked by anti-Rh6) project into the aMe, where they overlap with fibers from the PDF-positive LNv (magenta). (C) As true for WT flies, 1 H-B eyelet of clieya mutants consists of 4 photoreceptor cells. This can be judged from the 4 neurites (arrows) originating from it.
Next, we tested whether clieya mutants still have normal extraretinal H-B eyelets. The H-B eyelets are extraretinal visual organs composed of 4 Rh6-expressing photoreceptor cells that directly project to the fly’s circadian pacemaker center: the aMe (Yasuyama and Meinertzhagen, 1999; Helfrich-Förster et al., 2002) (Fig. 2B). The H-B eyelets have been shown to be present in eyeless flies and to project into the aMe (Hofbauer and Buchner, 1989; Helfrich-Förster et al., 2002), but so far, it has not been investigated whether these always find their targets in the aMe. Therefore, we immunostained 30 clieya brains with anti-Rh6 and anti-PDF. PDF is expressed in Drosophila’s most important circadian clock neurons, and it was shown that the terminals of the H-B eyelets overlap with the PDF neurons in the aMe of WT flies and eyeless so1 mutants (Helfrich-Förster et al., 2002, 2007). We found that clieya mutants possessed 4 H-B eyelet cells per brain hemisphere (Fig. 2C) that had quite variable locations but projected into the aMe in 59 of the 60 investigated hemispheres (Fig. 2B). Thus, there was just 1 brain that was only unilaterally innervated, indicating that photoreception via the H-B eyelets is still functional in clieya mutants.
Contribution of Different Photoreceptors to the Flies’ Responses to Moonlight
To find out which photoreceptor cells (and rhodopsins) in the compound eyes mediate WT-like responses to moonlight under increasing daylight intensities, we recorded the locomotor activity of the different photoreceptor mutants (Fig. 3). WTCantonS flies served as internal controls because these flies were ls-tim as all the mutants. clieya mutants without compound eyes but normal H-B eyelets served as negative controls.
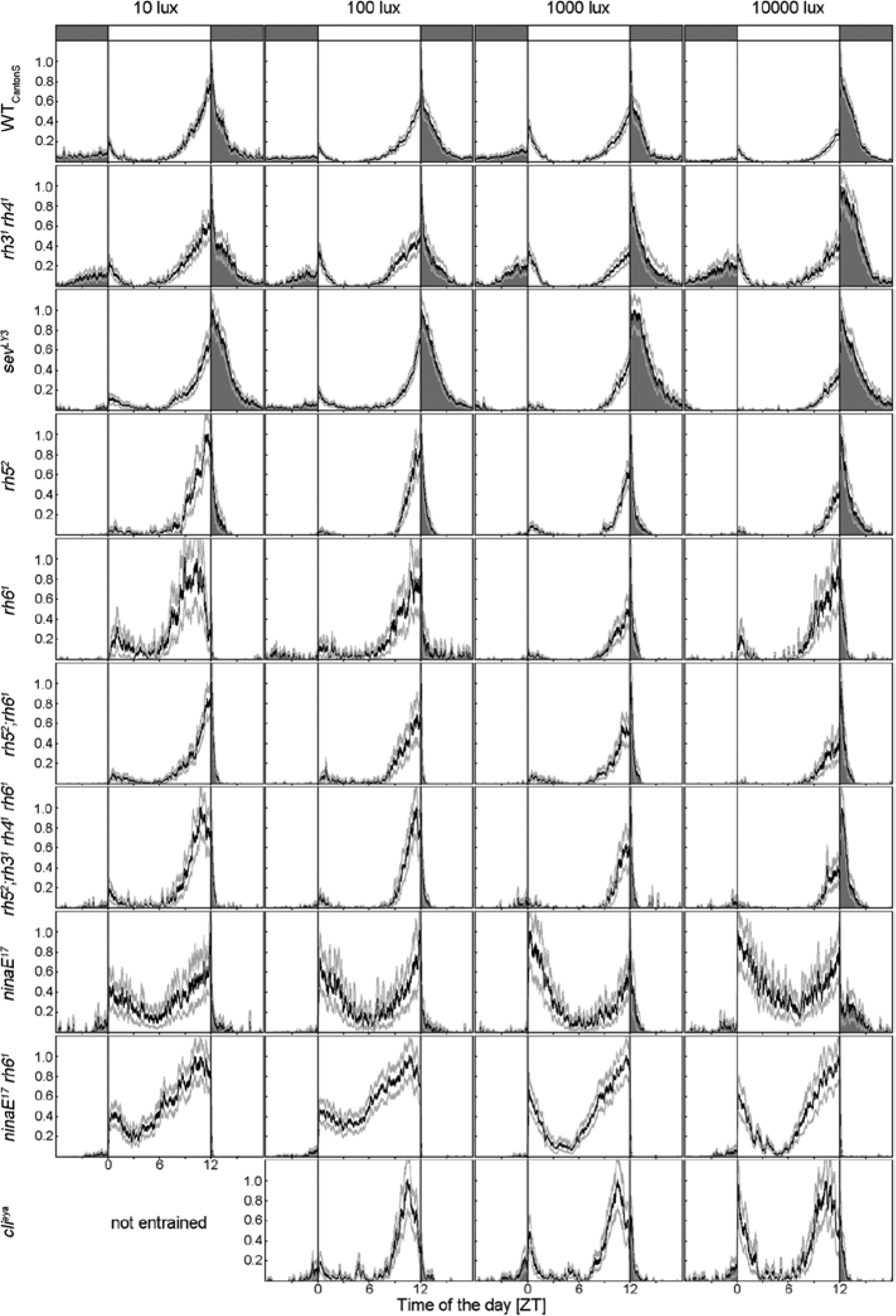
Mean normalized activity profiles (±SEM) of WTCantonS and different photoreceptor mutants under 12:12 LM cycles of different daylight intensities. clieya mutants (bottom) are completely eyeless; rh31 rh41 mutants lack the photopigments in the inner photoreceptor cell R7, and sevLY3 mutants lack the inner photoreceptor cell R7 as a whole; rh52, rh61, and rh52;rh61 mutants lack photopigments in R8; in rh52;rh31 rh41 rh61 mutants, photoreception is impaired in both inner photoreceptor cells (R7 and R8); ninaE17 mutants lack Rh1 in the outer photoreceptor cells (R1-R6); and ninaE17 rh61 mutants lack Rh1 and Rh6. Except for clieya that could not entrain to LM with 10-lux daylight intensity, all genotypes were able to synchronize to all simulated light conditions but differ mainly in their nocturnal activity. For detailed information, see text. Labeling as in Figure 1A.
As found for the WT strains, the different mutants showed quite different activity levels (Table 1). The severely impaired photoreceptor mutants ninaE17 rh61 were most active among the photoreceptor mutants, whereas rh52, rh61, and rh52;rh61 mutants showed very low activity levels, although these lacked just single rhodopsins. We conclude that the overall activity level is probably neither caused by deficits in photoreception nor by the s-/ls-tim polymorphism but by other genetic factors not of interest in the present study and did not consider these further.
Similar to WT flies, all photoreceptor mutants showed a WT-like bimodal activity pattern with a sometimes very weak M activity followed by a midday siesta and a pronounced E activity (Fig. 3). We noticed that the E activity was differently phased in the different photoreceptor mutants: It occurred earlier in the mutants with severely impaired photoreception as rh52;rh31 rh41 rh61, ninaE17, ninaE17 rh61, and clieya under LD and LM (Fig. 3A). This general phase advance of the E peak seemed to depend on photoreceptor input to the clock and will be addressed in detail in another paper (Schlichting et al., in preparation). In the present study, we were mainly interested in the effects of moonlight on the phase of the E peak and the nocturnal activity level. Therefore, we determined for each genotype whether moonlight significantly delayed the phase of the E peak and increased the nocturnal activity level.
Delay of the E peak upon moonlight
ANOVA revealed a significant delay of the E peak in all photoreceptor mutants except the eyeless flies and the ones that lacked photoreception in all inner or all outer photoreceptor cells (rh52;rh31 rh41 rh61, ninaE17, and ninaE17 rh61) (Fig. 4A). We conclude that all photoreceptor cells of the compound eyes contribute to delaying the E peak upon moonlight. Interestingly, photoreceptor cell R7 appeared to influence the delaying effect of moonlight, although this cell was not activated by our light (Suppl. Fig. S1). When comparing the phase of the E peak under moonlight in all mutants that responded with a delay, ANOVA followed by a post hoc test revealed that the time of the E peak was the same in rh31 rh41 and sevLY3 mutants (p = 1.0) and that it occurred significantly later than that of all the other strains (p < 0.003).
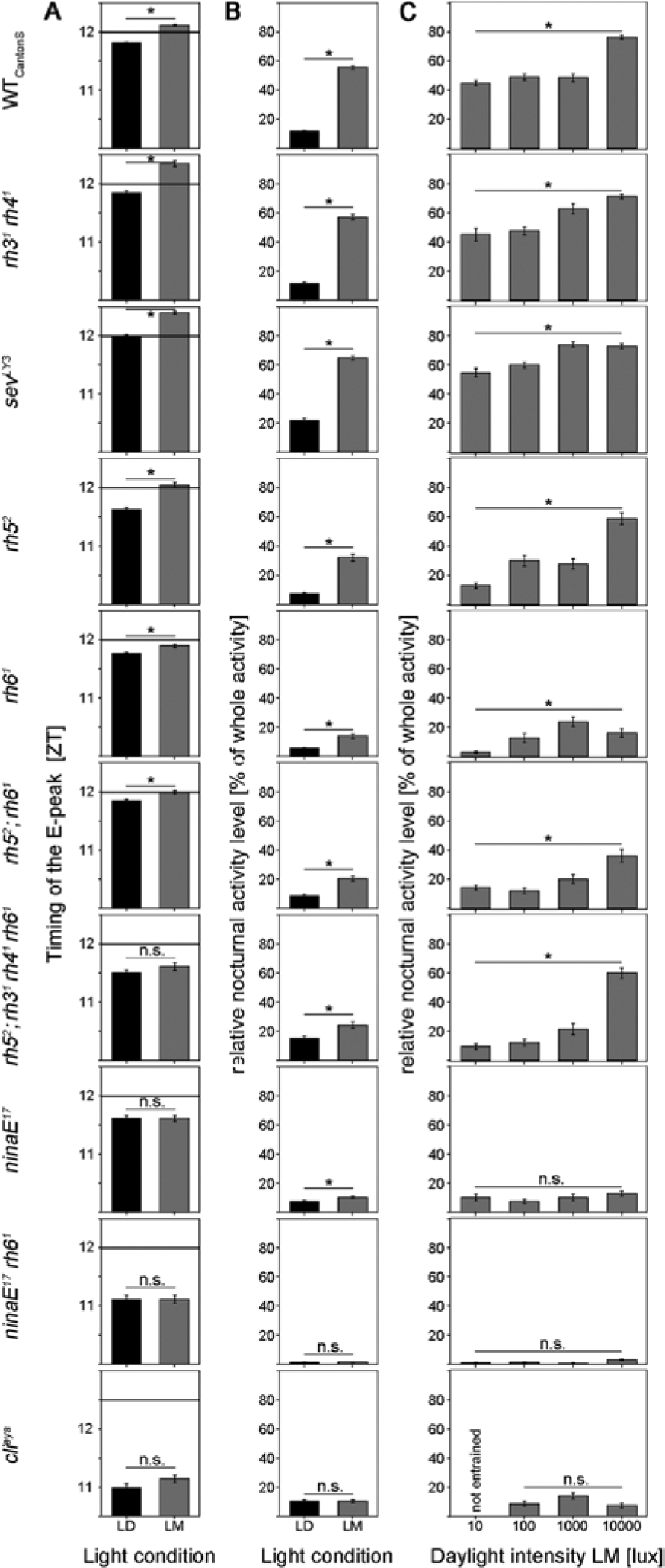
Timing of the E peak and relative nocturnal activity under LD and LM conditions in WTCantonS and 9 different photoreceptor mutants (alignment as in Fig. 2). (A) WTCantonS flies significantly delay the E peak upon moonlight simulation, whereas clieya mutants and mutants with severely impaired photoreception in the inner or outer photoreceptor cells of the compound eyes show no significant difference between LD and LM. (B) The relative nocturnal activity of WTCantonS and most photoreceptor mutants was significantly higher under LM than under LD. Only mutants without compound eyes (clieya) and mutants lacking rhodopsin 1 and 6 (ninaE17 rh61) show no rise in nocturnal activity on moonlit nights, indicating an important role of these 2 rhodopsins within the compound eyes for dim light detection. (C) Relative nocturnal activity in dependence of daylight intensity of 10, 100, 1000, and 10,000 lux. The higher the light intensity, the higher the nocturnal activity in WTCantonS (ANOVA: F3,188 = 64.168, p < 0.001). The same is true for mutants affecting the inner receptors. As soon as the outer receptors are out of function, no increase in the relative nocturnal activity is visible, indicating an important role of R1 to R6 and Rh1 for measuring daylight intensity. See text for further details.
Increase in nocturnal activity upon moonlight
Figures 3 and 4B show that moonlight seemed to stimulate nocturnal activity in a WT manner in the mutants that lacked photoreception in R7 (rh31 rh41 and sevLY3). The 2 mutants without functional photoreception in R7 had even the tendency for a higher relative nocturnal activity as compared to WTCantonS, but this was not significant. In rh52 mutants, moonlight provoked an increase in nocturnal activity that was about half as high as in WTCantonS (Figs. 3 and 4B). In all other mutants, the stimulation of nocturnal activity by moonlight was strongly reduced, or it was completely absent (Fig. 3). The latter was true for ninaE17 rh61 and clieya mutants (Fig. 4B). ninaE17 rh61 double mutants are furthermore exceptional with respect to their extremely low relative nocturnal activity (Figs. 3 and 4), although their general activity level was very high (Table 1). Already, the presence of Rh6 in the H-B eyelets was enough to bring the relative nocturnal activity back to the level of ninaE17, suggesting that the H-B eyelet does also contribute to normal nocturnal activity levels. Overall, we conclude again that all photopigments of the compound eyes contribute to the increase in nocturnal activity upon moonlight, whereby Rh1 and Rh6 seem to be most important.
Dependence of nocturnal activity on daylight intensity
clieya mutants, ninaE17 mutants, and ninaE17;rh61 double mutants did not change the ratio of diurnal/nocturnal activity with increasing daylight intensity (Fig. 4C), indicating that especially Rh1 is needed for measuring daylight intensity.
Discussion
The aim of this study was to investigate the importance of rhodopsins for a WT response to moonlight and increasing daylight intensities. First, we showed that the activity rhythms of 2 independent WT strains responded qualitatively in the same way to high daylight intensities as well as to moonlight simulation as did the laboratory strain WTCantonS: All 3 strains shifted activity into the night when artificial moonlight was present, and this was more prominent the higher the daylight intensity. This is especially interesting because the 3 WT strains expressed different proportions of the short (s) and long (ls) forms of the tim gene that play a major role in the circadian clock’s response to light (Kyriacou et al., 2008). The clock of s-tim flies was shown to be more light sensitive than that of ls-tim flies (Sandrelli et al., 2007; Tauber et al., 2007 ). Consequently, we may expect that the s-tim WTLindelbach flies shift activity stronger into moonlit nights than the ls-tim WTCantonS flies. Yet, this was not the case, suggesting that the input from the photoreceptor cells is more important for the present investigated light effects on the clock than is s-tim or ls-tim. We conclude that the evident abnormal responses to moonlight that we observed in the photoreceptor mutants are most likely due to the absence of particular photoreceptor cells (or photopigments) and not caused by different WT genetic backgrounds.
Second, we showed that the Rh6-expressing inner photoreceptor cell R8 together with the outer photoreceptor cells R1 to R6 are responsible for moonlight detection necessary for phase delaying of the E peak and stimulating general nocturnal activity. These photoreceptor cells are also the ones that finally determine the ratio of diurnal and nocturnal activity. The outer photoreceptor cells R1 to R6 are additionally needed for measuring daylight intensity.
It is already known that photoreceptor cells R1 to R6 are important for dim light detection in motion vision and phototaxis (Heisenberg and Buchner, 1977; O’Tousa et al. 1985; Yamaguchi et al., 2008; Zuker et al., 1985), whereas the inner photoreceptor cells were assumed to be less light sensitive and mainly responsible for color vision (Yamaguchi et al., 2010). This situation reminds us of the scotopic and photopic visual systems in mammals represented by the light-sensitive rods and the color-sensitive cones. Both systems signal via rod and cone bipolar cells on melanopsin-positive retinal ganglion cells that are light sensitive on their own and directly innervate the central clock in the suprachiasmatic nuclei (Hattar et al., 2003). Most interestingly, Altimus et al. (2010) found that, at higher irradiances when the classic rod pathway is not working anymore, rods signal through cones to the cone bipolar cells. This implies a direct interaction between the scotopic and photopic pathways in irradiance detection serving synchronization of the endogenous clock.
In the fruit fly, an interaction between the inner and outer photoreceptor cells was predicted for a long time for the phototactic response (Jacob et al., 1977) and was more recently confirmed (Yamaguchi et al., 2008, 2010). Whereas Jacob et al. (1977) proposed that R8 is inhibiting R1 to R6, Yamaguchi et al. (2010) could not find a general inhibition of the R1 to R6 receptor subsystem by R7/R8. This applies also for our results. If R8 would inhibit R1 to R6, rh52;rh61 mutants should be more light sensitive than WT flies and shift more prominently into moonlight. However, the opposite happened. Furthermore, we found that ninaE17 mutants without functional outer photoreceptor cells R1 to R6 still shift activity into moonlit nights, whereas ninaE17,rh61 double mutants are not able to do so anymore. This clearly indicates that Rh6 in R8 adds to moonlight detection. A direct functional connection between R8 and the outer photoreceptor cells was recently discovered by Wardill et al. (2012). These authors showed by sophisticated genetic manipulations of rhodopsin expression paired with electrophysiological and behavioral recordings that R8 signals either to the neighboring R6 and/or to R1 and by this way add to the optomotor responses elicited by R1 to R6. This supports our hypothesis that R8 and R1 to R6 work in the same direction in terms of dim light detection for synchronizing the flies’ activity rhythms (Fig. 5).
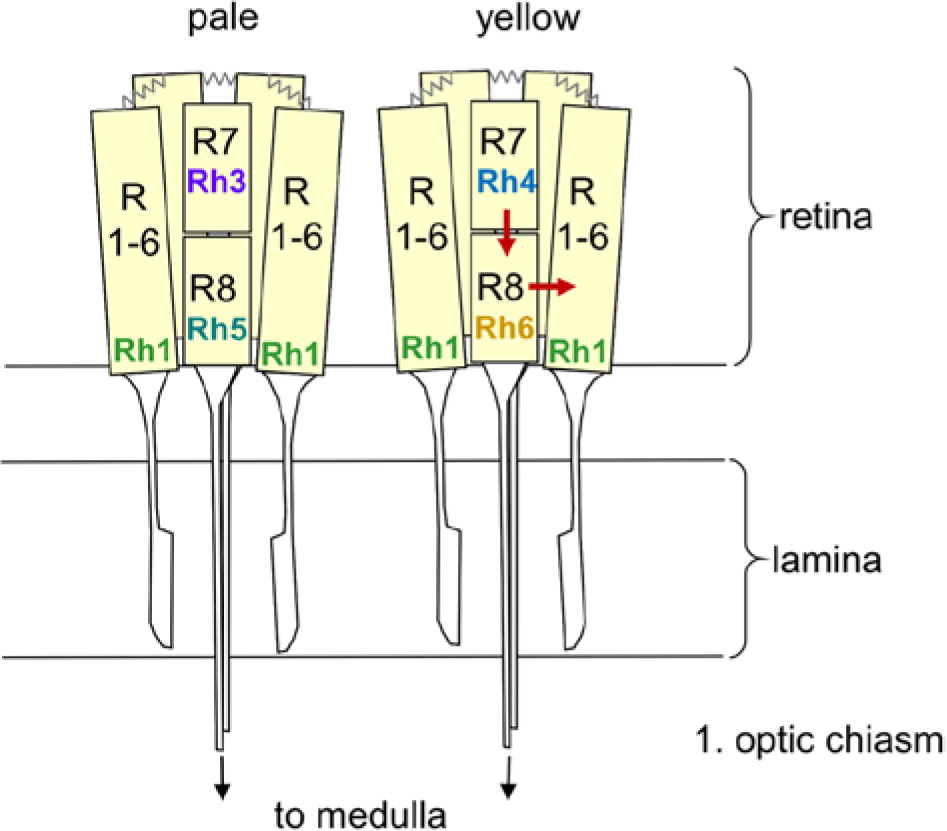
Rhodopsin expression in pale and yellow ommatidia of the fly retina. The 8 photoreceptor cells are shown in a schematic manner with the outer receptor cells R1 to R6 spanning the entire depth of the retina and the inner receptor cells R7 and R8 with R7 arranged above R8. R1 to R6 terminate in the fly’s first optic neuropil, the lamina, whereas R7 and R8 run toward the medulla. R1 to R6 are connected via gap junctions (zigzag lines). The arrows indicate the putative interaction between the Rh6-expressing R8 cell and R1 to R6 as well as between R7 and R8. See text for further details.
Wardill et al. (2012) found also that input from R7 cells converged on the outer photoreceptor cells. Our data do not permit a definitive conclusion about R7 because we did not stimulate the UV-sensitive rhodopsins Rh3 and Rh4 with our “white light” LEDs. Nevertheless, our results suggest that R7 may also be involved in the interaction between R8 (and/or R1-R6) but rather in the opposite direction. After ablation of R7 by the sevenless mutation and after elimination of Rh3 and Rh4, we did not see any reduction in moonlight sensitivity. On the contrary, sevLY3 and rh31 rh41 mutants showed a stronger shift of the E peak into moonlit nights and had the tendency to increase relative nocturnal activity more than WT flies. This result is especially remarkable for sevLY3 mutants because the R7 rhabdomere should serve as a light guide to the R8 rhabdomere: The rhabdomere tip of R7 is positioned in the focal plane of the facet lens, where the light flux is most intense, and subsequently, the light flux is trapped in the rhabdomere in an optical waveguide mode down to the rhabdomere of R8 (Snyder and Menzel, 1975). Without R7, R8 is expected to receive much less light, and as a consequence, photoreception of R8 should be reduced. Because we found the contrary, we have to conclude that at least regarding synchronization of the clock, intact (not excited) R7 cells inhibit R8 and/or R1 to R6 via unknown mechanisms. This inhibition seems to need the presence of the photopigments because it was similarly lost when R7 was present, but just Rh3 and Rh4 were absent. Future studies with light sources containing UV light are necessary to further unravel this interesting interaction.
Another interesting detail of our results is the fact that Rh5-expressing R8 cells seem to contribute only marginally to dim light detection. Although the number of Rh5-expressing R8 cells was increased in the absence of Rh6 (in rh61 and ninaE17 rh61 mutants), these mutants did not increase their response to moonlight. We could also exclude a prominent role of the H-B eyelet in moonlight detection. Although the H-B eyelets may contribute to general phasing of the E peak as well as to the relative nocturnal activity level of the flies, they were clearly neither sufficient for the shift of the E peak into moonlit nights nor for the nocturnal activity increase in response to moonlight.
In summary, we show here that the inner and outer photoreceptor cells of Drosophila’s eyes interact in adapting the flies’ activity patterns to variable diurnal and nocturnal light conditions. In mice, similar mechanisms are present in the retina, and among other factors, these determine whether the animal is nocturnal or diurnal (Doyle et al., 2008; McNeill et al., 2008). The latter may also be true in Drosophila, confirming the fruit fly as a suited model organism to unravel the mechanisms of synchronizing the circadian clock to light.
Footnotes
Acknowledgements
The authors thank Nina Vogt, Claude Desplan, Francois Rouyer, Ralf Stanewsky, Erich Buchner, Rodolfo Costa, and Christopher Schnaitmann for providing fly lines, Christiane Hermann and Pingkalai Senthilan for critical comments on the paper, and the German Research Foundation (DFG; Fo207/10-3 and SFB1047, INST 93/784-1) as well as the European Community (6th Framework Project EUCLOCK no. 018741) for funding. They also thank Craig Montell, Steve Britt, and Claude Desplan for providing antibodies as well as the Graduate School for Life Sciences Würzburg and the Hanns-Seidel-Foundation for supporting Rudi Grebler and Matthias Schlichting, respectively.
Conflict of Interest Statement
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Notes
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.