
Abstract
Circadian clocks are believed to provide the selective advantage of anticipation, thus allowing organisms to respond efficiently to stimuli at the appropriate moment. Disrupted circadian rhythms have been found to affect a variety of basic physiological processes. However, the importance of the circadian clock in regulating heart performance remains undetermined. We hypothesized that the circadian clock plays a crucial role in heart performance through the anticipation of daily workload. Echocardiography was employed to monitor heart function and structure in mice in a noninvasive, real-time manner. In wild-type mice, both the ejection fraction (EF) and the shortening fraction (FS), two important markers of cardiac function, show diurnal variation. In addition, the amplitude of the EF and the FS enlarges in response to forced exercise in a time-dependent manner. The diurnal variations in EF and FS are altered in mice with disruptions in circadian clock genes and are significantly attenuated under an imposed light regimen. Furthermore, it shows that the overexpression of peroxisome proliferator–activated receptor gamma coactivator 1 alpha (Pgc1α) under control of the muscle creatine kinase (MCK) promoter inhibited clock gene expression in the heart and muscle and decreased the expression of peroxisome proliferator–activated receptor alpha (Pparα), metabolic genes glucose transporter (Glut4), and acetyl-coA synthetase (Acs1). Pgc1α overexpression abolished the diurnal variation of EF. We thus propose that PGC1α might play an important role in circadian-mediated, impaired cardiac function by regulating the circadian rhythm of metabolic genes.
The circadian clock plays essential roles in the maintenance of physiological homeostasis and in the coordination of responses to external signals. In mammals, the circadian clock is composed of interlocking positive and negative feedback loops. The CLOCK and BMAL1 proteins form heterodimers and bind to E-box elements in the promoters of the two period and cryptochrome genes (Pers and Crys, respectively) to induce their expression. PER and CRY proteins negatively regulate their own expression by inhibiting the activity of BMAL and CLOCK. The second feedback loop is mediated by two nuclear receptors, RORα and Rev-erbα. Transcription of RORα and Rev-erbα is activated during the day by the CLOCK/BMAL1 complex. RORα and Rev-erbα exert positive and negative transcriptional effects on the Bmal1 gene through Rev-erbα/RORα–responsive elements (ROREs) in the Bmal1 promoter (Lowrey and Takahashi, 2004; Reppert and Weaver, 2002; Young and Kay, 2001). Circadian clocks are crucial not only for daily rhythms but also for maintaining the sleep/wake cycle (Xu et al., 2005; Xu et al., 2007), metabolic processes (Sahar and Sassone-Corsi, 2009; Wang et al., 2010), cardiac function (Young, 2009), vascular disease (Anea et al., 2009; Wang et al., 2008), and other physiological processes. The links between the circadian clock and a variety of physiological processes are just beginning to be unveiled.
Diurnal variation is known to occur in many aspects of cardiovascular function, including blood pressure, coagulation, and contractile function. Diurnal variation has also been observed in important clinical events, such as myocardial infarction and stroke (Durgan et al., 2010). Approximately 8% to 10% of the genes expressed in the heart exhibit a circadian expression pattern, including Pparα, acetyl-coA synthetase (Acs1), and the Glut4 glucose transporter (Anea et al., 2009; Durgan et al., 2006; Durgan and Young, 2010; Rudic, 2009; Storch et al., 2002). However, the biological significance of the circadian regulation of heart genes remains unclear. We hypothesized that if a gene important for heart metabolism (Lin et al., 2002) were itself a clock component, it would be able to provide insight into the molecular signaling pathways involved in this process. We thus theorized that Pgc1α might be a good candidate gene because of its role as a key component of the circadian oscillator that integrates the mammalian clock and energy metabolism (Lin et al., 2002; Lin, 2009; Liu et al., 2007; Sonoda et al., 2007). Therefore, we designed an experimental strategy utilizing genetically engineered mice and altered light regimens aimed at characterizing the molecular links between clock genes and cardiac function. The importance of PGC-1α is highlighted by the observation that PGC1α inhibits the expression of most circadian clock genes and the nuclear receptor genes Rev-erbα, Rorα, and Pparα. On the basis of these molecular changes, it is possible that pharmacological intervention strategies that coactivate PGC1α may hold therapeutic potential for improving cardiac performance.
Materials and Methods
Animal Models
Animal studies were performed in an Association for Assessment and Accreditation of Laboratory Animal Care (AAALAC) International–accredited SPF animal facility, and all animal protocols were approved by the Animal Care and Use Committee of the Model Animal Research Center, the host for the National Resource Center for Mutant Mice in China, Nanjing University. Mutant PER2S662G, PER2S662D (Xu et al., 2007), Pgc1α transgenic (Tg) (Lin et al., 2002), and overtime (Ovtm) (Siepka et al., 2007) mice and their wild-type littermates were tested between 2 and 3 months of age. Each of these lines has been backcrossed onto C57BL/6J mice for at least 10 generations. A minimum of two independent experiments were performed, and three to six mice were tested at each time point. All mice were maintained on a 12:12 light:dark (LD) cycle under the indicated conditions.
Gene Expression Analysis
The animals were housed in a temperature-controlled room with food (a chow diet containing 4% fat) and water available ad libitum. Mice were decapitated every 4 hours for 24 hours under a normal 12:12 LD cycle or constant dark as indicated, and the gastrocnemius and heart muscles were dissected, immediately frozen in liquid nitrogen, and stored at −80 °C. Total RNA was extracted from individual tissues using TRIzol (Invitrogen, Carlsbad, CA) and quantified using a spectrophotometer at OD 260 nm. Random hexamers were used to prime reverse-transcription reactions with Superscript III (Invitrogen). Real-time quantitative PCR was performed using an ABI 7300 with the SYBR green I reagent (Takara Bio Inc., Otsu, Japan).
Forced Exercise and Echocardiography
Mice were randomly assigned to either the sedentary group or the exercised group. The latter was accustomed to treadmill running. The treadmill was placed in the same room in which the mice were housed. Forced exercise was performed during the dark phase under dim red light to prevent light-induced phase shifts in the mice. Mice were acclimatized to the treadmill by running for 10 minutes at 10 m/min the day before the experiment. Every 4 hours over the course of a 24-hour period, mice were placed on a 6-lane treadmill, the speed of which was initially set to 10 m/min for the first 10 minutes and increased by 2 m/min every 2 minutes thereafter. Animals ran until exhaustion, which was defined as remaining on the shocker plate for more than 10 to 15 seconds. Echocardiography was performed on mice immediately after acute exercise or without exercise. Animals were anesthetized with 2.5% Avertin (10 µL/g, Sigma), their body temperature was maintained at 37 °C using a heating pad, and respiration was spontaneous. Echocardiography was performed using a Vevo 770 UBM (VisualSonics, Toronto, ON, Canada) that possessed a single-element mechanical transducer with a center frequency of 30 MHz and a frame rate of 30 Hz. Parasternal short-axis images of the heart were obtained by standard echocardiographic views. Two-dimensional M-mode images were acquired, and the measurements included the interventricular septum, the left ventricular internal diameter, the left ventricular posterior wall, and the ejection fraction.
Hematoxylin and Eosin Staining
Mice hearts were dissected and fixed in 4% PFA, embedded in paraffin, and sectioned. The sections were then stained with hematoxylin and eosin and were imaged using microscopy.
Statistics
To assess temporal changes, we calculated a fitted cosinor curve with a 24-hour rhythm based on the measured values. The cosinor curve was represented as y = M + A cos (ωt + φt), where M = MESOR, A = amplitude, ω = 2π/24, and φ = acrophase. The MESOR is the cosine-determined, overall 24-hour mean. The amplitude was defined as one half of the total excursion best approximating the rhythm. The acrophase represents the crest of the fitted cosine curve. Fluctuations in the variable studied were presumed to be cyclic and not random. The significance of the circadian rhythms was determined using the zero amplitude test. One-way analysis of variance (ANOVA) followed by Scheffe multiple comparison or Student t test was used to determine the statistical significance of observed differences in measured parameters. p < 0.05 was considered statistically significant.
Results
Cardiac Ejection Fractions Exhibit Daily Variations
To determine whether the heart has a 24-hour fluctuation in its adaptations to daily variations, ensuring an optimal response, echocardiography was employed to analyze the structure and function of the heart in sedentary and exercised mice at indicated zeitgeber time (ZT) points. The zero amplitude test was employed to clarify whether ejection fraction (EF) and shortening fraction (FS) exhibited significant circadian rhythms (fitting a cosinor curve). As shown in Figure 1A, EF, an indicator of heart contractile function measured by echocardiography, exhibited rhythmic changes over a 24-hour period in mice. FS, another marker of heart contractile function, exhibited a similar rhythmic pattern (Fig. 1B). The 4-hour-interval EF and FS data were fit to cosinor curves, and the MESOR, amplitudes, and acrophase were determined in sedentary mice with significant circadian rhythms (Suppl. Table S1). In response to forced exercise (running on a treadmill with increasing speed, 2 m/min every 2 minutes until exhaustion), both EF and FS showed significant fits to cosinor curves. The EF amplitude of exercised mice was significantly higher than that observed in sedentary mice (exercise: 3.83 ± 0.68 v. sedentary: 2.66 ± 0.69, p < 0.05), as was the MESOR (exercise: 64.33 ± 1.45 v. sedentary: 62.2 ± 1.03, p < 0.05) (Fig. 1C and 1D and Suppl. Table S1). In contrast, FS did not differ between sedentary and exercised mice. The exhibited rhythm was not correlated to the running distances at ZT 0 (361 ± 23 m) and ZT 12 (362 ± 17 m). Thus, the observed changes in EF did not result from different running distances. The variance in heart rate after forced exercise was not significantly different during the day compared with the night under our anesthesia conditions (Suppl. Fig. S1), which indicated that the state of anesthesia was similar among all mice in the experiment. We thus concluded that the timing of episodes of heart contractile function is the major contributor to the observed diurnal variation in EF. However, we could not fully exclude the possibility that anesthesia sensitivity over a 24-hour period may also affect EF variation.
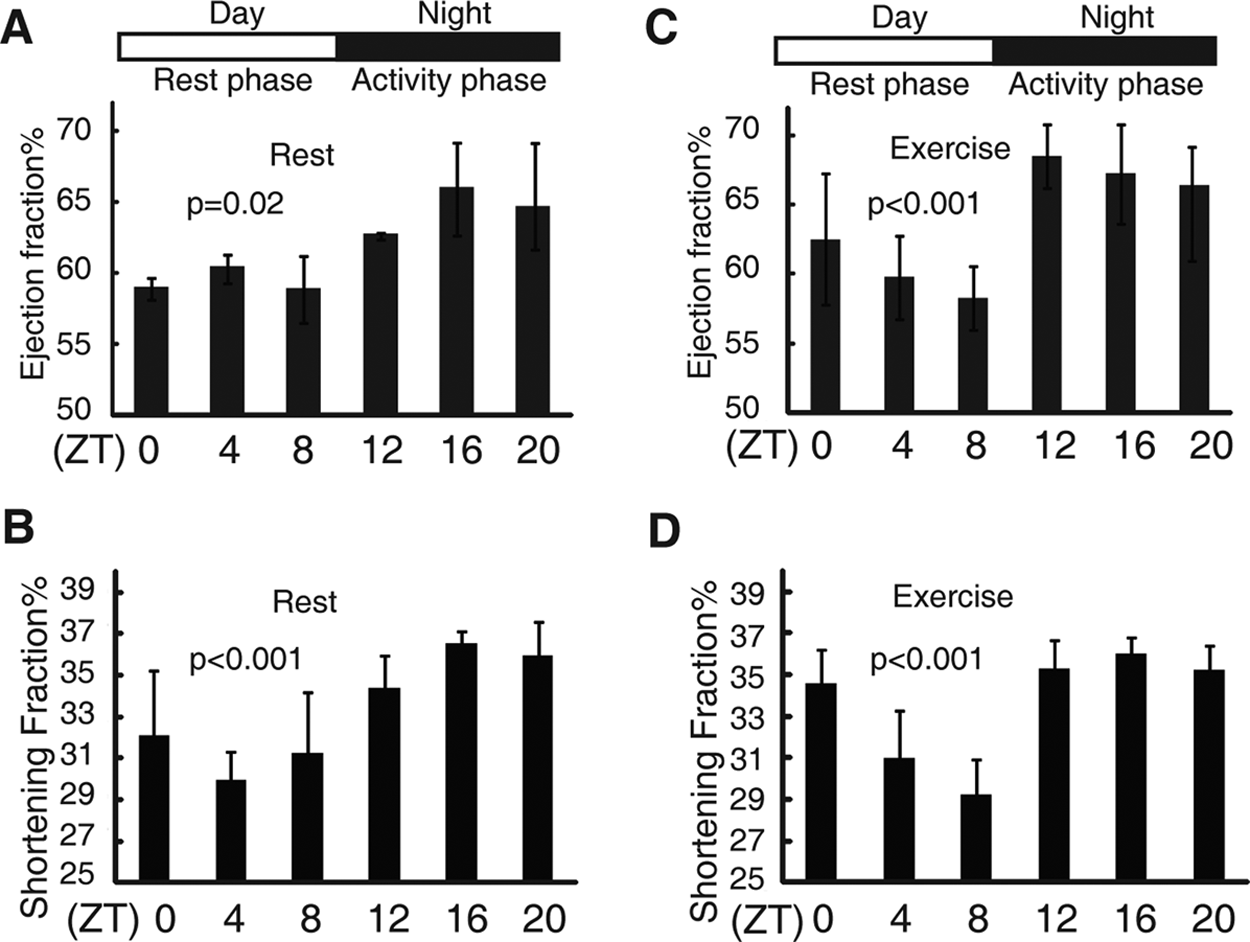
Diurnal variations in the ejection fraction (EF) and the shortening fraction (FS) of mice. Diurnal variations in the EF of wild-type mice under sedentary (A) and forced exercise (C) conditions. Diurnal variations in the FS of wild-type mice under sedentary (B) and forced exercise (D) conditions. Nonlinear regression was performed to fit cosinor curve to the observations. p < 0.05 indicated observations significantly fits to the curve. Values are shown as the mean ± SEM; n = 4 to 6 observations for each time point during the 24-hour period. The data shown are representative examples from at least two independent experiments with different cohorts of mice.
Disruption of the Circadian Clock Impairs Diurnal EF Variation
To further determine whether “anticipated” diurnal variations in heart contractile function are mediated by circadian rhythms, we examined the diurnal EF and FS variations in response to exercise in mice with various circadian periods. PER2S662G and PER2S662D transgenic mice are unique in that a single amino acid change in PER2S662 can either shorten or lengthen the circadian period (Xu et al., 2007), thus providing us with a unique opportunity to evaluate whether an event is circadian dependent or gene dependent. The acrophase of EF was significantly advanced from 0.52 ± 1.32 in wild-type mice to 21.48 ± 0.8 in PER2S662G mice that have a short circadian period (21.6 hours), as was the FS (1.16 ± 0.94 v. 20.66 ± 1.06) (Fig. 2E and Suppl. Table S1). However, the EF and FS were not rhythmic in PER2S662D mice, which exhibited only a single peak at ZT 16, suggesting that PER2S662 mutations may not only affect the phase of EF but may also contribute to impaired cardiac contractile function (Fig. 2A and 2B). Thus, Ovtm mice, which exhibit a τ = 26.2 hours, were employed to observe whether phase delays of EF and FS were associated with period lengthening (Siepka et al., 2007). The acrophases of EF and FS were significantly delayed to 3.75 ± 0.78 for EF and 4.83 ± 0.53 for FS in Ovtm mice (Fig. 2A, 2B, and 2E and Suppl. Table S1). These results suggest that “anticipated” diurnal variations are circadian-dependent events. To exclude the possibility that these circadian mutant mice exhibited alterations in cardiac structure and function, histological and echocardiographic analyses were employed. We found no obvious genotype differences (Suppl. Fig. S2 and Table 1).
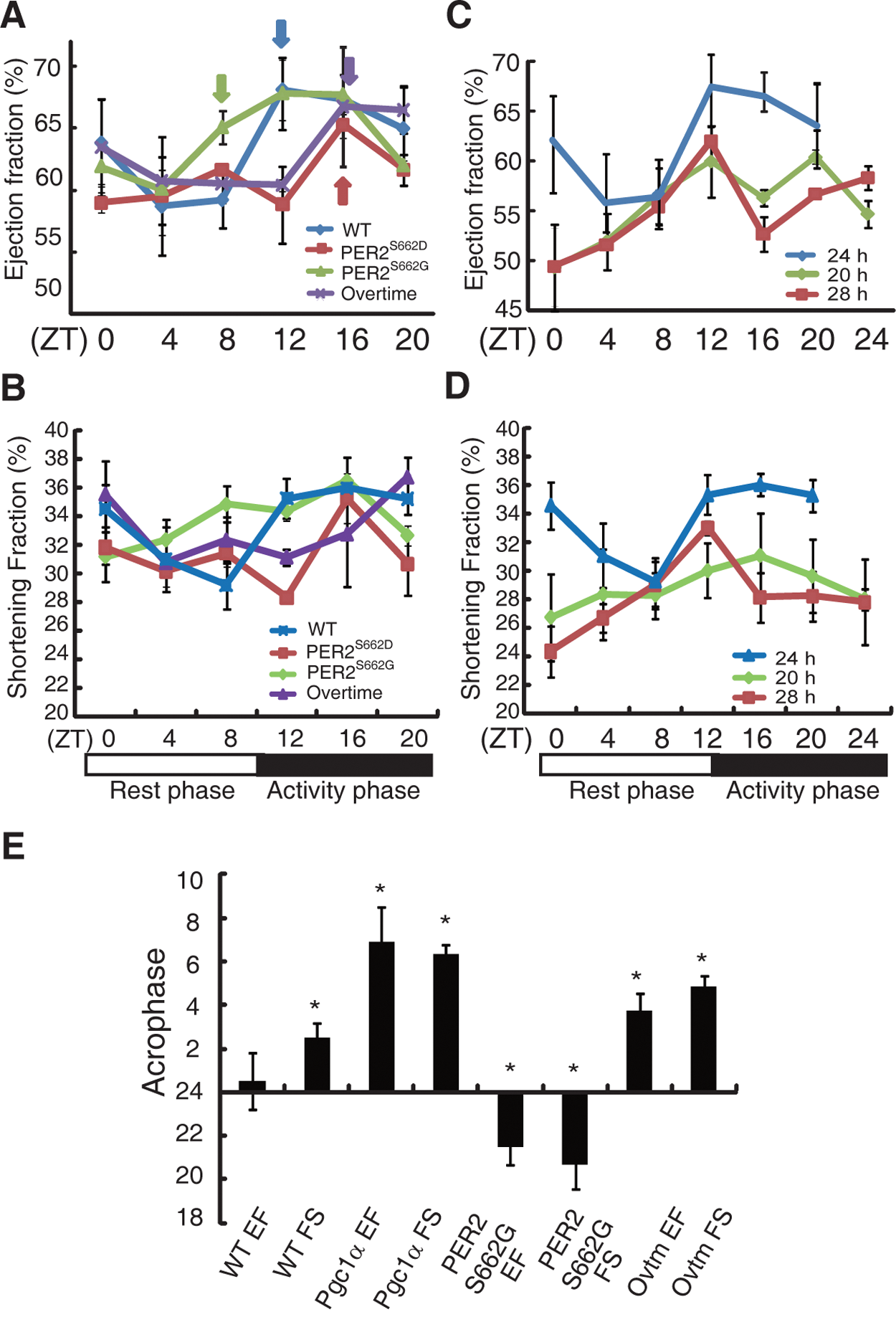
Dysfunctional circadian clock affects ejection fraction (EF) diurnal variation. Diurnal variation of EF (A) and shortening fraction (FS) (B) of wild-type (blue line), PER2S662G (green), PER2S662D (red), and ovtm (purple) mice. Diurnal variation of EF (C) and FS (D) of wild-type mice housed in 20-hour (light:dark 10:10) (green), 24-hour (light:dark 12:12) (blue), and 28-hour (light:dark 14:14) (red) light cycles under forced exercise. (E) Acrophase of EF and FS of mice after exercise. Values are shown as the mean ± SEM for four to six observations and were repeated in at least two independent experiments with different mice. Arrows represent onset of rising EF for each genotype. Asterisk represents acrophase difference for EF and FS comparing respectively between wild-type mice and indicated genotype. p < 0.01.
Echocardiographic analysis of different mouse genotypes (n = 10).
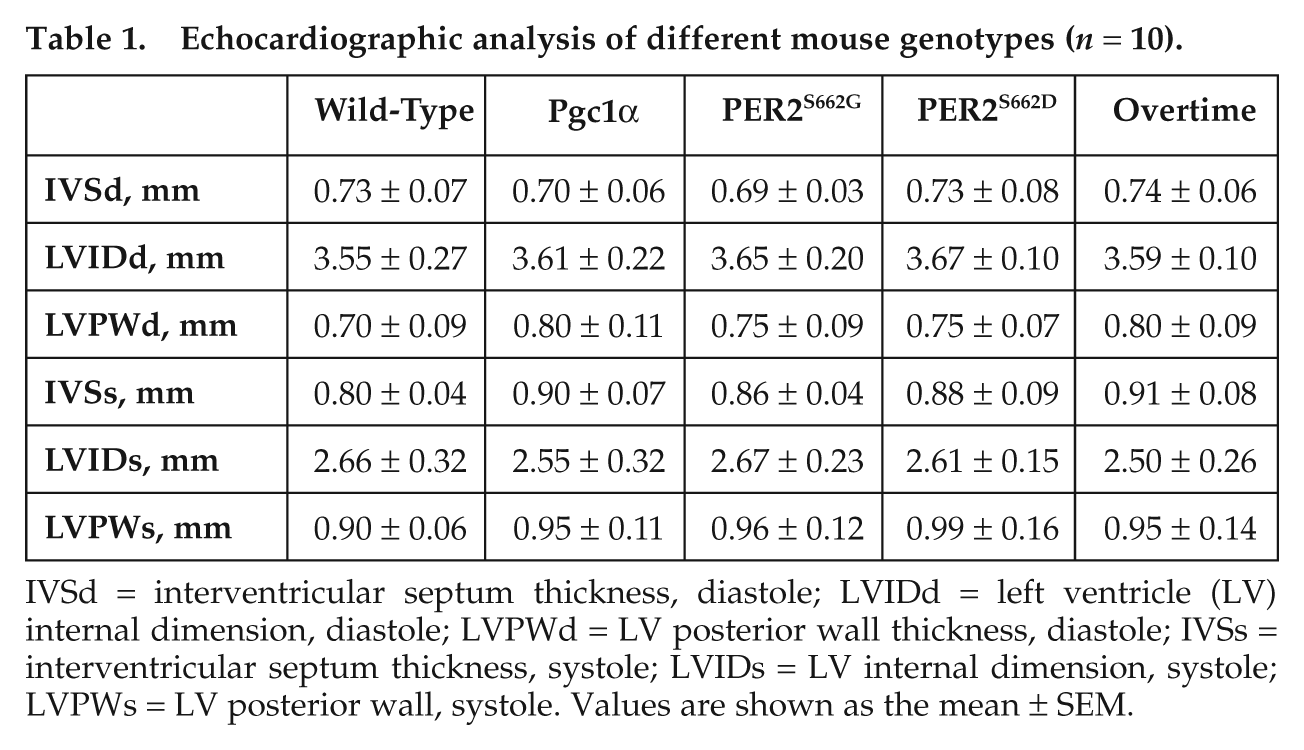
IVSd = interventricular septum thickness, diastole; LVIDd = left ventricle (LV) internal dimension, diastole; LVPWd = LV posterior wall thickness, diastole; IVSs = interventricular septum thickness, systole; LVIDs = LV internal dimension, systole; LVPWs = LV posterior wall, systole. Values are shown as the mean ± SEM.
Alterations of the LD cycle have been shown to re-entrain the circadian clocks in the suprachiasmatic nucleus (SCN) and in dependent peripheral tissues. To explore whether EF and FS are also affected in response to a non–24-hour (12:12) LD cycle, echocardiography was performed to measure the EF and FS after forced exercise at the indicated time points after entrainment on a 20-hour (10:10) or 28-hour (14:14) LD cycle for 4 weeks. The heart contractile capacity to respond to exercise was severely impaired (Fig. 2C and 2D). The MESOR was reduced to 56.08 ± 1.31 (EF) and 28.62 ± 0.95 (FS) on the 20-hour LD and to 55.97 ± 0.55 (EF) and 28.05 ± 0.24 (FS) on the 28-hour LD cycle compared with 64.33 ± 1.45 (EF) and 33.88 ± 1.05 (FS) on the 24-hour LD cycle (p < 0.001) (Suppl. Table S1), reflecting the entrainment of the circadian oscillator to different phases that suppress cardiac function.
The Circadian Clock Is Disrupted in Pgc1α Transgenic Mice
PGC1α is a key component of the circadian oscillator, which integrates the mammalian clock and energy metabolism (Liu et al., 2007). PGC1α expression is highly inducible in response to physiological cues, such as cold exposure and exercise (Finck and Kelly, 2006; Handschin and Spiegelman, 2006; Lin, 2009). PGC1α is therefore potentially a good model with which to study the links between circadian rhythms and energy metabolism in the heart after exercise. To avoid extremely high, nonphysiological expression levels of PGC1α, we selected the muscle creatine kinase (MCK) promoter instead of the alpha-myosin heavy chain (α-MHC) promoter. As expected, the transgenic mice overexpressing Pgc1α exhibited a 7.7-fold and a 2.9-fold increase in Pgc1α mRNA expression at ZT 8 in the muscle and heart, respectively, compared with wild-type littermates (Fig. 3A). PGC1α was maintained at a high level in the hearts of transgenic animals throughout the day (Suppl. Fig. S3). These mice developed normally, and echocardio-graphy and cardiac histology did not show obvious abnormalities (Fig. 3B and 3C and Table 1). We next examined the expression of circadian genes in Pgc1α transgenic mice because circadian gene expression has been shown to be altered in Pgc1α knockout mice (Liu et al., 2007). The expression of a majority of clock genes was inhibited in both heart and muscle tissues, although to varying extents (Fig. 4A and 4B). Furthermore, we monitored PER2:LUC oscillations in primary muscle cells by crossing Pgc1α transgenic mice to homozygous mPer2Luc knock-in reporter mice (Yoo et al., 2004). Luminescence was continuously measured in real time using PMT detector systems (Actimetrics, Wilmette, IL). Muscle cells in which Pgc1α was overexpressed exhibited an obvious elongation period (Tg: 24.83 ± 0.05 hours v. WT: 24.27 ± 0.04 hours, p < 0.01) (Fig. 4C). These data demonstrate that Pgc1α plays an important role in modulating circadian rhythms.
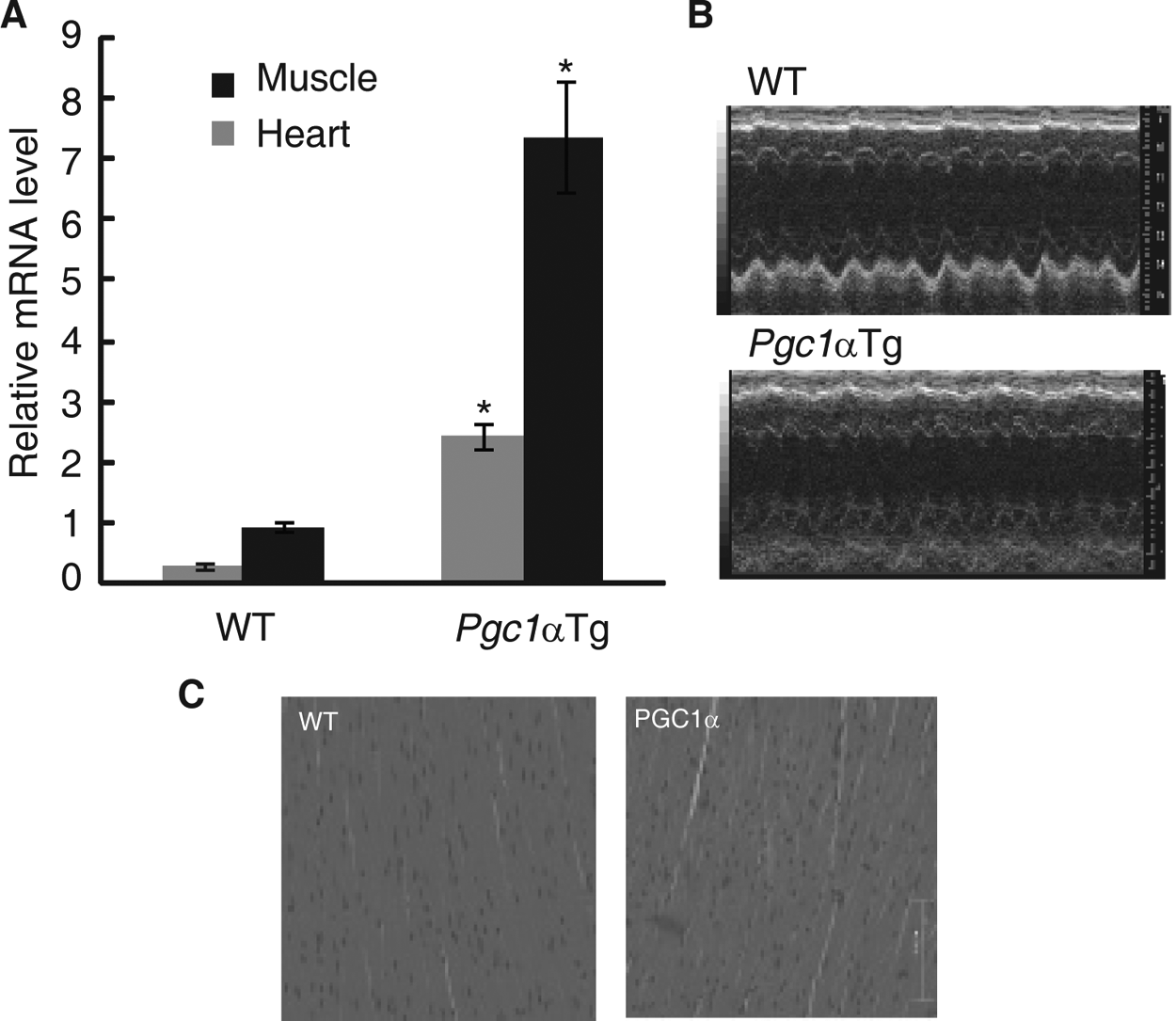
Analysis of Pgc1α transgenic mice. (A) Muscle creatine kinase (MCK) promoter induced Pgc1α overexpression. Relative Pgc1α mRNA expression level in muscle (black column) and heart (gray column) of wild-type and transgenic mice. Tissues were collected at ZT 8. Values are shown as the mean ± SEM for three independent experiments and were normalized to the corresponding Actin levels. Student t test was performed, and asterisk indicates significantly different mRNA expression levels of Pgc1α between wild-type and transgenic mice in the heart (gray bar) and muscle (black bar). p < 0.01. (B) Echocardiographic images of wild-type and Pgc1α transgenic mice. No obvious abnormalities in the hearts of Pgc1α transgenic mice were observed compared with the hearts of wild-type mice. Representative M-mode echocardiographic images of wild-type (top) and Pgc1α transgenic mice (bottom). (C) Representative examples of hematoxylin and eosin–stained left ventricular (LV) tissues from wild-type (left) and Pgc1α transgenic mice (right).
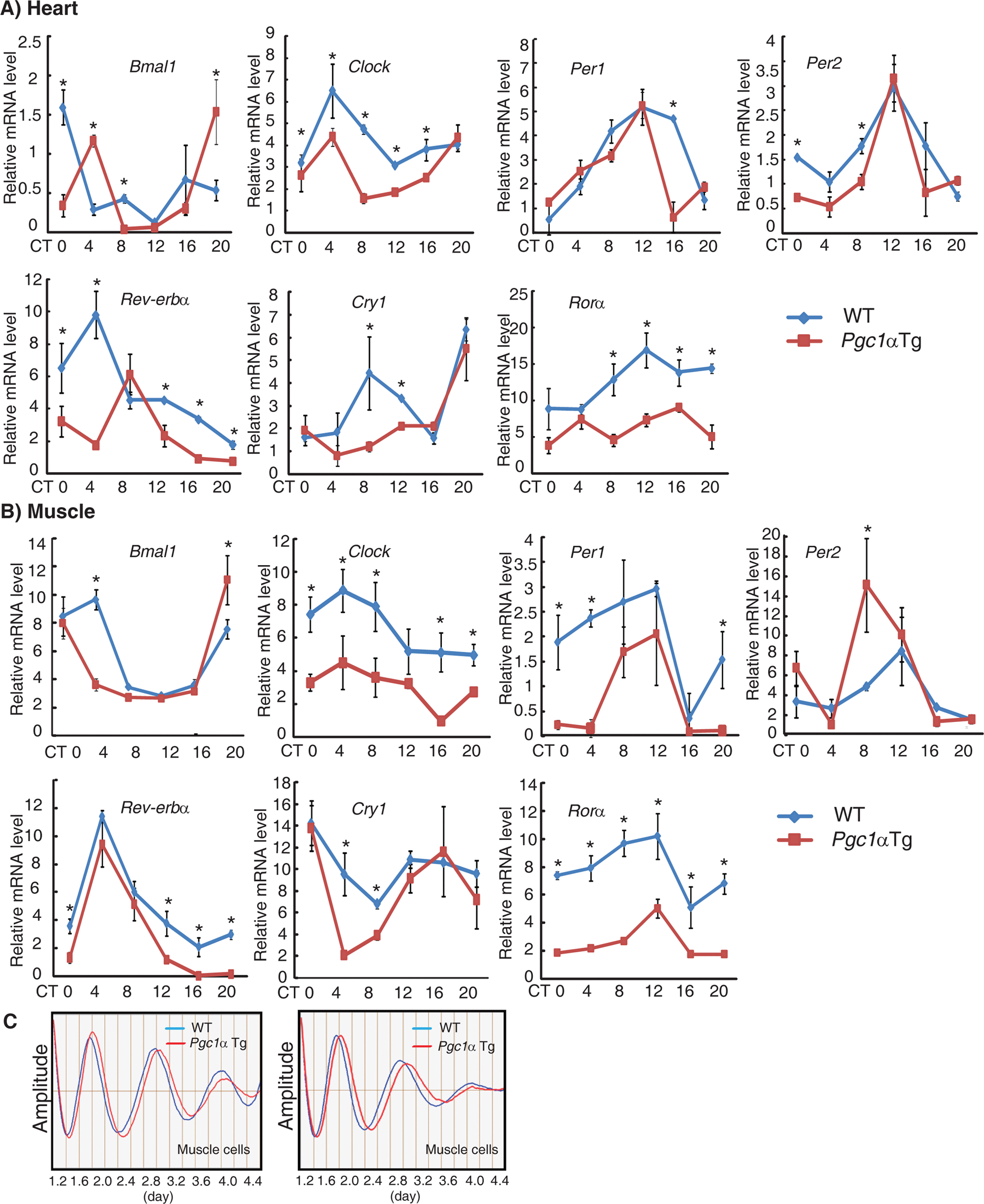
Ablation of the Pgc1α gene affects the circadian clock. Expression of clock genes was disrupted in the heart (A) and muscles (B). Representative mRNA profiles of clock genes at the indicated circadian times after the first day in constant dark. Values are shown as the mean ± SEM for three independent experiments and were normalized to the corresponding Actin levels. Scheffé multiple comparison tests assessed expression differences of clock genes between wild-type and Pgc1α transgenic mice. Asterisk indicated p < 0.05 between wild-type and Pgc1α transgenic mice at the indicative time point. (C) Period was prolonged in the primary muscle cells of Pgc1α transgenic mice. Two representative bioluminescence waveforms emitted by primary muscle cells from wild-type/mPer2Luc mice (blue) and Pgc1α transgenic/mPer2Luc mice (red).
Diurnal Variations in EF and FS Are Blunted in Pgc1α Transgenic Mice
Echocardiography was employed to observe EF and FS variations over a 24-hour period in Pgc1α transgenic mice. In sedentary Pgc1α transgenic mice, the EF and FS variations did not fit the cosinor curve (Fig. 5A and 5B). Furthermore, Pgc1α transgenic mice exhibited decreased MESOR (WT: 64.33 ± 1.45 v. Tg: 59.74 ± 1.93, p < 0.01 for EF; WT: 33.89 ± 0.61 v. Tg: 30.89 ± 0.49, p < 0.01 for FS) and phase delays (WT: 0.52 ± 1.32 v. Tg: 6.9 ± 1.59, p < 0.01 for EF; WT: 2.51 ± 0.64 v. Tg: 6.32 ± 0.46, p < 0.01 for FS) when subjected to forced exercise (running on a treadmill) (Fig. 2E, Suppl. Fig. S2, and Suppl. Table S1). Of note, heart performance was significantly compromised during the activity phase between ZT 12 and 20 in transgenic mice subjected to forced exercise (Fig. 5C and 5D).
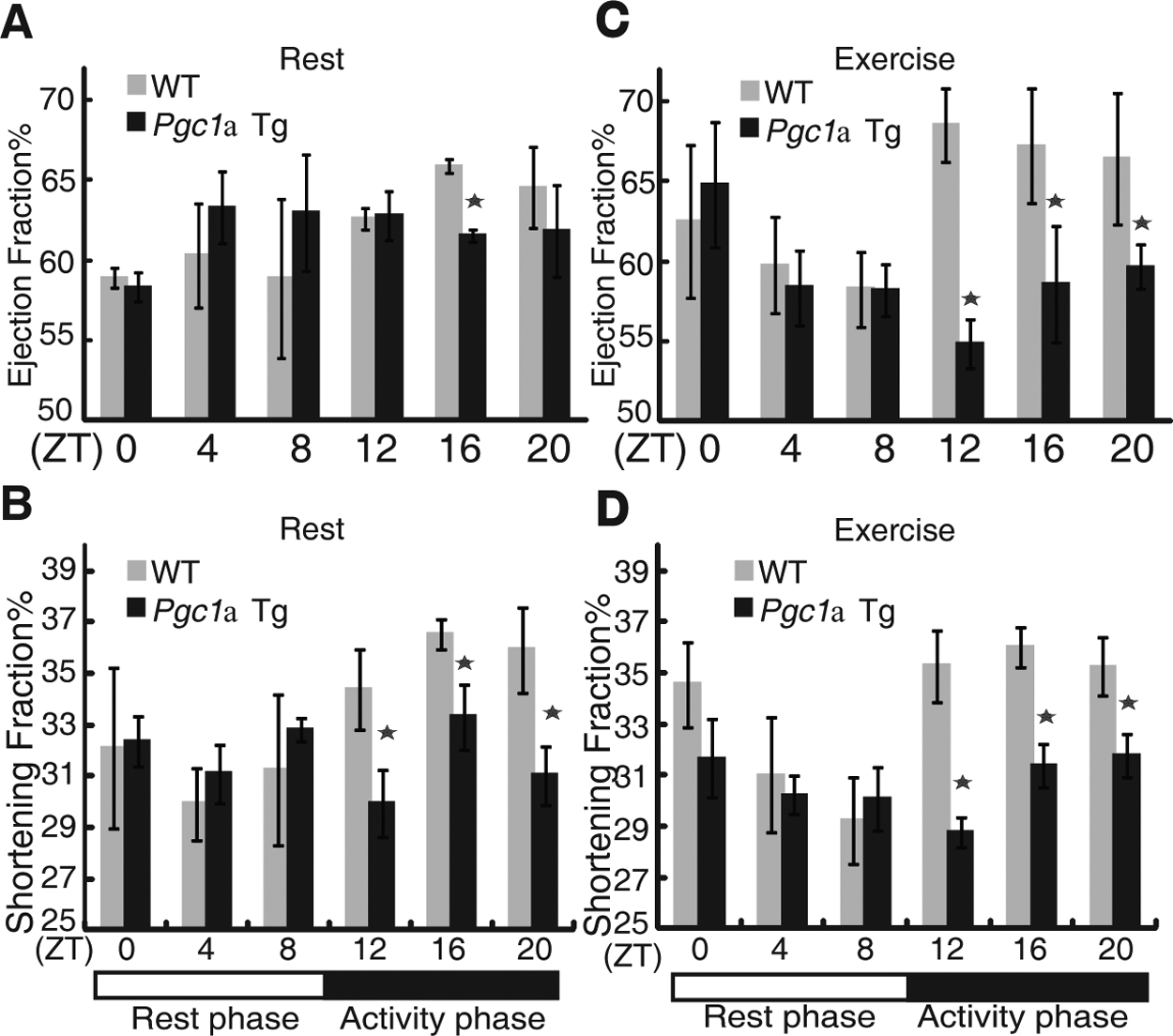
Diurnal ejection fraction (EF) and shortening fraction (FS) variation is blunted in Pgc1α transgenic mice. (A and C) EF in wild-type mice (gray column) and Pgc1α transgenic mice (black column) under sedentary (A) and enforced exercise (C) conditions. (B and D) FS in wild-type mice (gray column) and Pgc1α transgenic mice (black column) under sedentary (B) and enforced exercise (D). Scheffé multiple comparison tests assessed specific EF and FS differences between genotypes, and asterisk indicated p < 0.01 between wild-type and Pgc1α transgenic mice at the indicated time point.
Circadian Variations in EF and FS Are Modulated by Circadian Expression of Metabolic Genes
To gain a better understanding of the underlying mechanisms of the circadian oscillations of EF, we investigated the RNA expression profiles of genes involved in the metabolic pathways that are known to cycle in the heart (Durgan and Young, 2010). The expression profiles of Pparα, acetyl-coA synthetase (Acs1), and the Glut4 glucose transporter exhibited obvious circadian oscillations in wild-type mice. However, these circadian oscillations were significantly dampened in Pgc1α transgenic mice, especially in the activity phase (Fig. 6A). After forced exercise, the diurnal variations of Pparα and Acs1 increased in both wild-type mice and Pgc1α transgenic mice, but the induced levels of Pparα and Acs1 were severely attenuated in Pgc1α transgenic mice (Fig. 6A). Glut4 expression was inhibited in response to exercise (Fig. 6A). Interestingly, a higher percentage of altered expression appeared during the activity phase in response to exercise compared with that observed during the resting phase. These effects varied in parallel with the observed variations in the diurnal EF and raised the possibility that these genes directly regulate diurnal EF variations.
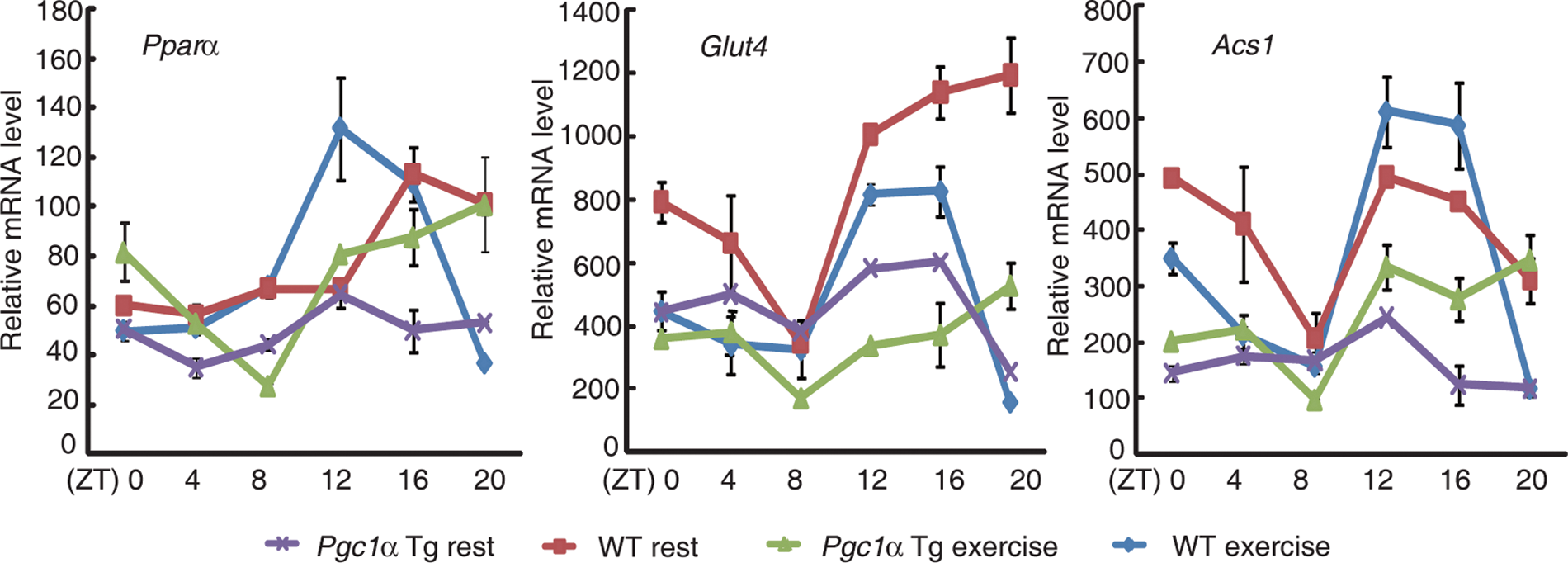
Circadian variation of ejection fraction (EF) and shortening fraction (FS) is modulated by PGC1α/PPARα. Expression of metabolic genes Pparα, Glut4, and Acs1 in the hearts of wild-type and Pgc1α transgenic mice under sedentary and enforced exercise conditions. Values are shown as the mean ± SEM for three independent experiments and were normalized to the corresponding Actin levels.
Discussion
Here, we present compelling evidence that the heart can sense the time of day and that PGC1α may function as regulating circadian rhythm of metabolic genes on EF. First, the EF showed diurnal variation that became pronounced in response to exercise. Second, disruption of the circadian clock altered the EF phase of oscillation, and entrainment to nonstandard light regimens (i.e., 20- or 28-hour LD cycles) attenuated the EF. Third, overexpression of Pgc1α impaired circadian rhythms and blocked exercise-induced increases in the EF during the activity phase, as evidenced by a reduction in metabolic genes expression.
Several laboratories have confirmed the diurnal oscillations in circadian clock gene expression in the rodent heart (Martino et al., 2004; Mohri et al., 2003; Naito et al., 2002; Young, 2009). Furthermore, it has been shown that the cardiomyocyte circadian clock influences myocardial gene expression, transcriptional responsiveness to fatty acids, triglyceride metabolism, and ischemia-reperfusion tolerance by generating cardiomyocyte-specific clock mutant mice (Durgan et al., 2006; Durgan et al., 2010). The results of these studies suggest that the cardiomyocyte circadian clock enables the heart to anticipate environmental stimuli, thus facilitating timely and appropriate responses (Young, 2009). Our observations further support the results regarding the roles of the cardiac circadian clock and the pathological consequences of circadian clock disruptions. In the genetically modified mice, the observed phase shifts in cardiac contractile function under LD cycles were similar to their “free-running” period. These results are in agreement with the parametric entrainment theory and indicate that the shorter periods observed in PER2S662G mice lead to phase advances and that the longer periods observed in Ovtm mice lead to phase delays under normal LD cycles. However, we observed a phase delay in Pgc1α transgenic mice, although luminescence data from primary cultured muscle cells from these animals displayed a period that was only one half hour longer than that of wild-type mice. These results indicate that the extent of phase shift alterations induced by disruptions of PGC1α extends beyond simply changing period length. These data provide further evidence that the circadian clock directly regulates diurnal EF and FS variations. Remarkably, the overexpression of Pgc1α disrupts normal circadian rhythms and impairs cardiac contractile function. In this case, PGC1α exerts its effects on the circadian clock and cardiac energy metabolism by serving as a transcriptional coactivator.
The link between the external environment and cellular metabolic responses is mediated by the activation of nuclear receptors. PPARα is a member of the PPAR family of nuclear receptor transcription factors and exhibits a remarkable circadian expression pattern in liver, adipose, and heart tissues. PPARα has also been linked to the regulation of the circadian gene Bmal1 through the direct binding of PPARα to a typical PPRE element in the Bmal1 promoter (Canaple et al., 2006). Thus, PPARα may affect phase shifts of the circadian genes Bmal1, Per3, and Rev-erbα (Goh et al., 2007). Recent evidence has emerged that PPARα plays an important role in connecting circadian biology to metabolism. We found that both Pparα and Pparα target genes were significantly downregulated in the activity phase when Pgc1α was overexpressed in cardiac tissue. Besides, glucose metabolic gene glut4 was also decreased. The time-dependent decrease expression of metabolic genes in parallel with the observed variations in the diurnal EF indicated that these genes may directly regulate diurnal EF and FS variations.
Footnotes
Acknowledgements
The authors extend appreciation to Bradford Lowell for Pgc1α transgenic mice, J.S. Takahashi for Overtime mice, and Fu and Ptácˆek laboratories for human PER2 transgenic mice. This work was funded by the National Science Foundation of China (30725011 and 30671039) and the Ministry of Science and Technology (2006CB943503 and 2010CB945102).
The author(s) have no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.