
Abstract
Chert and chert-phosphorite deposits of Lower Tal Formation have yielded new records of microfossil assemblage containing biomineralised articulated sponge Paradiagoniella sp., tubular microfossil Megathrix longus and other organic-walled microfossils from the Lesser Himalaya. Their occurrence indicates biostratigraphic correlation of Lower Tal Formation with Yanjiahe and Chiungchussu formations of Yangtze Platform in South China, bracketing the age between ~514 and 529 Ma. Their presence also suggests shallow marine oxic depositional environment. The geochemical characteristics of Lower Tal chert are compared with Cambrian chert from Yanjiahe and Yutrus formations (China). Their geochemical analyses indicate deposition in suboxic to oxic environments with clastic input close to the continental shelf. The present study suggests that Northern India was located near South China during the early Cambrian when suboxic to oxic conditions prevailed within the stratified Ocean. Here, abundant oxygen and silica-secreting organisms residing in the shallow marine regions assisted in the radical evolutionary change that occurred as the Cambrian Explosion of life.
Keywords
INTRODUCTION
An integrated palaeobiological and geochemical approach is required to understand the Cambrian Explosion, which is considered a fundamental step in the evolutionary history of life. Chemically stable fossiliferous cherts are potential rocks that can unravel the ancient oceans’ conditions that led to a Cambrian explosion of life (Fan et al., 2018; Zhang et al., 2020). In India, early Cambrian chert from the Tal Formation yielded microfossils (Mazumdar & Banerjee, 1998; Tiwari, 1999). After this, preliminary geochemical investigations were initiated (Mazumdar et al., 1999). Detailed palaeobiological and geochemical studies are required in purview of the aforesaid statements. The present study has been done in Cambrian cherts from the Lower Tal Formation exposed in new sections within Mussoorie (Masrana section) and Garhwal (Kaudiyala section) synclines of the Krol belt in the Lesser Himalaya. The study showcases new microfossil findings and palaeodepositional conditions of the Tal Formation. The biostratigraphic, palaeoenvironmental, and evolutionary significance of new palaeobiological records and their implications for Cambrian Explosion are discussed. The geochemical evidence of paleo-redox conditions, detrital/hydrothermal input, and tectonic setting from these fossiliferous deposits is studied. The geochemical and palaeobiological characteristics of Tal chert are compared with Cambrian chert from the Yangtze platform in South China (Yanjiahe and Chiungchussu formations) and the Akesu-Wushi area in Tarim Basin in Northwest China (Yutrus formation).
GEOLOGICAL SETTING
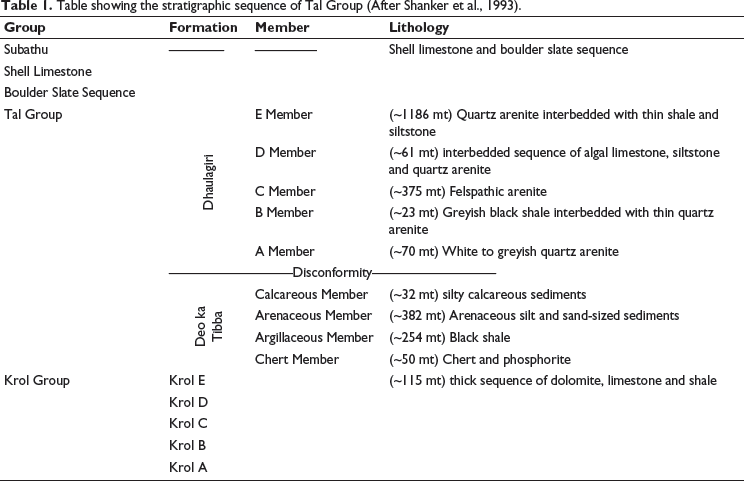
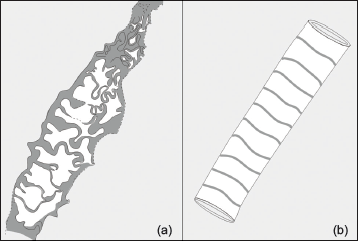
In India, Neoproterozoic-Cambrian sedimentary deposits of Baliana-Krol-Tal groups (Jain et al., 2020; Jiang et al., 2002; Shanker et al., 1993) are well exposed within various synclines of Krol Belt in Lesser Himalayas (Figure 1a). The siliciclastic early Cambrian Tal Group, which is rich in chert deposits, overlies Neoproterozoic carbonate rocks of the Krol Group (Jiang et al., 2002). Contact between the Krol and Tal groups is marked as the Precambrian-Cambrian boundary (Bhargava et al., 1998), which is considered a marker of the Cambrian Explosion. Tal Group is well exposed in Mussoorie and Garhwal synclines (Figure 1b and 1c). Tal Group is divided into Lower Tal/ Deo-ka-Tibba and Upper Tal/Dhaulagiri formations (Table 1). Lower Tal/ Deo-ka-Tibba Formation is subdivided into (a) Chert, (b) Argillaceous, (c) Arenaceous, and (d) Calcareous members (Shanker et al., 1993). The present investigation has been done on the Chert Member of Lower Tal Formation, where chert is exposed both in bedded and nodular forms associated with phosphorite and calcareous shale (Shanker et al., 1993).
(a) Blaini-Krol-Tal groups exposed in the Krol Belt of Lesser Himalayas (Modified after Shanker et al., 1993); (b) Geological map of Mussoorie syncline (redrawn after Shanker, 1975); (c) Geological map of the Garhwal syncline (redrawn after Kumar et al., 1983).
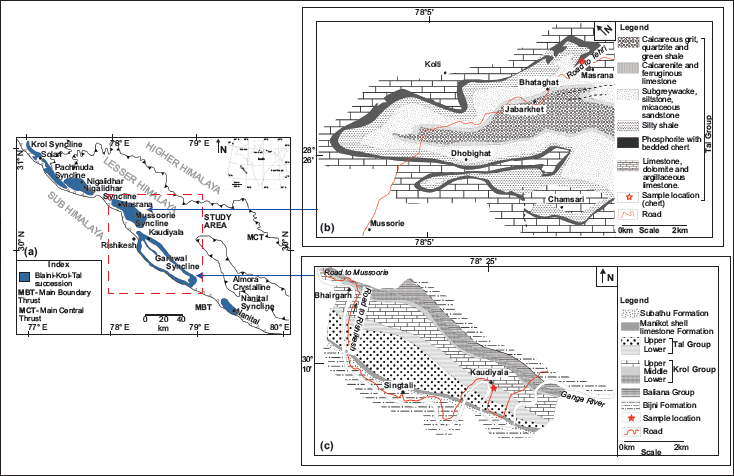
Table showing the stratigraphic sequence of Tal Group (After Shanker et al., 1993).
MATERIALS AND METHODS
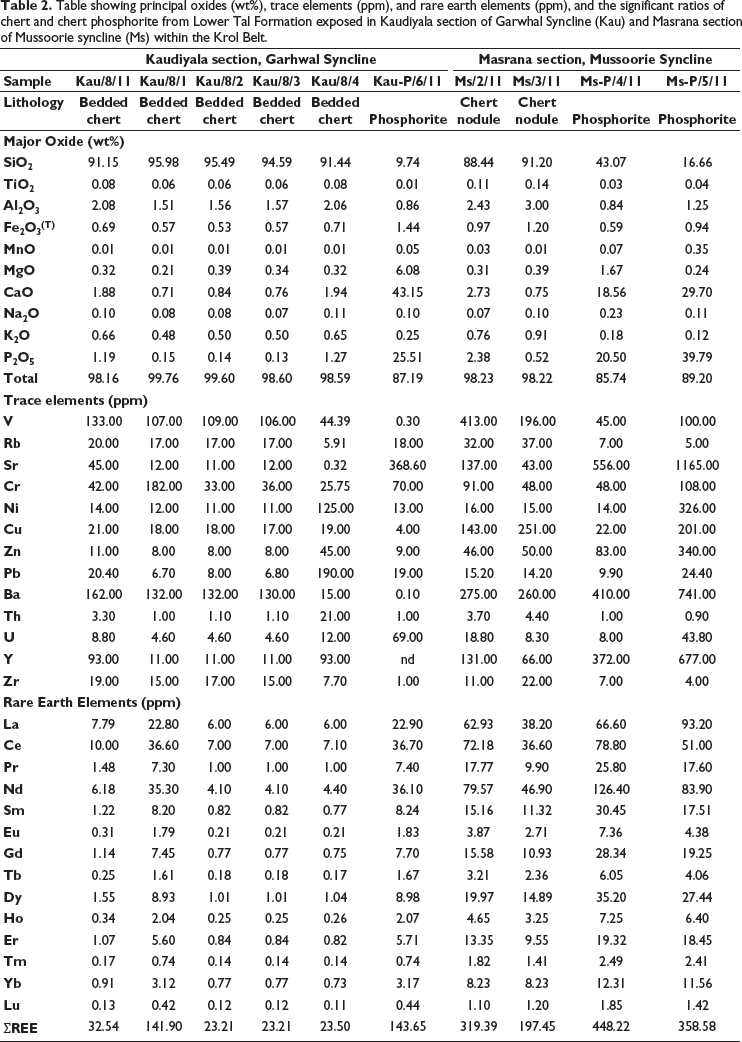
Ten chert and phosphorite samples were taken from Chert Member of the Lower Tal Formation (lithologs with sample locations shown in Figure 2a) at Masrana [30⁰27′3.8′′ N: 78⁰08′2′′ E; 02 chert and 02 phosphorite nodules (ms)] and Kaudiyala [30⁰04′28.59′′ N: 78⁰30′4.04′′ E; 05 bedded chert and 01 phosphorite (kau)] sections within Mussoorie and Garhwal synclines respectively (Figure 2b and 2c). These samples were cut so that one half was used for palaeobiological and the other for geochemical investigations. All analyses were done at the Wadia Institute of Himalayan Geology, Dehradun. Petrographic thin sections were scanned under Olympus BX61 for microfossils photographed using the attached camera DP71 and submitted to the WIHG repository (listed in section 7). The chemical composition of recovered microfossils was identified to substantiate their biological origin using Raman spectroscopic analyses through LabRAM HR-Horiba Jovin Yvon-High Resolution Laser Raman Micro Probe. Results of Raman shift are within ± 1–2 cm−1 w.r.t. standard values. Major oxides and rare earth elements (REEs) were analysed with XRF Spectrophotometer BRUKER S8 TIGER using the procedure of Saini et al. (2002). PerkinElmer SCIEX with an ICPM Spectrometer ELAN DRC-e was used for trace element analysis using the analytical protocol of Khanna et al. (2009). International rock standards SBR/CK and J Ch-1 were reference materials. Instruments were calibrated with WIHG in-house standards BRS and JG-2 for shale and international standard J Ch-1 for chert samples. Replicate analyses of samples and reference standards were done to assess analytical reproducibility and accuracy of data. Accuracy (%RSD) for major and minor oxides is <5%, and trace elements is <12%. Analytical error is within ±5%. Average precision is better than 1.5% as reported by Islam et al. (2011).
(a) Generalised litholog of Krol belt along with the litholog (1) of fossiliferous Masrana section in Mussoorie syncline and (2) Kaudiyala section in Garhwal Syncline; (b) Photograph of the outcrop showing Masrana section with the inset showing the exposed chert nodules; (c) Photograph of the outcrop showing Kaudiyala section with the inset showing the exposure of chert and chert phosphorite.

RESULTS AND DISCUSSION
Petrography and mineralogy
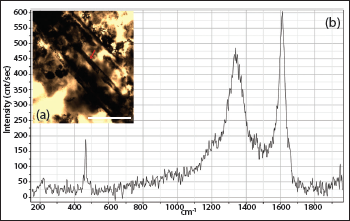
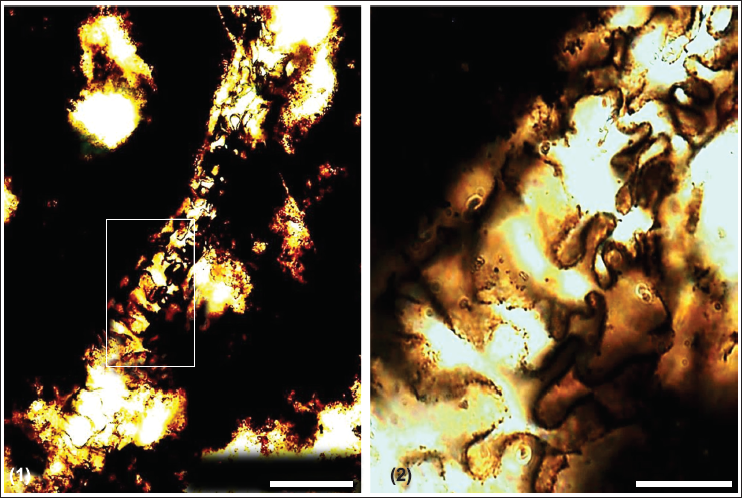
In thin section, chert samples comprised cryptocrystalline quartz, replaced calcite remnants with minor muscovite and clay as impurities (Figure 3a). An exceptional presence of fossils and their permineralisation is observed. Raman spectra of the axial canal and sponge spicule attest their biogenicity, showing major peaks and overall trend similar to disordered organic carbon and phosphate (Figure 4a and 4b). Phosphorite samples comprise greyish to dark brown phosphatic material within peloidal structures (Figure 3a–3c). Layered apatite rims are observed (Figure 3d). Abundant sponge spicules and acritarchs, with some deformed acritarchs, are seen irregularly dispersed. The shale layers associated with chert consist of very fine quartz grains, minor sericite, muscovite flakes and dispersed deformed acritarchs in thin section.
Photomicrographs of chert and phosphorite from the Lower Tal Formation showing (a) cryptocrystalline quartz and replaced calcite remnants with corroded boundaries; (b, c) pelloidal texture; (d) Growth layers in apatite rim; Scale for a = 100 µm; b = 100 µm; c = 20 µm; d = 50 µm.

Palaeobiological investigations and interpretation
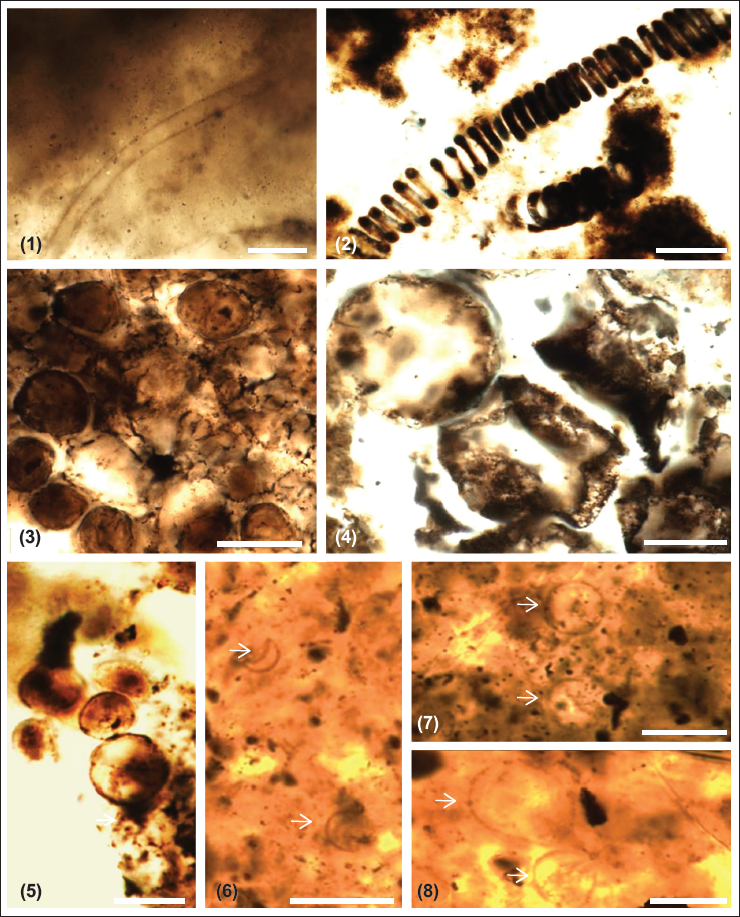
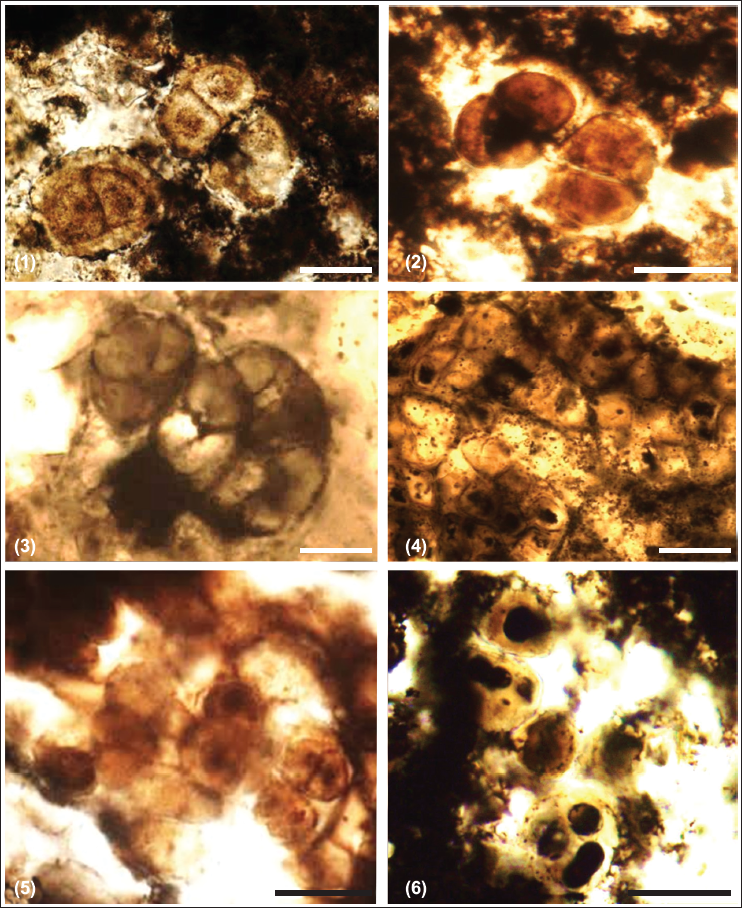
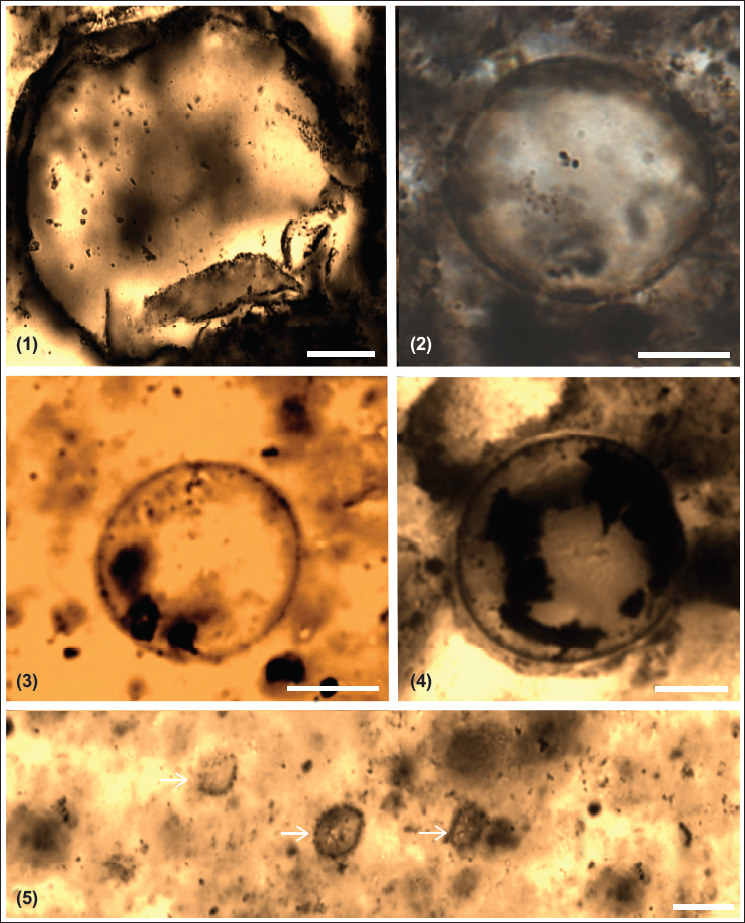
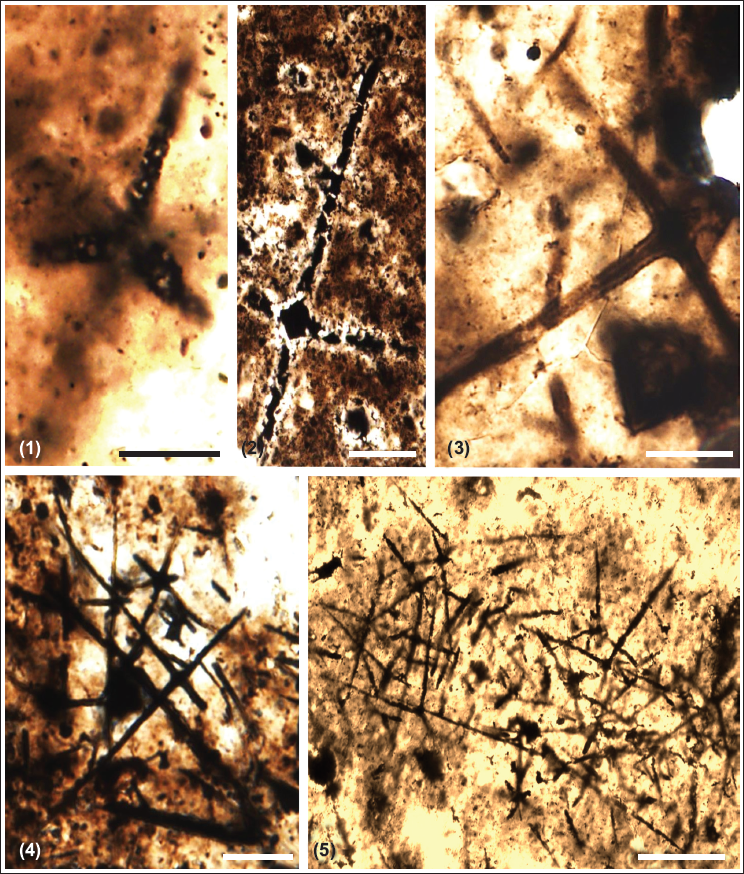
Chert and phosphorite have yielded new microfossil assemblage with organic walled non-septate filamentous cyanobacteria: Siphonophycus robustum, Obruchevella parva and coccoidal cyanobacteria: Paleopleurocapsa wopfneri, P. reniforma, Sphaerophycus medium, Glodeodiniopsis lamellosa, Archaeophycus yunannensis, Myxococcoides grandis, M. minor, Hurnoniospora microreticulata, H. psilata, Eontophysalis dismallakesensis; sphaeromorphic acritarchs: Leiosphaeridia minutissima and L. jacutica; acanthomorphic acritarchs: Asteridium tornatum; tubular microfossil Megathrix longus and biomineralised microfossil including sponge Paradiagoniella sp. along with various sponge spicules belonging to class Hexactinellida from chert and chert-phosphorite samples from Lower Tal Formation (Plates 1–5 and Figure 5). Systematic description of recovered microfossils and repository details is given in the supplementary data.
Biostratigraphic correlation of Lesser Himalaya, India and Yangtze Platform, South China, showing equivalence of Lower Tal, Yanjiahe and Chiungchussu formations with age range bracketed between ~514 and 529 Ma.

Organic-walled microfossils, evaluation of age and depositional environment
The presence of small acanthomorphic acritarch (SAA; <25 mm) Asteridium is an indicator of early to mid Meishucunian age (equivalent to Nemakit-Daldynian and early Tommotian) (Joshi et al., 2022; Tiwari, 1999). Asteridium is used to form Asteridium-Heliosphaeridium-Comasphaeridium (AHC) assemblage zone in basal Cambrian deposits of China (Moczydlowska, 1991; Yao et al., 2005). Presence of Obruchevella indicates lineage across Ediacaran-Cambrian boundary (Ahluwalia & Bhargava, 1987; Schopf, 1992; Sharma & Shukla, 2012; Tiwari, 1996). It is considered a relic of oscillatoriacean cyanobacteria similar to the modern cyanobacterium genus Spirulina (Knoll, 1992b; Luchinina, 1975; Yankauskas, 1989), which grows in saline and alkaline environments (Shih et al., 2013) and is obtained at pH 6.5–8.0 (Bano & Siddiqui, 2004). Also, the pH of modern Oscillatoria mats ranges between 7.4 and 7.9 (Revsbech et al., 1983). Another important form is Paleopleurocapsa, a fossil analogue of the modern unicellular nitrogen-fixing cyanobacterium genus Pleurocapsa (Knoll et al., 1975). It is found in alkaline water and hot springs (53°C; pH 8.5) within a narrow range of salinities (Shih et al., 2013). Spirulina and Pleurocapsa survive in pH≥ 8.5 (Klanchui et al., 2017). This indicates that the pH of early Cambrian Ocean water where fossiliferous Tal cherts deposited was 6.5 ≥ 8.5.
Tubular and biomineralised microfossils, depositional environment, and biostratigraphic correlation
In India, the Lower Tal Formation of the Lesser Himalaya has chert and phosphorite within interbedded black shale, indicating a shallow subtidal depositional environment. These sedimentological characters are similar to those of the Cambrian Yutrus and Yanjiahe formations of Tarim basin in NW and the Yangtze platform in southern China, respectively (Dong et al., 2009; Yu et al., 2009). Large tubular microfossil Megathrix, articulated sponge Paradiagoniella, and various hexactinellid sponge spicules in Lower Tal are significant biostratigraphically. Megathrix is reported from the basal Cambrian Yanjiahe Formation in the Three Gorges area of the Yangtze Platform (Meishucunian Stage/Stage 2; ~529 Ma) (Dong et al., 2009; Okada et al., 2014). Paradiagoniella is reported from China’s early Cambrian Chengjiang biota of Chiungchussu Formation (Nangaoan/Stage 3; ~521 Ma) (Chen et al., 2015). Hence, the age of the Lower Tal Formation is bracketed between ~521 and ~529 Ma (Figure 6). Megathrix is considered to be Oscillatoriaceae Cyanophyta (Shang et al., 2016; Yin, 1987), but Yao et al. (2005) suggested it to be the stem group of cnidarians, indicating the evolution of metazoans during that period. The sponge genus Paradiagoniella of family Protospongiidae is a paraphyletic group that includes members of the total group of Hexactinellids (Botting & Butterfield, 2005). The presence of sponges and ample spicules suggests the presence of plenty of oxygen.
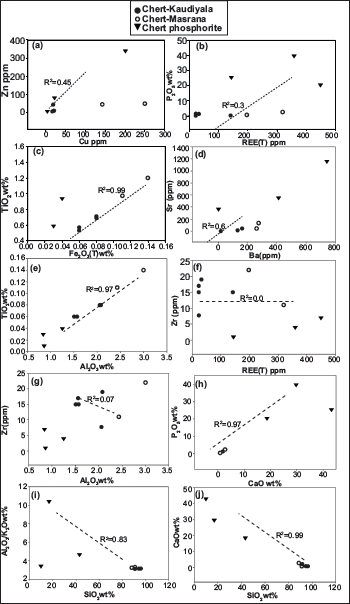
Bivariate diagrams for chert and phosphorite from Tal Formation (a) Cu vs Zn; (b) REE(T) vs P2O5 (c) Fe2O3 (T) vs TiO2; (d) Ba vs Sr; (e) Al2O3 vs TiO2; (f) REE (T) vs Zr (g) Al2O3 vs Zr; (h) CaO vs P2O5; (i) SiO2 vs Al2O3/K2O; (j) SiO2 vs CaO.
Geochemical investigations and interpretations
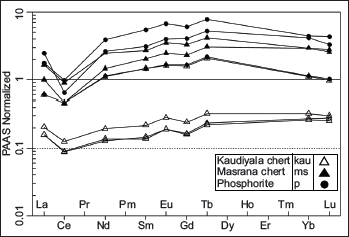
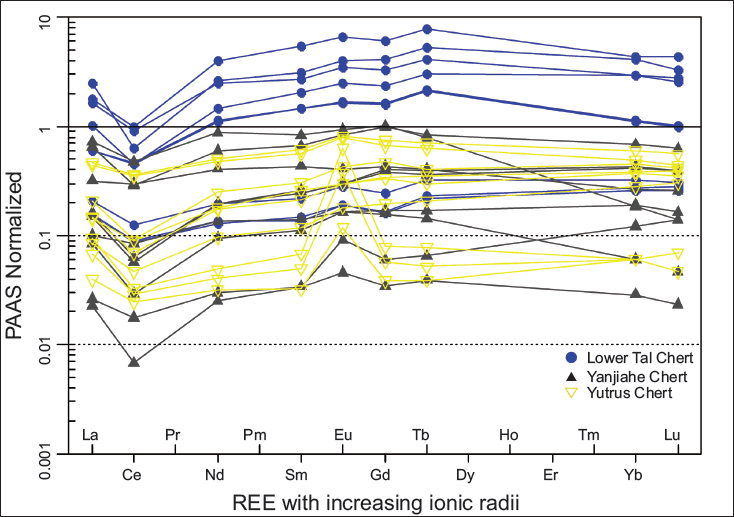
Major oxides, REEs, and trace elements for chert and phosphorite samples of the Lower Tal formation taken from new sections at Masrana and Kaudiyala are listed in Table 2. REEs have been normalised against post-Archean Australian shale (PAAS) (Figure 7). PAAS normalised REE patterns are similar to marine water (Elderfield & Greaves,1982). For chert samples, positive Eu and negative Ce anomalies are displayed for both Kaudiyala (Eu/Eu* = 1.14; Ce/Ce* = 0.65) and Masrana (Eu/Eu* = 1.15; Ce/Ce* = 0.41) sections. Depletion of LREE and MREE relative to HREE is observed. Depleted LILE (in ppm) is present with average values of Th (4.78), U (10.24), Rb (24.94) with respect to PAAS and average values (ppm) of Ba = 190.85; Sr = 53.03 and Zr = 15.62. Average values (in ppm) of transition metals Ni (25.02) and Cr (66.63) have depleted values in comparison to PAAS. For phosphorite samples, average values (ppm) of Th = 0.98, U = 47.45, Rb = 12, Ba = 287.8, Sr = 614.55, Ni = 91.5, Cr = 74, and Zr = 3.25 (with Ms-P/5/11 showing Ba = 741 and Sr = 1165). Positive correlation is observed (Figure 8) between Sr and Ba (r2 = 0.6); Zr and Al2O3 (r2 = 0.7); Zn and Cu (r2 = 0.45).
Post-Archean Australian shale (PAAS) normalised REE patterns of chert and chert phosphorite from Masrana and Kaudiyala sections of Lower Tal Formation.
(a) Eu/Eu* vs Ce/Ce* and (b) REE(T) vs Y/Ho plot for Lower Tal and Yanjiahe chert. The curve represents a mixing trend between the hydrogenous (modern water) and terrigenous (lithogenous) components (McLennan, 2001); percentages provided as the amount of hydrogenous component (Chen et al., 2015; Zhang et al., 2016).
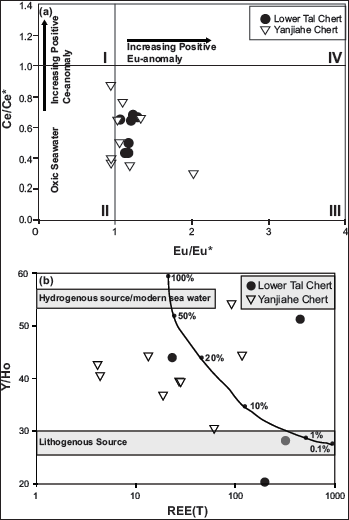
Table showing principal oxides (wt%), trace elements (ppm), and rare earth elements (ppm), and the significant ratios of chert and chert phosphorite from Lower Tal Formation exposed in Kaudiyala section of Garwhal Syncline (Kau) and Masrana section of Mussoorie syncline (Ms) within the Krol Belt.
Palaeoredox conditions
The ratios of redox-sensitive elements like V/Cr indicate oxic (<2), dysoxic (2–4.25) or suboxic to anoxic (>4.25); V/(V+Ni) indicate oxic (<0.46), dysoxic (0.46–0.6) or anoxic (>0.6); Th/U indicate oxic (>2) and anoxic (0–2) conditions of deposition (Fu et al., 2014; Jones & Manning, 1994; Rimmer et al., 2004; Wignall & Twitchett, 1996). For Lower Tal samples, average ratios of V/Cr (0.79), V/(V+Ni) (0.78), and Th/U (0.25) indicate suboxic to oxic depositional conditions. Ce3+ oxidises to insoluble Ce4+, resulting in its depletion (negative anomaly) in seawater and relative enrichment (positive anomaly) in associated deposits under oxic conditions (DeBaar, 1991; Sholkovitz et al., 1989). Mazumdar et al. (1999) linked negative Ce anomaly to rapid Ce removal from phosphate-rich water upwelling into an oxic compartment in the shallow marine water column. Negative Ce-anomaly [Masrana (Ce/Ce* = 0.41) and Kaudiyala (Ce/Ce* = 0.65)] (Figure 7) for Tal samples indicates oxic conditions. Positive Eu-anomaly (Masrana = 1.15; Kaudiyala = 1.14) indicates a low Eh, alkaline, diagenetic depositional environment with influence of hydrothermal fluids on dissolved REE load of seawater (Balaram et al., 2015; Bau & Alexander, 2009; Mazumdar et al., 1999). Chondrite normalised value of Eu shows a negative anomaly (0.75) for both Masrana and Kaudiyala samples, suggesting selective Eu depletion due to oxidation/or weathering (Chen et al., 2015). The Eu/Eu* vs Ce/Ce* plot (Figure 9a) also shows deposition in the oxic regime. Again, higher Y/Ho values (81.77) in these samples suggest an oxic shallow marine environment (Nozaki et al., 1997). Y/Ho vs REE(T) plot (Figure 9b) indicates a combination of hydrogenic and terrigenous sources.
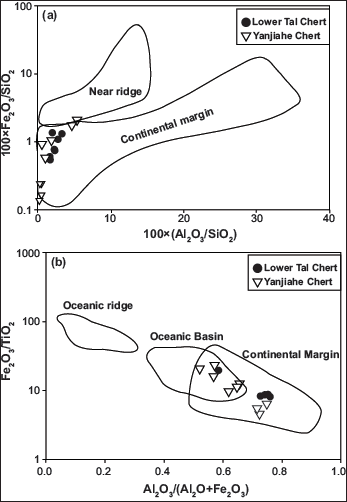
Discrimination diagrams (a) {100(Al2O3/SiO2) vs 100(Fe2O3/SiO2); (b) Al2O3 (Al2O3+Fe2O3) vs Fe2O3/TiO2]} showing the tectonic regions of chert deposition based on the major oxides. Various fields are after Murray (1994).
Detrital input
For Tal samples, values of major oxide ratios SiO2/Al2O3 (30.40–63.56), K2O/Al2O3 (0.30–0.32), Al2O3/TiO2 (21.43–26.17) and REE(T) (23.21–319.39 ppm) indicate detrital input (Garcia et al., 1994; Ling et al., 2013; McLennan, 2001; Sugitani et al., 1996). Also, these values are quite comparable to PAAS (23.2) and hemipelagic sediments (21–23) (Nance & Taylor, 1976). Sympathetic relation of Al2O3/K2O ratio to SiO2wt% (r2 = 0.83) again indicates presence of detrital component (Figure 8i). A positive correlation between P2O5 and CaO (r2 = 0.97) and REE(T) (r2 = 0.3) indicates the presence of carbonate fluorapatite (Figure 8h). CaO show positive correlation (r2 = 0.99) with SiO2 (Figure 8j). SiO2/Al2O3 value (Avg = 32.92) is higher than continental crust (~3.6) indicating SiO2 supply from multiple sources (terrestrial, biological and hydrothermal) (Jia et al., 2018).
Tectonic setting
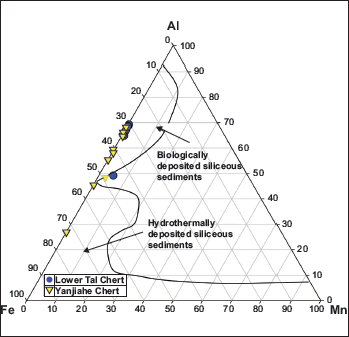
Average value of MnO/TiO2 = 0.12 for Lower Tal cherts indicates their deposition in a continental shelf and margin setting (>0.5 = open ocean, <0.5 = continental slope and margin; Adachi et al.,1986). Also, values of Al2O3/(Al2O3+Fe2O3) ratio range between 0.71 and 0.75 (Avg = 0.73), indicating closeness to continental margin. The majority of samples fall in the continental margin field within discrimination plots [100(Al2O3/SiO2) vs 100(Fe2O3/SiO2) and Al2O3 (Al2O3+Fe2O3) vs Fe2O3/TiO2] of Murray (1994) (Figure 10). Al/(Fe+Mn+Al) ratios change from 0.01 for hydrothermal origin to 0.60 for biological sedimentation (Adachi et al., 1986). Al/(Al+Fe+Mn) values of Lower Tal chert (Avg = 0.67) and phosphorite (Avg = 0.47) suggest that the majority of samples fall in the biogenic field. The same is displayed in the Fe-Al-Mn ternary plot (Figure 11).
Fe-Al-Mn ternary plot for cherts from Lower Tal and Yanjiahe formations. Various fields are after Adachi et al. (1986).
The post-Archean Average Shale (PAAS) normalised REE pattern for Lower Tal and Yanjiahe chert shows a normal seawater pattern with decreasing HREE and negative Ce anomaly.
COMPARISON WITH CAMBRIAN YURTUS (NW CHINA) AND YANJIAHE (SOUTH CHINA) FORMATIONS
Geochemical characteristics of samples from Lower Tal chert [Kaudiyala (Kau) = 05 and Masrana (Ms) = 02)] were compared with Cambrian Yutrus [(Xiaoerbulake (XCM) = 05; Sugantebulake (SCM) = 04)] and Yanjiahe [(Gunshiao (gy) = 03, Luojiacum (ly) = 03, Qiaojiaping (qy) = 03)] formations. The correlation of their significant ratios is listed in Table 3. MnO/TiO2 ratios for Lower Tal (0.10–0.15) and Yanjiahae (0.02–0.39) are <0.50, which indicates a continental margin setting (Shimizu et al., 2001). Al2O3/(Al2O3+Fe2O3) ratios for Lower Tal (0.71–0.74) and Yanjiahae (0.56–0.69) are >0.50; Fe2O3/TiO2 for Lower Tal (8.6–9) and Yanjiahae (7.1–21.9) are <50. These values indicate a continental margin environment with the influence of terrigenous input (Liao et al., 2019). These ratios signify that Yanjiahe’s Gunshiao and Lower Tal cherts are similar. The average Si/(Si+Al+Fe+Ca) ratio for Lower Tal (0.92–0.95) cherts suggests biogenic origin of silica and diagenetic input (Murray, 1994). Average Al/(Al+Fe+Mn) ratio values in cherts for Lower Tal and Yanjiahae formations range from 0.65 to 0.68 and 0.50 to 0.63, respectively. These values are close to average shale composition (0.61), reflecting the contribution of continental and non-hydrothermal sediments (Baltuck, 1982). In the Al-Fe-Mn diagram of Adachi et al. (1986), all samples fall in the non-hydrothermal zone (Figure 11). In the Eu/Eu* vs Ce/Ce* plot, Tal cherts fall in the oxic regime (Figure 9). Tectonic discrimination plots infer continental shelf and marginal sea with shallow water conditions for all formations (Figure 11).
Table showing correlation of significant major oxide, trace element and REE ratios for chert from Lower Tal (present study), Yanjiahe (Chang et al., 2009) and Yutrus (Yu et al., 2009) Formations.
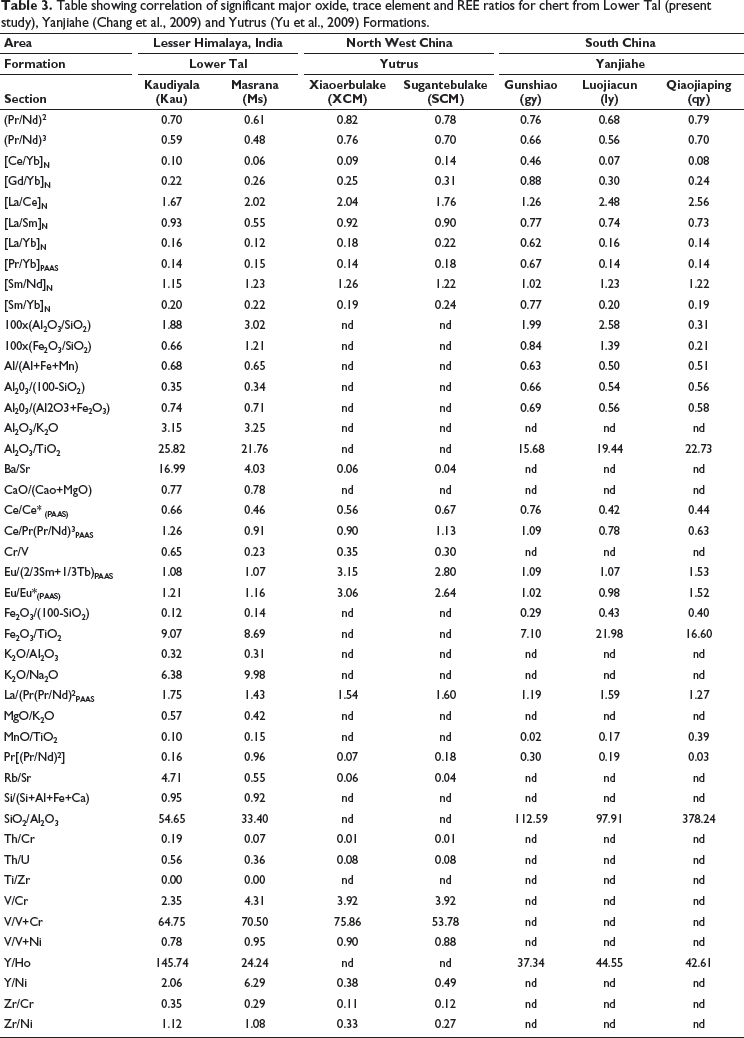
PAAS normalised REE pattern for Tal, Yutrus, and Yanjiahe formations shows a typical seawater pattern with decreasing HREE (Figure 12). Values of Eu/Eu* ratios for Lower Tal (1.07–1.29), Yutrus (1.01–3.37), Yanjiahae (0.95–2.02) show positive anomalies indicating hydrothermal input. Value of Ce/Ce* ratios for Lower Tal (0.29–0.68), Yutrus (XCM = 0.46–0.69; SCM = 0.49–0.83), Yanjiahae (gy = 0.65–0.87; ly = 0.37–0.50; qy = 0.30–0.66) indicate negative Ce anomaly, highlighting preferential removal of Ce to host rocks during diagenetic loss and oxidised seawater (Bau & Dulski, 1996; Alibo & Nozaki, 1999; Chen et al., 2015). Positive Eu anomaly and higher values of Zn and Cu indicate hydrothermal influence on REE budget in cherts of Yanjiahe (Qiaojiaping section) and some part of the Yutrus formations. The geochemical dataset suggests suboxic-oxic conditions during the Cambrian. Conceivably, a redox-stratified ocean with oxic surface waters overlying anoxic bottom water existed during deposition of the Lower Tal, Yutrus (NW China), and Yanjiahae (south China) formations.
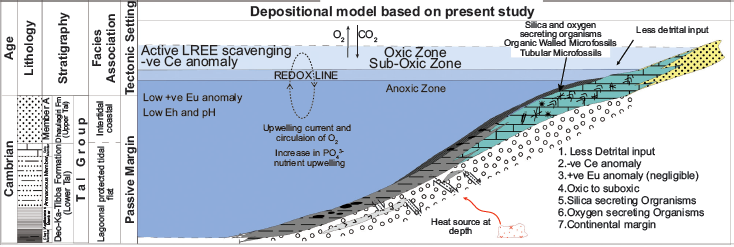
Depositional Model for Tal Group based on the present study.
CONCLUSIONS
The significant outcomes of palaeobiological and geochemical studies of chert from the Tal Formation are listed below:
This study presents the first reports of the articulated sponge fossil Paradiagoniella sp. and the large tubular microfossil Megathrix longus from India. These are significant additions to the metazoan microfossil record of the Lower Tal Formation of Krol Belt in the Lesser Himalaya. The presence of Paradiagoniella and Megathrix indicates a biostratigraphic correlation of the Lower Tal Formation with early Cambrian Chiungchussu (Nangaoan/Stage3; ~521–529 Ma) and Yanjiahe formations (Meishucunian Stage/Stage2; ~529–514 Ma) of Yangtze Platform in South China (Chen et al., 2015; Dong et al., 2009), bracketing the age range as ~514–529 Ma. The geochemical characteristics suggest that the formation of chert and phosphorite in the Tal Formation was caused by (a) biological input of silica, (b) with the influence of clastic and hydrothermal input, and (c) under oxic shallow marine conditions. The presence of cyanobacteria Obruchevella and Pleurocapsa indicates a saline and alkaline environment with pH conditions between 6.5 and ≥ 8.5. Moreover, the presence of sponges strongly indicates an oxic paleoenvironment. During the Cambrian, a stratified ocean with oxic surface waters overlying anoxic bottom eventually led to the Cambrian Explosion (Figure 13).
A. Line diagram of specimen in Plate 5 a; b. Reconstruction of specimen of Megathrix longus.
SYSTEMATICS
Filamentous and coccoidal microfossils
Kingdom: Eubacteria Woese and Fox, 1977
Phylum: Cyanobacteria Stainer et al., 1978
Class: Hormogoneae Thuret, 1875
Order: Oscillatoriales Elenkin, 1949
Family Oscillatoriaceae (S.F. Gray) Kirchner, 1900
Type species: Obruchevella Parva Reitlinger, 1948, emend. Golovenok and Belova, 1989, emend. Burzin, 1995
Plate 1, Figure 2
Description: Tight to loosely coiled hollow tubular spiral filament with ~150 µm length and 8 µm diameter. The stretched, elongated form has inner and outer diameters varying from ~20 to 38 µm and ~36 to 54 µm, respectively.
Material: One well-preserved specimen from the Masrana section.
Remarks: This form is much larger than O. parva reported from the Infrakrol Formation (Tiwari & Pant, 2004).
Occurrence: Obruchevella Parva are reported from Neoroterozoic and Early Cambrian deposits of China, India, Svalbard, Australia, Siberia and Canada (Allison & Awramik, 1989; Chuanming et al., 2007; Knoll, 1992a; Sharma & Shukla, 2009, 2012; Sharma et al., 2016; Tiwari, 1996; Tiwari & Pant, 2004).
Repository Number: WIMF/A 1925
Family: Pleurocapsaceae Geitler, 1925
Genus: Palaeopleurocapsa Knoll et al., 1975
Type species: Palaeopleurocapsa wopfneri Knoll et al., 1975
Plate 2, Figure 3
Description: Cells having fine-grained single-layer walled (~0.5 µm thick) and diameter ranging from ~5 to 12 µm are present in two semi-parallel rows. An outer spheroidal membranous layer envelops these closely packed cells making ~25 µm diameter colony.
Material: Two well-preserved specimens from the Masrana section.
Remarks: The diameter range of this specimen’s vesicles (5–12 µm) is smaller than that of the type species (4–26 µm).
Occurrence: Palaeopleurocapsa wopfneri is reported from Mesoproterozoic deposits of Australia and India (Buick & Knoll, 1999; Kumar & Venkatachala, 1998).
Repository Number: WIMF/A 1919
Type species: Palaeopleurocapsa reniforma Ogurtsova and Sergeev, 1987
Plate 2, Figure 4
Description: Tightly packed ellipsoidal to spherical cells with fine to medium grained ~1µm thick single walls and diameter ranging from ~10 to 30 µm are seen present in sub-parallel rows. Individual cells contain an opaque inclusion of ~1.0 to ~1.5 µm diameter. Two to four cells are present within a common envelope, forming a vesicle with diameter ranging from ~20 to 50 µm. These vesicles are seen forming a kidney-shaped colony (length ~300 µm; width ~150 µm).
Material: Two well-preserved specimens were observed from the Masrana and Kaudiyala sections.
Remarks: The diameter of the enveloping vesicles of these forms is slightly greater than that of specimens observed from South Kazakhstan (Knoll et al., 1975).
Occurrence: Palaeopleurocapsa reniforma is reported from Neoproterozoic deposits of South Kazakhstan (Knoll et al., 1975; Sergeev & Schopf, 2010).
Repository Number: WIMF/A 1920
Genus: Sphaerophycus Schopf, 1968
Type species: Sphaerophycus medium Horodyski and Donaldson, 1980
Plate 2, Figure 2
Description: Spheroidal to ellipsoidal vesicles occur solitary, or as dyads and triads with diameter ranging from ~10 to 25 µm.
Material: Ten well-preserved specimens observed from the Kaudiyala section.
Occurrence: Sphaerophycus medium is observed from Proterozoic deposits in North America, Siberia, India, Canada, China, Australia and Northern Europe (Hofmann & Grotzinger, 1985; Kumar & Srivastava, 1995; Schirrmeister et al., 2016; Sergeev, 2006; Yun, 1985).
Repository Number: WIMF/A 1921
Genus: Gloeodiniopsis Schopf, 1968
Synonymy: Bigeminococcus Schopf and Blacic, 1971
Eozygion Schopf and Blacic, 1971
Eotetrahedrion Schopf and Blacic, 1971
Type species: Gloeodiniopsis lamellosa Schopf, 1968, emend. Knoll and Golubic, 1979
Plate 2, Figure 1
Description: Spheroidal to ellipsoidal vesicles occur solitary, or as dyads and triads, with a diameter ranging from ~8 to 12 µm are present within a lamellated envelope. The width of the lamellated part of the envelope is ~2 to 5 µm. With the outer envelope, the diameter of specimens is ~45 to 50 µm. Dark inclusions are present in the central parts of individual cells.
Material: Two specimens were observed in the Kaudiyala section.
Occurrence: Gloeodiniopsis lamellosa is reported from Proterozoic deposits of Northwest Canada, Australia, and China (Schopf, 1968; Sergeev et al., 2012; Yuan & Hoffman, 1998).
Repository Number: WIMF/A 1922.
Genus: Myxococcoides Schopf, 1968
Type species: Myxococcoides grandis Horodyski and Donaldson, 1980
Myxococcoides grandis
Description: Single-walled solitary spheroidal cells with diameter ranging from ~30 to 50 µm. Sporadic dark granular inclusions are also present within the cells.
Material: Thirteen specimens were found in the Kaudiyala section, and six specimens were found in the Masrana section.
Occurrence: Myxococcoides grandis is reported from Proterozoic sediments of Canada, China, Siberia, Russia and India (Fuxing et al., 1983; Kumar & Srivastava, 1995; Sergeev et al., 2007; Shi et al., 2017; Veis et al., 2006).
Repository Number: WIMF/A 1926.
Genus: Myxococcoides Schopf, 1968
Type species: Myxococcoides minor Schopf, 1968
Plate 1, Figure 5
Description: Spheroidal vesicles with a diameter ranging from ~8 to 10 µm, having spheroidal to sub-spheroidal dark inclusions (~1 to 5 µm) in the centre are seen.
Material: Twelve preserved specimens from Kaudiyala and Masrana sections.
Occurrence: Myxococcoides minor is reported from Proterozoic deposits of India, Australia, China, and Russia (Ahn & Lee, 2003; Chongyu, 1990; Kumar & Srivastava, 1995; Pandey & Kumar, 2013; Sergeev, 2001; She et al., 2013; Tiwari & Pant, 2004).
Repository Number: WIMF/A 1927.
Type species: Eoentophysalis dismallakesensis Horodyski and Donaldson, 1980
Description: Three to four small spherical vesicles are present within an outer envelope ranging from ~20 to 25 µm. With an envelope, the diameter ranges from ~26 to 30 µm. The outer wall is psilate.
Material: Ten specimens from the Kaudiyala section.
Remarks: E. dismallakesensis can be differentiated from E. belcherensis by the larger diameter of its constituent vesicles.
Occurrence: Eoentophysalis dismallakesensis is reported from Mesoproterozoic deposits of North America and Semri Group of India (Horodyski & Donaldson, 1980; Sharma & Sergeev, 2004).
Repository Number: WIMF/A 1928.
Genus: Huroniospora Barghoorn and Tyler, 1965, emend. Awramik and Barghoorn, 1977
Type species: Hurnoniospora microreticulata Barghoorn and Tyler, 1965
Description: Solitary, spherical to ellipsoidal, and sometimes compressed vesicles observed with a diameter varying from ~2 to 8 µm.
Material: Eighteen well-preserved specimens were observed in the Masrana section.
Occurrence: Hurnoniospora microreticulata has been reported from Proterozoic deposits of Canada, India, and Australia (Moreau & Sharp, 2004; Pandey & Kumar, 2013; Tappan, 1980).
Repository Number: WIMF/A 1929.
Type species: Huroniospora psilata Barghoorn and Tyler, 1965
Plate 1, Figure 6
Description: Spherical smooth vesicles ranging in diameter from 8 to 18µm.
Material: Two specimens from the Masrana section.
Remarks: These forms are smaller in size than those described from the Infrakrol Formation (Tiwari & Pant, 2004; Venkatachala et al., 1990).
Occurrence: Huroniospora psilata is reported from Proterozoic deposits of China, Canada, Australia, South Africa, and India (Schopf & Barghoorn, 1969; Shixing & Huineng, 1992; Tappan, 1980; Tiwari, 1996; Tiwari & Pant, 2004; Venkatachala et al., 1990; Yun, 1984).
Repository Number: WIMF/A 1930.
Acritarchs
Subgroup: Sphaeromorphitae Downie et al., 1963
Genus: Leiosphaeridia Eisenack, 1958, emend. Downie et al., 1963, emend. Turner, 1984, emend. Yankauskas, 1989
Type species: Leiosphaeridia minutissima (Naumova, 1949), emend. Yankauskas, 1989 (in Yankauskas, 1989)
Description: Solitary, single-walled spheroidal vesicles with a diameter of ~20 to 60 µm are present. The wall of vesicles is ~1 to 2 µm thick and smooth to fine-grained.
Material: Five well-preserved specimens from Masrana and Kaudiyala sections
Remark: These forms are similar to the type species Leiosphaeridia minutissima (Yankauskas, 1989).
Occurrence: Leiosphaeridia minutissima is reported from the Neoproterozoic deposits of India, Canada, Mexico, Sweden, and Siberia (Butterfield & Chandler, 1992; Gaucher et al., 2008; Loron, 2016; Nagovitsyn & Kochnev, 2015; Tiwari, 1996).
Repository Number: WIMF/A 1932.
Plate 3, Figure 1
Description: A Large isolated spherical vesicle of ~108 µm diameter is present. The vesicle wall has a fine-grained surface and broad crenulations.
Material: One well-preserved specimen was observed in the Masrana section.
Remarks: This specimen resembles Leiosphaeridia jacutica (Yankauskas, 1989).
Occurrence: Leiosphaeridia jacutica is reported from Proterozoic sediments of Australia and Russia (Grey, 2005; Vorobeva et al., 2009).
Repository Number: WIMF/A 1933.
Group: Acritarcha Evitt, 1963
Genus: Asteridium Moczydłowska, 1991
Type species: Asteridium tornatum (Volkova, 1968), ememd. Moczydłowska, 1991
Plate 3, Figure 5
Description: Oval to spheroidal vesicle with a diameter ranging from 5 to 8 µm. Small <1 µm processes similar to knobs are present on surface of vesicles.
Material: Nicely preserved, three specimens recovered from the Masrana section.
Remarks: Specimens are similar to previous reports from the Tal Formation in the Korgai syncline in India (Tiwari, 1999). They are slightly smaller than those recovered from Spitzbergen (Knoll & Swett, 1987).
Occurrence: A. tornatum is reported from India and Svalbard (Knoll & Swett, 1987; Tiwari, 1999)
Repository: WIMF/A 1980.
Tubular microfossil
Genus: Megathrix Yin L., 1987, emend.
Type species: Megathrix longus Yin L., 1987, emend. Unnamed filamentous fossils Yin L., 1986
Megathrix longus, Yin L., 1987
Megathrix sp. A, Yin L., 1987
Megathrix sp. B, Yin L., 1987
Megathrix sp. C, Yin L., 1987
Megathrix sp. D, Yin L., 1987
Megathrix sp., Yin L., 1987
Megathrix longus, Yin L., 1987; Yin et al., 1992
Megathrix longus, Yin L., 1987; Yin L., 1997
Megathrix longus, Yin L., 1987; Yin et al., 2003
Description: A Solitary, large, curved, and compressed tube is present. The length of the tube is ~310 µm, and its maximum diameter is ~48 µm. It has unequal, closely spaced, irregularly crenulated cross walls.
Material: One well-preserved specimen from the Masrana section. Peculiar corrugated cross walls are present in this specimen.
Remarks: Yao et al. (2005) suggested that corrugated cross walls are never present in a filament. M. longus possesses corrugated and perforated cross-walls, whereas Oscillatoriopsis-like cyanobacteria possess complete transverse septa. So, the present form should not have cyanobacterial affinity. Also, M. longus is analogous to Sinocyclocyclicus guizhouensis, which is interpreted as a stem group cnidarian from the Neoproterozoic Doushantuo Formation in South China (Chen et al., 2002; Xiao et al., 2000; Xue et al., 1992). Reconstruction of the present form is shown in Figure 5.
Occurrence: Megathrix longus is an index fossil which is observed from Cambrian/Meishucunian Stage deposits of the Yangtze Gorges area and the Yurtus and Xishanblaq formations in Tarim in China (Dong et al., 2009; Yao et al., 2005; Yin, 1987).
Repository Number: WIMF/A 1934.
Biomineralised microfossils
Description: Several scattered siliceous spicules are present in the assemblage. These spicules are commonly present in isolation or groups. Spicules ranging in size from ~501 to ~1200 µm and above have been classified as first-order spicules, followed by a range of ~200 to ~500 µm of second-order. The size range between ~100 and ~200 µm is grouped into the third order. Spicules ranging from ~50 to ~100 µm are classified into fourth-order spicules, followed by ~40 to ~50 µm length-sized fifth-order spicules. Spicules varying in size from ~10 to ~50µm are classified into the sixth order. Their length ranges from ~100 to ~1105 µm, with the latter being the largest. They are in acicular forms with ~50 to 600 µm length and ~ 2 to 4 µm width and hexactins (with six rays) with a size range of ~10 to 50 µm (microscleres) and 50 to ~1105 µm (megascleres). Hexactin shows well-preserved four to six rays at right angles to each other that taper distally with sharp ends. A typical cross-shaped structure is formed at the junctions of these spicules. In one specimen, the transverse section of an isolated spicule is ~350 µm long and ~45 µm wide. Most of the spicules contain secondary material (phosphatic) that shows exceptional preservation of axial canals.
Remark: Spicules are scattered, so reconstructing the sponge framework is not feasible. Individual spicules are identified based on the well-preserved axial canal, triaxon symmetry, and cross shape at the ray junction.
Number of specimens: 75 spicules are measured from the Masrana and Kaudiyala sections.
Occurrence: Hexactinellid spicules are reported from Neoproterozoic deposits of Mongolia, India, North America, Australia, and Russia (Brasier et al., 1997; Mazumdar & Banerjee, 1998; Rigby, 1987; Rozanov & Zhuravlev, 1992; Tiwari, 1999).
Repository Number: WIMF/A 1936–1950.
Kingdom: Animalia
Phylum: Porifera Grant, 1836
Class: Hexactinellida Schmidt, 1870
Order: Reticulosa Reid, 1958
Super family: Protospongioidea Finks, 1960
Family: Protospongiidae Hinde, 1887
Genus: Paradiagoniella Chen et al., 2015
Type species: Paradiagoniella
Description: Hexactinellid spicules/stauractines are observed arranged mutually, forming an articulated sponge skeleton. First/- and second-order spicules are oriented in parallel or subparallel arrangement. Second-order spicules are arranged in an irregular quadrilateral habit. Lower-ranked, smaller stauractines show oblique orientation and chaotic occurrence. The smallest oxeas (straight or curved diactines) are scattered within thin stauractines and are sometimes seen as interlocked. They are not in parallel arrangement with principal stauractines. The present specimen has stauractines that are ranked in size with first-order principal stauractines arranged irregularly, but forming a local subquadrule oblique to the body axis. Smaller stauractines of lower orders are embedded in the space between principal stauractines and are oriented independently of principal stauractines. It also consists of rare hexactines.
Material: One well-preserved specimen reported form Kaudiayala Section
Remarks: Paradiagoniella n. gen. can be differentiated from other protospongiids in the presence of oxeas in its skeletal structure (Chen et al., 2015). This specimen is quite comparable to Paradiagoniella magna described by Chen et al. (2015) from the Chengjiang Formation in South China, which has large principal stauractins present obliquely diagonal to the longitudinal axis of the sponge body with six orders of irregularly arranged quadrules and subquadrules. It differs from P. magna, which contains stauractines of different orders articulated obliquely and interlocked with diactines to form an irregularly arranged skeleton.
Occurrence: Paradiagoniella is observed in China from the Cambrian Chengjiang biota (Chen et al., 2015).
Repository Number: WIMF/A 1951–1953.
Footnotes
Acknowledgements
The authors are thankful to the Director of the Wadia Institute of Himalayan Geology, Dehradun, India, for providing the necessary working facilities.
Authors Contribution
Meera Tiwari (MT), Meenal Mishra (MM), Harshita Joshi (HJ); Conceptualization: MT, HJ; Formal analysis: MT, MM, HJ; Investigation: MT, HJ; Supervision: MT; Writing- original draft: HJ; Writing-review and editing: MT, MM.
Consent to Participate
Not applicable.
Consent for Publication
Not applicable.
Data Availability Statement
Microfossil specimens are present in repository of Wadia Institute of Himalayan Geology. The datasets used or analysed during current study are available with the corresponding author and can be obtained on reasonable request, and all data generated or analysed during this study are included in the manuscript.
Declaration of Conflicting Interests
The authors declared no potential conflicts of interest with respect to the research, authorship and/or publication of this article.
Ethical Consideration
Not applicable.
Funding
The authors disclosed receipt of the following financial support for the research, authorship and/or publication of this article: Harshita Joshi received a Research Fellowship from the Wadia Institute of Himalayan Geology, Dehradun.