
Abstract
The Sonia Sandstone of the Jodhpur Group, Marwar Supergroup in western Rajasthan, has unveiled many fossil assemblages, significantly advancing our comprehension of the Ediacaran biotic diversity and palaeoecological dynamics. Recent field discoveries within the Sonia Sandstone include a diverse suite of Ediacaran fossils, such as Tirasiana qingzhenensis, Hiemalora stellaris, Pteridinium sp., Finkoella ukrainica, Finkoella oblonga, Arumberia banski, coiled form, bizarre form A, Quaestio simpsonorum, and jellyfish-like impressions. These fossils, preserved predominantly as positive epirelief on the bedding plane, exhibit an extraordinary range of morphological intricacies, suggesting a multifaceted and dynamic Ediacaran ecosystem. This unprecedented palaeontological record sheds light on the ecological complexity and evolutionary trajectories during the Ediacaran Period, contributing to the broader understanding of early metazoan diversification and sedimentary biogenic processes.
INTRODUCTION
The Marwar Supergroup encapsulates a pivotal geological interval, spanning late Ediacaran to the early Cambrian Period (Kumar & Pandey, 2008, 2010; Kumar et al., 2009; McKenzie et al., 2011; Pandey & Sharma, 2017). This time duration was instrumental in shaping the contemporary biosphere with complex and modular Ediacaran organisms, more akin to animals, that appeared for the first time in the Earth’s history (Bobrovskiy et al., 2018; Peng et al., 2020; Xiao & Narbonne, 2020). However, the affinity of the Ediacaran fossils is still debatable. Many of these forms were described under various groups: Vendobionta (Seilacher, 1989, 1992), protists (Zhuravlev, 1993), prokaryotes (Steiner & Reitner, 2001), mesozoans (McMenamin, 1998), lichen (Retallack, 1994), fungi (Peterson et al., 2003; Retallack & Landing, 2014), fungi, lichens and encrusting algae (Droser et al., 2014); macroalgae (Pandey & Sharma, 2017; Wan et al., 2020). Furthermore, another debate is about the preservation; where and in what environment did they evolve and preserve? Recent findings of field data and geochemical proxies, such as REE, are helpful to reveal marine versus freshwater Ediacaran fossils bearing horizons (Retallack, 2024). It is postulated that during the Neoproterozoic Oxygenation Event (NOE), which occurred approximately 850–540 million years ago, atmospheric oxygen reached values near to those of the present day (Lenton et al., 2014). This oxygen level and availability of micronutrients were adequate to support the emergence of complex eukaryotes, which experienced significant diversification around 600 million years ago (Sahoo et al., 2012; Wan et al., 2020; Xiao & Laflamme, 2009). Consequently, the Cambrian Period, marked by the rapid emergence of numerous new and more complex species, likely has its roots in the late Ediacaran period. One of the hallmark evolutionary traits of the Cambrian Period was the advent of skeletal fossils, a process that began in the late Ediacaran (Maloof et al., 2010; Wood, 2011). Such diversity of the Ediacaran–Cambrian transition is marked in the Marwar Supergroup. Over the past decade, numerous fossils from the Sonia Sandstone have been discovered, establishing the basin’s lower boundary as the Ediacaran period (see Pandey & Sharma, 2017 and references therein). The present study represents the suite of new Ediacaran fossils reported from the Sonia Sandstone (Jodhpur Group), which not only enriches the diversity of Ediacaran macrofossils in the Marwar Supergroup but also enhances our understanding of the origin and diversification of life on the onset of the Cambrian explosion.
GEOLOGICAL CONTEXT
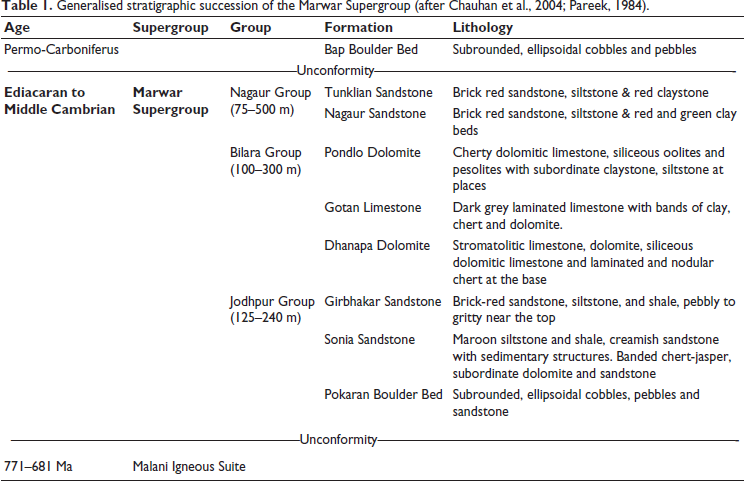
The Marwar Supergroup, formerly known as the Trans-Aravalli Vindhyan (Heron, 1932) (Figure 1), is situated west of the Aravalli Mountain range and distinct from the Vindhyan Supergroup rocks of the Chambal Valley region (Kumar, 2012). It unconformably overlies the Malani Igneous Suite and older metamorphic formations (Pareek, 1981, 1984) and dates back to 779 ± 5 Ma (Gregory et al., 2009). The Marwar Supergroup comprises thick deposits of argillo-arenaceous and carbonate facies in the Jodhpur-Khatu-Bikaner-Barmer, Nagaur-Phalodi regions of western Rajasthan, with an approximate thickness of 1000 meters (Chauhan et al., 2004; Pandey & Bahadur, 2009; Pareek, 1981, 1984). The Marwar Supergroup is lithostratigraphically classified into three groups and eight formations (Table 1). The three groups are the Jodhpur Group, the Bilara Group, and the Nagaur Group (Figure 2). The calcareous Bilara Group is sandwiched between the two siliciclastic units, such as the Jodhpur and Nagaur Groups (Pareek, 1984). Further, the Jodhpur Group is subdivided into three formations: the Pokaran Boulder Bed, the Sonia Sandstone, and the Girbhakar Sandstone. Similarly, the Bilara Group comprises the Dhanapa Dolomite, Gotan Limestone, and Pondlo Dolomite. The Nagaur Group represents the youngest succession of the Marwar Supergroup, overlies the Bilara Group, and is further divided into the Nagaur Sandstone and Tunklian Sandstone.
Generalised stratigraphic succession of the Marwar Supergroup (after Chauhan et al., 2004; Pareek, 1984).
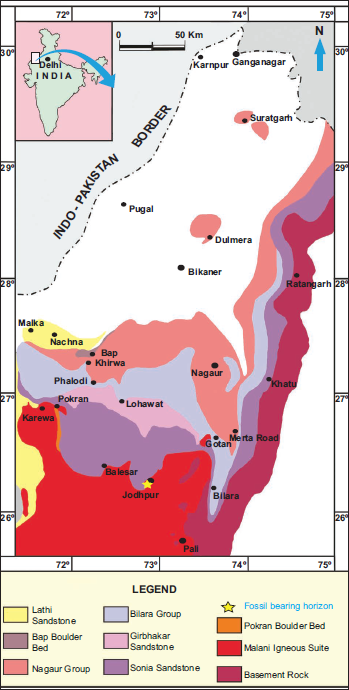
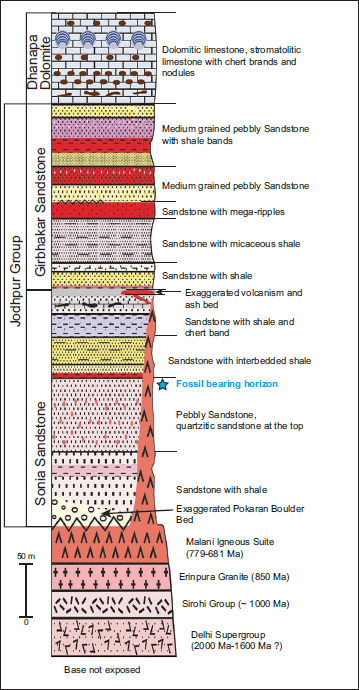
In the southwest portion of the basin, close to Pokaran town, the 4 m thick Pokaran Boulder Bed is exposed. However, the Sonia Sandstone lies above the Malani Igneous Suite in other areas. A detailed sedimentological investigation of the Sonia Sandstone was performed by Sarkar et al. (2008), who described three different marine facies: lower (A), middle (B), and upper (C). The upper and lower (C and A) facies symbolise a wave-dominated shallow to deep neritic environment—middle facies (B), characterised as a supralittoral zone of the backshore–foreshore environmental condition. The variability in palaeoslope gradients, lack of vegetation, and base-level fluctuations influenced the fluvial-to-marine transition of the Sonia Sandstone. The fossil-bearing horizon of the Sonia Sandstone must have been deposited within the shallow, moderate-energy foreshore environment, marking the ancient Sonia Sea (Pandey & Sharma, 2017).
The Sonia Sandstone: An Ediacaran fossil-Lagerstätten
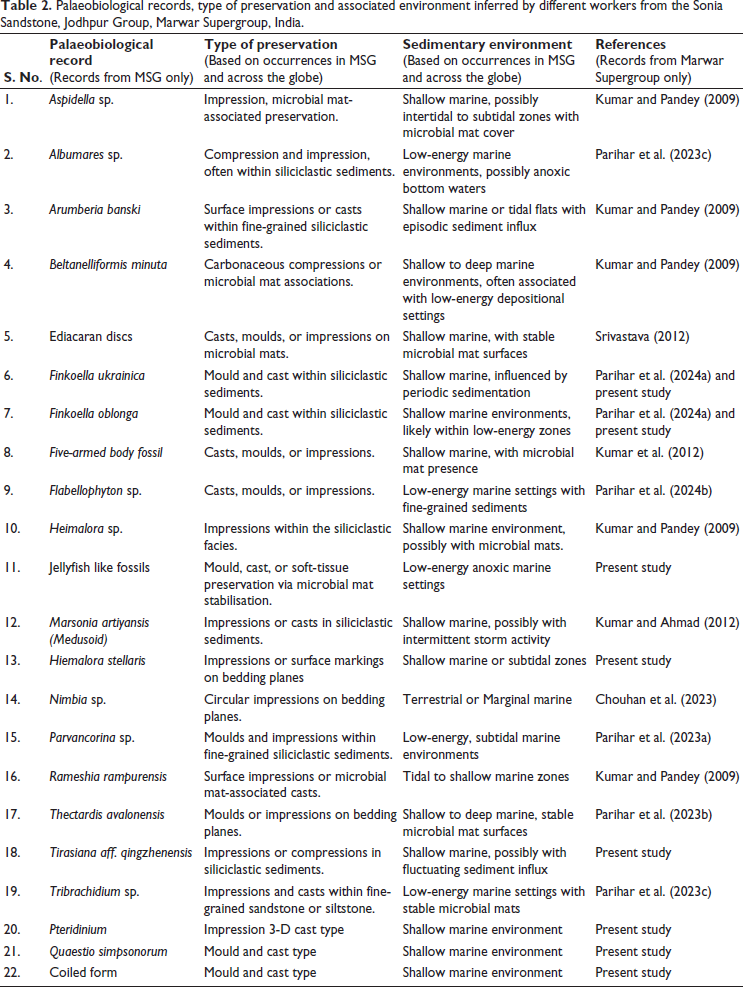
Over the past two decades, the Sonia Sandstone has gained prominence as an Ediacaran fossil-bearing unit offering valuable insights into early life and evolution (Table 2). The very first promising report by Raghav et al. (2005) documented the medusoid body fossil Marsonia artiyansis in the lower part of the Sonia Sandstone, which was later reviewed by Kumar and Ahmad (2012). The Ediacaran body fossils, including Aspidella sp., Hiemalora sp., Beltanelliformis minuta, and organo-sedimentary structures Arumberia banksi and Rameshia rampurensis, were reported by Kumar and Pandey (2009). However, the original specimen of Arumberia banksi was recently examined by Retallack and Broz (2020), who regarded it as a quilted body plan (body fossil) of the late Ediacaran-aged Class Vendobionta. Additional discoveries, such as a possible five-armed echinoderm fossil (Kumar et al., 2012) and large, non-carbonaceous Ediacaran discs (Srivastava, 2012, 2014), further underscore the rich biosphere within the Sonia Sandstone. The organic-walled microfossils and acritarch assemblage comprise the Lophosphaeridium rarum- Granomarginata prima zone recovered from the BGW-A well (subsurface profile) by Prasad et al. (2010) and inferred as Late Ediacaran age for the Sonia Sandstone. Noteworthy features include microbial mat structures (Kumar & Ahmad, 2014; Samanta et al., 2011) and preserved putative giant seaweeds that may have supported the existence of large-sized Ediacaran macroalgal fossils (Kumar et al., 2009; Kumar & Ahmad, 2016; Pandey & Sharma, 2017). Although iconic fossils like Charniodiscus, Dickinsonia, and Kimberella are absent, recent findings from the Sonia Sandstone reflect close affinity with the White Sea and Nama assemblages, which enhance our understanding of early metazoan and metaphyte life on Earth.
Palaeobiological records, type of preservation and associated environment inferred by different workers from the Sonia Sandstone, Jodhpur Group, Marwar Supergroup, India.
This study highlights a diverse array of Ediacaran fossils from the Jodhpur Sandstone, including Tirasiana qingzhenensis, Hiemalora stellaris, Finkoella ukrainica, Finkoella oblonga, coiled fossils, Arumberia banski, an enigmatic form ‘A’ and jellyfish-like fossils. Most fossils are preserved as positive epireliefs, while some appear as negative hyporeliefs. The fossil assemblage not only reinforces the biostratigraphy of the Sonia Sandstone but also provides insights into the evolutionary processes and environmental factors that shaped early life.
Arumberia banski Glaessner & Walter, 1975
(Plate 1, Figures 1 and 2)
Background Information: Arumberia banski was initially described as a cnidarian body fossil (Glaessner & Walter, 1975), which was later reinterpreted as an inorganic feature, specifically a flute cast formed on microbial mat surfaces (McIlroy & Walter, 1997). Over the years, affinity has been inferred by different workers, such as close to Vendobionts (Ernietta and Pteridinium) (Bland, 1984), to trace fossils (McIlroy et al., 2005), and to organo-sedimentary structures (Kumar & Ahmad, 2014; Kumar & Pandey, 2008; Liu et al., 2011). Arumberia is considered one of the significant fossils documented in the Late Ediacaran Period worldwide (Ahmad & Kumar, 2014; Kumar & Pandey, 2008, 2009; McMahon et al., 2022; Retallack & Broz, 2020). Two Recent critical reviews of Arumberia have been conducted by Retallack and Broz (2020) and McMahon et al. (2022), and discussed the significance.
Description: The specimens observed in the field were identified as Arumberia banski, preserved as concave ellipsoidal structures with the deepest part situated near one end of a rippled surface (Plate 1, Figures 1 and 2). They exhibit ridges radiating from the deepest point toward the margins, occasionally bifurcating. Some ridges form at angles of 10°–20°, with finer ridges appearing between them. Individual ridges measure 4.0–12 cm in length and 0.8–2.4 cm in width, with a spacing between 0.2 and 0.4 cm. The preservation was observed on both the top and bottom of the Sonia Sandstone.
Paleontological Significance: This study recorded exceptionally well-preserved specimens of Arumberia banski in the Sursagar Mine area, marking the first confirmed report from this locality. However, Sharma and Mathur (2014) described Arumberia-like mat structures from the nearby Bhuri Beri region earlier. Arumberia banski was also reported from the lower part of the Sonia Sandstone exposed at Chhoti Khatu hillock section in the Nagaur district of Rajasthan (Kumar & Pandey, 2009).
Arumberia is considered a valuable biostratigraphic marker for the Ediacaran–Cambrian transition (Bland, 1984). The type specimen, recently re-examined by Retallack and Broz (2020), was interpreted as a quilted body structure of the Vendobionta class, suggesting biological similarities with other quilted organisms like Ernietta. These features indicate complex ecological interactions and structural adaptations during the late Ediacaran period. McMahon et al. (2022) propose that it represents extinct sessile filamentous organisms surviving from the terminal Ediacaran into the early Cambrian. This reinforces the hypothesis that Arumberia-bearing Sonia Sandstone layers date back to the late Ediacaran. Furthermore, the occurrence of Arumberia in both Ediacaran and Cambrian deposits, such as the Arumbera Formation, challenges the notion of a severe extinction event in the Ediacaran–Cambrian transition, suggesting the survival and adaptive nature of some of the Ediacaran fossils. Notably, Arumberia banksi was preserved in the same foreshore lithofacies, where Sonia Seaweeds were documented by Kumar and Ahmad (2016) and Pandey and Sharma (2017).
Tirasiana qingzhenensis Yang, 2010
(Plate 1, Figures 3 and 4)
Background Information: Tirasiana qingzhenensis is a type of Ediacara-like fossil originally described from the Lower Cambrian Taozichong Formation in Qingzhen County, Guizhou Province, South China. The occurrence of this taxon in Cambrian strata provides valuable insights into the evolutionary transition at the Ediacaran–Cambrian boundary (Yang, 2010). The Ediacaran fossils are traditionally associated with the Ediacaran Period. However, the presence of T. qingzhenensis alongside Cyclomedusa sp. in Chinese Cambrian deposits indicates the persistence of some Ediacaran taxa into the Phanerozoic, contrary to the belief that the terminal Neoproterozoic mass extinction completely eradicated this biota. The fossils exhibit discoidal morphologies and other structural features strongly suggesting a biological origin. Their stratigraphic position is constrained by the underlying Anabarites-Protohertzina Zone and the overlying appearance of the trilobite Zunyidiscus sp., confirming their Early Cambrian age in the Chinese section.
Description: Tirasiana qingzhenensis (Plate 1, Figures 3 and 4) preserved as simple convex discoid impressions on the bedding plane of the Sonia Sandstone, Jodhpur Group. These fossils exhibit a distinct morphology, characterised by a central nipple-like protuberance measuring 2.0 cm in diameter and approximately 1.0 mm in height, surrounded by three to five radially arranged ridges that slightly widen outward and form the body outline. A total of four to five specimens were observed in the field in the Sursagar mine area of Jodhpur district. They left an impression on the bedding plane of fine-grained sandstone facies. The present specimen from the Sonia Sandstone of the Marwar Supergroup has been reported for the first time.
Palaeontological Significance: The discovery of Tirasiana qingzhenensis has significant implications for understanding the evolutionary history and behavioural traits of early metazoans in the context of the Marwar Supergroup. This fossil marks its presence in the Ediacaran Sonia Sandstone, demonstrating a continuity of life forms across the Ediacaran–Cambrian boundary. This challenges the traditional narrative that the Ediacaran biota was entirely replaced by Cambrian life. T. qingzhenensis exhibits notable morphological variability, such as nipple-like protuberances and annular ridges, adding to the diversity of known Ediacara-type fossils. The extended stratigraphic range of Ediacaran-type organisms into the Cambrian period suggests that some lineages of this biota might have contributed to the ‘Cambrian explosion’ of life forms, albeit in modified skeletal or morphological forms. This underscores their widespread distribution and potential ecological significance during the Late Ediacaran-Early Cambrian transition.
Hiemalora stellaris Fedonkin, 1980
(Plate 1, Figure 5)
Background information: Genus Hiemalora was first identified in the Vendian sequence of the White Sea, Russia. Later, this species was reported from various Ediacaran global localities including Canada, Siberia, Ukraine, Norway, India, and Australia (Burzynski & Narbonne, 2015; Farmer et al., 1992; Fedonkin, 1980; Gureev, 1988; Hofmann et al., 2008; Kumar & Pandey, 2009) which ranges from 565 to 550 Ma (Chen et al., 2014). Hiemalora and its affinity are still debatable. According to specific interpretations, Hiemalora is either a body fossil with unknown affinities or a hydrozoan (Fedonkin, 1980), whereas Anderson and Conway Morris (1982) interpreted it as having unknown affinities. Later, Hofmann et al. (2008) considered it as a discoidal holdfast with radiating root-like rays. Morphologically, the diameter of the central disc of Hiemalora can also vary and/or sometimes be absent in the rock record (Serezhnikova, 2007). Only two species have been identified: Hiemalora pleiomorpha from Siberia (Vodanyuk, 1989) and Hiemalora stellaris from Russia (Fedonkin, 1980, 1982). Parihar et al. (2021) reported Hiemalora stellaris from the Sonia Sandstone, but the close examination reveals that the preservation is insufficient to assign the structures to the genus and species level.
Fossil Description: The Present specimen of the genus Hiemalora (Plate 1, Figure 5) is preserved on the bedding plane of the fine to medium grained Sonia Sandstone exposed in the Sursagar mine area, Jodhpur district, Rajasthan, and morphologically similar to Hiemalora stellaris exhibits externally radiating, closely spaced, fairly elongated, narrow rays of varying length attached to the rim. Rays are typically unbranched, rectilinear to slightly sinuous, but they hardly cross over one another. The fossils measure 8 cm in diameter and about 0.4 mm in elevation. 8–10 rays are protruding outside, outlining the body. The length of the ray ranges between 4–6 cm, and its width is about 0.7 cm.
Palaeontological significance: Hiemalora stellaris from the Sonia Sandstone constitutes a remarkable Ediacaran fossil find, illuminating the emergence of complex multicellular life. This discoidal fossil, preserved as prominent positive relief on medium to fine-grained sandstone surfaces, exhibits a distinct central circular body encircled by radiating, tentacle-like appendages. Their presence Hiemalora, within the shallow marine foreshore depositional setting supports it as a benthic, soft-bodied organism that thrived during the deposition of the Ediacaran Sonia Sandstone. The identification of Hiemalora not only enriches our understanding of palaeoenvironmental dynamics but also enhances our understanding of the evolutionary trajectory of the Ediacaran biota, corroborating analogous fossil records from Russia and Newfoundland (Fedonkin, 1980; Gehling et al., 2000; Hofmann et al., 2008).
Pteridinium sp.
(Plate 1, Figure 6)
Background information: Pteridinium is a significant genus of the Ediacaran biota (erniettomorph taxon), distinguished by its unique three-dimensional body plan exhibiting triradial symmetry recorded from late Ediacaran successions in South Australia, Russia, and Namibia. Diagnostic features include a distinctive quilted pattern of parallel, tubular, or rib-like structures arranged in rows, forming a leaf-like or frondose appearance (Meyer et al., 2014; Narbonne, 2005). Three vanes joined along an axis called the ‘seam’ make up the distinctive body layout of the Pteridinium. According to Jenkins (1992), each vane comprises a single row of segments fastened to the seam, or maybe two overlapping rows.
Description: The specimens (n = 2) were recorded from the Sonia Sandstone exposed in a hillock near Ostran village, further west of Bhopalgarh town in Jodhpur district, Rajasthan. The specimen measures 11 cm in total length and 9 cm in width, with eight visible vanes. Each vane ranges from 6 to 8 cm in length and 1.6–2.5 cm in width. However, a half-broken specimen was found in the field, in which one side of the lateral vanes is missing (Plate 1, Figure 6). Pteridinium is generally preserved as impressions within fine-grained sedimentary rocks, often associated with microbial mats, which are key in promoting fossilisation (Gehling, 1999). Likely benthic, Pteridinium is thought to have inhabited low-energy marine environments associated with siliciclastic sediments (Darroch et al., 2022), similar to the conditions observed in the Sonia Sandstone of the Marwar Supergroup. One specimen is deposited in the repository of the museum of Birbal Sahni Institute of Palaeosciences (specimen no. BSIP-42516; statement no. 1666).
Palaeontological Significance: Pteridinium holds significant importance in understanding early multicellular life and the ecological structure of the Ediacaran period. The phylogenetic placement remains controversial, with hypotheses suggesting affiliations with Vendobionta (a proposed clade of Ediacaran organisms), cnidarians, or stem-group metazoans (Xiao & Laflamme, 2009). Its association with microbial mats suggests a detritivorous or absorptive feeding mode, leveraging microbial interactions in nutrient cycling (Darroch et al., 2022). The intricate body plan of Pteridinium challenges the simplistic view traditionally associated with Precambrian life, providing insights into the evolution of tissue differentiation and multicellularity (Laflamme et al., 2013; Seilacher, 1992). According to Darroch et al. (2018), Pteridinium is one of the few taxa that survived until the end of the Ediacaran Period, although many Ediacaran creatures went extinct close to the White Sea-Nama assemblage boundary (~548 Ma). Fossil-bearing facies with ripple marks, cross bedding, and many other sedimentary features suggest a dynamic depositional environment influenced by tides and storms. In the present study, a half-broken/incomplete specimen of Pteridinium was reported from the Sonia Sandstone. However, completely preserved specimen needs to be explored for further understanding.
Finkoella ukrainica Martyshyn & Uchman, 2021
(Plate 4, Figures 1 and 2)
Field photographs of the Sonia Sandstone, Jodhpur Group, Marwar Supergroup. 1: Arumberia banksi preserved on the rippled surface (1: space between two sets of ridges, 2: ridges radiating from the deepest point toward the margins, and bifurcating (see sketch), 3: length and width of the ridges of A. banksi). 2: Arumberia bearing a huge rock slab within the Sonia Sandstone (see inset in Plate 1, Figure 1); 3: Tirasiana qingzhenensis preserved as positive epirelief. 4: Line diagram of Tirasiana qingzhenensis. 5: Hiemalora stellaris. 6: Incomplete specimen of Pteridinium sp. Scale: 2. hammer head = 17 cm; 5. diameter of coin = 2.3 cm; 6. width of hammer head = 2.8 cm.

Field photographs are from the Sonia Sandstone, Jodhpur Group, and Marwar Supergroup. 1: Finkoella ukrainica preserved as positive epirelief. 2: Line diagram of Finkoella ukrainica showing its oblong sac-like morphology. 3: Finkoella oblonga displaying longitudinal lines. 4: Line diagram of Finkoella oblonga. 5: Two specimens of coiled fossils with loose whorls preserved on the same bedding plane (line diagram provided for clarity). 6: Quaestio simpsonorum, 7: Bizarre Form ‘A’ 8: Jellyfish-like fossil preserved as negative epirelief. Scale: 4, 7. Diameter of lens cover = 5.5 cm; 5 8. Diameter of coin = 2.3 cm.

Background Information: Finkoella ukrainica represents a sack-shaped fossil of an organism resembling Ediacaran tunicates. Described for the first time in 2021 from Ukraine, the genus comprises two species: F. ukrainica (the type species) and F. oblonga (Martyshyn & Uchman, 2021). The genus name honours S. V. Finko, an amateur palaeontologist who provided the holotype specimen of F. Ukrainica from Ukraine. While earlier reports by Parihar et al. (2024a) documented fossils from a similar horizon, the specimens analysed here exhibit superior preservation.
Fossil Description: Four specimens of F. ukrainica (Plate 4, Figures 1 and 2) were observed in the field. These fossils exhibit oval to elliptical forms, ranging up to 12 cm in length (average of 8 cm) and 3.8 cm in width (average of 2.8 cm). The fossils often include a short, neck-like projection and/or shallow sinuses, with a circular depression or protrusion frequently appearing within their convex or concave features. Smaller examples lack the neck-like projection and additional surface structures. One end is more elongated, while the other is rounded and features a protruding neck that widens unevenly into sharp-lobed margins. These fossils are preserved on top of the bedding plane of the Sonia Sandstone, exposed in the Sursagar area of Jodhpur district, Rajasthan.
Palaeontological Significance: The discovery of F. ukrainica enhances our understanding of Ediacaran biota and provides insight into early metazoan evolution. Its sack-like morphology suggests an affinity with early tunicates, shedding light on the structure and diversity of soft-bodied Ediacaran organisms. The presence of Finkoella species in Ukraine and India points to a broader geographical range during the Ediacaran Period, potentially indicating environmental connections across distant regions.
Finkoella oblonga Martyshyn & Uchman, 2021
Background Information: Finkoella oblonga is another species within the genus, characterised by its elongated sack-like morphology. The name ‘oblonga’ derives from the Latin term for ‘elongate’, reflecting the species’ shape. Finkoella oblonga was recorded from the Ediacaran shallow-marine deposits of Ukraine for the first time (Martyshyn & Uchman, 2021).
Fossil Description: The fossils of F. oblonga (Plate 4, Figures 3 and 4) are cucumber-shaped, measuring approximately 12 cm in length and 7 cm in width. They exhibit longitudinal ridges and grooves, with about seven such features running parallel to the fossil’s margins. These grooves turn at the ends to form concentric rings, which are more prominent on one side while appearing fragmented on the other. The surface displays numerous small corrugations lacking an organised pattern. Edges of the fossil are either slightly elevated or depressed, often showing a double ridge around 1 mm wide. These specimens are preserved on the bedding plane of the Sonia Sandstone in a hillock section near Bhopalgarh town in Jodhpur district, Rajasthan.
Palaeontological Significance: The distinct morphology of F. oblonga highlights the diversity of Ediacaran body plans and suggests varying ecological adaptations. Its sack-like form supports potential affinities with early tunicates, providing valuable information on chordate-related evolution. The presence of Finkoella fossils in Ukraine and India illustrates the wide palaeogeographical distribution of these organisms during the Ediacaran period. Moreover, the fossils contribute to understanding the sedimentary environments and taphonomic conditions that allowed for preserving soft-bodied organisms in the Ediacaran Period.
Quaestio simpsonorum Evans et al. 2024
(Plate 4, Figure 6)
Background information: Quaestio simpsonorum was recently described as an Ediacaran fossil from the Rawnsley Quartzite of the Nilpena Ediacara National Park (NENP) in South Australia. It is preserved as a shallow circular impression, characterised by a prominent, curved ridge resembling a backwards question mark. The ridge displays a distinctive left-right asymmetry or handedness, suggesting bilateral organisation. The organism likely exhibited anterior-posterior (AP) polarity and possible dorsoventral (DV) differentiation, indicating a level of body organisation not commonly observed among Ediacaran taxa. Morphologically inferred as a mobile, soft-bodied metazoan with chirality, potentially possessing sensory capabilities and coordinated movement. These characteristics imply an advanced level of morphological complexity compared to most Ediacaran organisms, reflecting a significant evolutionary innovation during the late Neoproterozoic.
Fossil description: Quaestio simpsonorum is an intriguing Ediacaran fossil characterised by a central depression or feature resembling a shallow imprint, possibly indicating the original structure or organism preserved within the sedimentary rock (Plate 4, Figure 6). The fossil is preserved as a negative epirelief on the bedding plane of the Sonia Sandstone in the Sursagar area of Jodhpur district, Marwar Supergroup. It exhibits prominent lines forming the outline of the body, with the form measuring approximately 9–10 cm in length and an outline width of about 0.5 cm. The maximum depression observed is 1.6 cm. The centrally located circle is not well preserved. The overall morphological traits represent plausible remains of a holdfast or soft-bodied organism. The concentric outline and radiating grooves or lines may imply a symmetry, potentially reflecting its original shape or anatomy, suggesting an organic origin. Only two field specimens were recorded and observed, of which one is incomplete. More field scrutiny is required for further understanding.
Palaeontological significance: The discovery of Quaestio simpsonorum within the Marwar Supergroup represents a pivotal finding for understanding the evolution of early metazoan body plans and motility in the late Precambrian. Also highlights the emergence of advanced morphological features such as bilateral symmetry, anterior-posterior polarity, and dorsoventral differentiation within shallow marine settings. The fossil’s characteristic curved ridge and associated trace fossils point to a mobile and epibenthic lifestyle, suggesting that motility and directional movement had evolved well before the Cambrian explosion. The occurrence of Quaestio simpsonorum in the Marwar Supergroup enriches the Ediacaran biota’s diverse record in India and aligns with global evidence of complex multicellular life from the same period. This discovery challenges the traditional view of the Ediacaran as predominantly static. It reinforces the idea of significant behavioural and morphological evolution before the Phanerozoic, marking a crucial step towards diversifying animal body plans.
The coiled fossil (Plate 4, Figure 5) appears coiled with a central circular object, resembling Aspidella Billings (1872), but features a single loose whorl. It is preserved as a positive epirelief with 1–3 mm relief, the fossil’s diameter is 4 cm, and the width of the coiled body ranges from 1 to 3 cm with a central depression of 0.5 cm relief. A total of three specimens have been observed in the field (Sursagar mine, Jodhpur district, India). Present specimens closely resemble the horizontal spiral trace fossil described from the Brioverian Series (Ediacaran–Cambrian) in Brittany, NW France (Gougeon et al., 2022). Furthermore, similar to the Grypania spiralis described from the Ediacaran Doushantuo Formation of South China (Wang et al., 2016). Thus, in the present status, more specimens are required to describe the coiled form systematically.
(Plate 4, Figure 7)
Form A, under the category of bizarre form, is preserved on top of the bedding plane of the Sonia Sandstone with symmetrical ripple in the Sursagar area of Jodhpur district. Form A is characterised by a filament/ribbon-like form, is highly entangled, and the length cannot be measured correctly. The width of the filaments ranges between 0.5 and 1.0 cm. The underlying ripple mark has not been disturbed, which rules out its inorganic origin and suggests it was preserved after the formation of the ripple bed. It displays an intricate and irregular outline, suggesting a network or branching structure. The outline appears delicate, but uneven, with potential lobes or extensions radiating outward. It resembles impressions like features, which may be the remnant of microbial mats/seaweed or a delayed remnant of jellyfish or related forms. This form closely resembles Streptichnus, recorded from Namibia’s Ediacaran(?)-Cambrian succession (Jensen & Runnegar, 2005). However, these filaments neither radiate from the central area of the fossil organisation, nor tightly coiled and preserved fairly in three dimensions.
(Plate 4, Figure 8)
Background Information: A jellyfish-like fossil showcases a fossil impression resembling a jellyfish, preserved on top of the bedding plane of the Sonia Sandstone in the Sursagar area of Jodhpur district. The fossil exhibits a circular outline with faint radial symmetry, characteristic of soft-bodied organisms. The central portion appears slightly raised or more defined, potentially representing a jellyfish’s bell structure (more akin to Exumbreller surface). At the same time, the surrounding areas may correspond to its tentacles or, most likely, oral arms of dead jellyfish. The impression suggests that the fossil was preserved in a fine-grained sediment layer, likely under a low-energy depositional environment. Such a structure is often associated with soft-bodied organisms of the Ediacaran biota. In the absence of frondose arrangement, and presence of bulbous holdfast system, the present specimen is more akin to being inferred as a dead jellyfish with oral arms rather than the genus Arkrophyllus (Grimes et al., 2024). However, the specimen was observed in the field, but a thorough field investigation with more similar specimens is required to establish its affinity.
Description: A Total of 2 specimens have been observed in the field, preserved as negative epirelief. The total diameter of the structure is 24–26 cm, and protruding tentacle-like projections are 22 to 28 cm in length. No ripple or sedimentary features were observed on the bedding plane except for the microbial mat.
Palaeontological Significance: The present specimen is nicely preserved on the bedding plane. Microbial mat observed structure on the same bedding plane, which must have provided cohesiveness to get preserved within the fine-grained sandstone.
DISCUSSION
The present fossil assemblage, viz. Tirasiana qingzhenensis, Hiemalora stellaris, Pteridinium sp., Finkoella ukrainica, Finkoella oblonga, Arumberia banski, jellyfish-like impressions, coiled fossils, and one bizarre form offer critical insights into the palaeobiological and palaeoenvironmental dynamics of the Ediacaran Sonia Sandstone. The observed fossils represent a broad spectrum of morphological diversity, underscoring the ecological complexity of the Ediacaran period. For instance, Arumberia banski and Hiemalora stellaris illustrate the adaptability in varied Ediacaran depositional environments, ranging from shallow marine subtidal to intertidal settings (McMahon et al., 2022). The Ediacaran mat ground was pivotal in their preservation, highlighting the interplay between biological and sedimentary processes (Kumar & Pandey, 2008, 2009; McMahon et al., 2022). In the present study, the first occurrence of Tirasiana qingzhenensis from the Sonia Sandstone is particularly intriguing. It challenges the conventional understanding of the terminal Ediacaran mass extinction, as previously reported from the Cambrian strata in China. Similarly, the sack-like structures of Finkoella ukrainica and Finkoella oblonga imply potential affinities with early tunicates, expanding our knowledge of early metazoan evolution. Pteridinium sp. is well known in late Ediacaran strata of Namibia, South Australia, South Africa, the White Sea region, and for the first time from the Marwar Supergroup. However, a complete fossil specimen has yet to be discovered.
The fossil assemblage aligns closely with the Nama assemblage of the Ediacaran, reflecting similar ecological and depositional conditions characterised by siliciclastic sediments in coastal to shallow marine environments, unlike the Avalon and White Sea assemblages, which are predominantly deep-water biotas. The Sonia Sandstone captures the transitional environments of tidal flats, providing a unique perspective on Ediacaran life forms. The discoveries of new fossil assemblages from the Sonia Sandstone have broader implications for understanding the biogeographical distribution of Ediacaran biota and their role in the Cambrian explosion. The diverse suite of fossils reported here not only enriches the palaeontological record of the Marwar Supergroup but also establishes the Sonia Sandstone as a critical Ediacaran fossil lagerstätte, offering valuable insights into the evolutionary, ecological, and taphonomic processes of the late Precambrian.
CONCLUSIONS
The Sonia Sandstone of the Jodhpur Group has previously yielded significant Ediacaran fossil records, viz., Heimalora, Aspidella, Nimbia, Albumares, Parvancorina, five-armed body fossil, medusoid impressions (Marsonia artiyensis), trace fossils, and Microbially Induced Sedimentary Structures (MISS) and Ediacaran giant seaweed (see Pandey & Sharma, 2017 and references therein). These findings have provided foundational insights into the depositional history and palaeoecological conditions of the Marwar Supergroup. The present study enriched the existing fossil assemblage, comprising Tirasiana qingzhenensis, Hiemalora stellaris, Pteridinium sp., Finkoella ukrainica, Finkoella oblonga, Arumberia banski, coiled forms, bizarre forms A, Quaestio simpsonorum, and jellyfish-like impressions. Out of which, Quaestio simpsonorum, bizarre forms A, Tirasiana qingzhenensis, Hiemalora stellaris, Pteridinium sp., are being reported for the first time. Arumberia banski is reported from another locality and shows its temporal existence within the Sonia Sea, representing a characteristic foreshore depositional environment. In contrast, Hiemalora stellaris, jellyfish-like impressions, and other medusoid forms indicate deposition in calm, shallow marine settings, occasionally influenced by episodic sediment influx that facilitated rapid burial and exceptional preservation. The presence of Finkoella ukrainica and Finkoella oblonga is indicative of deposition in soft, fine-grained substrates typical of coastal shallow marine zones. At the same time, the bizarre forms and coiled impressions provide further evidence for fluctuating depositional conditions in a transitional environment between intertidal and subtidal regimes (Pandey & Sharma, 2017). When viewed in a broader palaeobiogeographical context, the overall fossil assemblage of the Sonia Sandstone displays strong affinities with the Nama (in specific) and White Sea assemblages (in rare instances) of the Ediacaran Period. Fossils such as Pteridinium sp. are diagnostic of shallow marine to intertidal environments, a hallmark of Nama assemblages. Similarly, the medusoids, including Hiemalora stellaris and jellyfish-like impressions, align with the low-energy depositional settings and episodic sedimentation that characterise the Nama biota. These findings reinforce the ecological and taphonomic parallels between the Sonia Sandstone and globally recognised Ediacaran assemblages, thereby underscoring its significance in understanding the evolution and distribution of early complex life.
Footnotes
Acknowledgements
The authors thank the Director of BSIP for providing facilities for the investigation and for permission to publish this work (BSIP/RDCC/85/2024-25). SA gratefully acknowledges the Council of Scientific and Industrial Research CSIR, New Delhi, for the Pool Scientist Fellowship in support of this research.
Declaration of Conflicting Interests
The authors declared no potential conflicts of interest with respect to the research, authorship and/or publication of this article.
Funding
The authors disclosed receipt of the following financial support for the research, authorship and/or publication of this article: This research was financially supported by the DST-BSIP, Lucknow and Council of Scientific and Industrial Research (CSIR), New Delhi (Senior Research Associate, Fellow ID 3510103).