
Abstract
The present work recognised a well-preserved suite of trace fossils including Catenichnus isp., Cruziana problematica, Cruziana tenella, Diplichnites isp., Monomorphichnus lineatus, Planolites montanus, Rusophycus isp., ?Rosselia socialis, serially repeated Rusophycus eutendorfensis intergrading Cruziana tenella, and ?Treptichnus cf. pedum from the Lower Cambrian Member-A of the Dhaulagiri Formation (Tal Group) in the Mussoorie syncline. Trace fossils from the Member-A of the Dhaulagiri Formation are grouped into the Cruziana ichnofacies, and trace makers mainly are epi- and endobenthic polychaete worms, trilobites and arthropods. Sedimentological studies indicate the presence of lenticular and flaser bedding, trough cross laminations, trough cross stratifications, and micro-conglomerate. Facies analysis shows that the trace fossils are preserved in a heterolithic succession of silty shale-siltstone—and minor sandstone facies of a mixed flat environment of a tidal flat setting. The shallow penetration depth, the small size of burrows (especially Rusophycus eutendorfensis intergrading Cruziana tenella), and low ichno-diversity are indicative of a stressed environment during the deposition of the Member-A of the Dhaulagiri Formation. Distribution of trace fossils concerning the Ediacaran-Cambrian boundary in the Mussoorie syncline indicates that so far traces are not recorded near the boundary interval, and known records are mostly from the Arenaceous Member (Deo-Ka-Tibba Formation), which is much above the Ediacaran-Cambrian boundary interval.
INTRODUCTION
Integrated sedimentological and ichnological studies provide identification of facies and the inter-relationship between organisms at the sediment-water interface and paleo-depositional affinities (e.g., Buatois & Mángano, 2011; Knaust, 2017; Pemberton et al., 1992). At the water-sediment interface and within the sediments, the activity of organisms is mainly controlled by depositional energy conditions, substrate consistencies, depositional rates, oxygenation, water turbidity and salinity (Bann et al., 2004, 2008; Buatois & Mángano, 2011; Buatois et al., 2005; Knaust, 2017; MacEachern et al., 2007; Pemberton et al., 1992). The Cambrian trace fossils, though widely reported from the Tal Group in the Mussoorie syncline (Lesser Himalaya) confined to documentation of ichnogenera and ichnospecies, and lack integrated sedimentological-ichnological studies, and their bearing on the Ediacaran-Cambrian boundary.
The present study provides a detailed sedimentological and ichnological characterisation of Member-A of the Dhaulagiri Formation (Tal Group) in the Mussoorie syncline, Lesser Himalaya, Uttarakhand. Our study documents sedimentary facies in the Member-A, ichnological assemblages and interprets their paleoenvironment. The present described ichnofossils are stratigraphically significant as they are from the Member-A of the Dhaulagiri Formation, which constitutes the lowermost part of the Dhaulagiri Formation.
GEOLOGICAL SETTING AND LITHOSTRATIGRAPHY
The Lesser Himalaya (LH) lies south of the Main Central Thrust (MCT) and north of the Main Boundary Thrust (MBT) (Figure 1A). In LH, the Cambrian rocks classified under the Tal Group (Auden, 1934; Bhargava et al., 1998; Kumar & Dhaundiyal, 1979; Middlemiss, 1887; Shanker et al., 1993; Singh et al., 2019) (Figure 1B) occur along the core of Nigalidhar, Korgai, Mussoorie, Garhwal and questionably in Nainital (Figure 1A) of the Inner Krol Belt (Bhargava, 1972).
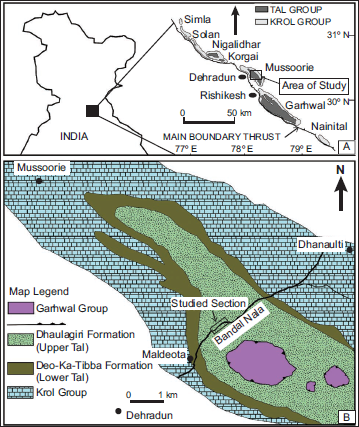
Lithostratigraphically, the Tal Group in the Mussoorie syncline is broadly divided into Deo-Ka-Tibba (Lower Tal) and Dhaulagiri (Upper Tal) formations based on a disconformity between them (Shanker et al., 1993). This classification has also been used in the Garhwal syncline (Uttarakhand) (Kumar & Dhaundiyal, 1979). However, in Nigalidhar and Korgai synclines (H.P.), the Tal Group is divided into three formations (Bhargava et al., 1998; Singh et al., 2019). Bhargava and Singh (2022) presented a correlation of various formations and members of the Tal Group from Nigalidhar to Nainital synclines and revealed that the Tal Group is thicker and more completely preserved in the Nigalidhar syncline. Bhargava and Singh (2022) presented the lithostratigraphic correlations of the Tal Group in Nigalidhar and Mussoorie synclines in Krol-Tal Belt, which are followed in the present work (Figure 2).
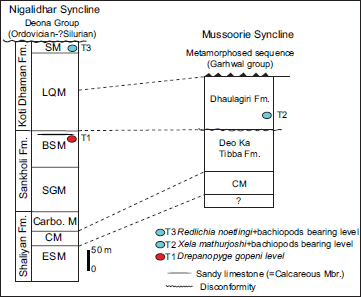
In the Mussoorie syncline, the Ediacaran-Cambrian boundary is proposed at the juncture of the Krol and Tal groups based on the negative carbon isotope excursion (Aharon et al., 1987). The small shelly fossils (SSFs) of Anabarites trisulcatus–Protohertzina anabarica zone of Fortunian age (Cambrian) occur in the Chert Member of the Deo-Ka-Tibba Formation (Tal Group) (Azmi et al., 1981; Bhatt et al., 1983; Brasier & Singh, 1987; Juyal, 1979). Trace fossil Treptichnus pedum, indicating the base of the Fortunian age (Cambrian), has yet to be discovered from the upper part of the Krol Group or from the lower part of the Tal Group. The Lower Cambrian trilobite Xela mathurjoshi is known from the grey-greenish shale of Dhaulagiri Formation (Jell & Hughes, 1997; Mathur & Joshi, 1989).
SECTION DETAIL AND SEDIMENTARY FACIES
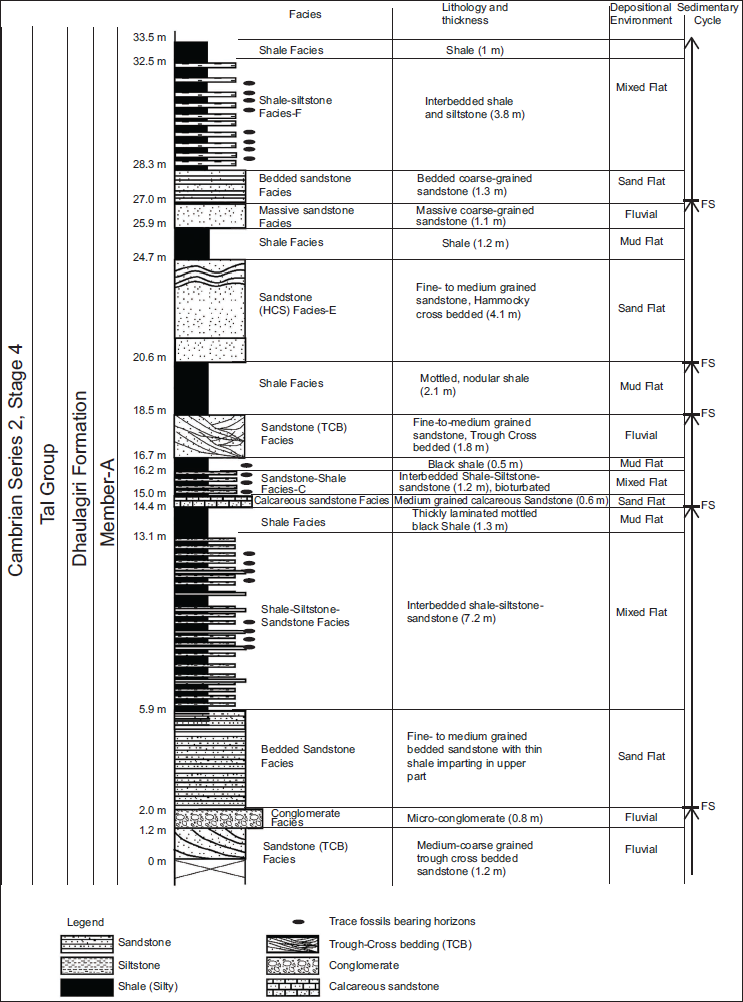
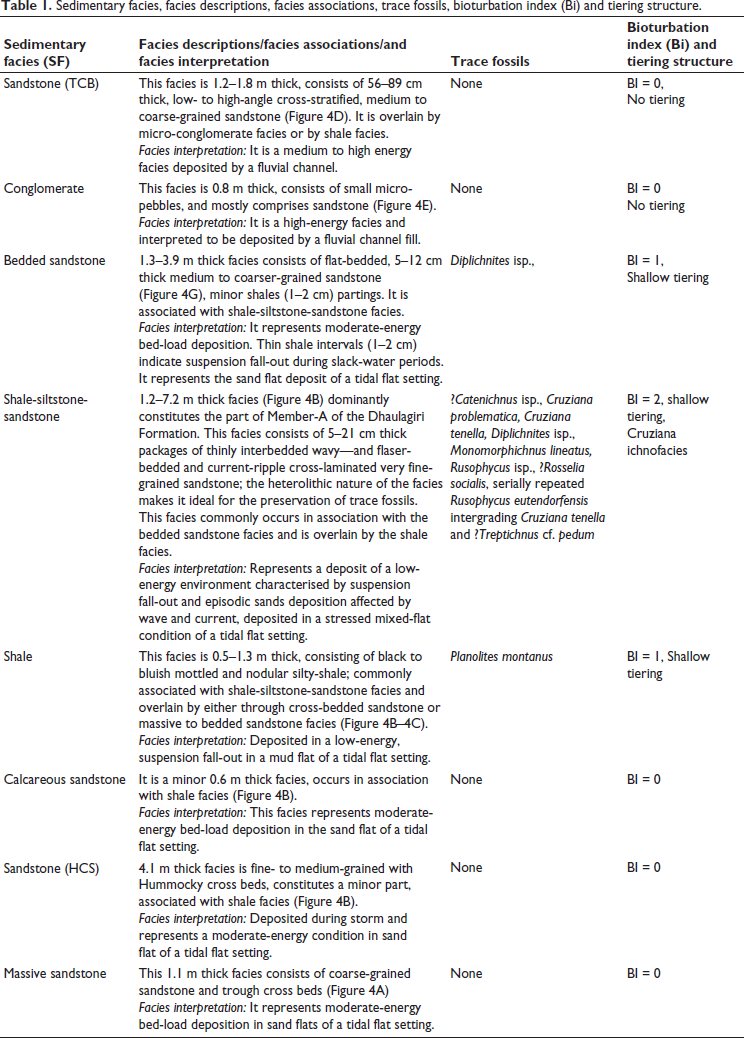
The studied section lies in the central part of the Mussoorie syncline along the Maldeota-Dhanaulti road (Figures 1 and 3). The 33.5 m thick section of the Member-A of the Dhaulagiri Formation displays interbedded trough cross-bedded (TCB) sandstone, massive to bedded sandstone, interbedded shale-siltstone-sandstone, medium- to coarse sandstone, thin to thick shale and calcareous sandstone. Eight lithofacies, that is, sandstone (TCB) facies, conglomerate facies, bedded sandstone facies, shale-siltstone-sandstone facies, shale facies, calcareous sandstone facies, sandstone (Hummocky Cross Bedded [HCB]) facies and massive sandstone facies, are mainly recognised (Figure 3 and Table 1). These facies are interpreted to represent mud flat, mixed flat and sand flat facies of the tidal flat setting and fluvial facies (Figure 3 and Table 1).
Characteristic lithology, thickness, sedimentary structures, facie, cycle, trace fossils and depositional environment of the Member-A of the Dhaulagiri Formation (Tal Group) Mussoorie syncline (Uttarakhand).
Sedimentary facies, facies descriptions, facies associations, trace fossils, bioturbation index (Bi) and tiering structure.
SYSTEMATIC ICHNOLOGY
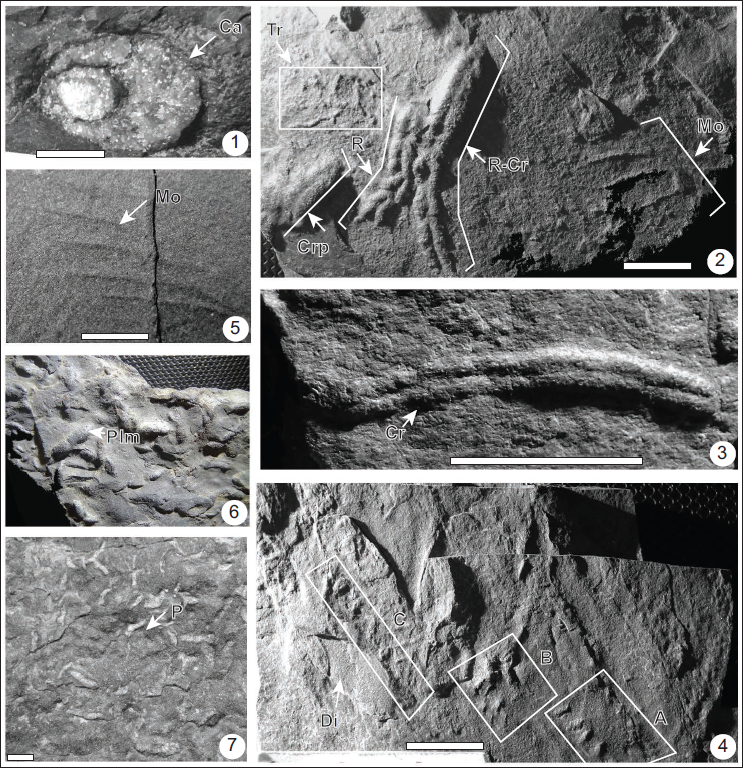
Catenichnus ?isp.
(Plate 1.1)
Material: One fine-grained sandstone slab containing two specimens.
Description: Vertical to slightly inclined cylindrical burrow opening comprises two elliptical eccentric rings, diameter is 13.7 mm; central shaft contains filling material (sandstone) different from the host rock (Plate 1.1).
Remarks: Due to poor preservation, this specimen is identified only at ichnogeneric level. Both the rings of the burrow opening are slightly eccentric, which may be due to the inclined nature of the U-shaped tube. However, the other opening is not visible, which constrains us to identify this as Catenichnus confidently.
Cruziana problematica Schindewolf, 1928
(Plate 1.2 (Crp))
Material: One fine-grained sandstone slab contains Cruziana problematica (Crp.), serially repeated Rusophycus and integrated Rusophycus and Cruziana, ?Treptichnus pedum and Monomorphichnus lineatus.
Description: Straight, bilobed, ribbon-like burrows with a central furrow and faint transverse striations; length is 18.2 mm and width 5.6 mm, and the central groove is 1 mm deep. The structure is present as a positive hyporelief on the sole of the bed.
Remarks: The present specimen is classified under Cruziana problematica (Mángano et al., 2002). Jensen (1997) considered it a junior synonym of Cruziana tenella. The presence of transverse scratch marks confirms its designation as Cruziana problematica (Schlirf et al., 2001). Cruziana problematica is generally attributed to vagile small arthropods and vermiform deposit feeders (Mángano et al., 2002).
(Plate 1.3)
Material: One medium-grained sandstone slab with one specimen.
Description: Small, bilobed, straight to gently winding burrow, with rare transverse striations, 23.5 mm long and 2.1 mm wide, longitudinally divided by a central shallow to deep non-continuous groove. The burrow is preserved as a positive relief on the sole of a bed (Plate 1.3).
Remarks: Our specimen shows similarities to Cruziana tenella, described by Jensen (1997) and Kesidis et al. (2019), in its small size and rare transverse striations. Fillion and Pickerill (1990) described a similar trace under the Cruziana problematica. Jensen (1997) interpreted C. problematica as a junior synonym of C. tenella. The trace fossil Cruziana is discriminated from other, similar traces, for example, ichnospecies of Didymaulichnus (Young, 1972) by the presence of striation on the lobes (Keighley & Pickerill, 1996). The discussed specimen is, however, too narrow and has a low relief to be considered as Didymaulichnus. The undulations in the horizontal plane suggest the foraging behaviour of the producers (e.g., Gaillard et al., 2005).
(Plate 2.1 (m-o))
Material: One fine-grained sandstone slab contains three poorly preserved specimens of Cruziana isp. and four specimens of Rusophycus isp., preserved as a positive hyporelief.
Description: Straight, bilobed, ribbon-like burrows with a central furrow (Plate 2.1 m-o); length varies from 11.5 to 19.6 mm, and width from 4.2 to 9.6 mm, showing L/W ratio 2.04–2.86.
Remarks: Cruziana is considered a result of combined locomotion and feeding strategy (Kesidis et al., 2019; Seilacher, 2007), and it occurs in shallow water marine to freshwater settings mostly in Palaeozoic (Bromley & Asgaard, 1979).
Diplichnites isp.
(Plate 1.4)
Material: One silty-sandstone slab contains three distinctive sets (A to C) of paired to unpaired, small parallel to sub-parallel rows of symmetrical to asymmetrical, crescent-shaped to spindle-shaped elongated ridges.
Description: The Set A (Plate 1.4) is 13.5 mm in length and 8.9 mm in width; it comprises paired rows of six small ridges or imprints preserved as hyporelief and separated by a distance of 4.3 mm. Individual imprints in a row are 1.6 mm apart. The set B (Plate 1.4) is 7.4 mm long, 6.2 mm wide, and comprises paired rows of five imprints preserved as positive hyporelief and separated by a distance of 1.6 mm. In another row, the individual imprints are 0.9 mm apart. The set C (Plate 1.4) is unpaired and comprises 15.6 mm long row of five imprints separated from each other by a distance of 2.8 mm. In each set, the ridges are oriented normal or slightly oblique to the axis of the trackway.
Remarks: Diplichnites has been attributed to locomotion trails of trilobites (Seilacher, 1955). Diplichnites occurs widely from the lower Cambrian to the Permian (Crimes et al., 1982; Häntzschel, 1975). Diplichnites from the Palaeozoic are attributed to arthropods, whereas Mesozoic occurrences are associated with benthic crustaceans (Häntzschel, 1975; Weber et al., 2007).
Monomorphichnus lineatus Crimes, Legg, Marcos and Arboleya, 1977
(Plate 1.2 (Mo), 1.5)
Material: Two fine-grained sandstone samples; one sample contains other traces of serially repeated Rusophycus and intergrading Cruziana, Treptichnus pedum and Cruziana problematica.
Description: The set includes individual straight to slightly curved ridges arranged parallel to each other, but each shifted longitudinally from the previous marking. Ridges range from 5.8 mm to 11.6 mm in length with an average width of 1.2 mm (Plate 1.2 (Mo). The distance between the ridges varies from 10 mm to 11.2 mm. The structure is preserved as positive hyporelief (Plate 1.2 (Mo). Another set (Plate 1.5) comprises five straight to slightly curved and elongated ridges ranging from 8.7 mm to 30.5 mm; average width ranges from 0.5 mm to 1.2 mm; the central ridge is thickest 30.5 mm, ridges are separated by a distance 2.0 mm to 3.5 mm, preserved as hyporelief.
Remarks: The presence of a single series of ridges classified these under Monomorphichnus lineatus. M. lineatus (Crimes et al., 1977) differs from M. bilineairs (Crimes, 1970) by lack of paired ridges and from M. multilineatus (Alpert, 1976) in that central ridges are deeper than the outer ridges (Fillion & Pickerill, 1990; Gibbs et al., 2016). Initially, Monomorphichnus was thought to be produced by the endopodite claws of trilobites (Crime, 1970; Martinsson, 1965), but Keighley and Pickerill (1996) reported M. cf. lineatus from the Carboniferous non-marine deposits of eastern Canada that demonstrated that it can be produced by non-marine crustaceans also. Monomorphichnus is interpreted as a trace fossil likely produced by trilobites and other arthropods engaged in swimming or grazing activities (Crimes, 1970, 1987; Mikuláš, 1995).
Planolites montanus (Richter, 1937)
(Plate 1.6–1.7)
Material: Two fine-grained silty-shale and silty-sandstone samples with dozens of specimens.
Description: Small, smooth, straight to slightly sinuous, horizontal to slightly inclined, with coarse filling burrows preserved as full relief (Plate 1.6, 1.7). These vary in length from 6.4 mm to 14.2 mm and in width from 2.3 mm to 4.9 mm. These burrows are commonly found in bundles, with individual burrows exhibiting random orientations. Transverse annuli are preserved in some burrows. Another specimen (Plate 1.7) is cylindrical to sub-cylindrical, horizontal, unlined burrows having an irregular surface, burrow fill is coarser than the host sediment. Burrows’ width varies from 1.8 to 2.7 mm, and length from 6.3 to 13.6 mm.
Remarks: Planolites montanus has a tortuous course consisting of both horizontal and inclined segments, and notably lacks ornamentation. Planolites is interpreted as a feeding structure formed by the vagile infaunal vermiform organism (Buatois & Mángano, 1993; Fillion & Pickerill, 1990; Frey, 1970; Hofmann et al., 2012; Howards & Frey, 1975; Nicholson, 1873; Pemberton & Frey, 1982).
Rusophycus isp.
(Plate 2.1)
Material: One fine-grained sandstone slab contains three poorly preserved specimens of Cruziana isp. and four specimens of Rusophycus isp., preserved as a positive hyporelief.
Description: Rusophycus as small coffee bean-shaped lobes (Plate 2.1 (a-d) varies in length from 8.0 mm to 14.6 mm and in width from 6.9 mm to 8.4 mm, all showing L/W ratio range from 1.15 to 1.73. The lobes taper towards the posterior side. The median furrow between the lobes is clearly visible in some burrows.
Remarks: Rusophycus differs from Cruziana in its L/W ratio. This ratio is <2 in Rusophycus and >2 in Cruziana (Keighley & Pickerill, 1996). Further species-level identification is not possible as the surface markings are not well preserved. These are similar to Rusophycus dispar (Linnarsson, 1869) in having wider posterior ends. The traverse scratches are not decipherable due to poor preservation. Rusophycus is interpreted as a resting trilobite trace (Crimes, 1975; Häntzschel, 1975).
?Rosselia socialis (Dahmer, 1937)
(Plate 2.2)
Material: One fine-grained silty-sandstone slab contains one specimen of ?Rosselia socialis.
Description: Concentric circles with a central opening are preserved as concave epirelief on bedding plane, 25.4 mm in diameter (Plate 2.2), and circles are composed of finer-grained sandy and clayey-silty material.
Remarks: The upper transverse section is visible only; the lack of a vertical cross-section limits its confident identification.
Treptichnus pedum (Seilacher, 1955)
(Plate 2.3)
Material: One fine-grained sandstone slab contains Cruziana problematica (Crp.), serially repeated Rusophycus and integrated Rusophycus and Cruziana, Treptichnus pedum and Monomorphichnus lineatus traces.
Description: Horizontal, row of smooth, straight to slightly curved short 5–6 segments connected at low angles forming a zigzag pattern; probes are added inward of the previous segment and connect to neighbouring segment by approximately 45°. The overall length of the trace is 13 mm, and the average width is 0.8 mm (Plate 2.3).
Remarks: The first appearance datum of Treptichnus pedum in association with small shelly fauna (SSFs) is used to demarcate the Ediacaran-Cambrian boundary (Buatois & Mángano, 2011; Geyer & Landing, 2016; Narbonne et al., 1987). In the Indian Himalaya, the broad significance of Treptichnus pedum was discussed by Singh et al. (2017), who concluded that all the reports so far from the Himalaya indicate a younger age rather than the Ediacaran-Cambrian boundary interval. Treptichnus is considered to be formed by the activity of priapulid worms (Vannier et al., 2010), and it shows feeding behaviour (cf. Buatois et al., 1998; Crimes et al., 1977; Jensen, 1997; Seilacher & Hemleben, 1966). The present discussed specimen is very small but preserves characters confidently grouped under the Treptichnus pedum.
(Plate 1.2 (R-Cr, R))
Material: One fine-grained sandstone slab contains Cruziana problematica (Crp.), serially repeated Rusophycus eutendorfensis and integrated Rusophycus eutendorfensis and Cruziana tenella, Treptichnus pedum and Monomorphichnus lineatus traces.
Description: The sample contains two structures showing the behaviour of trace maker, that is, (a) structure comprises serially repeated six to seven Rusophycus eutendorfensis (Plate 1.2(R) and, (b) five serially repeated Rusophycus eutendorfensis intergrading to Cruziana tenella (Plate 1.2 (R-Cr). The serially repeated structure comprises six to seven Rusophycus eutendorfensis (Plate 1.2 (R) is 22.6 mm in length and 4.0 mm in width; the length of individual Rusophycus eutendorfensis varies from 2.6 mm to 5.7 mm, and width from 2.7 mm to 4.0 mm. The anterior specimen usually overlaps with the successive ones. The surface of Rusophycus eutendorfensis is smooth. The second structure (Plate 1.2 (R-Cr)) comprises five Rusophycus eutendorfensis arranged in a series and laterally intergrading into Cruziana tenella, creating a transitional spectrum. The structure is 40 mm in length and 4.3 mm in width; the length and width of individual Rusophycus eutendorfensis vary from 2.8 to 6.0 mm and 3.7 to 4.3 mm, respectively. Cruziana tenella measures 16.5 mm in length and 4.3 mm in width. The surfaces of both Rusophycus eutendorfensis and the intergrading Cruziana tenella are smooth.
Remarks: Kesidis et al. (2019) and Brandt (2007) discussed the serially repeated/or multiple Rusophycus and intergrading behaviour of Rusophycus into Cruziana and attributed this behaviour to (a) as detritus-feeding (Cruziana) alternating with forays (Rusophycus) for prey (Brandt, 2007) and (b) energy-efficient sediment processing at a resource-rich location (Kesidis et al., 2019). Orlowski et al. (1971) attributed integration to the mating activity. According to Brandt (2007), the serially repeated Rusophycus suggests a low sedimentation rate, as the trace maker requires lifting off the substrate to move forward or backwards in calm water conditions. It is generally understood that serial repetition of burrows of similar size mainly reflects the activity of a single individual. Kesidis et al. (2019) suggested an epibenthic lifestyle, and the low height of Rusophycus and Cruziana indicates a search for food rather than a resting phase.
STRATIGRAPHIC DISTRIBUTION OF TRACE FOSSILS CONCERNING EDIACARAN-CAMBRIAN BOUNDARY IN MUSSOORIE SYNCLINE
Although, a significant number of trace fossils are known from the Tal Group of rocks in Mussoorie syncline (Banerjee & Narain, 1976; Joshi & Tiwari, 2014; Rai, 1987; Rai & Singh, 1983; Singh, 2011; Singh & Rai, 1983; Singh et al., 2014; Tiwari & Parcha, 2006; Tiwari et al., 2013) but so far these trace fossils have not been analysed in term of Ediacaran-Cambrian boundary in the Mussoorie syncline. Moreover, most of the recorded trace fossils from the Mussoorie syncline are subject to revision (Singh et al., in review). The confirmed ichnogenera are presently considered for their utility in the evaluation of metazoan life after the Ediacaran-Cambrian boundary in the Tal Group of the Mussoorie syncline (Lesser Himalaya).
In the Mussoorie syncline, the Ediacaran-Cambrian boundary is expected at the basal Chert-Phosphorite Member of the Deo-Ka-Tibba Formation (See Braiser & Singh, 1987). Body fossils of brachiopods and trilobites of Stage 4 are known from the Dhaulagiri Formation. Body fossils have yet to be discovered from the Deo-Ka-Tibba Formation.
Various authors presented a view of close relationship between the western part of the Yangtze block (South China) and the Himalayan margin early in the Cambrian period and this assumption is mainly based on similarities in litho- and biostratigraphic framework of these areas (Braiser & Singh, 1987; Hughes et al., 2005; Jiang et al., 2002). Herein, we present comparative studies of the distribution of trace fossils in the Tal Group of the Mussoorie syncline (Lesser Himalaya) and equivalent rocks of the Yangtze block (South China), and evaluate their significance in terms of metazoan life after the Ediacaran-Cambrian boundary in the Lesser Himalaya.
Text Table 2 shows the distribution of trace fossils from the various formations in early Cambrian of South China (after Weber et al., 2007; Zhu, 1997) and so far recorded confirmed trace fossils from the Tal Group of the Mussoorie syncline (Joshi & Tiwari, 2014; Rai, 1987; Singh, 2011; Singh & Rai, 1993; Singh et al., 2014; Tiwari & Parcha, 2006; Tiwari et al., 2013, and Present work).
Comparative account of trace fossil records in lower Cambrian of Yangtze block (South China) and Mussoorie syncline (Lesser Himalaya, India).
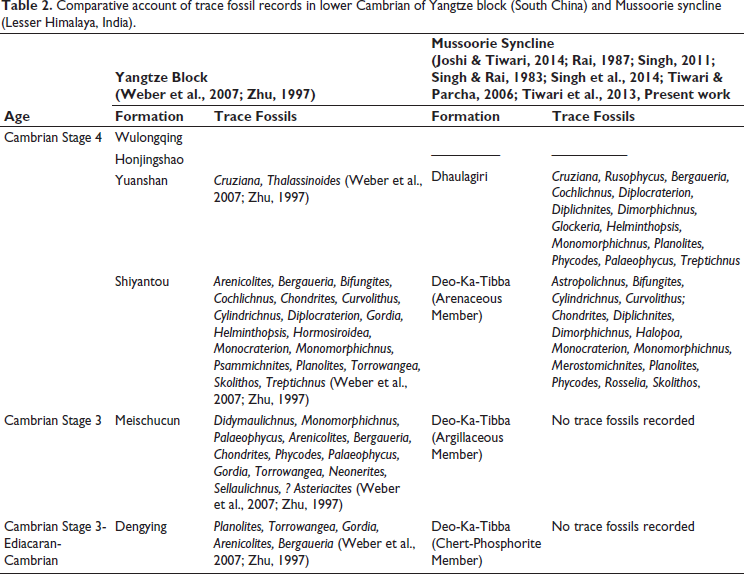
The Chert-Phosphorite Member (Deo-Ka-Tibba Formation) in Mussoorie syncline yielded SSFs of Anabarites trisulcatus-Protohertzina anabarica assemblage zone of Fortunian age (Braiser & Singh, 1987). The lithostratigraphic and biostratigraphic correlations suggest the Chert-Phosphorite Member is equivalent to the Dengying Formation in Yunan, South China, which also contains the Anabarites trisulcatus-Protohertzina anabarica assemblage zone (Chen et al., 1995; Hughes et al., 2005; Weber et al., 2007; Zhu, 1997). However, recently, the limestone of the Dengying Formation yielded the oldest known Cambrian brachiopod–trilobite association, specifically the trilobite Parabadiella cf. huoi and the brachiopod Eoobolus incipiens, both of Cambrian Age 3, in South China (Zhang et al., 2021). Zhang et al. (2021) placed the Xihaoping Member (from which the trilobite Parabadiella cf. huoi and brachiopod Eoobolus incipiens are reported) under the Dengying Formation, however, Figure 4 (Zhang et al., 2021) correlated Xihaoping Member with the basal part of the Yuanshan Formation. Suppose the placement of Xihaoping Member under Dengying Formation is valid. In that case, the correlation of Anabarites trisulcatus-Protohertzina anabarica assemblage zone of Chert-Phosphorite Member with the Dengying Formation is questionable. It may also be possible that Zhang et al. (2021) mistakenly placed it under Dengying Formation or that the Dengying Formation ranges in age from the Fortunian to Stage 3.
Sedimentary structures and facies recorded in the Member-A of the Dhaulagiri Formation (Mussoorie synclines), (A) trough cross bedded sandstone facies; (B) shale-siltstone-sandstone facies with lenticular and wavy bedding (f) and (C) trace fossils yielding horizons (TF); (D) trough cross bedded sandstone facies (TCB); (E) micro-conglomerate facies; (F) Hummocky cross-bedded (HCB) sandstone facies; (G) bedded sandstone facies. The length of the hammer head is 17 cm.

Moreover, the Dengying Formation yielded trace fossils such as Planolites, Torrowangea, Gordia, Arenicolites and Bergaueria (Weber et al., 2007; Zhu, 1997). However, to date, the Chert-Phosphorite Member of the Deo-Ka-Tibba Formation has not yielded any trace fossils.
The overlying Argillaceous Member of the Deo-Ka-Tibba Formation in the Mussoorie syncline has not yielded body or trace fossils. The equivalent rocks, that is, Meischucun Formation in Yunnan (South China), yielded abundant trace fossils (see Table 2) but lack traces of Cruziana and Rusophycus.
Hughes et al. (2005) correlated the uppermost part of the Deo-Ka-Tibba Formation with the uppermost part of the Sankholi Formation of Nigalidhar syncline (Lesser Himalaya). This level yielded a trilobite of Drepanopyge gopeni (516-517 Ma) in the Nigalidhar syncline (=Drepanuorides zone of the Honjingshao Formation of Yunnan, South China) (Bhargava et al., 1998; Hughes et al., 2005). However, no such trilobites are known from the Mussoorie syncline. Some impressions of trilobites are recorded from the uppermost part of the Arenaceous Member (Deo-Ka-Tibba Formation) in the Mussoorie syncline; however, they are not identifiable to the generic level due to poor preservation (Rai & Singh, 1983).
We differ with this view that the uppermost part of the Deo-Ka-Tibba Formation is equivalent to the uppermost part of the Sankholi Formation (Nigalidhar syncline). As in the Mussoorie syncline, a disconformity is well identified between the Deo-Ka-Tibba and overlying Dhaulagiri formations (Singh, 1979). We suggest herein that much of the upper part of the Deo-Ka-Tibba Formation either eroded or was non-deposited before the disconformity (see Bhargava & Singh, 2022). This is validated with the trace fossils herein. We correlate the Arenaceous Member (Deo-Ka-Tibba Formation) with the Shiyantou Formation (Yunan, South China). These formations share diverse and typical traces like Psammichnites gigas, Diplichnites, Dimorphichnus, and Monomorphichnus, which indicate activity by large mollusks and trilobite/similar arthropods, respectively (Banerjee & Narain, 1976; Rai, 1987; Singh, 2011; Singh & Rai, 1983; Weber et al., 2007; Zhu, 1997).
In the Mussoorie syncline, the definite traces of Rusophycus and Cruziana are only known from the Dhaulagiri Formation (Singh et al., 2014; and present work), which shows the correlation with the first appearance of traces of Rusophycus and Cruziana in Yuanshan Formation in Yunnan (South China), where they occur below the lowest trilobite in Yuanshan Formation (Weber et al., 2007; Zhu, 1997). The Yuanshan Formation contains trilobites of Parabadiella and Eoredlichia-Wutingaspis zones (Zhang et al., 2021). These trilobites have not been reported from the Lesser Himalaya. Besides Cruziana and Rusophycus, the Dhaulagiri Formation richly contains other trace fossils (Table 2). We correlate the Dhaulagiri Formation (Mussoorie) with the entire Yuanshan and up to the middle part of the Honjingshao formations. This is validated by the known occurrence of trilobite Xela mathurjoshi in the Mussoorie syncline. The only trilobite known from the Cambrian of Mussoorie syncline is Xela mathurjoshi, which is known from the upper part of the Dhaulagiri Formation (see Jell & Hughes, 1997; previously described as Redlichia noetlingi by Mathur & Joshi, 1989). Hughes et al. (2005) correlated this occurrence with the Redlichia noetlingi in Nigalidhar syncline and stated that it is part of Palaeolenus lantenoisi zone of South China. We disagree with this assignment. The genus Xela (Jell, in Bengtson et al., 1990) is known from the Lower Cambrian Pararaiajaneae zone of South Australia, which is equivalent to Yunnanaspis-Yiliangella to Drepanuroides zones of the Honjingshao Formation in South China. The Palaeolenus zone of South China occurs in Wulongqing Formation (see Zhang et al., 2021). The rocks equivalent to the Wulongqing Formation are not recorded in the Mussoorie syncline (Table 2). We correlate the occurrence of Xela mathurjoshi in the Dhaulagiri Formation equivalent to a level in the Lower Quartzite Member of the Koti Dhaman Formation, where recently well-preserved trilobites of Drepanuroides zones are recorded (Singh et al., in preparation).
Trace fossils from the Member-A of the Dhaulagiri Formation (Tal Group) of the Mussoorie syncline. (1): Catenichnus isp. (ca), (2): Cruziana problematica (Crp), CAS/M/2003, serially repeated Rusophycus eutendorfensis (R) and intergrading Rusophycus eutendorfensis and Cruziana tenella (R-Cr), and Monomorphichnus lineatus (Mo); and Treptichnus pedum (Tr); (3): Cruziana tenella (Cr); CAS/M/2043 (4): Diplichnites isp. (Di), CAS/M/2023, A-C, (5): Monomorphichnus lineatus (Mo); CAS/M/2033; (6): Planolites montanus (Plm), CAS/M/2013;(7): CAS/M/2001. All scale bars = 1 cm.
Trace fossils from the basal part of Member-A, Dhaulagiri Formation (Tal Group) of the Mussoorie syncline. (1): Preferentially oriented Rusophycus isp. (A–D) and Cruziana (M-O), CAS/M/2008; (2): Rosselia socialis (Ro), CAS/M/2011, (3): ?Treptichnus cf. pedum (Tr), CAS/M/2003. All scale bars = 1 cm.

CONCLUSIONS
Thus, the distribution of so far recorded trace fossils in the Tal Group of the Mussoorie syncline shows that the trace fossils belong to Cambrian Stage 4, and none of them are indicative of the Ediacaran-Cambrian boundary. A thorough search is required in the lower part of the Tal Group to identify the trace fossils from Chert-Phosphorites and Argillaceous members. The traces Rusophycus and Cruziana occur higher in the stratigraphy, and these occurrences are consistent with the global occurrences.
Footnotes
Declaration of Conflicting Interests
The authors declared no potential conflicts of interest with respect to the research, authorship and/or publication of this article.
Funding
The authors disclosed receipt of the following financial support for the research, authorship and/or publication of this article: This work was supported by the Paleontological Society International Research Program (PalSIRP), USA [Sepkoski Grant, 2021] to BPS; the Indian National Science Academy (INSA) [INSA/SP/SS/2022/482] to ONB; and the National Testing Agency (NTA), Ministry of Human Resource Development (MHRD), Government of India [Junior Research Fellowship, NTA Ref. No. 191620104020] to DK.