
Abstract
Deciphering past vegetation composition can be upgraded by examining modern pollen rain and extant vegetation. Here, we compare pollen dataset of 15 surface soil samples from the forested area and open-land and crop-land areas of the Kukrail Reserve Forest, to understand the association between the modern pollen dispersal and the vegetation cover in the Central Ganga Plain. The palynoassemblage recovered from the surface soil displays the tropical moist and dry deciduous woodland comprising Syzygium, Butea, Holoptelea, Terminalia, Emblica, Sapotaceae and Meliaceae indicating warm and humid climate in response to rainfall heterogeneity in the study site. Extensive pollen morphological studies have also been conducted on 15 selected plant taxa for generating the modern pollen flora from this region, which could aid in the accurate identification of fossil pollen. It has been observed that a significant correlation can be seen between the local herbaceous vegetation and pollen data. The excessive incidence of anthropogenic marker pollen (cereal, average value: 17.26%) alongside with cultural pollen taxa like Brassica, Coriandrum and Solanaceae demarcate agricultural impact around the region. The multivariate statistical analysis like box-plot and principal component analysis signifies a sizable dissimilitude and similarity in pollen spectra from different depositional sites. Careful interpretation of past vegetation extent using fossil pollen assemblages is required, especially when the vegetation is treeless (open-land and crop-land areas) due to the fact that a large potential of pollen-source region and subsequently a higher percentage of long-distance anemophilous pollen have been recorded. Therefore, this study has been initiated to analyse recent pollen dispersal and preservation in various depositional settings of tropical reserve forest. Moreover, the account on cultural and anthropogenic pollen indicator could provide valuable insights into the extent of agricultural practices and the degradation of pristine forests and surrounding areas. Thus, the present pollen data would aid in accurately understanding the palaeoecological changes in the tropical forests and surrounding regions of Central Ganga Plain.
INTRODUCTION
Paleoecologists working with sediment data in unexplored areas face the challenge of decoding novel fossil pollen assemblages (Bradley, 1985; Tripathi & Pandey, 2023). The relationship between vegetation and pollen assemblages is now not linear due to variations in pollen production, dispersal and deposition owing to the over-representation of some pollen taxa, whilst others are under-represented or absent (Cushing, 1967; Prentice, 1985; Moore et al., 1991; Sugita, 1993; Jackson & Lyford, 1999). Surface soils, sediments and different pollen traps such as moss polsters, spider webs and honey comprise pollen and spores that reflect the extant vegetation of a vicinity and help in the interpretation of fossil pollen records (Wright, 1967; Flenley, 1973; Moore, 1978; Liu & Lam, 1985; Fall, 1992; Birks & Birks, 2000).
Over the previous two decades, however, growing zeal in palaeoecological reconstruction of past tropical environments has led to the establishment of modern pollen analogues from the tropical region (Tripathi et al., 2023; Tripathi & Pandey, 2023; Prasad & Quamar, 2023). The Central Ganga Plain (CGP) is one of the most populated areas of the world due to favourable monsoonal conditions with high diversity in vegetational succession that supports the fertile soil, availability of water and local weather (Singh & Awasthi, 2011).They are the important physiographic features of the Indian subcontinent having monsoon-dependent agrarian economy. The Ganga Plain is one of the most substantially developed fluvial landscapes of the world and constitutes a primary phase of the Himalayan Foreland Basin with several lakes, ponds (commonly referred as ‘Taal’) and sedimentary deposits in the interfluve location for the palaeoclimatic research during the Late Quaternary (Singh, 1996; Singh et al., 1999; Srivastava et al., 2003).
Tripathi et al. (2016), Saxena et al. (2017) and Ranjan & Tripathi (2024) conducted research on modern pollen deposition in and around Lucknow and adjoining areas. In the present communication, our study would elaborate and provide an insight into the potential of generating pollen flora and formulating surface palynological studies exclusively from the forest-land and open-land and crop-land areas of Kukrail Reserve Forest, Lucknow. This database will help in understanding any variations recorded in pollen deposition vis-à-vis surrounding vegetation (Figure 1). Moreover, the modern pollen spectra generated would definitely help in the reconstruction of past environment and vegetation shifts in the CGP.
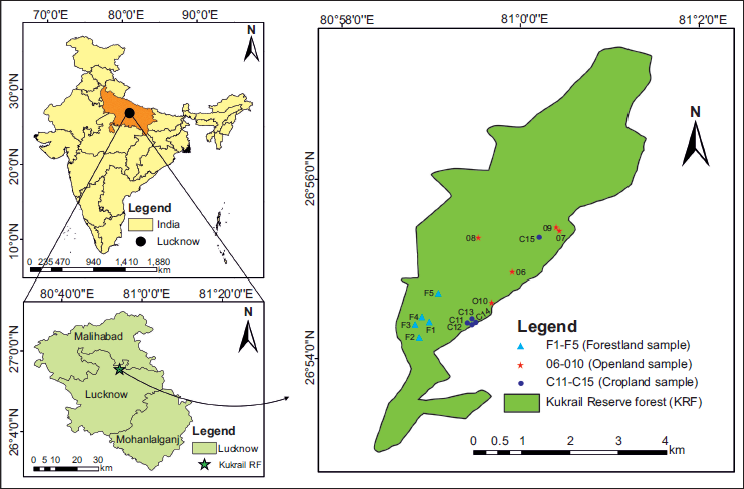
Location map showing study areas.
Pollen morphological analysis is one of the most potential biotic proxy to reconstruct the palaeoenviromental conditions and alteration in climatic condition of any particular region (Bradley, 1985) and helps in ascertaining the extent of propinquity and analogy among taxonomically related plant taxa (Walker & Doyle, 1975; Doyle, 2005; Garg et al., 2021; Goncalves-Esteves et al., 2022). Additionally the past anthropogenic and arboricultural practices could also be traced by pollen morphological investigation of cultivated and arboreal taxa of a region, respectively (Tripathi et al., 2021). In India, extensive pollen morphological research has been conducted in the North-West Himalayas and Maharashtra (Gupta & Sharma, 1986; Nayar, 1990). Nevertheless, there have been some publications on the pollen morphology of tropical regions of India over time (Chauhan & Bera, 1990; Tripathi et al., 2017, 2019, 2022; Quamar et al., 2017, 2022) to enrich the pollen identification reference from different regions of India. Thus, the objectives of the present study are (a) to establish a modern pollen analogue from 15 surface sediment samples collected in a transect from different depositional settings (forested, open-land and crop-land) of Kukrail Reserve Forest, Lucknow District (Figure 1), and (b) to investigate pollen morphological characters of 15 important plant taxa of the Lucknow District using light microscopy (LM) alongside with respective phenological information as a background information in studying modern pollen rain.
The study includes the palynological interpretation of aforesaid surface soil samples to observe the relationship between the modern pollen assemblages and current vegetation patterns in the CGP. To attain this objective, the multivariate principal component analysis (PCA) and box plot were also applied to the pollen frequency dataset to analyse the principal pollen taxa representing the different depositional sites. Furthermore, the generated modern pollen analogue could aid in deciphering the palaeoclimate and past vegetation changes in the CGP.
The study area is frequently affected by the anthropogenic activities such as buildings and roads near the reserve forest areas along with the expansion of crop-land at the outskirt, leading to soil erosion and habitat loss for economically and medicinally important plant species. As a consequence, natural vegetation has been replaced by rolling grasslands. Therefore, it is essential to consider pollen morphological and phenological data, particularly the flowering period, as crucial factors in sustaining the scattered vegetation in and around the reserve forest of Lucknow city. The modern pollen analogue in the forests, grasslands and human-impacted areas aids in the interpretation of the past ecology of that region (Huntley, 1993) and also helps in solving the important issues regarding pollen productivity, dispersal and preservation studies (Calcote, 1995, Li et al., 2009; Tripathi et al., 2016; Zhang et al., 2017).
STUDY SITE
Lucknow is the capital city of Uttar Pradesh which is geographically situated between 26.30°–27.10° North and 80.30°–81.13° East with an elevation of ~123 m above sea level (Figure 1). The city is located on the Indo-Gangetic foreland basin which is traversed by three mighty rivers, that is the Indus, the Ganga and the Brahmaputra. The CGP represents the middle part of the Indo-Gangetic foreland basin and mainly comprises sandy silty sediments (Goswami & Yhokha, 2010). It exhibits a very low relief and gentle regional slope and the present-day geomorphology of the region is a result of the complex interaction between climate changes associated with base level changes and tectonic activity during Late Pleistocene-Holocene (Srivastava et al., 2003). The Ganga Plain, one of the largest alluvial tracts of the world, abounds with numerous potential lakes and sedimentary deposits for the Quaternary palaeoclimatic studies (Singh, 1996; Srivastava et al., 2003). The modern pollen analogue study was conducted in and around the Kukrail Reserve Forest of Lucknow, the capital city of Uttar Pradesh, India. The sampling sites are located between 26°54′–26°56′ N latitudes and 80°58′–81°35′ E longitudes using the Global Positioning System (Figure 1). A large area of Uttar Pradesh is covered by a deep layer of alluvium spread by the slow-moving rivers of the Ganges system which is extremely fertile ranging from sandy to clayey loam. The soils in the southern part of the state are generally mixed red and black or red-to-yellow.
Kukrail Reserve Forest is located in Khurram Nagar/Indranagar adjoining Shivpuri colony on a picnic spot road at a distance of 9 km from the centre of the city (Figure 2 (1)). The present modern pollen exploration was performed in and around tropical deciduous forest of Kukrail Reserve Forest (Figure 1), for assessing the relationship between modern pollen rain and extant vegetation.
Field Photographs in Kukrail reserve forest (RF), Lucknow district. 1–3. View of tropical dry deciduous forest inside Kukrail RF; 4. Swamp inside Kukrail RF; 5. Collection of surface soil in the forest area of Kukrail RF; 6. The luxuriant growth of shrubby and herbaceous taxa at the outskirts of Kukrail RF.

CLIMATE AND VEGETATION
The climate of Uttar Pradesh is the tropical monsoon type, with warm weather year-round. Average high temperatures in Lucknow range from about 70°F (20°C) in January to over 100°F (38°C) in May and June. High temperatures of about 120°F (50°C) have been recorded at Gonda, northwest of Faizabad. As the state is a landlocked system, moisture-laden Indian Summer Monsoon (ISM) reaches here during the months of June to September. The ISM is responsible for ~75% of precipitation during these four months. An average of 50 years of meteorological data shows that Lucknow receives an average rainfall of ~1010 mm/year (IMD, Amausi station, Lucknow). There is a huge difference in the summer and winter temperatures as summer temperatures can easily go up to 47°C associated with intense heat waves while winter temperatures can go to a minimum of ~2°C. Chilly winds caused by western disturbances also characterise the winters. About 80% of the rainfall occurs through the southwest monsoon, lasting from about June to September. Periodic failure of monsoons results in drought conditions.
The CGP supports a diversity of different types of vegetation, such as tropical sal forest, mixed deciduous forest, tropical teak forest and dry deciduous forest. These deciduous forests are also referred sometimes to as riparian fringe forests (Champion & Seth, 1968). Lucknow has a tropical mixed to dry deciduous vegetation (Tripathi et al., 2016; Figure 2 (2&3)). The perennial lakes and other water bodies are in abundance in and around the study region, which supports rich aquatic vegetation mainly comprised of Nymphaea, Nymphoides, Potamogeton, Lemna, Trapa, Polygonum, Cyperus, and Ipomoea (Figure 2 (4)).
MATERIAL AND METHODS
Pollen Extraction from Living Samples
To observe the detailed pollen micro-morphometry of important plants growing in and around the reserve forest, the maceration was applied to the well-identified floral parts of the 15 studied species belonging to different genera and families. The flowers were collected from the Kukrail Reserve Forest and adjoining areas in Lucknow to study pollen grains’ morphological characters under LM. The samples were chemically treated for palynological observations following Erdtman (1963) which has been elaborated below.
The freshly collected flowers of 15 plant species were identified during field observation. They were kept in water for an hour to facilitate hydration which permits easy liberation of pollen grains from the antheridium. The selection of the aforementioned plant species for the pollen morphological investigation was made based on their luxuriant growth in and around the reserve forest. Some of the common and important living plant samples, including herbs, shrubs, and trees, were taken to extract and examine the morphological characters of pollen grains under LM to make the study more comprehensive and familiarise oneself with the morphological characteristics of pollen of different families for accurate identification. The material was placed in a centrifuge tube (15 ml) and gently smashed using a thin glass rod to loosen the pollen grains from the anther wall into the water. Subsequently, sieving was performed through a 150 µm mesh sieve for the removal of any macro-remains like petals and anther cover, and the filtrate was then centrifuged to decant excess water. The remaining water content from the obtained filtrate was decanted off, followed by the glacial acetic acid treatment that is again decanted off after the centrifugation. The acetolysis mixture (9:1 ratio of acetic anhydride and sulphuric acid) was prepared and carefully poured into the sample-laden centrifuge tube (Hesse & Waha, 1989). Subsequently the centrifuge tubes were boiled inside a water-filled beaker kept on a hot plate for about 5–10 minutes with regular stirring with a glass rod to dissolve the cellulose left in the pollen wall (Erdtman, 2013). Afterward, the catalysing mixture was centrifuged and decanted; then the samples were treated again with glacial acetic acid. Subsequently, the acetolysed samples were washed with water two to three times to remove any leftover acid in the sample. The sample in the centrifuge tube was then centrifuged again to remove the extra water and the left sample with pollen grains was transferred into small plastic/glass vial tubes for further investigation.
The acetolysed samples were preserved by adding 50% glycerine with 2–3 drops of phenol to avoid microbial decomposition. The final material (pollen) was mounted on a glass slide with polyvinyl and sealed with a DPX mount to prepare a permanent slide (Erdtman, 1963). The pollen micro-morphological characters of 15 plant species were observed through a light obscuration pattern and identified by referring to the published literature (Erdtman, 1952; Rao & Shukla, 1975; Nayar, 1990). The terminology for pollen morphology was adopted from Gupta and Sharma (1986) and Punt et al. (2007). The photomicrographs for the pollen and spores were taken using an Olympus SC-50 digital camera attached to an Olympus BH-2 microscope (Plate 1). A list of major plant taxa growing in the Kukrail Reserve Forest of Lucknow District is provided in Table 1.
Pollen recovered from the living polliniferous samples of kukrail Reserve Forest. 1 & 2. Cassia fistula, 3 & 4. Bauhinia purpurea, 5. Butea monosperma, 6. Senna occidentalis, 7. Bougainvillea, 8. Rosa alba, 9. Dianthus barbatus, 10. Vernonia cinera, 11. Zinnia elegans, 12. Alternanthera tenella, 13. Justicia simplex, 14. Dendocalamus strictus, 15. Eragrastis tenella, 16. Nympheaea.

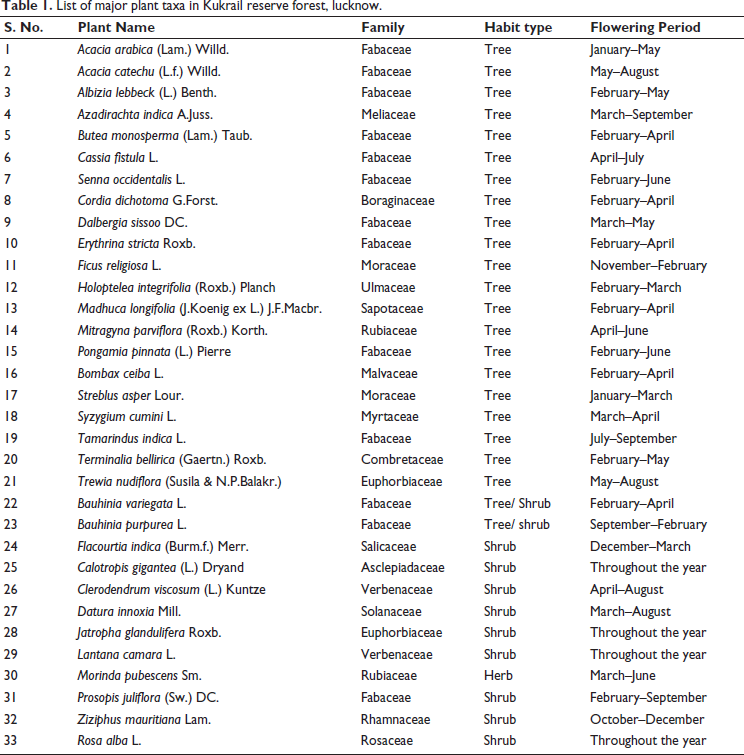
List of major plant taxa in Kukrail reserve forest, lucknow.
Pollen Extraction from Soil Samples
To understand Kukrail Reserve Forest and adjoining areas concerning modern pollen rain and pollen preservation on subsurface soil in relation to extant vegetation, the sampling sites were carefully selected to cover different land use zones. The collected surface samples were obtained from March to April and have been procured in a transect from various vegetational settings including within the forest (F 1–-5), across the open-land (O 6–10) and crop-land (C 11–15) areas from south to north of the reserve forest (Figure 1). The surface samples in the pollen spectra are designated by the site initial ‘K’ and numbered accordingly. The soil samples were collected using the fine-edged stainless steel trowel below 2–5 cm of surfaces to avoid surface rootlet and vegetal contamination and packed tightly in a ziplock plastic bag (Figure 2 (5)). The surface samples were chemically treated for the palynological observations and have been discussed in detail below:
The soil sample, weighing approximately 10–20 g, was treated with a 10% potassium hydroxide solution to separate pollen and dissolve humic acid. The resulting mixture was then filtered through a 150 µm mesh size sieve to remove large debris. Afterwards, the samples underwent three washes with distilled water to eliminate alkalis through decantation. Subsequent treatment involved the use of 40% hydrofluoric acid (HF) in plastic containers, followed by two washes with distilled water to remove silica and excess HF. Finally, the pollen residue was mixed with 50% glycerine and a small amount of phenol to protect it from microbial decomposition.
The range of total pollen count varies from 150 to 400. The non-pollen palynomorphs, like fungal remains and fern spores, have been excluded from the total pollen sum to avoid the over-representation of palynoassemblage. However, their relative frequencies have been noted for the overall preservation status of the microbiota. For the preparation of the pollen spectra and cluster analysis (CONISS), TILIA software (ver. 1.7) was used, and the cluster analysis aided in the delineation of major pollen zones (Figure 3). The pollen zones separated through the cluster analysis were named ‘KS’ after the Kukrail Spectra. The pollen spectra were further modified using the CoralDraw-17 software program. The pollen types found in the surface soil were grouped into trees, shrubs, terrestrial herbs, marshy/aquatic plants and pollen transported from high-land regions via fluvial and aeolian activities. The pollen grains were accurately identified using modern reference pollen slides from the BSIP Herbarium and published pollen morphology literature (Chauhan & Bera, 1990; Bera et al., 2009). The monolete and trilete spores are consistently recovered as fern taxa but are not included in the total pollen sum to avoid misinterpretation due to overdominance. An Olympus BX-50 Microscope with an attached DP-26 Olympus camera was used to photograph and count palynomorphs (Plate 2). Temporary pollen slides were prepared using glycerol to identify and count pollen and non-pollen palynomorphs. An average frequency of the recovered pollen taxa from the three sampling sites has been provided in Table 2.
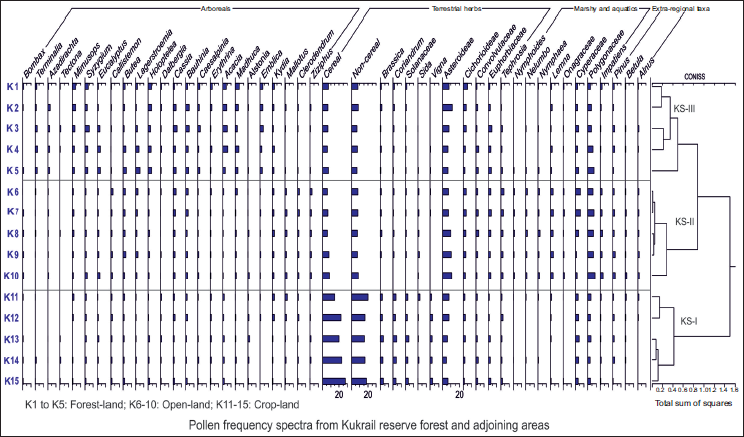
Pollen frequency spectra from different depositional sites of Kukrail reserve forest.
Palynomorphs extracted from the surface soil samples of Kukrail reserve forest. 1. Bombax, 2. Terminalia, 3. Barringtonia, 4. Acacia, 5. Cassia, 6. Syzygium, 7. Callistemon, 8. Sida, 9. Convolvulaceae, 10. Chenopodiaceae, 11. Vigna, 12. Onagraceae, 13. Non-cereal, 14. Cereal, 15. Solanaceae, 16. Brassica, 17. Potamogeton, 18. Degraded Non-cereal, 19. Unidentified, 20. Degraded monolete, 21. Trilete spore, 22. Ascospore of Cookeina, 23. Diploidia & Diporotheca, 24. Glomus.

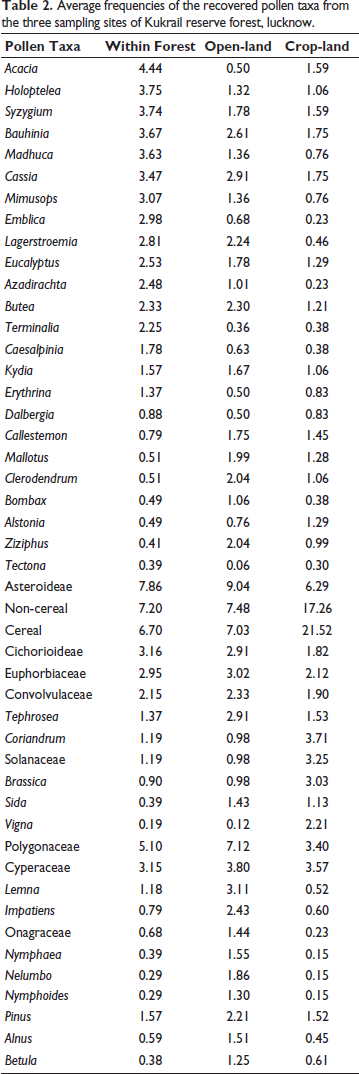
Average frequencies of the recovered pollen taxa from the three sampling sites of Kukrail reserve forest, lucknow.
Principal Component Analysis
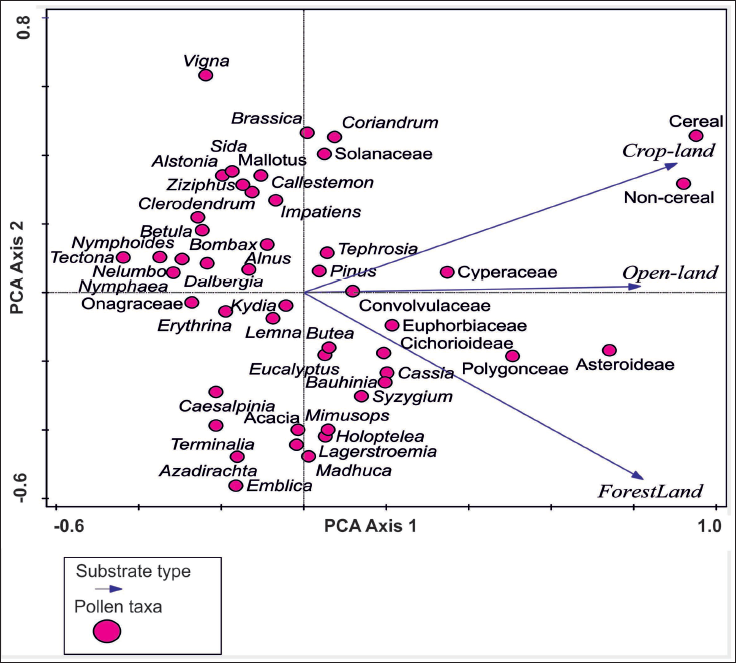
The statistical examination of pollen frequencies obtained from surface sediment samples collected from different depositional sites of Kukrail Reserve Forest, such as the forest area, open-land, and crop-land, have been analysed using PCA with the Canoco model 5 software (Smilauer & Leps, 2014; Figure 4). A p value of ≤.05 was used to determine the significance of the data. The value was imported into Corel Draw-12 software for minor editing of data labels.
PCA ordination plot showing interrelationships of recovered pollen taxa and surface soil samples collected from different depositional settings of the Kukrail reserve forest, lucknow.
Box-plot Statistical Analysis
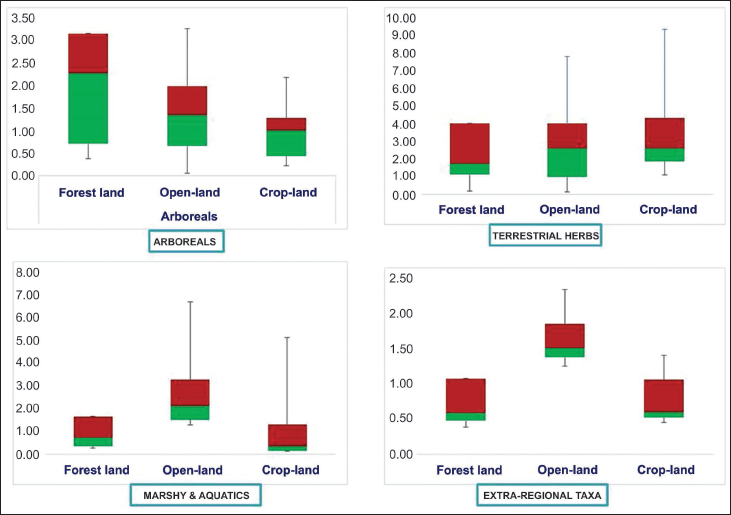
The box-plot graphs were utilised to compare and present the predominant vegetation groups across various pollen collection sites within the Kukrail Reserve Forest and its neighbouring areas. This statistical approach aided in standardising the abundance of pollen in the obtained datasets using a five-number summary, including the minimum, the maximum, the sample median and the first and third quartiles (Rousseeuw et al., 1999). The recovered pollen data are categorised into major vegetational groups (arboreals, terrestrial herbs, marshy and aquatics, and extra-regional taxa) to calculate the minimum to maximum percentile of pollen found in forest-land, then open-land and then crop-land. Subsequently, the box plot was created in a Microsoft excel worksheet using the stacked column option and then modified in Microsoft PowerPoint (Williamson et al., 1989; Figure 5).
Box plots showing the distribution of arboreals, terrestrial herbs, marshy, aquatic and extra-regional taxa in the Kukrail reserve forest of lucknow based on the modern pollen precipitation.
Pollen Morphological Results
A total of 15 pollen taxa under the Poaceae, Asteraceae, Fabaceae, Acanthaceae, Amaranthaceae, Nymphaeaceae, Nyctanginaceae, Caryophyllaceae and Rosaceae families were studied. The living floral samples were collected from the Kukrail Reserve Forest of Lucknow District. The main goal of collecting this pollen-based dataset is distinguishing between modern and fossil pollen grains found in surface and sedimentary profile samples. Thus, in this communication, exact pollen morphometric evaluation with the help of LM of 15 pollen taxa has been undertaken for the generation of palyno-morphological data, which could aid in the reconstruction of palaeoecology. The description of pollen taxa is provided below:
Monoporate. Spheroidal, 40–42 µm. Pore annulate. Exine 1.2 µm; sexine thicker than nexine, psilate, Tegillate. Flowering period: once in 25–45 years. Monoporate, spheroidal, 30–32 µm. Pore annulate. Exine 0.9 µm, sexine and nexine thickness is almost equal, retico-granulate. Flowering period: March–September. Tricolporate, spheroidal 42–44 µm. Colpi long and narrow. Exine 0.9 µm, sexine is thicker than nexine, granulate or rugulate; Tenuimarginate. Flowering period: May–September. Panporate, spheroidal 15–17 µm. Exine 0.6 µm; nexine is thicker as compared to sexine, punctate on the ridges, slight reticulation on the cavity, characterised by ridges with spines all over the ridge boundary. Flowering period: June–December. Tricolporate, spheroidal, 32–34 µm. Colpi indistinct. Os indistinct and slightly elliptical. Exine 5 µm (including spines); sexine thicker than nexine, echinolophate. Tegillate. Spinate, spines 3 µm in length. Flowering period: August–November. Tricolporate, prolate, 27×19 µm. Colpi long and narrow. Os subcircular, 2.5×63 µm. Exine 0.5 µm, sexine as thick as nexine at poles but relatively thicker at equator, bacullate, branched bacula coalescing to form a fine reticulum, Punctate. Flowering period: Throughout the year. Monocolpate, prolate-spheroidal, 36×32 µm. Exine 1.2, sexine slightly thinner than nexine, scorbiculate. Tegillate. Spinate, spines, 4 µm, blunt, densely placed. Flowering period: June–September. Tricolporate. Prolate-spheroidal (25.9×22.9 µm). Colpi long (length 27.3 µm, width 2.5 µm), tenuimarginate. Os distinct, lalongate (2.2×2.3 µm). Exine 0.7 µm thick, punctate sexine pattern. Flowering period: April–July. Tricolporate. Sub-prolate (42.3×34.9 µm). Colpi long and broad (length 42.9 µm, width 4.6 µm), crassimarginate. Os distinct, lalongate (3.3×3.6 µm). Exine 1.9 µm thick, sexine thicker than nexine, fossulate sexine pattern. Flowering period: February–June. Tricolporate. Prolate, 70×50 µm; Colpi long and broad (length 64.9 µm, width 9.3 µm), crassimarginate. Os distinct, lalongate (12.5×7.0 µm). Exine 3.4 µm thick, sexine thicker than nexine, striato-reticulate sexine pattern. Flowering period: September–February. Tricolporate, subprolate, 34×31 µm. Colpi very long and narrow, margins thickened. Os lalongate, 2.5×2 µm. Exine 2 µm; sexine thicker than nexine, reticulate with bacula suspended in the lumina. Tegillate. Flowering period: February–April. 3–4 zonocolporate, suboblate to spheroidal to prolate spheroidal, 36×35 µm. Exine 0.9 µm, nexine is thicker than sexine. Densely reticulate with granules within reticulum. Flowering period: October–March. Porate, presence of 6 pores, spheroidal, 35–37 µm. Exine 1 µm, sexine is thicker than nexine. Verrucate. Flowering period: February–May. Porate, spheroidal 25–27 µm. Exine 1.1 µm. Spinate with holes on the base of the spine; Fossulate. Flowering period: June–October. Tricolporate, subprolate, 33×30 µm, colpi long, narrow, constricted at equator, margins thickened, apices obtuse. Exine 0.9 µm, sexine as thick as nexine, psilate, Tegillate. Flowering period: Throughout the year.
MODERN POLLEN RAIN
In order to establish a modern pollen analogue for the tropical deciduous forest of Kukrail Reserve Forest, we need to collect and analyse surface soil samples from various areas within the forest, as well as adjacent open-land, crop-land, lakes and swampy areas. This analysis will help us understand how the forest components are represented in modern pollen spectra, as well as their dispersal and preservation in the sediment compared to their actual presence in the regional forest types. The comparative database created from pollen deposition and existing vegetation analysis using surface soil samples can provide a modern analogue for accurately assessing fossil pollen spectra, which would aid in reconstructing past vegetation and climate in the CGP region.
Pollen Spectra (Within Forest)
The palynological study of the samples procured from the forest-land reveals that arboreals taxa account for an average of 50.35%, which is comparatively higher than non-arboreal taxa (47.12%) and extra-regional taxa (2.54%) (Figure 3). The abundant non-arboreal taxa are grasses including non-cereal (7.20%) and cereal (6.70%) with a combined value of 13.9%. Other terrestrial herbs such as Asteroideae (7.86%) were found in higher value (>4%), followed by the fair occurrence (2–4%) of Cichorioideae (3.15%), Euphorbiaceae (2.95%) and Convolvulaceae (2.15%). Herbaceous taxa like Tephrosia (1.37%), Coriandrum (1.19%) and Solanaceae (1.19%) were encountered in lower frequencies (1–2%), while other cultural pollen taxa like Brassica, Sida and Vigna were found in scarce frequencies (<1%). Polygonaceae and Cyperaceae represents the marshy taxa with higher and fair values of 5.10% and 3.15%, respectively, followed by Onagraceae in scattered frequency (<1%). The aquatic taxa were represented by Lemna (1.18%) in lower value followed by Nymphaea and Nelumbo in scattered values. Extra-regional taxa were also encountered in the area with lower values of Pinus (1.57%) followed by Alnus and Betula in scarce amount (<1%). The frequent occurrence of ferns, including both monolete and trilete (degraded) spores, were identified within the forest soil. Fungal spores such as Diporotheca, Diploidia and Nigrospora were also encountered.
Among arboreals, tree taxa like Acacia is predominant with high value of 4.44%, followed by Holoptelea (3.75%), Syzygium (3.74%), Bauhinia (3.67%), Madhuca (3.63%), Cassia (3.47%), Mimusops (3.07%), Emblica (2.98%), Lagerstroemia (2.81%), Eucalyptus (2.53%), Azadirachta (2.48%), Butea (2.33%) and Terminalia (2.25%) in fair values (2–4%) followed by Caesalpinia (1.78%), Kydia (1.57%) and Erythrina (1.37%) in lower values, whereas the rest of the deciduous arboreals like Dalbergia, Mallotus, Bombax, Alstonia and Tectona were encountered in scattered frequencies (<1%). The shrubby taxa were represented by Clerodendrum (0.51%) and Ziziphus (0.41%).
Pollen Spectra (Open-land)
The palynological analysis of surface soil samples collected from open-land areas shows the dominance of herbaceous taxa (non-arboreals), which is around 61.82%, followed by arboreals (33.20%) and extra-regional taxa (4.98%), respectively (Figure 3). The grasses among non-arboreals were present in high abundance, comprising non-cereals (7.48%) and cereal (7.03%) with a combined value of 14.51%, followed by other terrestrial herbs like Asteroideae (9.04%) in high frequencies. The pollen taxa like Cichorioideae (3.02%), Euphorbiaceae (2.91%), Convolvulaceae (2.91%) and Tephrosia (2.33%) were present in fair values (2–4%), followed by Coriandrum pollen (1.43%) in lower frequency, while other cultural pollen like Solanaceae, Brassica, Sida and Vigna was found in scarce frequencies (<1%). Marshy taxa were represented by Polygonaceae (7.12%), Cyperaceae (3.80%), Impatiens (2.43%) and Onagraceae (1.86%), whereas aquatic taxa were represented by Lemna (3.11%) in fair value, followed by Nymphaea (1.55%), Nelumbo (1.44%) and Nymphoides (1.30%) in lower values. Extra-regional taxa were also encountered in the area with fair value of Pinus pollen (2.21%), followed by Alnus (1.51%) and Betula (1.25%) in lower frequencies (1–2%). The fungal remains like Helminthosporium were found in abundance.
Among arboreals, a rapid reduction in arboreal taxa such as Acacia (2.94%) has been observed. Besides, Holoptelea (2.61%), Syzygium (2.30%), Bauhinia (2.24%), Madhuca (2.04%), Cassia (2.04%) and Mimusops (1.99%) were encountered in fair values, while tree taxa like Emblica (1.78%), Lagerstroemia (1.78%), Eucalyptus (1.75%), Azadirachta (1.67%), Butea (1.36%), Terminalia (1.36%), Caesalpinia (1.32%), Kydia (1.06%) and Erythrina (1.01%) were found in lower values. Some other tree taxa, such as Dalbergia, Mallotus, Bombax, Alstonia and Tectona, were found in dispersed patterns (<1%). Shrubby taxa were scantly (<1%) found in open-land pollen assemblage and are represented by Clerodendrum (0.50%) and Ziziphus (0.36%).
Pollen Spectra (Crop-land)
The palynological analysis of surface soil samples procured from the crop-land showed the average value of non-arboreal taxa around 74.53%, which is dominant over arboreal taxa (22.89%; Figure 3). The moderate presence of extra-regional taxa (2.58%) was also witnessed. The non-arboreals were represented by grasses in higher abundance comprising cereal (17.26%) and non-cereal (21.52%) pollen with a combined value of 38.78%, followed by other terrestrial herbs like Asteroideae (6.29%), Cichorioideae (3.71%), Euphorbiaceae (3.25%) and Convolvulaceae (3.03%) in higher values, followed by the occurrence of Tephrosia (2.21%) and Coriandrum (2.12%) in fair frequencies. The other cultural pollen taxa, like Solanaceae (1.90%), Brassica (1.82%), Sida (1.53%) and Vigna (1.13%), were found in lower frequencies. Among the marshy taxa, Polygonaceae (3.40%) and Cyperaceae (3.57%) were dominant, followed by the occurrence of Impatiens and Onagraceae in scarce frequencies (<1%), whereas aquatic taxa like Nymphaea, Nelumbo and Nymphoides are present in very scarce frequencies (<1%). Extra-regional taxa were represented by Pinus (1.52%) with lower values, while Alnus and Betula were encountered in very scarce frequencies (<1%). The parasitic fungal spores of Solanum plants like Diploidia and Diporotheca, along with some coprophilous fungal spores like Ascospore of Cookiena, Sordaria and Podospora, were also observed in high abundance.
Among arboreals, the rapid reduction in Acacia and Holoptelea (1.75%) has been observed. Besides, Syzygium and Bauhinia (1.59% each), Madhuca (1.45%), Cassia (1.29%) and Mimusops (1.29%) were encountered in lower values. In comparison, other prominent tree taxa of the reserve forest like Emblica (1.28%), Lagerstroemia (1.21%), Eucalyptus (1.06%), Azadirachta (1.06%) and Butea (1.06%) were also found in lower values. The tree taxa like Terminalia (0.99%), Caesalpinia and Kydia (0.83% each), Erythrina (0.76%), Dalbergia, Mallotus, Bombax, Alstonia and Tectona were encountered in scattered frequencies (<1%). Shrubby taxa were represented by Clerodendrum (0.38%) and Ziziphus (0.23%) in scanty amounts (<1%).
Principal Component Analysis
The PCA score plot displays values of 0.7320 for Axis 1 and 0.1552 for Axis 2. The cumulative variation explained by the PCA score plot is 73.20% for Axis 1 and 88.72% for Axis 2. The pollen taxa observed in the soil samples from forest-land, open-land and crop-land in and around Kukrail Reserve Forest account for a total variance of 43.66%. The PCA score plot separated crop-land, open-land and forest-land into two clusters in which the dominant pollen taxa of crop-land and open-land showed closer similarity and lay in one quadrant, while the dominant pollen taxa of forest-land lay in another quadrant of the same side. After analysing the PCA score plot, we noticed that the pollen taxa such as Coriandrum, Solanaceae, Tephrosia, Pinus, Cyperaceae, cereal and non-cereal were accountable for the resemblance between soil samples from crop-land and open-land, whereas the key pollen taxa which represent the tropical forest-land were Butea, Eucalyptus, Bauhinia, Cassia, Syzygium, Holoptelea, Mimusops, Lagerstroemia, Terminalia and Madhuca.
The pollen taxa responsible for significant differentiation between soil samples of crop-land/ open-land from forest-land were cereal, non-cereal, Coriandrum, Solanaceae, Tephrosia, Pinus, Impatiens and Cyperaceae. Cereal and non-cereal pollen were exceptionally higher in crop-land surface soil. The taxa which differentiate forest-land from crop-land/open-land were Convolvulaceae, Euphorbiaceae, Butea, Eucalyptus, Bauhinia, Cassia, Syzygium, Holoptelea and Madhuca, Mimusops, Lagerstroemia, Erythrina and Terminalia. The pollen taxa like Madhuca, Emblica, Terminalia, Azadirachta, Lagerstroemia and Erythrina were the indicators of forest-land. Obviously, there is relatively less representation in crop-land and open-land, which was well noted in the PCA loading plot (Figure 4). At the same time, the pollen taxa which were the indicator of crop-land/open-land were cereal, non-cereal, Vigna, Coriandrum, Solanaceae, Brassica, Sida, Cyperaceae, Ziziphus, Bombax, Dalbergia and Nelumbo.
DISCUSSION
The pollen morphological investigation of 15 plant taxa from the Kukrail Reserve Forest and surrounding areas was initiated to examine the primary pollen features like aperture number, position and character (according to NPC classification), which are best examined under LM and have useful implications in increasing the taxonomic resolution of fossil pollen identification, particularly for palaeofloral and palaeoecological records (Farooqui et al., 2019). The sexine pattern is also examined closely since it is regarded as the secondary pollen morphological feature for the identification of most of the genera, species and subspecies (Garg et al., 2021).
The structure of pollen is closely associated with the process of pollination (Garg & Rao, 1997; Rao et al., 1999). Pollen with thick surface ornamentation (like in Vernonia cinerea, Justicia simplex, Bauhinia purpurea and Zinnia elegans) are more prone to being stuck on the body parts of pollinating insects as opposed to smooth pollen grains (e.g., Poaceae). Moreover, the pollen wall thickness and the number of apertures play an important role in pollen germination on the stigmatic surface (Walker & Doyle, 1975; Dajoz et al., 1995). Pollen morphological characters, thus, suggest alternative reproductive strategies for successful pollen transfer to the stigma for their conservation and proliferation ex situ and/or in situ, coupled with authentic identification for both their systematics and palaeoecological implications (Garg & Rao, 1996; Tripathi et al., 2022). Pollen micro-morphometric analysis of important plant taxa under LM is crucial for accurate identification and critical for detecting the fossil counterpart in the sediment of that region for the reconstruction of palaeoflora vis-à-vis past climatic changes.
The major arboreal taxa recovered from the surface samples are Acacia, Butea, Bauhinia, Emblica, Lagerstroemia, Mimusops, Bombax, Syzygium and Ziziphus. The remaining constituents of the reserve forest, including Erythrina, Flacourtia, Mitragyna, Pongamia and Trewia, are either poorly or not represented in the varied pollen substrates. The reason may be low pollen productivity owing to the entomophilous nature of pollination, as well as irregularity of the monsoon during recent years, which governs the flowering period of these taxa. The low pollen dispersal efficiency as well as poor preservation of pollen in the sediments and differential recognition of poorly preserved grains also cannot be ruled out (Quamar & Chauhan, 2013). These pollen grains might have low sporopollenin content in their exines, which could have acted in a negative way to degrade the pollen grains of these plants (Havinga, 1984). The pollen of teak (Tectona grandis) has been recorded in trace amounts owing to its sporadic presence on the forest floor; however, its low pollen dispersal efficiency also cannot be excluded (Quamar & Bera, 2015). The high pH value of soil, as well as the microbial and chemical degradation of the pollen grains of these taxa in the sediments, might have been detrimental factors contributing to the scarcity of pollen of these plants in the sediments (Sharma, 1985; Bera et al., 2008).
The pollen assemblages in the surface samples indicate the presence of tropical mixed deciduous vegetation under warm and humid climates in and around the region with scattered patches of vegetation in open-land areas. These tropical mixed arboreal taxa are noticeable as long as the study is limited to the forest areas. However, the representation of the arboreal taxa like Acacia, Holoptelea, Syzygium, Madhuca, Bauhinia, Cassia and Mimusops decreases proportionately in samples taken away from the forest, such as in open-land and crop-land areas. Some herbaceous and shrubby pollen types, including Tephrosia, Euphorbiaceae, Convolvulaceae, Asteroideae, Cichorioideae, Clerodendrum and Ziziphus, do not indicate significant changes in most cases inside or outside the forest. These plants generally thrive post to forest clearance or, due to other biological influences, require further investigation.
The modern pollen analogue generated in the present study also suggests the biodegradation of palynomorphs and other microbiota in swampy sediments. It emphasises the complexity of interpreting pollen diagrams constructed from unique forest stands with minimal or no tree taxa. Some of the key pollen constituents of tropical deciduous forests like Erythrina, Adina and Flacourtia were found missing, owing to their production and dispersal biases. In such conditions, the closely associated arboreals, as well as non-arboreals, should be considered to study the various developmental phases of the forest on both spatial and temporal scales. The generated modern pollen analogue, thus, presents strong background data to decipher the past vegetation and climate changes in the tropical forests of Kukrail Reserve Forest.
Pollen Inferences from Forest-land
The surface soil samples from the forested area have been palynologically investigated from the Kukrail Reserve Forest. This study showed that there were an approximately equal proportion of tree pollen (arboreals; 50.35%) and non-tree pollen (non-arboreals; 47.12%). The pollen taxa like Holoptelea, Syzygium, Bauhinia, Madhuca, Cassia, Mimusops, Emblica, Erythrina, Bombax, Butea and Terminalia indicate the existence of factual forest composition under tropical dry deciduous forest along with Ziziphus and Clerodendrum, which are incorporated as the prominent shrubby taxa in the palynoassemblage. Among the Combretaceae family, the Terminalia pollen with unique polycolporate aperture conditions are frequently identified and noticed in the palynoassemblage and are representative of tropical mixed deciduous forests. The terrestrial herbs such as Cichorioideae, Euphorbiaceae, Tephrosia, Coriandrum, Solanaceae, Brassica and Sida were represented at fair to lower values.
Interestingly, the grass pollen is found in appreciable frequencies even within the forest area (up to 14%). The occurrence of cereal pollen, along with other cultural pollen taxa like Brassica, Sida and Vigna, indicates the acceleration of pastoral activities, whereas non-cereal pollen, in general, represents vast open grassland, though the same may occur at patches within the forest and may be attributable to other anthropic factors. The monolete spores show higher values as compared to trilete spores in almost all the samples. However, some monolete ferns are not of a local nature, which needs further investigation (Tripathi et al., 2020; Tripathi & Pandey, 2023). Extra-regional taxa have marked in lower frequency (2-3%) in forest samples as compared to open-land samples (4–5%), attributed to the obstacles created by the trees for the wind activity. However, crop-land samples record the extra-regional taxa in lower frequencies. In the forest sediment, grass pathogens belonging to the Ascomycetes and Deuteromycetes are commonly found. The fungal spores, like Nigrospora, Diporotheca and Diploidia, were frequently observed in forest soil, indicating a humid depositional environment (Tripathi et al., 2021). The modern pollen analogue provides clear evidence that the distribution pattern of pollen has enabled the deduction of the pollen/vegetation relationship within the forest.
Conventional modern pollen surface samples with a checklist of plants in the immediate vicinity of the sites have been used as the basis for interpreting the past records extensively in the Indian subcontinent (Anupama et al., 2000; Dixit & Bera, 2011, 2013; Quamar & Stivrins, 2021; Quamar, 2022). Quantitative estimates of the plants in the immediate vicinity and up to 1 km around the site, combining field surveys and remote sensing, take this one step further in providing distance-weighted plant abundances and are important for understanding the pollen dispersal and deposition characteristics of particular taxa in an environment. It is necessary to know how the pollen is spatially dispersed and deposited at the site of preservation, with reference to its source taxon. Once the spatial frameworks are attached to the pollen analysis, investigation of a range of important issues, particularly environmental and climate change, would be possible with improved reliability and confidence, which further facilitate the use of palaeoecological data to address issues in ecology, climatology, palaeontology and archaeology (Tripathi et al., 2022).
Pollen Inferences from Open-land
The pollen data of five surface soil samples procured from the open-land areas of the Kukrail Reserve Forest reflect the relative reduction in arboreal taxa (33.20%) as compared to the forest-land soil samples. Ziziphus and Clerodendrum are represented as the shrubby taxa in the palynoassemblage with sporadic values and without any significant changes. Unlike arboreal taxa, the non-arboreals (61.82%) are consistently recorded in relatively high frequencies. The herbaceous taxa are represented by Asteroideae, Cichorioideae, Euphorbiaceae, Convolvulaceae, Tephrosia, Coriandrum, Solanaceae and Brassica (3–9%). The extra-regional taxa like Pinus, Betula and Alnus attain improved value in the open-land samples, attributed to the changing face of the forest (outskirt-open-land) where strong wind found actively carrying thermophilic pollen taxa from nearby or distant higher reaches and finally got incorporated in sediment. However, the extra-regional taxa were found in relatively low frequencies in the crop-land samples. The fungal remains were found in fair frequencies in open-land soil samples owing to the location of adjacent crop fields, where air is mostly charged with pathogens and finally incorporated in the sediment.
Pollen Inferences from Crop-land
An important feature of the palynoassemblage recovered from the crop-land is the reasonable enhancement of non-arboreal taxa (75%), especially the crop taxa, including cereal, non-cereal pollen and other cultural pollen taxa (Figures 3 and 5). Moreover, the crop-land samples procured further off the forest stand show less or sporadic frequencies of tree pollen taxa. However, there is no evident change in the frequency of a few terrestrial herbaceous taxa in this case. The marshy and aquatic taxa are well represented because of their local origin as they luxuriantly grow in and around the swampy areas in the proximity (Tripathi et al., 2016). The poor dispersal efficiency, low pollen productivity owing to entomophily as well as biodegradation of pollen in sediments could be the reason for the irregular representation of certain tree pollen taxa like Erythrina, Madhuca, Mitragyna, Pongamia, Trewia and Flacourtia, which are frequently occurring along the forest. The herbaceous plant taxa, especially Asteroideae, Cichorioideae and Euphorbiaceae, with local abundance, also create uncertainty and consequently serve a distorted picture due to the over-representation of their pollen in the soil. Therefore, such factors must be considered precisely while interpreting the fossil pollen assemblages for the reconstruction of past ecology.
Interpretation from PCA Analysis
The score plot of the PCA depicted the two pollen cluster separations between forest-land and open-land plus crop-land surface soil in which open-land soil samples show a closer resemblance with the crop-land surface soil and falls in one quadrant, while the forest surface soil falls separately in another quadrant of the same side. The PC score plot reveals the overall two-cluster separation between the tree pollen taxa and non-tree pollen taxa (including grasses, terrestrial herbs, marshy taxa and shrubs; Figure 4). The relatively high frequencies of tree pollen taxa like Erythrina, Kydia, Acacia, Terminalia, Azadirachta, Emblica, Lagerstroemia, Syzygium, Butea, Bauhinia, Holoptelea, Madhuca, Cassia and Mimusops could be well observed in forest samples. It was clearly observed in the PCA score plot that the characteristic pollen retrieved from forest soil samples include the dominance of core arboreal taxa of the extant forest in the region, which shows that the forest soil is a potential substrate for preserving extant pollen and can serve as a modern analogue for depicting past vegetation vis-à-vis climatic history of this region.
The dominance of grasses along with other cultural pollen taxa like Brassica, Solanaceae, Coriandrum and Tephrosia near the crop-land quadrant indicates the agricultural and anthropogenic activity in the region according to the PCA score plot. The occurrence of marshy taxa like Impatiens, Cyperaceae and Onagraceae near the open-land arrow indicates that these open-lands include small swampy areas with perennial water systems. According to the PCA analysis, it is concluded that the differential values in arboreal and non-arboreal pollen taxa in forest-land and open-land (including crop-land) is responsible for the groupings.
Inferences from the Box-plot Analysis
According to comparative box-plot analysis, arboreal pollen taxa in forest-land have greater frequencies and diversities, whereas terrestrial herbs are relatively more abundant in crop-land (Figure 5). The frequency and diversity of marshy, aquatic and extra-regional taxa are relatively more dominant in crop-land soil samples.
Biological Degradation of Pollen and Spores In and Around Kukrail Forest
The occurrence of degraded pollen spores is not a common phenomenon, but it is still reported in some parts of the world. Bera et al. (2008) published a record of degraded pollen spores from Assam, northeast India. Here in our study, among the degraded forms, pollen of Malvaceae, Cyperaceae, Potamogeton and Bombax were less noticed compared to fern spores. The most common degradation pattern found in pollen spores is the circular, rosette-to-branching perforations (wedge-shaped) and crumpled broken forms. Sometimes, the identification of pollen and spores could be very difficult or impossible.
It is mandatory to consider pollen data from all ecosystems, covering all latitudes, to apply the technique for the global biome distribution that is used for data model comparison in the framework of global change projects (Prentice et al., 1996; Tripathi & Pandey, 2023). The generated modern pollen analogue data from the Kukrail Reserve Forest of the CGP may be useful in providing the background information for past ecological studies, especially in tropical dry deciduous forests, which are more prone to urbanisation and anthropogenic activities.
CONCLUSIONS
The newly generated modern pollen analogue from Kukrail Reserve Forest areas of Lucknow District portray the occurrence of tropical dry deciduous forest mainly composed of Terminalia, Butea, Emblica, Syzygium, Bombax and Holoptelea under warm and humid climate in response to local rainfall variations. Moreover, for the accurate identification of fossil pollen types in the surface soil and sediments, the pollen morphological characteristics of 15 plant species have been studied in detail. Overall, the non-arboreal taxa (especially grass pollen and terrestrial herbs) are dominant in the surface palynoassemblage. The relatively lower values of arboreal taxa (trees and shrubs) were observed in open-land and crop-land areas. The frequencies of prominent tree taxa in the forest palynoassemblage were appreciable when compared to the open-land and crop-land. The multivariate PCA and box-plot analysis also complement the generated modern pollen data. The extant vegetation and surface pollen data quite cohere, and therefore, this modern pollen analogue serves as a potential baseline data for the reconstruction of past vegetation and climate change in the CGP.
Furthermore, the frequent encounter of degraded pollen and spores was observed, attributed to the parallel occurrence of different fungal spores in abundance in the Kukrail soil samples, which requires detailed investigation. This pollen preservation bias would also be expected in Holocene pollen records of northern India and, hence, could provide information on the actual vegetation when analysing fossil pollen samples. It is an imperative need to conserve this reserve forest to protect its unique biodiversity in the wake of ongoing anthropogenic pressure and climate change. Therefore, this study could provide insight into the potential of formulating palynological studies with reference to extant vegetation to assess the preservation of microbiota on the surface soil of the Kukrail Reserve Forest. Overall, such modern analogue studies would definitely assist in scheming restoration strategy and remediation action in and around the reserve forest to conserve a healthy environment for both flora and fauna.
Footnotes
Acknowledgements
Thanks are due to the Director, Birbal Sahni Institute of Palaeosciences, for the permission and for providing all the necessary facilities for carrying out this research project (Permission no. BSIP/RDCC/Publication no. 39/2024-25). We are highly gratified to HOD, Department of Geology, Lucknow University, for the encouragement and permission. We would also like to thank Ms. Arya Pandey for the statistical assistance.
Declaration of Conflicting Interests
The authors declared no potential conflicts of interest regarding the research, authorship, and/or publication of this article.
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: The Birbal Sahni Institute of Palaeosciences, Lucknow, and the Department of Geology, Lucknow University, financially supported this research.