
Abstract
The Yellow Flagstone is the oldest member of the Goradongar Formation exposed in the Pachchham Island, Kachchh Basin. The members exposed in four sections are mapped for trace fossils, and 17 ichnofossils have been documented. The concept of ichnoguild (IG) is applied to reveal the infaunal tiering structure and the strategies organisms adapt to exploit the resources. Ten IG of similar characteristics and ecological functions have been identified. Five are shallow-tier IGs (Asterosoma IG, Gyrochorte IG, Nereites IG, Planolites IG and Bolonia IG), three middle-tier IGs (Rhizocorallium IG, Arenicolites-Diplocraterion IG and Zoophycos-Teichichnus IG) and two deep-tier IGs (Skolithos Arenicolites IG and Thalassinoides-Ophiomorpha IG). The analysis of three ichnological tiers and their IGs, namely shallow, middle and deep-tiers across the stratigraphy of Yellow Flagstone Member suggests a transition from a lower shoreface environment to offshore transition environments. The transition is supported by an increase in deep-tier IG and the marginal increase in the ichnodiversity and density. Changes in the composition and abundance of IGs over time can indicate shifts in sedimentation patterns, sea level fluctuations and paleoenvironmental conditions within the basin. Thus, the findings suggest an environmental shift from shoreface to offshore transition indicative of a transgression during the deposition of upper Bajocian to lower Bathonian Yellow Flagstone Member in the Kachchh Basin. The study also examines the temporal and spatial changes in the IGs, contributing to a better understanding of ecological changes within the basin. The findings contribute to the broader knowledge of paleoecology and provide valuable data for sedimentological and stratigraphic interpretations in sedimentary basins.
Introduction
Ichnological analysis has gained significant prominence as a potent tool for assessing paleoenvironmental changes and related alterations in recent times. It has become an essential component that bolsters sedimentological and stratigraphic interpretations, making it indispensable for studies on sedimentary basins (Buatois & Mángano, 2011; Knaust, 2012).To evaluate these changes, the ichnological record contributes significantly to our understanding of paleoecologic analysis (Buatois et al., 1998; Buatois & Mángano, 1993; Carmona et al., 2004; Crimes, 2000; Crimes & Fedonkin, 1994; Jensen et al., 2005; Mángano & Droser, 2004; Orr, 2001; Seilacher, 1956, 1977; Seilacher et al., 2005; Uchman, 2006). Considering distinctive traces of soft-bodied organisms may yield a more accurate picture of species richness and feeding-type diversity within fossil communities (Rhoads, 1975).
Bromley (1990) established the ‘Ichnoguild’ (IG) concept, which can be used to assess eco-space colonisation throughout geologic time in specific ecosystems (Buatois et al., 1998). An IG considers three factors: bauplan, food source and space utilisation (Bromley, 1990, 1996). The recurring patterns of the IGs reveal activity levels within the substrate and feeding style and reflect colonisation events of different levels. It provides valuable information for understanding the patterns of eco-space utilisation through geological time as well as useful tools for elucidating the adaptive strategies displayed by benthic producers (Bromley, 1994; Buatois et al., 1998; Buatois & Mángano, 2003).
The Kachchh Basin preserves priceless geological and paleontological marvels, providing significant insights into the Mesozoic Rift evolution and sedimentation (Desai & Chauhan, 2021). The middle Jurassic sediments are well exposed in the Kachchh Basin allowing tracing of the rift basin evolution (Biswas, 1980, 2016b; Desai & Chauhan, 2021) (Figure 1). Biswas (2016a, 2016b) provides a detailed updated summary of the structure, tectonic framework, stratigraphy and evolution of the basin. In the Kachchh Mainland, the older three Mesozoic formations, viz., Jhurio, Jumara and Jhuran, have gained preference due to their richness of body fossils, trace fossils and well-documented outcrops (Chauhan et al., 2022). The majority of research on Kachchh’s ichnology concentrates on stratigraphic formations or members (Choudhary et al., 2023; Darngawn et al., 2019; Desai, 2012, 2013, 2016; Desai & Biswas, 2018; Desai & Chauhan, 2021; Desai et al., 2008; Desai & Saklani, 2012, 2015; Ghare & Kulkarni, 1986; Joseph & Patel, 2015; Joseph et al., 2020; Kulkarni & Ghare, 1989, 1991; Padia et al., 2022, 2024; Patel et al., 2008, 2009, 2014; Shringarpure, 1986; Solanki et al., 2017). However, very few studies have evaluated the ichnological history of the entire basin (e.g., Badve & Ghare, 1978; Desai & Biswas, 2018; Fürsich, 1998; Howard & Singh, 1985).
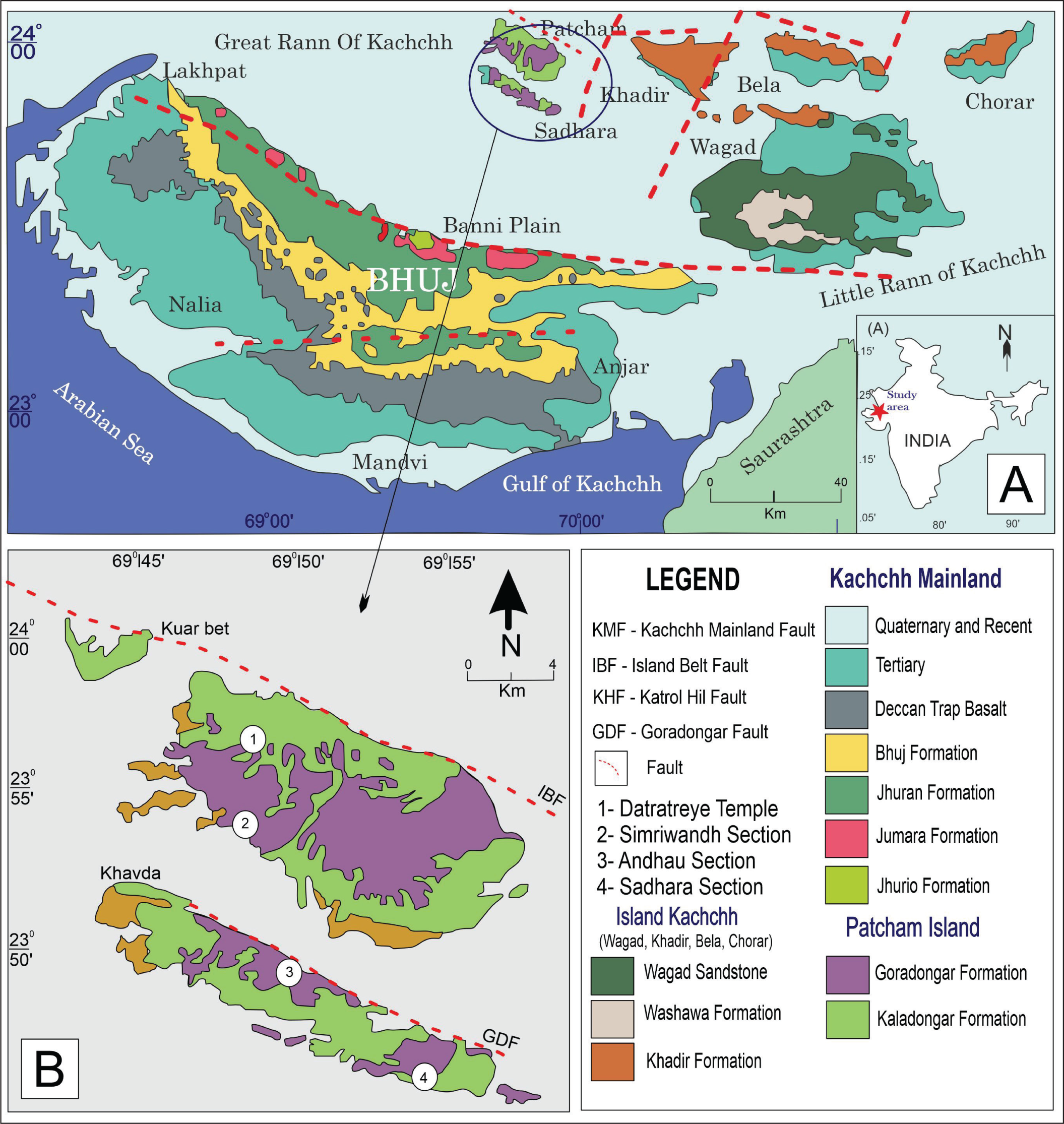
(A) Outline map of India showing the location of the study area; (B) Regional geological map of Kachchh basin (modified after Biswas & Deshpande, 1970); (C) Geological map of the study area (modified after Biswas & Deshpande, 1973; Patel et al., 2013) with the sections of Yellow Flagstone Member studied.
The Yellow Flagstone Member is the oldest member of the Goradongar Formation exposed on Pachchham Island, Kachchh Basin (Figure 2). The member is characterised by interbedded shale and flaggy limestone, also called flagstone, which is easily recognisable in Pachchham Island due to its bright golden yellow colour (Fürsich et al., 2023). With its well-preserved ichnofossils, it offers an exceptional opportunity to establish the ecological dynamics and behaviour of the organisms that once colonised the sediment. The present article is based on extensive ichnological studies of Yellow Flagstone Member at the Dattatreya temple, Sadhara dome, Andhao and Simriwandh sections. The temporal and spatial changes in the IG of different sections of Yellow Flagstone Member have helped decipher the rift basin’s distal and proximal parts.
Material and Methods
Methodology
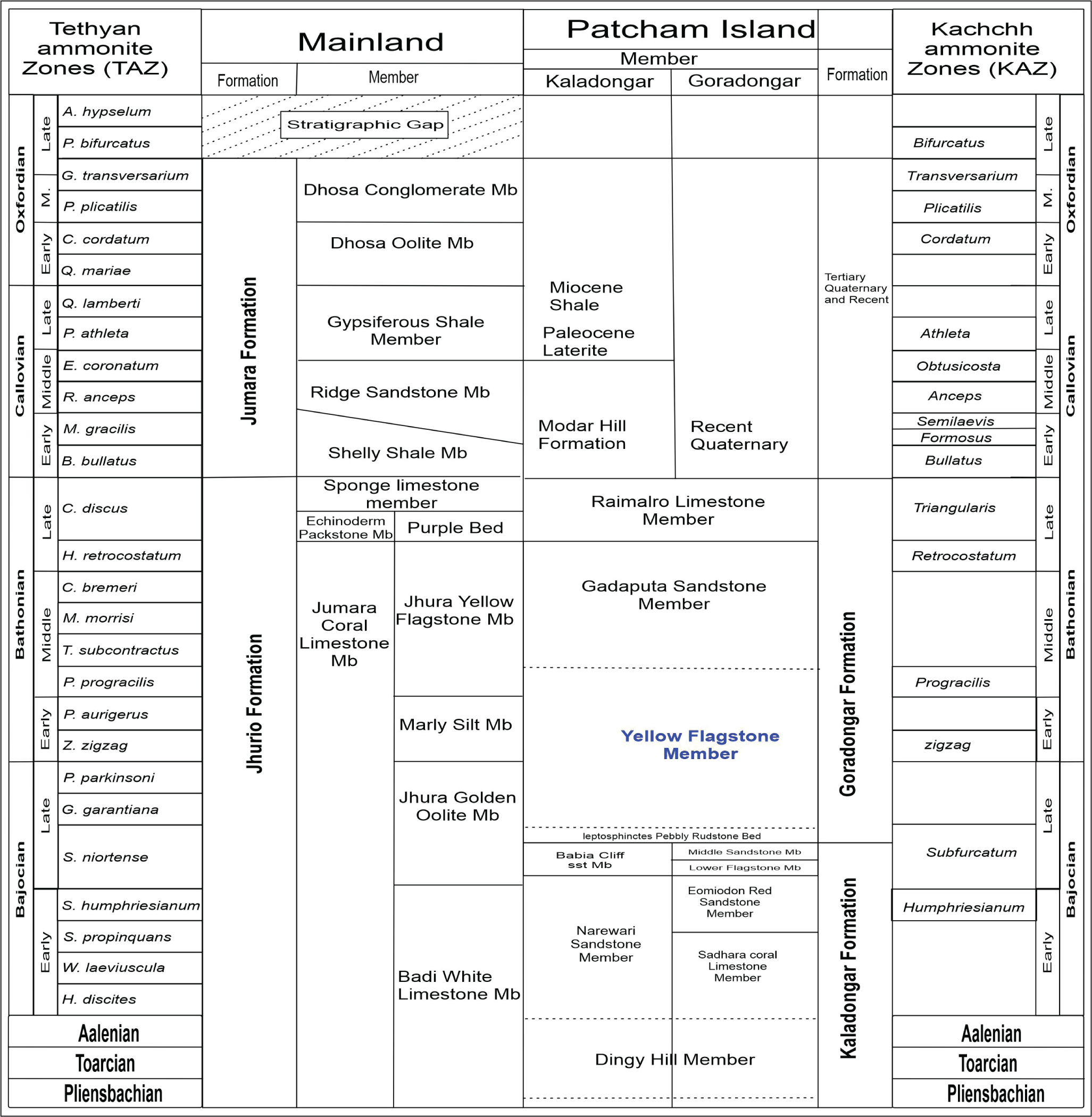
The data presented here were collected during several fieldwork excursions to the Yellow Flagstone Member exposed on Pachchham Island. Standard ichnological methodologies were employed to identify and categorise trace fossils. Bed-scale identification and descriptions of trace fossils were utilised to gain ichnological field data. Dimensions, lining, infilling, cross-cutting relationships, density and diversity have been examined. Previous works on sedimentology and palaeontology (Biswas, 1977; Fürsich et al., 1994; Joseph & Patel, 2015; Pandey & Fürsich, 1998) are correlated with our ichnological findings and a composite litho-log has been prepared (Figure 2). The classification of ichnofossils includes ethology and trace morphology. The lithostratigraphy of Pachchham Island is followed after Biswas (2016a) and Fürsich et al. (2023).
Mesozoic lithostratigraphy of the Kachchh Basin updated after Biswas (2016, 1977, 1993); and Pachchham island Stratigraphy Fürsich et al. (2001, 2023).
Site Selection
The Yellow Flagstone Member contains exceedingly diverse and extraordinarily well-preserved trace fossil assemblages exposed in different sections on Pachchham Island. The member serves as a marker horizon all over the Island.
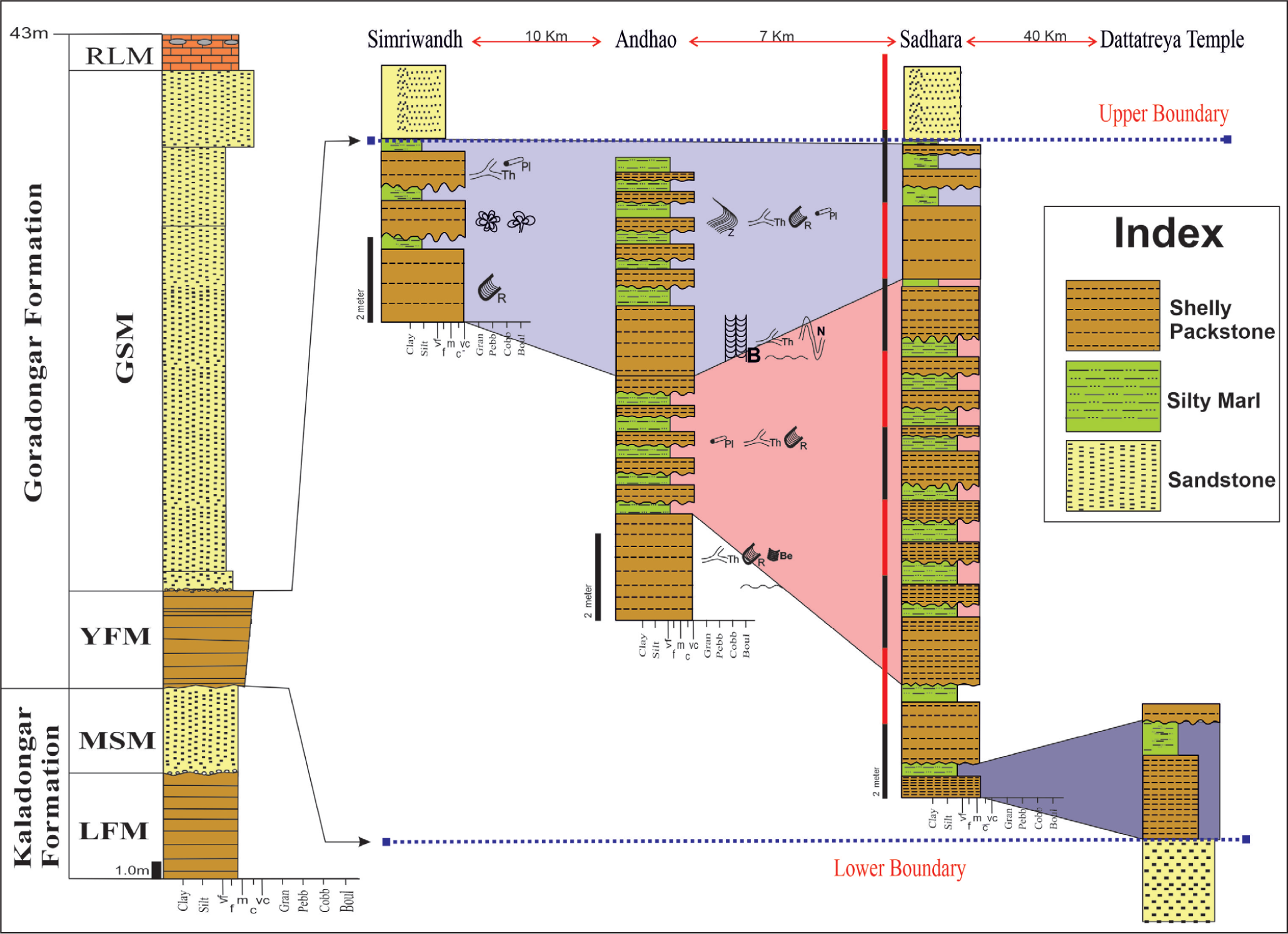
The investigation focused on trace fossils at four locations (Figure 1C): Dattatreya temple, Simriwandh, Andhao and Sadhara Dome. Lithostratigraphic correlation of the member was done based on tracing local marker beds over short-spaced lithological columns along with Ichnological and previously published data (Figure 3)
Lithostratigraphic correlation of the Yellow Flagstone Member (RLM = Raimalro Limestone Member, GSM = Gadaputa Sandstone Member, YFM = Yellow Flagstone Member, MSM = Middle Sandstone Member, LFM = Lower Flagstone Member).
The Dattatreya temple section (23056’02.6”N; 69048’ 50.8”E) is located on the top of Kaladongar Hills near the bus stand of Dattatreya temple. Here, the Flagstone Member sits on top of the Babia Cliff Sandstone Member from the Kaladongar Formation. The section records the facies change from sandstones to flaggy limestones and shales. Bioturbation is widespread. Trace fossils such as Rhizocorallium and Gyrochorte are abundant and well preserved on the ripple-laminated bioturbated flagstones.
Simriwandh section (23052’20.5”N; 69049’21.7”E). It is a small river section near Simriwandh Village. It has a symmetrical plunging anticlinal fold that exposes the Yellow Flagstone Member. The succession consists of medium- to fine-grained yellow flaggy calcareous sandstones with an erosional base.
The section starts with ripple cross-laminated thin flagstones with shale intercalations. Asterosoma and Rhizocorallium are exceptionally well-preserved on these symmetrical ripple surfaces. Beds in the upper part are thickly bedded with planar laminations and trace fossils such as Thalassinoides and Ophiomorpha. The section shows the contact between the flagstone and the middle Gadaputa Sandstone Member. The well-preserved trace fossils occur in high abundance and diversity.
Andhao section (23052’20.3”N; 69049’20.8”E). The distance between Simriwandh and Andhao section is 11 km towards the south on Sadhara Road. At Andhao, 3 m of flaggy sandstones with shale intercalations and abundant ichnofossils are exposed. The members can be divided into two parts. The lower part is a thick yellow calcareous sandstone (flagstone), followed by thin beds of shale and sandstone with erosional contacts. The lower beds are highly bioturbated and rich in trace fossils like Thalassinoides, Rhizocorallium, Planolites and Palaeophycus. The upper beds are rich in locomotion and feeding traces, such as Nereites, Bolonia, Zoophycos and Gyrochorte.
Sadhara section (23045’98”N; 69054’20.24”E). The Sadhara dome is the southern end of Pachchham Island, which exposes rocks of the Goradongar Formation. The Yellow Flagstone Member is exposed in the road section connecting the Sadhara and Juna villages. The beds are inclined and show intercalations of flagstone and shale with an erosional base. The beds are highly bioturbated by Rhizocorallium, Thalassinoides and Planolites.
Geological Setting
Different sedimentation histories in the Kachchh Basin (Mainland, Island Belt and Wagad Uplift) led to different lithostratigraphic schemes for each area. In the Island Belt, Pachchham Island (Figure 2), Kaladongar and the Goradongar Formations are exposed, respectively, in the northern Kaladongar and southern Goradongar hill ranges (Biswas, 2016; Fürsich et al., 2023). They deposited during the syn-rift phase of the basin.
The Goradongar Formation consists of the basal Yellow Flagstone Member (Containing Leptosphintes Pebbly Rudestone as a base), the middle Gadaputa Sandstone Member and the upper Raimalro Limestone Member. The formation is composed of yellow, thin-bedded, fossiliferous, partly bioturbated, partly laminated, fine-sandy calcareous sandstones, limestones and marl (Fürsich et al., 1994; Patel et al., 2013, 2014). The formation records transgressive deposition on an overall inner and mid-shelf homoclinal ramp and was deposited in distal shore settings of the Bathonian age (Patel et al., 2013).
The Yellow Flagstone Member, the focus of our study, is composed of flaggy, grey and yellow limestones containing fossiliferous and golden oolitic bands, resembling ‘member F’ of the ‘Jhurio Formation’ (Biswas, 1977, 2002). The flagstone has an erosional base and the contact is sharp. Individual cycles start with thickening upward bioturbated flagstone (Fürsich, 1998). The bioturbation intensity increases upwards and reaches its maximum at the top. The beds exhibit large-scale low-angle cross-bedding, small-scale ripple bedding, oscillatory ripples and scattered shells. The intercalated siltstone is parallel laminated. Brachiopods and gastropods along with rare ammonites are present. The member is highly bioturbated and is rich in trace fossils such as Rhizocorallium, Asterosoma, Zoophycos and Nereites.
Age of the Yellow Flagstone Member
As the Goradongar Formation is poor in age diagnostic ammonites its age are revised periodically. Two ammonite levels are recorded from Yellow Flagstone Member (Pandey & Fürsich, 1998). The older level is a shelly, fine-sandy marly micrite (Fürsich et al., 1994; Pandey & Agrawal, 1984; Pandey & Westermann, 1988), while the upper level is a rubbly, bioturbated fossiliferous micrite, about 1.2 to 1.5 m above the first level (Pandey & Callomon, 1995). This faunal horizon coincides with the type horizon of the Middle Bathonian P. Arkelli of Madagascar representing the Progracilis Zone of Europe (Pandey & Callomon, 1995). The age of the Goradongar Formation is revised recently by Fürsich et al. (2023) now spanning from the Late Bajocian G. grantiana zone to the Late Bathonian C. discus zone. The basal part also includes the marker bed of Leptosphinctes Pebbly Rudstone bed (Fürsich et al., 2023).
Ichnotaxonomy
The Yellow Flagstone Member is characterised by a high diversity ichnofauna consisting of 18 ichnotaxa, namely: Asterosoma radiciformis, Arenicolites variabilis, Bolonia lata, Diplocraterion isp, Gyrochorte comosa, Gyrophyllites geryonides, Nereites missouriensis, Ophiomorpha nodosa, Palaeophycus tubularis, Phycodes palmatum, Planolites beverleyensis, Rhizocorallium commune var. irregulare, Skolithos linearis, Taenidium serpentinum, Teichichnus rectus, Thalassinoides horizontalis and Zoophycos villae. The main ichnotaxa are described in the following.
Ichnospecies: Asterosoma radiciformis von Otto, 1854
Ichnospecies: Asterosoma ludwigae Schlirf, 2000
Horizontal to inclined burrows, star-like arranged bulbs, or bulbs that bud from a circular to elliptical tube in a dichotomous to fan-like pattern; burrow wall with or without longitudinal, subangular furrows and striae (Häntzschel, 1975; Schlirf, 2000). The possible producers of Mesozoic Asterosoma are decapod crustaceans (Häntzschel, 1975). The concentric laminae represent the feeding behaviour, while the massive tubes represent the locomotion behaviour (Schlirf, 2000). Two ichnospecies of Asterosoma are present in the Simriwandh section. A. radiciformis differs from the A. ludwigae in its radiating and branching system. In A. radiciformis the bulb length varies from 4 to 6 cm; the width is 1–1.5 cm. A. ludwigae, has a non-radiating longitudinal structure. The long axis of the bulbs is 2–3 cm, the short axis is 1.5–2 cm and the height is 0.6 cm. The overall size of the burrow is 10 cm in length and 2 cm in width.
Asterosoma is interpreted as a feeding structure of a vermiform organism (Chamberlain, 1971; Pemberton et al., 2001; Seilacher, 2007; Vossler & Pemberton, 1989) and is commonly found in fully marine environments, ranging from shoreface to offshore (Farrow, 1966; Frey & Howard, 1970; Vossler & Pemberton, 1989).
Ichnospecies: Arenicolites variabilis Fürsich, 1974a
U-shaped lined burrows, oriented vertically to inclined to the bedding plane without spreite. The arms of the U-tubes are straight or curved and parallel to each other. In cross-sections, they also appear as varying J-shaped morphologies. Burrowing depth varies between 10 and 15 cm, with a burrow diameter of 6–10 mm and a distance between two arms of 10–25 mm. Arenicolitess is the burrow of a suspension feeder (Hakes, 1976) welling and feeding structures of crustacean-like creatures’ (Goldring, 1962). These are distinguished from Diplocraterion by the absence of spreiten (Fürsich, 1974a). Although it has been found in various environments, including non-marine ones (Gillette et al., 2003), it is most commonly found in shallow marine environments.
Ichnospecies: Bolonia lata Meunier, 1886
Horizontal, straight, unbranched, bilobate trace fossil with a somewhat heart-shaped outline in cross-section with positive epirelief. The width of the ribbon varies between 1.6 and 4 cm, and the thickness of individual pads is between 0.2 and 0.5 cm. B. lata, described and documented by Schlirf (2000), is a close match for the ribbons. The trace fossil is consistent with a pascichnia ethological class and is likely created by the lateral movement of an infaunal deposit-feeder through the sediment. Bolonia resembles Protovirgularia, Gyrochorte, Scolicia.
A thorough revision of Scolicia clarifies the genus and distinguishes it from Bolonia. The only ichnospecies of Bolonia is B. lata, Meunier (1886). The paleoenvironment from which B. lata has been reported so far is nearshore marine, influenced by storm waves.
Ichnospecies: Diplocraterion isp
U-shaped burrows with spreite, with a tube diameter varying from 2 to 8 mm and a distance between the two arms up to 30 mm. In the longitudinal section, the depth of the burrow reaches 9 cm. Burrows are lined with burrow fill material different from the host sediments. Fürsich (1974b) provided a detailed account of significant features related to ichnospecies of Diplocraterion. Such dwelling burrows were made by suspension-feeding organisms in high-energy settings (Goldring, 1962).
Ichnospecies: Gyrochorte comosa Heer, 1865
On top of sandy beds, plait-like ridges possibly were produced by a polychaete worm that pushed sediment sideway, back and upwards as it travelled through the substrate horizontally (Fürsich et al., 2017; Heinberg, 1973; Seilacher, 2007). The width of the G. comosa trail ranges from 3 to 6 mm and the length is approximately 40 cm. Gyrochorte is distinguished primarily by the vertical dimension of convex epireliefs and the corresponding hyporeliefs within the same bed (de Gibert & Benner, 2002). There are two ichnospecies of Gyrochorte, one having a simple bilobate horizontal burrow, that is, G. comosa, and the other is Gyrochorte variabilis (Fürsich et al., 2017) in which the bilobated traces are a fan like arranged. The observed specimen has elongated, sinuous bilobate ridges and furrows prominently on the upper bedding planes. Gyrochorte typically occurs in sandy facies of moderately energetic, near-shore and shallow marine environments (de Gibert & Benner, 2002).
Ichnospecies: Gyrophyllites geryonides von Huene, 1901
Radial trace fossils with numerous, more or less horizontal, swollen leaf-like lobes radiating from a vertical shaft having negative epirelief. The radiating lobes occur at one or several levels and are primarily unbranched (after Fu, 1991; modified by Uchman, 1998). Preservation of the radial lobes as negative epirelief suggests that they were filled with softer material. Gyrophyllites has been interpreted as fodinichnion (e.g., Fu, 1991; Fürsich & Kennedy, 1975) of a wormlike organism such as a polychaete or echiuran worm (Strzeboński & Uchman, 2015).
Ichnospecies: Nereites missouriensis (Weller, 1899)
Winding to regularly meandering, more or less horizontal trails, consisting of a median back-filled tunnel (core) enveloped by an even to lobate zone of reworked sediment (mantle) (Uchman, 2006). According to Chamberlain (1971), worm-like organisms generate Nereites by excavating lobes in front of them by backfilling and making central furrows as they advance.
Nereitabletes occur as shallow tier traces colonising soft substrates and are interpreted as feeding traces of deposit feeders. Ichnospecies of Nereites are differentiated partly based on the course of the burrow (Uchman, 1995). N. missouriensis (Weller, 1899) has a relatively unprogrammed course (Conkin & Conkin, 1968; Rindsberg, 1994; Wetzel, 2002). Seilacher (1974) and Chamberlain (1980) suggested that such differences in behaviour might represent the evolution of a lineage of trace makers. Unfortunately, the general course of the burrow, though significant, sheds scant light on the identity of its trace maker, such as whether it was propelled by a muscular foot, jointed legs, tube feet or other means of locomotion.
Ichnospecies: Ophiomorpha nodosa Lundgren, 1891
An intricate 3D network of sinuous burrow system preserved in full relief with Y-shaped branching. The outer wall has a knobby structure whereas the inner wall is smooth. The animal shapes, presses and smoothes these pellets into the burrow wall. It reinforces the burrow wall and makes sediment disposal easy. Individual cylindrical tubes are 10–30 mm in diameter, with oval cross-sections and around 100 mm long. O. nodosa burrows were formed in incohesive and loose sediment, which made the pelleted wall lining necessary. These are dwelling burrows of suspension-feeding crustaceans of shallow marine environments. The etiology of the Ophiomorpha tracemaker is complex and may be a variable combination of deposit and/or suspension feeding behaviours (e.g., Ekdale, 1992; Uchman & Gaździcki, 2006). Single pellet linings are characteristic of O. nodosa, whereas O. borneensis has a double-pellet wall (Frey et al., 1978; Leaman et al., 2015). Thalassinidean shrimps construct similar structures today (e.g., Frey et al., 1978). Ethologically, O. Nodosa probably represents a dwelling structure. At the ichnogenus level, Ophiomorpha is registered in a broad environmental range, from shallow-water deposits represented mainly by O. Nodosa to deep-sea environments represented mainly by O. rudis (Ksiazkiewicz, 1977; Tchoumatchenco & Uchman, 2001).
Ichnospecies: Palaeophycus tubularis Hall, 1847
Smooth-walled, unornamented, straight to sinuous cylindrical burrows, thinly but distinctly lined (Frey & Howard, 1970; Pemberton & Frey, 1982). Burrow fill is similar to the host rock. Cross-sections are circular to oval varying between 6 and 15 mm in diameter. The fill is structureless and passive. The ichnogenus Palaeophycus was first described by Hall in 1847. Five ichnospecies based on their morphology are (1) P. Striatus, (2) P. tubularis, (3) P. heberti, (4) P. alternatus and (5) P. sulcatus. According to Fillion and Pickerill (1984), the burrow morphology, that is, thin lining, gently curved and smooth walls, are consistent with P. tubularis. P. tubularis is a dwelling structure of facies-crossing form generated by polychaetes or annelids (Pemberton & Frey, 1982).
Ichnospecies: Phycodes palmatum Hall, 1852
Thick, rounded branches originating in a palmate or digitate form from nearly the same point (Fillion & Pickerill, 1990). It differs from P. Curvipalmatum by its large size and lack of re-curvature. Vermiform annelids are possible producers of Phycodes (Fillion & Pickerill, 1990).
Ichnospecies: Planolites beverleyensis Billings, 1872
(Plate 1, Figure 7)
Unlined, subcylindrical to cylindrical, unbranched burrows, parallel to oblique to the bedding with smooth walls, preserved as full reliefs or convex hyporeliefs and structureless burrow fills. The burrow diameter varies from 4 to 10 mm. P. beverleyensis differs from Planolites montanus in its curved to arcuate courses and more penetrative nature (Pemberton & Frey, 1982). Since Planolites (unlined) tunnels are considered temporary structures used for mobility and/or eating (fodinichnion), there is no need to reinforce the burrow wall. Planolites probably represent active sediment backfilling in an ephemeral burrow (Pemberton & Frey, 1982; Pemberton & Jones, 1988).
Ichnospecies: Rhizocorallium commune var. auriforme (Hall, 1843)
Rhizocorallium has different morphological forms, ranging from small, straight structures to very long, sinuous ones, which can be either planispiral or trochospiral. In addition, when the diameter of the burrow expands, the limbs can show a minor divergence in the distal region (away from the apertures), creating a pear- or fan-shaped structure. R. commune var. Auriforme are U-shaped spreiten-burrows, parallel or oblique to bedding planes. They lack retrusive or trochospiral. Short and commonly slightly inclined (10–20°) to bedding. Knaust (2013) suggested that the producers of R. commune var. auriforme are suspension and deposit-feeding polychaetes. These are common in middle-outer ramp settings, lagoons and tidal flats in Mesozoic shallow marine carbonate and siliciclastic deposits (Knaust et al., 2012; Pemberton et al., 2001, 2012; Rodríguez-Tovar et al., 2012).
Ichnospecies: Skolithos verticalis Hall, 1943
Unbranched, vertical to steeply sloped shafts, straight to slightly curved, cylindrical to sub-cylindrical, lined or unlined, with or without a funnel-shaped top, all indicate a Skolithos. The burrow is approximately 6–10 mm in diameter, vertical or slightly sloped, the burrow fill is homogeneous with the host rock, the burrow wall is distinct and smooth, and the burrow fill lacks structure. The specimens from the study are identical to those described by Alpert (1974) as S. verticalis, which is characterised by inclined, relatively short burrows compared to S. linearis. Skolithos verticalis burrows are shorter, smaller, more inclined and curved, and rarely crowded. Weathering leaves the burrow as a hole in the sediment. Skolithos are interpreted as dwelling burrows of suspension-feeding organisms (e.g., Patel & Desai, 2009); commonly occurring in high-energy, shallow marine, nearshore environments (Knaust, 2017).
Ichnospecies: Thalassinoides horizontalis Myrow, 1995
The three-dimensional burrow systems of Thalassinoides are characterised by the prevalence of smooth-walled, mainly cylindrical to elliptical tunnels of various dimensions with Y or a T shaped branching network lacking vertical shafts. Thalassinoides, typically thicken at the points of branching (e.g., Howard & Frey, 1985). T. saxonicus (Geinitz), T. ornatus (Kennedy), T. Paradoxicus (Woodward), T. suevicus (Rieth) and T. Horizontalis (Myrow) are the five most often accepted ichnospecies (Kim & Pickerill, 2003). T. horizontalis differs from other ichnospecies in having a dominant horizontal polygon network and a lack of vertical shafts. Thalassinoides is a facies-crossing ichnotaxon commonly found in oxygenated marine environments and soft and firm sediments (e.g., Ekdale et al., 1984; Rodríguez-Tovar et al., 2008; Schlirf, 2000).
Ichnospecies: Taenidium serpentinum Brady, 1947
(Plate 1, Figure 8)
Cylindrical, unbranched, thinly lined, simple, parallel to inclined to bedding, sinuous burrow with meniscate fills. The burrow diameter is 10 to 17 mm. The burrow contains thick meniscate segments. The fill and host material may be identical or contrasting. The menisci are approximately 1–3 mm thick, with inter-meniscal segments being much wider (8–13 mm thick) and feebly concave. Taenidium is interpreted as produced by vagile, deposit-feeding animals (D’Alessandro & Bromley, 1987). During the Jurassic, the ichnospecies occurred under a broad range of energy conditions in Kachchh, varying from shallow marine, storm-influenced environments (Fürsich et al., 2018) to a low-to-intermediate energy ramp (Fürsich, 1998) to inner shelf/prodelta environments (Desai et al., 2008).
Ichnospecies: Teichichnus rectus Seilacher, 1955
Unbranched, straight or slightly sinuous, irregular, horizontal burrow. Its diameter ranges from 6 to 11 mm and is broadly U-shaped. The burrow appears to consist of parallel, more-or-less horizontal to undulating laminae in the lateral view, producing a retrusive spreite structure with a tube on top. Laminae are simple and convex, while tubes may be cylindrical or elliptical in cross-section. Burrow fill is identical to the matrix. The described forms lie horizontally on the bedding plane. Based on the morphology of the laminae, T. rectus can be used to categorise the current specimens. Five species of Teichinchnus are present. The current species T. rectus is a retrusive, spreiten burrow (Seilacher, 1955). It differs from the spiral burrow of T. flexuosus, branched structure of T. palmatus and the double gutter-shaped lamina morphology of T. Patens (Schlirf, 2000).
Ichnospecies: Zoophycos villae Massalongo, 1855
The endichnial burrows are fan-shaped, comprising radiating arcuate laminae and branching at an acute angle. Few specimens show bulging at the end. The length of the structure is 25 cm and comprises spreiten structures comprising J- or U-shaped burrows of varying length and width. The spreiten form laminae, surrounded by a marginal tube, spirally coiled around a central axis, and lined with primary and secondary lamellae. The laminae exhibit characteristic backfill structure during marginal tube lateral displacement (Häntzschel, 1975; Olivero, 2003; Uchman, 1999). The laminae exhibit characteristic backfill structure during marginal tube lateral displacement (Häntzschel, 1975; Olivero, 2003; Uchman, 1999).
Ichnoguilds
The Yellow Flagstone Member displays relatively simple tiering structures. The shallow tier IGs are densely bioturbated, and the mottled background indicates deposition in transitional offshore facies.
The shallow-tier trace fossils Asterosoma, Gyrochorte, Planolites, Gyrophyllites and Nereites are cross-cut by mid-tier Rhizocorallium, Arenicolites, Diplocraterion, Teichichnus and Zoophycos and deep tier Skolithos, Arenicolites, Thalassinoides and Ophiomorpha. Ten ichnoguilds are identified and are described below.
Shallow-Tier Ichnoguilds (Plate 1)
Asterosoma IG: This monospecific IG comprises stationary, shallow-tier, horizontal to inclined burrows of deposit-feeding worm-like organisms. This IG is present only in the Simriwandh section in high abundance. The Asterosoma IG indicates nutrient-rich substrates deposited during stable low-energy periods (Pemberton et al., 2001).
Nereites IG: The Nereites is a shallow-tier, feeding trace fossil present in high density in the Andhao section with Taenidium. Complex grazing traces, such as Nereites, indicate a quiet but oxygenated environment (Pemberton & MacEachern, 1995).
Gyrochorte IG: This IG is dominated by horizontal, crawling and feeding structures, such as Gyrochorte, Planolites and Palaeophycus, respectively. It is found in the Simriwandh section with Rhizocorallium. Gyrochorte typically occurs in sandy facies of moderately energetic, near-shore and shallow marine environments (de Gibert & Benner, 2002). Bioturbation is moderate to high, indicating relatively slow sedimentation and little physical reworking. The guild occurs in the lower shoreface, below the fair-weather wave base but above the storm-wave base, in quiet conditions.
Bolonia-Taenidium IG: This IG is dominated by horizontal, straight, unbranched, bilobate trace fossil B. lata and T. serpentinum and G. geryonides. This IG is present only in the Andhao section. B. lata (Schlirf, 2000) are ribbon structure, heart-shaped outlines in cross-section with positive epirelief and T. serpentinum have well-spaced, arcuate menisci in the form of a serpent with sharp boundaries and unlined (Keighley & Pickerill, 1994). The trace fossil was most likely created by the lateral journey of an infaunal deposit-feeder through the sediment. The palaeoenvironment from which B. lata has been reported so far is nearshore marine, influenced by storm waves. Taenidium is found from shallow marine, storm-influenced environments (Fürsich et al., 2018) to a low-to-intermediate energy ramp (Fürsich, 1998) to inner shelf/prodelta environments (Desai et al., 2008).
Planolites IG: This IG is characterised by a palaeo-community of mobile, shallow to mid-tier, subsurface deposit-feeding vermiform organisms. The intricate behaviour of deposit feeders, which simultaneously seek food and shelter, makes the present IG. It is dominated by the actively filled burrow of P. beverleyensis and semi-permanent, middle-tier, passively filled lined, suspension feeder or predator structures produced by vagile vermiform organisms burrows attributed to P. tubularis. Vagile deposit-feeder organisms mainly produce this shallow-to-mid-tier IG in flagstone and siltstone beds, presenting low ichnodiversity and moderate to high density in the Simriwandh and Andhao sections. This IG indicates the prevalence of low to moderate hydrodynamic energy conditions, primarily in lower shoreface to transitional offshore settings.
Shallow-tier trace fossil from the Yellow Flagstone Member (Goradongar Formation), Pachchham Island; 1. (i) Asterosoma ludwigae (As. i) and (ii) Asterosoma radiciformis (As. ii); 2. Phycodes palmatus (Py), Planolites (Pl) and Skolithos (Sk); 3. Bolonia lata (Bl) 4. Gyrophyllites geryonides (Gp); 5. Nereites missouriensis (Ne); 6. Gyrochorte comosa (Gy); 7. Planolites beverleyensis (Pl); 8. Taenidium serpentinum (Te).

Middle-Tier Ichnoguilds (Plate 2)
Rhizocorallium IG: This IG is composed of abundant species of Rhizocorallium. This IG is present in all the four studied sections of Yellow Flagstone Member, but in the Simriwandh section, the density and abundance are highest. Shallow burrowing deposit feeders dominate this IG in thinly bedded flagstone and calcareous silty shale. The most abundant species is R. commune var. irregulare. Other feeding structures observed include Palaeophycus, Planolites, Thalassinoides and Skolithos. As proposed by Knaust (2013), the producers of R. commune var. irregulare are deposit-feeding polychaetes. Rhizocorallium reflects periods of minimum disturbance and probably a high influx of organic matter within the sediment (Fürsich & Schmidt-Kittler, 1980). This IG indicates marine conditions and possibly a sediment-feeding mode of life in the well-lit, clear water, photic zone of the lower shoreface to offshore (Patel & Patel, 2015).
Zoophycos IG: This IG is dominated by middle-tier, fan-shaped trace fossils Z. villae and T. rectus. They are present in high-density bioturbated beds in the Andhao section. The Zoophycos producer fed at the sediment-water interface and excreted faecal pellets at depth to maintain a food source for periods of scarce nutrient flux (Bromley, 1991; commonly being associated with quiet-water conditions and dysoxia; Löwemark & Schäfer, 2003; MacEachern et al., 2012; Martin, 2004). It shows the dominance of permanent deposit-feeding strategies and is preserved in stable offshore settings and storm-dominated lower shoreface to transitional offshore.
Arenicolites-Diplocraterion IG: This IG is made up of the middle-tier, vertical U-shaped burrows, Arenicolites and Diplocraterion. The Arenicolites are oriented vertically to incline to the bedding plane and lined burrows without spreite. The arms of the U-tubes are straight or curved and parallel to each other, interpreted as the work of suspension-feeding polychaetes (Fürsich, 1974a; Hakes, 1976; Patel & Desai, 2009). They are crustacean-like creatures living and feeding structures (Goldring, 1962). Arenicolites are distinguished from Diplocraterion by the absence of spreiten (Fürsich, 1974a). Diplocraterion parallelum are unlined, with retrusive spreite, burrow fill and spreite sediments similar to the host sediments. The burrows are probably made by annelids or crustaceans for permanent domicile purposes at distinct stratigraphic levels, suggesting the opportunistic behaviour of tube-dwelling suspension-feeding organisms (Dam, 1990). This IG represents the behaviour of suspension-feeding organisms in argillaceous to sandy shifting substrates of the upper shoreface environment.
Middle-tier trace fossil from the Yellow Flagstone Member (Goradongar Formation), Pachchham Island; 1. Rhizocorallium Commune var. irregulare (Rh); 2. Highly bioturbated bed of Rhizocorallium (Rh) in Simriwandh section; 3. Bioturbated bed with Planolites (Pl), Palaeophycus (Pa) in Andhao section; 4. Zoophycos villae (Zo) in Andho section; 5. Bioturbated Rhizocorallium (Rh) with Skolithos (Sk) and Arenicolites (Ar); 6. Arenicolites (Ar) with Thalassinoides (Th); 7. Diplocraterion parallelum (Di); 8. Bioturbated bed in Simriwandh with Rhizocorallium (Rh), Skolithos (Sk), Phycodes (Py), Planolites (Pl) and Palaeophycus (Pa).

Deep Tier (Plate 3)
Skolithos IG: This IG is characterised by deep-tier, suspension-feeding organisms. This IG comprises S. linearis, isp and O. nodosa. S. linearis is unbranched, vertical to steeply sloped, straight to slightly curved, cylindrical to sub-cylindrical and lined (Schlirf & Uchman, 2005). This IG suggests a prevalence of high hydrodynamic energy conditions during the colonisation of foreshore and shoreface settings (Frey, 1990). The ichnodiversity is low. The uniform burrow diameters range from 3 to 20 mm, and the fill is mostly structureless, medium sand, similar to the host rock.
Thalassinoides-Ophiomorpha IG: This IG is made up of three-dimensional systems of lined burrows with smooth burrow walls of T. horizontalis. Burrow branching is irregular to dichotomous, typically Y- and T-shaped in a horizontal plane and enlarges at bifurcations. Crustaceans (Frey et al., 1984) constituted these burrows, which serve as combined dwelling and feeding burrows. O. nodosa is full relief, vertical shafts and interconnecting horizontal-to-inclined tunnels forming three-dimensional, branching burrow networks. The burrow walls are heavily reinforced, and the burrow fill resembles the host sediment commonly colonising shallow marine environments.
Deep-tier trace fossil from the Yellow Flagstone Member (Goradongar Formation), Pachchham Island; 1. Deep-tier Skolithos (Sk), Diplocraterion (Di) and Arenicolites (Ar) with shallow tier Planolite (Pl) and Palaeophycus (Pa) in Simriwandh Section; 2 and 3. Highly bioturbated bed in Sadhara and Andhao sections; 4. Ophiomorpha Nodosa (Op) from Simriwandh Section; 5. Teichichnus rectus (Ti) with Thalassinoides (Th) in Dattratreya section.

Tiers, Diversity and Density of the Trace Fossils in the Yellow Flagstone Member
The Middle Bathonian Yellow Flagstone Member reflects the lower shoreface to offshore transition environment (Figures 4, 5). The rocks of this member are exposed in four sections on Pachchham Island and correspond to a shift towards offshore during transgression. The basal beds with trace fossils are found in the Dattatreya section, followed by the youngest beds in the Simriwandh section.
In the Dattatreya and Sadhara sections, older beds of the Yellow Flagstone Member are exposed. These rocks show bioturbation of middle and deep-tier trace fossils (Skolithos, Diplocraterion, Arenicolites, Palaeophycus and Planolites) belonging to the Skolithos ichnofacies. These are low in diversity and show the adaptive behaviour of suspension-feeding organisms. Towards younger beds, there is a general increase in density, diversity and suspension-feeding habits. Andhao and Simriwandh section show rocks with shallow to middle-tier trace fossils (Asterosoma, Rhizocorallium, Thalassinoides, Palaeophycus, Planolites, Phycodes, Taenidium and Nereites) belonging to the Cruziana ichnofacies. These sections correspond to nutrient-rich substrates in low-energy environments. The predominance of Asterosoma, Nereites and Zoophycos IGs attests to optimum marine conditions. The predominance of shallow-tier IGs corresponds to stable low-energy periods in an oxygenated environment in the photic zone of the lower shoreface to offshore (Patel & Patel, 2015).
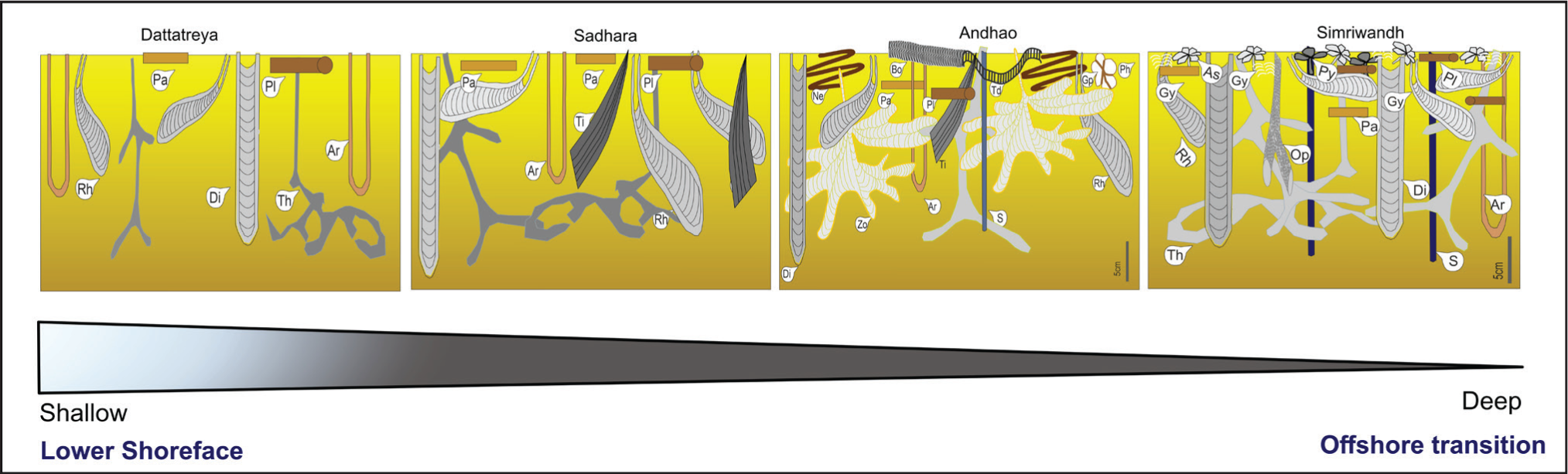
Ichnoguilds of the Yellow Flagstone Member in the studied sections and their distribution of depositional settings. Ichnoguilds are arranged along a shallowing-deepening trend (below).
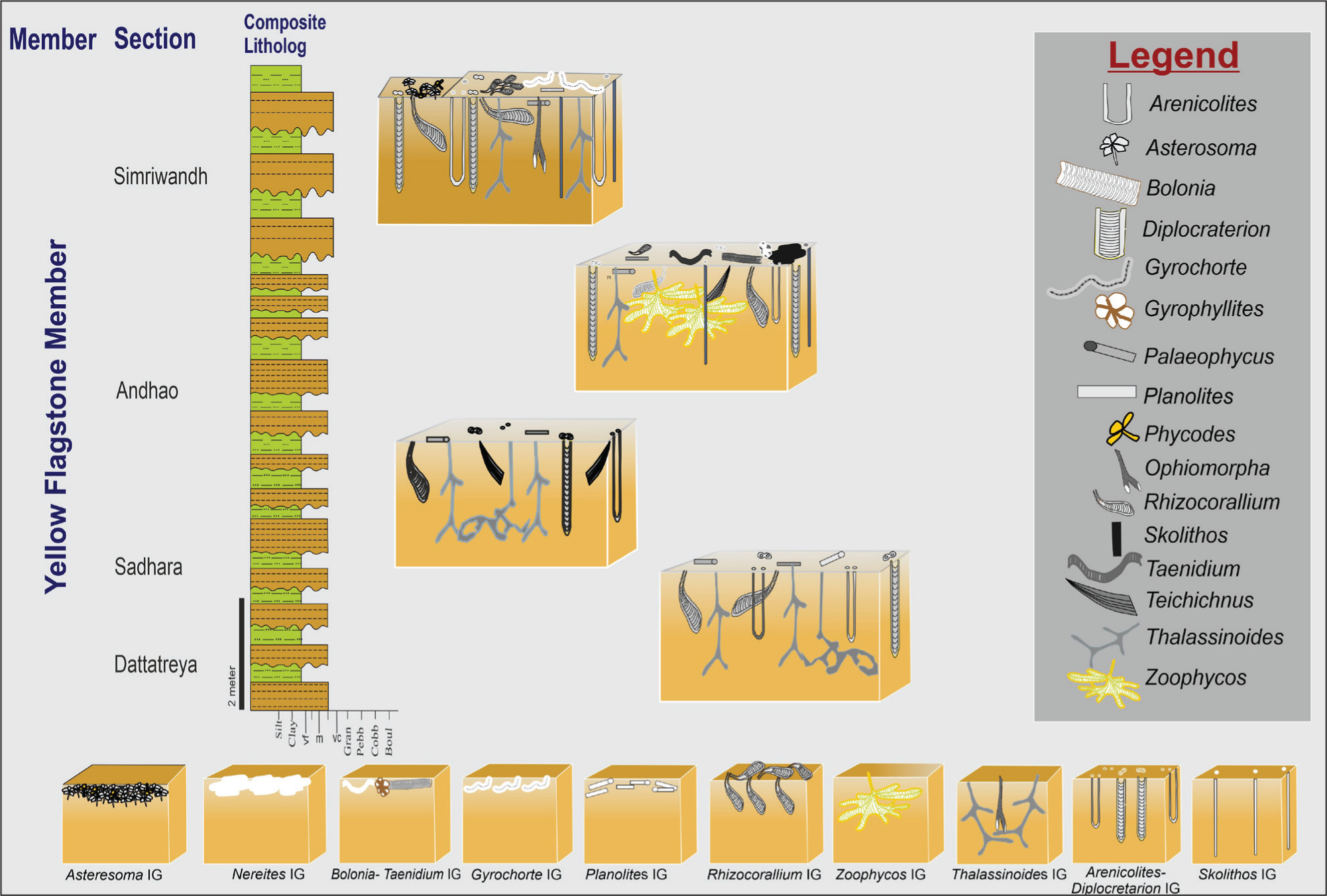
Composite Ichnoguild showing variation from lower shoreface to offshore transition in Yellow Flagstone Member.
The trace fossil distribution in the Bathonian Yellow Flagstone Member unravels the palaeoecological controls in the offshore transition environment in the transgressing sea. For a better understanding, the trace fossil tiering, diversity and density are plotted against the lithostratigraphy of the Member (Figure 6).
Trends in tiering, diversity and density of the trace fossil in the Yellow Flagstone Member.
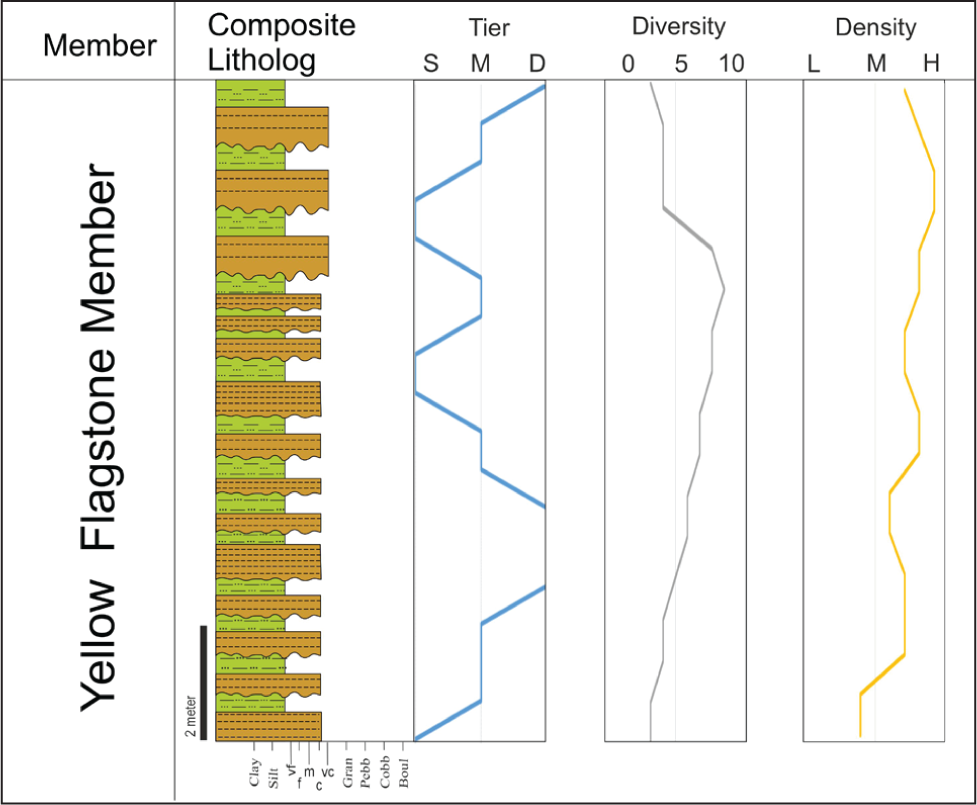
Tiering suggests that infaunal communities ranging from simple to complex dominate. The gradient intensified and varied more offshore. The older succession of the member shows low diversity and suspension feeding lifestyles, with the predominance of the Thalassinoides-Arenicolites, Diplocraterion and Rhizocorallium IGs in the Dattatreya temple, and Sadhara dome sections representing more distal settings. Slowly, due to transgression, the sediments shift from high to moderate energy with storm-dominated lower shoreface to the well-oxygenated transitional offshore. The ichnofossils representing the offshore transition are deposit feeders with high density and diversity in the Andhao and Simriwandh sections. These two sections show shallow to middle-tier IG. The Asterosoma IG, Gyrochorte IG, Planolites IG, Gyrophyllites IG and Nereites IG represent the shallow tier whereas the Rhizocorallium IG, Arenicolites- Diplocraterion IG and Zoophycos IG are Middle tier.
The diversity and density of trace fossils indicated low diversity and density in the Dattatreya section but it attains maximum diversity in the Andhao and then the diversity again decreases in the Simriwandh section. Similarly, the density of trace fossils is lowest in the Dattatreya section and increases up-section. Additionally, the younger bed of the succession at Simriwandh shows a decrease in the density and diversity and deep-tier IG because during this time the transgression stopped and the overlying Gadaputa Sandstone Member shows a gradual change from flagstone to sandstone. This shift of facies is well reflected by the trace fossil data.
Conclusions
The trace fossil distribution and IG analysis of the Yellow Flagstone Member offer evidence of a transitional offshore depositional environment during transgression. Our data support the earlier sedimentological and sequence stratigraphical interpretation of the Yellow Flagstone Member. The offshore transitional environment exhibits more variability due to the alternations of severe storm events and periods of calm weather. The offshore transition occurs under the fair-weather wave-base. The composition of trace fossils undergoes visible alterations along an onshore-offshore transect. The shallower onshore area is characterised by a lower diversity and an increased abundance of traces left by suspension feeders. Conversely, in the offshore area, there is a greater abundance of deposit-feeders exhibiting a higher level of diversity. The environmental conditions from the lower offshore to offshore areas display more pronounced fluctuations due to the alternation of intense storm events and calm periods. The trace fossils belong to the Cruziana-Skolithos ichnofacies, which differ in composition and affect the abundance of ichnotaxa and the degree of sediment disturbance by organisms.
Footnotes
Declaration of Conflicting Interests
The authors declared no potential conflicts of interest with respect to the research, authorship and/or publication of this article.
Funding
The authors received no financial support for the research, authorship and/or publication of this article.