
Abstract
The palaeobiogeography and age of the upper calcareous part of the Nimar Sandstone (NS), Bagh Group is determined based on ostracod fossils. The rocks of the NS directly rest over the Precambrian basement rocks outcropping near Pipaldehla, Gola Chhoti (Sajwani), Dhekal Badi, Ranapur, Udaygarh (Kanas) and multiple locations in Jhabua District. The present paper is the first report on ostracod fauna from Pipaldehla (N 22° 46’ E 74° 39’), Jhabua District, Madhya Pradesh, India. A total of 15 species of ostracods belonging to 11 genera have been identified. The assemblages of ostracods from the western and eastern exposures of the Bagh Group have been compared. The overall ostracod fauna suggests an early Turonian age and shows affinity to the ostracod assemblage of (1) the lower part of the Nodular Limestone of the Bagh-Jeerabad area, Dhar District, Madhya Pradesh (eastern exposures), and (2) also with the westernmost exposures of the Kawant-Rajpipla area of Gujarat.
Introduction
The Upper Cretaceous Bagh Group of rocks is popularly known as Bagh beds. The Bagh Group of rocks, composed of thick siliciclastic and calcareous sediments, were deposited during the late Cretaceous (late Cenomanian-Coniacian) global sea-level rise in a shallow epicontinental sea or an embayment that invaded the western and central parts of India from the west along the Narmada valley (Figure 1) (Bhattacharya et al., 2020; Jaitly & Ajane, 2013; Keller et al., 2021; Kumari et al., 2020; Saha & Shukla, 2022; Saha et al., 2010; Singh, 1981; Singh & Srivastava, 1981). The Bagh Group comprises three formations, which in ascending order are: Nimar Sandstone (NS), Nodular Limestone and Coralline (Bryozoans) Limestone (Jaitly & Ajane, 2013; Tripathi, 2006). Bose (1884) subdivided the NS into the ‘Lower Member’ consisting mainly of conglomerate, sandstone and carbonaceous clay, and the ‘Upper Member’ comprising calcareous sandstone with marine fossils.
(A) Inset map of India showing the distribution of the Deccan Traps and the Precambrian rift system in India; Abbreviations are as follows: NR = Narmada Rift; GR = Godavari Rift; MR = Mahanadi Rift; (B) Distribution of Bagh Group, Lameta Formation, Deccan Traps and Inter-trappean sequences (modified from Kumari et al., 2020).
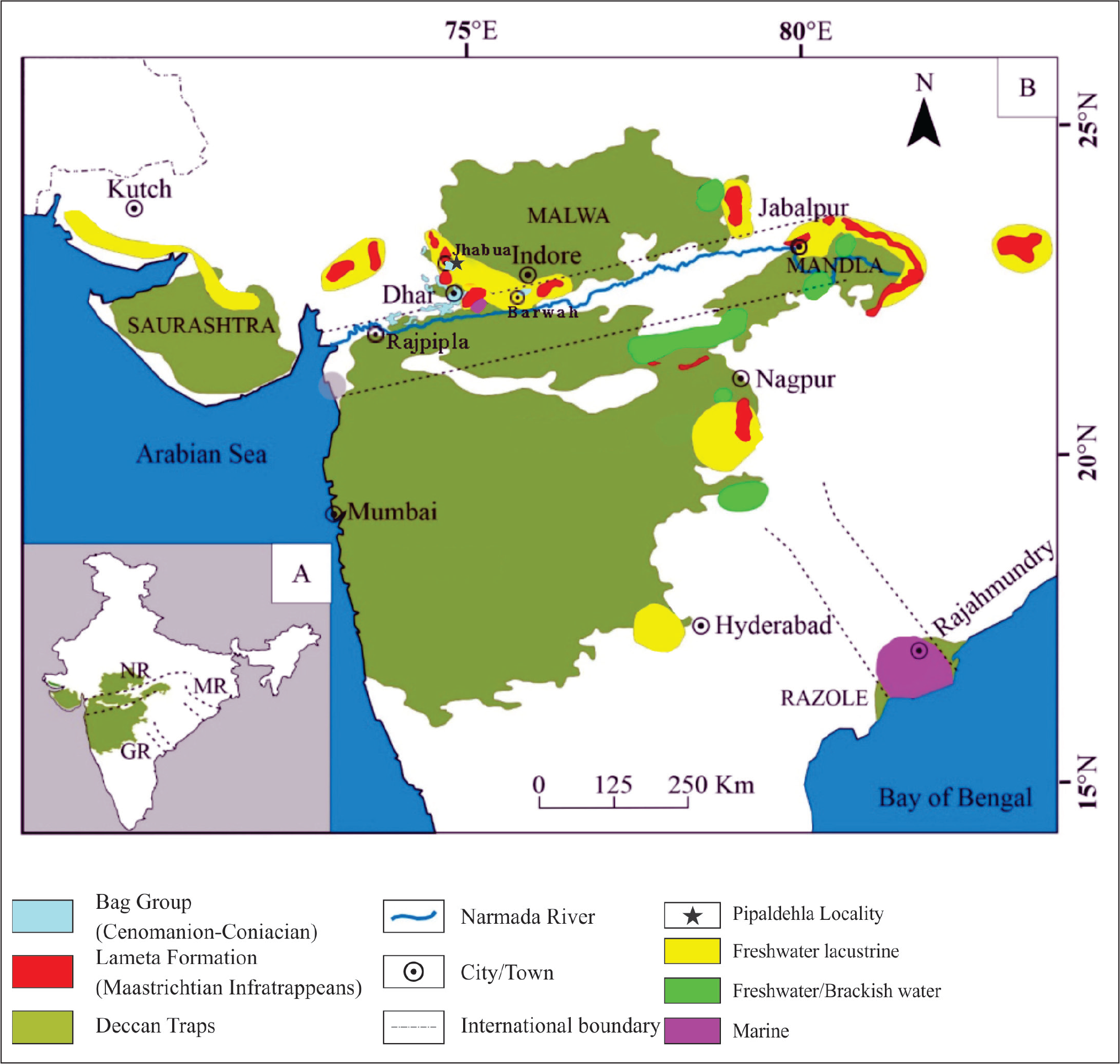
At Pipaldehla, NS directly overlies the Precambrian basement, while the overlying sediments (Upper Cretaceous) occur as a linear belt of restricted distribution. To enhance our knowledge of the ostracods of the NS of the Bagh Group, samples were collected from different sections of Pipaldehla (N 22° 46’ E 74° 39’), Ranapur, Dhekal Badi, Gola Chhoti and Udaygarh (Kanas) by the authors during fieldwork in Jhabua District. However, in the present work, the authors first reported the presence of ostracods from the northernmost exposure of the NS at Pipaldehla village about 7 km east of Jhabua town (Figure 2). A total of 15 taxa were recorded, out of which six are common with ostracods reported earlier from the late Cenomanian of the Kawant-Rajpipla area (Chaudhary & Nagori, 2019; Keller et al., 2021), while 14 taxa are common with ostracods of the Bagh-Jeerabad area (Chaudhary & Nagori, 2019). Based on the ostracods age and palaeobiogeography are also discussed.
Lithostratigraphy and photo illustrations of key lithologic horizon of the studied Pipaldehla section, Bagh Group, Jhabua District, Madhya Pradesh, India.

Previous Work
The top part of NS is calcareous and full of megafossils, mainly bivalves, gastropods, echinoids and microfossils viz., calcareous algae, rare foraminifers and ostracods. It is worth noting that ostracods from the Bagh Group of eastern and western exposures of different localities were previously described and recorded by Jain (1961, 1975b), Roy Chowdhary and Sastri (1962), and Guha and Ghosh (1970). Recently, ostracods from these beds were described and illustrated (Chaudhary, 2017; Chaudhary & Nagori, 2019; Chaudhary et al., 2017a, 2017b, 2019; Keller et al., 2021). So far studies on calcareous algae are concerned, considerable work has been carried out in the past. Badve and Nayak (1983) first time reported five species of the algal genus Halimeda from this locality. Afterwards, Badve and Nayak (1984) recorded nine species of algae and coined the term Jhabotrigonia–Turritella bed for the calcareous top part of NS at Pipaldehla.
Sanganwar and Kundal (1996) recorded 10 species of the melobesioidean calcareous algae from the same locality. Kundal and Sanganwar (1998), while dealing with calcareous algae from the calcareous top of the NS, recorded 72 species belonging to Cyanophyta, Rhodophyta and Chlorophyta. Based on these algal assemblages they discussed the stratigraphical, palaeogeographical and palaeoenvironmental significance and assigned Cenomanian-Turonian ages to this lithounit of Bagh Group and late Albian for part of NS at the base. Further, they concluded that the calcareous top of the NS might have been deposited in tropical waters at a depth of 10–12 m from below low tide level in moderate energy setting and turbulence with normal salinity of water.
Foraminiferal studies from this locality were carried out by Nayak (1987) and first time recorded following taxa: Miliola aff. Miliola saxorum (Lamark) and species of the genus Lenticulina, Pararotalia, Alabamina and Gavelinella and shown their Albian-Cenomanian-Campanian affinity. Foraminifers from other parts of the Bagh Group were recorded (Jain, 1961; Roy Chowdhary & Sastri, 1962; Sharma, 1976; Singh, 1950). However, during the last two decades of the twentieth century, considerable detailed work was carried out (Rajshekhar, 1982, 1984, 1987, 1991, 1995; Rajshekhar & Atpalkar, 1995) from the eastern and western exposures of the Bagh Group.
Evolution of the Basin
During the late Cretaceous (late Cenomanian-Turonian-Coniacian) marine incursion, which influenced farther inland regions (Rajpipla, Gujarat, up to Barwah near Indore, Madhya Pradesh), several hundreds of kilometres into central India. This transgression is linked with the breakup of Greater India from Madagascar ~90 Ma ago (Kumari et al., 2020). This late Cretaceous dispersal event was due to spreading along the central Indian Ridge, which opened up the Mascarene Basin (Chatterjee et al., 2013). The southward drift of Madagascar concerning the Indian subcontinent exposed the western coast of India. This breakup is associated with Marion plume activity that resulted in the eruption of the Mahajanga Flood Basalt, reactivation of the Narmada fault and subsequently opening of the Narmada Basin and allowed seawater to inundate the pericratonic rifts (Biswas, 1987, 1999, 2003; Jafar, 2016). The Narmada rift had been reactivated and opened due to the extensional tectonics (separation of Madagascar) at that time (~90 Ma) and acted as the pathway for this transgression. The average sea level was higher than the present-day mean sea level throughout the Cretaceous. It reached a maximum of ~250 m at the beginning of Turonian between 93.9 and 91.6 Ma (Gradstein & Ogg, 2020; Haq, 2014). The temporal coincidence of tectonics as a result of the separation of Madagascar from India, and the subsequent global sea-level rise, allowed seawater to transgress onto the Indian subcontinent during the late Cenomanian (Jafar, 2016; Keller et al., 2021). Oyster beds, the calcareous top part of Nimar Sandstone Formation (NSF), Nodular Limestone and Coralline (Bryozoan) Limestone Formations of the Bagh Group are well-preserved evidence of transgression during late Cenomanian-Turonian-Coniacian age (Chaudhary & Nagori, 2019; Keller et al., 2021; Ruidas et al., 2018).
The Bagh Group of rocks rests over Precambrian Basement rocks of Lower Proterozoic to Archean ages. The continental sediments, NSF overlying the Precambrian basement, cover a very large part of the area, while the overlying marine sediments show restricted distribution forming linear belts and were deposited in an intracratonic rift basin (Biswas, 1987; Kumar et al., 2019; Tripathi, 2006) following the Precambrian tectonic grain. Three separate marine basins in a linear belt were identified by Tripathi (1995). The ENE-WSW trending basins are the Bagh-Jeerabad basin (Madhya Pradesh) and Kawant basin (Gujarat) while the third Jobat-Dohad basin follows the NE-SW Aravalli trend.
The distribution and thickness variation of marine strata across the basins suggests that these basins were probably half grabens, where southern or southwestern faults were active (Tripathi & Lahiri, 2000). The reactivation of Pre-Cambrian tectonic grain during the anticlockwise movement of the Indian plate at the Cretaceous Period resulted in the formation of tectonic basins following the Precambrian trends and generated Cretaceous triple point junction (Tripathi, 1995, 2006; Tripathi & Lahiri, 2000).
Material and Methods
A total of five fresh samples, each of approximately 200 g, were collected at an interval of 2 feet from the 7-foot-thick hard calcareous sandstone (NS), which is full of molluscan shells. All the samples were processed for ostracods by standard procedure. About 100 g of each sample was soaked in water overnight, and a spoonful of sodium carbonate was added and boiled for more than two hours. The disaggregated samples were washed under a jet of water using 30, 72 and 150 B.S.S. mesh-size sieves. The ostracods were picked from the washed residue of a 72-mesh sieve under a stereo-zoom binocular microscope. Ostracod species were photographed under SEM (Zeiss-Gemini 300) at IIT Roorkee. All the illustrated specimens are deposited in the Micropaleontology Laboratory, Department of Geology, Mohanlal Sukhadia University, Udaipur, with repository number SUGDMF No. 1458–1472.
Age of the Bagh Group
These beds are very rich in invertebrate fauna, and on this basis, different ages have been assigned. The majority of earlier workers, including Duncan (1865, 1887), Vredenburg (1907, 1908), and Chiplonkar (1939a, b), assigned Cenomanian age based on echinoids, ammonites and lamellibranchs. Based on the above fossil groups (Bose, 1884; Fourtau, 1918; Mukherjee, 1934 in Fermor) assigned Albian to Senonian ages. Taylor and Badve (1995), based on the new bryozoan genus Chiplonkaria, concluded that Bagh Group sedimentation took place during the Turonian-Coniacian Period. Based on ostracods Jain (1961, 1975b), Chaudhary (2017), Chaudhary et al. (2017a, 2017b, 2019), and Chaudhary and Nagori (2019), it was concluded that the deposition of Bagh sediments lasted from late Cenomanian to Coniacian. However, foraminifers suggest an age ranging from Aptian to Maastrichtian, but tentatively Turonian as a dominant element for the Bagh Group (Rajshekhar, 1987, 1991, 1995). Nayak (1987) based on planktonic and benthic foraminifers of the calcareous top portion of NS assigned Albian-Cenomanian age. Further, while re-examining the collection of Nayak by Rajshekhar (1995), the presence of planktonic foraminifera genera like Hedbergella, Ticinella and Discorbis was found. So far benthic foraminifers are concerned they reveal the presence of Miliammina and Gavelinella. Based on the above fauna a middle Albian to late Cenomanian age has been proposed for the deposition of this lithounit.
Rajshekhar (1995) recorded 23 genera belonging to benthic and planktonic foraminifers from the Coralline Limestone Formation. On this basis, he assigned an age ranging from Aptian to Maastrichtian but Turonian was the dominant element. In the Jeerabad area (Man Valley) marl and Coralline Limestone overlies Nodular Limestone, which is very rich in mega as well microfossils (foraminifers and ostracods), suggesting a late Coniacian age (Chaudhary & Nagori, 2019).
On the other hand, Dassarma and Sinha (1975) described rich assemblages of lamellibranches, gastropods, ammonites, echinoids and shark teeth from eastern and western exposures of the Bagh Group. They concluded that different marine patches of the Bagh Group exposed along Narmada Valley are not of the same geologic age. According to them Bagh Group from Barwah to east of Alirajpur ranges in age from Cenomanian to Turonian, while the Group in the west of Alirajpur to Kawant–Rajpipla, which is lithologically different than that of the eastern part, shows younger ages, that is, Coniacian and ranging at least up to uppermost Cretaceous. The arenaceous series designated as NS also has a composite character regarding its origin and age. Badve and Nayak (1984) designated the term Jhabotrigona-Turritella bed for the calcareous top part of NS at Pipaldehla.
The environment of deposition predicted for the lower part of NS is fresh water (fluvial), whereas the upper calcareous sandstone represents a shallow marine setting (Fluvio-Marine estuarine deposit) (Bhattacharya et al., 2020; Bose & Das, 1986; Singh & Gosh, 1977; Singh & Dayal, 1979; Tripathi, 2006).
Palaeobiogeography
The disintegration of Gondwanaland and their separation into different parts initiated roughly ~180 Ma. During the late Jurassic (about 158 Ma), Madagascar and India rifted away from Africa and the movement was almost directly south and nearly parallel to the African coastline. Such genetic communication was still possible in shallow marine environments until the earliest Cenomanian, that is, about 100 Ma (Puckett et al., 2016).
During the early late Cretaceous, Africa, Madagascar and Indian-Seychelle plate remained as the major tectonic elements of the southern-central Tethys, with Africa having already been separated from a Madagascar-India-Seychelle ‘plate’. A rapid northward drift of India together with the Seychelle block may be due to extensional processes that occurred between 90 and 80 Ma and may have caused changes in the palaeoceanographical currents, which influenced the distribution of the ostracod faunas. Until about 90 Ma, India remained close to Madagascar, but during Campanian, seafloor spreading initiated between India and Madagascar via the Carlsberg Ridge resulted from the rapid movement of India away from Madagascar (Dingle, 1988; Luger, 2003; Puckett et al., 2016).
The Southern Gondwanaland oceans fostered a distinctive ostracod fauna that can be recognised in Bajocian-Cenomanian strata, that is, South Gondwana Fauna (Dingle, 1988). It contains numerous endemic forms, with the most characteristic species belonging to the Progonocytheridae. A major faunal change occurred between the late Cenomanian and early Turonian times, when new taxa migrated from the equatorial South Atlantic, via the Temperate South Atlantic, and ousted the extant South Gondwana Fauna.
Some palaeoceanographical and palaeogeographical changes, such as opening pole change at 105 Ma (early Albian), probably led to the deep-water passages forming along the line of the Falkland Plateau-Southern Africa fracture zone, allowing the dysaerobic temperate South Atlantic to be flushed by oxygenated waters, which favoured the appearance of new taxa and rapid distribution in the Albian, along the Southern margins of Gondwana (Ballent & Whatley, 2006). At the same time, some old or original element of the South Gondwana fauna such as Rostrocytheridea made their way to Madagascar, India, New Zealand and Australia. During the latest Jurassic-Berriasian, the opening of a shallow intermittent epicontinental seaway between southern South Africa and Southern Argentinian Patagonia favoured faunal interchange.
The genus Rostrocytheridea is strictly gondwanine in its distribution (Ballent & Whatley, 2007). Dingle (2009) reported one species of the genus, that is, Rostrocytheridea pukehouensis from a cool, outer shelf/upper slope at Pukehou, the latest Maastrichtian of New Zealand. In India, Rostrocytheridea appears in the Albian-Turonian of Jaisalmer Basin (Andreu et al., 2007; Singh, 1997), while in Narmada Basin it appeared in late Cenomanian time up to Coniacian and represented by six species (Chaudhary et al., 2019).
According to the available records, Rostrocytheridea was an opportunistic, retrothermal organism that existed in both cool and warm, normal salinity and shallow water environments. This is evident from the studies conducted by Ballent and Whatley (2007), and Dingle (2009). Furthermore, the presence of Rostrocytheridea in the Antarctic, which is typically a tropical region, and the co-occurrence of the thermophile Cytherelloidea Alexander, once again support the idea of warm climates in high-latitude regions during the late Cretaceous period. In the Bagh Basin, most of the Cytherelloidea species are endemic. However, the discovery of Cytherelloidea oudiapurensis Jain (1975b), which bears resemblance to Cytherelloidea sp. 1 described by Rosenfeld and Raab (1974) from the Judea Group (upper Cenomanian) of Israel, suggests the migration of at least a few species from one basin to another via shallow sea routes. The environmental range of Bairdoppilata extends from very shallow to very deep waters (Maddocks, 1969; Morkhoven, 1963), though they tend towards greater abundance in warm and shallow carbonate environments (Horne, 2005; Kornicker, 1961). Dingle (1984, 1996) considered the genus Makatinella to be endemic to the Aptian to Cenomanian marine deposits of South Africa. Andreu et al. (2007) reported species of the genus from the Turonian of the Jaisalmer Basin of northwestern India. Chaudhary (2017) reported three species viz. Makatinella bilthanaensis, Makatinella punctata and Makatinella thuatiensis (Jain) from the Bagh Group.
1. Cytherelloidea oudiapurensis Jain, SUGDMF No. 1458. A complete carapace, left valve view, scale bar = 100 µm; 2. Cytherelloidea thuatiensis Jain, SUGDMF No. 1459. A complete carapace, left valve view, scale bar = 100 µm; 3. Bairdoppilata sp., SUGDMF No. 1460. A complete carapace, right valve view, scale bar = 100 µm; 4. Ovocytheridea baghensis Chaudhary and Nagori, SUGDMF No. 1461. A complete carapace, right valve view, scale bar = 100 µm; 5. Perissocytheridea batei (Jain), SUGDMF No. 1462. A complete carapace, right valve view, scale bar = 100 µm; 6. Rostrocytheridea baghensis (Jain), SUGDMF No. 1463. A complete carapace, right valve view, scale bar = 100 µm; 7. Rostrocytheridea sp. cf. R. Cerasmoderma Ballent and Whatley, SUGDMF No. 1464. A complete carapace, left valve view, scale bar = 100 µm; 8. Rostrocytheridea sp., SUGDMF No. 1465. A complete carapace, right valve view, scale bar = 100 µm; 9. Neocytherideis reymenti Jain, SUGDMF No. 1466. A complete carapace, right valve view, scale bar = 100 µm; 10. Curfsina? hanumapuraensis Chaudhary and Nagori, SUGDMF No. 1467. A complete carapace, left valve view, scale bar = 100 µm; 11. Haughtonileberis thuatiensis (Jain), SUGDMF No. 1468. A complete carapace, right valve view, scale bar = 100 µm; 12. Makatinella punctata Chaudhary, SUGDMF No. 1469. A complete carapace, right valve view, scale bar = 100 µm; 13. Makatinella thuatiensis (Jain), SUGDMF No. 1470. A complete carapace, right valve view, scale bar = 100 µm; 14. Veeniacythereis raoi (Jain), SUGDMF No. 1471. A complete carapace, left valve view, scale bar = 100 µm; 15. Nigeroloxoconcha baghensis Chaudhary and Nagori, SUGDMF No. 1472. A complete carapace, left valve view, scale bar = 100 µm.

The genus Nigeroloxoconcha is a typical common constituent of South Tethyan faunas (Gondwanian), occurring in Africa and South America. It appears in the Aptian of Morocco and evolves from the Albian to Miocene in Morocco, Algeria, Tunisia, Gabon, Niger, Ivory Coast, Nigeria, Egypt and Libya (Viviers et al., 2000). Species are also known in the Cenomanian-Santonian of Brazil (Piovesan et al., 2013). Andreu et al. (2007) reported Nigeroloxoconcha sp. from the Turonian of Jaisalmer Basin. Chaudhary and Nagori (2019) reported two species of the genus from the Bagh Group of Madhya Pradesh, India.
The records of Rostrocytheridea and other ostracod genera provide clear evidence of links between Western Australia, Argentina, the Antarctic Peninsula, South Africa, Madagascar and India during the late Cretaceous. The Cenomanian-Turonian species were common to the Jaisalmer Basin, Narmada Basin (Bagh Group) and the southern margin of the Tethys enable us to link up the Jaisalmer Basin, West Rajasthan shelf and Narmada Basin to the ‘North African-Middle East palaeobiogeographical province, established during the early Cretaceous and extending to the South-Tethyan margin in the Cenomanian, as defined by Andreu (1993), or to the South Tethyan ostracod province proposed by Luger (2003). Andreu et al. (2007), name a new province the Cenomanian-Turonian ‘South Tethyan ostracode province’, incorporating at one time the North African, Middle East Province, India, Madagascar, South Africa and South America.
During the early Turonian, just after the end of the Oceanic Anoxic Event 2 (OAE2), the taxonomic diversity decreases, while the level of oxygenation and the palaeoproductivity increase (along with the supply of nutrients); the water becomes hypoxic in the outer ramp environments that spread over the whole platform. The climate becomes hot and humid (Andreu et al., 2013). At Pipaldehla, ostracods are rare and non-specialised, associated with calcareous algae, benthic foraminifers and lamellibranches.
In the late Cenomanian-early Turonian marine communication is relatively easy between the carbonate platforms of the Kawant–Bilthana area with that of the Jhabua and Bagh–Jeerabad area, with the number of ostracod species in common ranging from 5 to 11, respectively.
Systematics
Subclass OSTRACODA Latreille, 1806
Order PODOCOPIDA Müller, 1894
Suborder OSTRACODA Latreille, 1806
Family CYTHERELLIDAE Sars, 1866
Genus Cytherelloidea Alexander, 1929
Cytherelloidea oudiapurensis Jain, 1975b, pp. 193–194, pl. 1, figs. 6a–b; text fig. 1.
Cytherelloidea oudiapurensis Jain; Chaudhary et al., 2017b, p. 220, pl. I, figs. 5–6.
Cytherelloidea oudiapurensis Jain; Chaudhary and Nagori, 2019, p. 522, pl. 1, fig. 7.
Description: Carapace subovate in lateral view; the species is ornamented by strong anterior and posterior rib; posterodorsally posterior rib continues into an inner sinuate rib below dorsal margin, this inner rib makes a hair pin bend and a very short rib from the dorsal part of inner loop bends downwards; one more short rib present in between ventral part of two inner loop and ventral margin.
Age range: Late Cenomanian to Turonian.
Cytherelloidea indica Jain, 1961, p. 341.
Cytherelloidea thuatiensis Jain, 1975b, p. 196, pl.1, figs. 10a–c, text fig. 4.
Cytherelloidea thuatiensis Jain; Chaudhary et al., 2017b, p. 222, pl. II, figs. 6–7.
Cytherelloidea thuatiensis Jain; Chaudhary and Nagori, 2019, p. 522, pl. 1, figs. 11.
Description: Carapace elongate, ovate in lateral outline; valve surface ornamented with spiral rib with strong anterior, ventral and posterior parts of the outer loop and dorsal and ventral parts of the inner loop; the rib starts from the anterodorsal margin.
Age range: Late Cenomanian to Coniacian.
Suborder PODOCOPA Sars, 1866
Superfamily BAIRDIACEA Sars, 1888
Family BAIRDIIDAE Sars, 1888
Genus Bairdoppilata Coryell et al., 1935
Description: Carapace elongate, subtrapezoidal in lateral outline; dorsal margin straight in middle, otherwise gently sloping anterodorsally and posterodorsally; overlap distinct, left valve larger than right.
Age range: Early Turonian.
Superfamily CYTHERACEA Baird, 1850
Family CYTHERIDEIDAE Sars, 1925
Subfamily CYTHERIDEINAE Sars, 1925
Genus Ovocytheridea, Grekoff, 1951
Ovocytheridea baghensis Chaudhary and Nagori, 2019, p. 524, pl. 2, figs. 6–9.
Description: Carapace subovate in lateral view, dorsal margin arched/convex, sloping down posteriorly; anterior margin broadly rounded, posterior margin acuminate and projected downward. Left valve larger than right, overlapping all along the margin. Surface of each valve smooth.
Age range: Early Turonian to late Turonian.
Genus Perissocytheridea Stephenson, 1938
Brachycythere batei Jain, 1975b, p. 200, pl. 3, figs. 31a–b, 32a–b.
Majungaella? batei Jain; Andreau et al., 2007, pl. 9, fig. 18.
Majungaella? batei Jain; Chaudhary and Nagori, 2019, p. 526, pl. 3, fig. 8.
Description: Carapace elongate, subrectangular in lateral outline. Dorsal margin straight, sloping down posteriorly; ventral margin convex in middle, otherwise straight. Anterior margin broadly rounded, posterior margin narrowly rounded. Surface ornamented with transverse ribs in anterior, median and posterior regions, while longitudinal ribs occur in ventral part and inner costal area pitted.
Age range: Early Turonian and Coniacian.
Genus Rostrocytheridea Dingle, 1969
Macrocypris? baghensis Jain, 1961, p. 341.
Acuticytheretta baghensis Jain, 1975b p. 202, pl. 1, figs. 12a–c, 13; pl. 3, fig. 35.
Rostrocytheridea baghensis Jain; Chaudhary et al., 2019, pp. 76, 78, pl. I, figs. 1–5.
Rostrocytheridea baghensis Jain; Chaudhary and Nagori, 2019, p. 524, pl. 2, figs. 10–12.
Description: Carapace large, elongate; surface ornamented with normal pores and with fine and delicate punctuations, which along ventral area are disposed subparallel to the margin, with fine ornamentation of ribs which are concentrically disposed parallel to the margin.
Age range: Early Turonian-Coniacian.
Rostrocytheridea cerasmoderma Ballent and Whatley, 2007, pp. 117–118, figs. 2, I-M, W.
Rostrocytheridea sp. Cf. R. Cerasmoderma Ballent and Whatley; Chaudhary et al., 2019, p. 78, pl. I, figs. 6–9.
Rostrocytheridea sp. cf. R. cerasmoderma Ballent and Whatley; Chaudhary and Nagori, 2019, pp. 524–525, pl. 2, figs. 13–14.
Description: Carapace medium to large, subtriangular in lateral view with greatest length below mid-height and greatest height in middle. Valve surface with coarse reticulate punctate network subparallel to margins.
Age range: Early Toronian-early Coniacian
Rostrocytheridea sp. Chaudhary et al., 2019, p. 80, pl. II, figs. 13–15.
Rostrocytheridea sp. Chaudhary and Nagori, 2019, p. 525, pl. 3, fig. 4.
Description: Carapace medium size, subtriangular in lateral view; valve surface consists of 9–10 fine horizontal delicate riblets and in between fine punctuation in the middle 2/3rd part of valve surface, rest of the marginal area smooth.
Age range: Late Cenomanian-early Turonian.
Family PROGNOCYTHERERIDAE Sylvester-Bradley, 1948
Subfamily PROGNOCYTHERINAE Sylvester-Bradley, 1948
Family NEOCYTHERIDEIDAE Puri, 1957
Genus Neocytherideis Puri, 1952
Neocytherideis reymenti Jain, 1975a, pp. 52, 54, figs. 1G; 3S.
Neocytherideis reymenti Jain; Jain, 1975b, p. 203, pl. 2, figs. 27a–b.
Neocytherideis reymenti Jain; Chaudhary and Nagori, 2019, pp. 527–528, pl. 3, fig. 16.
Description: Carapace subcylindrical; anterior margin obliquely rounded with a downwardly directed beak-like projection with extremity just above ventral margin; posterior slightly produced into a short subdorsal caudal process. Surface ornamented with reticules arranged parallel to anterior and ventral margins.
Age range: Late Cenomanian to early Turonian and Coniacian.
Family TRACHYLEBERIDIDAE Sylvester-Bradley, 1948
Genus Curfsina Deroo, 1966
Planileberis? sp. Indet Jain, 1975b, pp. 208–209, pl. 3, figs. 41a–b.
Curfsina? Hanumanpuraensis Chaudhary and Nagori, 2019, pp. 529–530, pl. 4, figs. 12–14.
Description: Carapace elongate, subtrapezoidal in lateral outline; dorsal margin straight, ventral margin medially concave. The anterior margin is broadly rounded and denticulate. The posterior end is angular, compressed and denticulate. The surface is marked with two ridges, one each in the dorsal and ventral region; a prominent anterior marginal ridge, and a distinct subcentral tubercle. Valve surface smooth.
Age range: Late Cenomanian to early Turonian and Coniacian.
Genus Haughtonileberis Dingle, 1969
Curfsina thuatiensis Jain, 1975b, p. 206, pl. 2, figs. 25a–b, 26a–c; pl. 3, figs. 40a–b.
Haughtonileberis thuatiensis Jain, 2019; Chaudhary and Nagori, p. 532, pl. 4, figs. 17–18.
Description: Carapace elongate, subtrapezoidal in lateral view; dorsal margin straight sloping down posteriorly, ventral margin concave. The anterior margin is broadly rounded, posterior margin triangular with an apex below mid-height. Anterior and posterior rims are prominent; three ribs, one each in dorsal, median and ventral region, anterior and median ribs joined posteriorly by a sort vertical rib; rest of surface finely reticulate. Eye spot and muscle-node weakly developed.
Age range: Late Cenomanian to Coniacian.
Genus Makatinella Dingle, 1984
Makatinella punctata Chaudhary, 2017, p. 10, pl. 1, figs. 5–8.
Makatinella punctata Chaudhary; Chaudhary and Nagori, 2019, p. 534, pl. 5, fig. 2.
Description: Carapace elongate, subrectangular in lateral view; anterior marginal rim broad, with a furrow behind it, posterior rim somewhat narrow; valve surface covered with fine reticulations and pits. Reticulation meshes are slightly raised forming a faint loop in the middle.
Age range: Early Turonian to Coniacian.
Leniocythere? thuatiensis Jain, 1975b, p. 208, pl. 3, figs. 42 a–b.
Makatinella thuatiensis Jain; Chaudhary, 2017, p.10, pl. 1, figs. 9–10.
Makatinella thuatiensis Jain; Chaudhary and Nagori, 2019, p. 534, pl. 5, fig. 3.
Description: Carapace elongate, subrectangular in lateral outline. The left valve is slightly larger than the right valve, overlapping distinctly along the anterior and posterodorsal corners. The surface of each valve with a high anterior marginal rim and a deep sulcus in front of it. Valve surface marked with fine pits and three indistinct low longitudinal ridges, one each in dorsal, median and ventral regions, median and ventral ridge forming a loop.
Age range: Early Turonian to Coniacian.
Genus Veeniacythereis Gründel, 1973
“Cythereis” raoi Jain, 1975b, pp. 206–207, pl. 3, figs. 28 a–b; 29a–b; 30 a–b.
Veeniacythereis raoi Jain; Chaudhary and Nagori, 2019, p. 534, 536, pl. 5, figs. 4–5.
Description: Carapace elongated; left valve with a prominent anterior ‘hinge ear’; anterior marginal rim strong; surface marked with three longitudinal ridges, one each in dorsal, median and ventral regions and entire surface reticulate including ribs. Eye spot and muscle node well developed. The lower half of anterior and posterior margins with spines/denticles.
Age range: Early Turonian to Coniacian.
Suborder CYTHEROCOPINA Gründel, 1967
Superfamily CYTHEROIDEA Baird, 1950
Family LOXOCONCHIDAE Sars, 1925
Subfamily NIGEROLOXOCONCHINAE Reyment, 1963
Genus Nigeroloxoconcha Reyment, 1963
Protojonesia sp. cf. P. bolliaformis (Veen) Jain, 1975b, p. 201, pl. 3, figs. 34 a–b.
Nigeroloxoconcha baghensis Chaudhary and Nagori, 2019, pp. 538, 540, pl. 5, figs. 11–12.
Description: Carapace medium-sized, subovate in lateral outline; anterior margin rounded, posterior drawn out in an upwardly directed caudal process at posterodorsal margin. The dorsal margin is straight, sloping down posteriorly. The ventral margin is strongly convex. Valve surface smooth or with indistinct pits; a distinct depression in posteroventral and ventral regions.
Age range: Early Turonian to Coniacian.
Conclusions
In the present work, 15 species of ostracods belonging to 11 genera are recorded. In general, the average number of specimens of each species is very less but the overall assemblage as a whole is very helpful in determining the precise age of the top part of calcareous sandstone at Pipaldehla. All these ostracod taxa are abundantly found in the Nodular Limestone of the Bagh-Jeerabad region, where they flourished during the whole Turonian period, probably due to the increased depth of the sea and good ecological conditions. During the early Turonian period, which occurred just after the end of the OAE2 era, taxonomic diversity decreased while the level of oxygenation and palaeoproductivity increased. These changes were accompanied by an increase in the supply of nutrients. The climate during this time was hot and humid, as evidenced by the algal assemblage reported by Kundal and Sanganwar (1998). This suggests that the calcareous top of the NSF was deposited in tropical waters at a depth of 10–12 meters below low tide level. The deposition occurred in a moderate energy setting with moderate turbulence and normal salinity of water.
After analysing the ostracods in this bed, we have concluded that they share a close resemblance to the ostracod fauna found in the lower section of the Nodular Limestone in the east, as well as the oyster-bearing limestone in the west of the Bagh Group. Probably these might have been deposited during the same time when the lower part of Nodular Limestone was deposited in the eastern part. It is believed that the first marine transgression occurred from the west during the late Cenomanian Period, resulting in the deposition of Oyster beds and oyster-bearing limestone in the Rajpipla-Kawant area of Gujarat. As the transgression progressed towards the eastern part of the Narmada Valley, these marine sediments were deposited in other linear belts of graben or half-graben. In the Jhabua area, marine incursion took place during the early Turonian period, and the top of the calcareous sandstone exhibited the presence of calcareous algae, ostracods, foraminifers and other mega fossils. Among these, calcareous algae and ostracods are indicative of the early Turonian age.
Footnotes
Acknowledgements
The authors are thankful to the Department of Geology, Faculty of Earth Sciences, Mohanlal Sukhadia University, Udaipur for extending their support in this work by providing laboratory facilities. We thank anonymous reviewers for their critical and constructive suggestions to improve the manuscript.
Declaration of Conflicting Interests
The authors declared no potential conflicts of interest concerning the research, authorship and/or publication of this article
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: The authors are thankful to the Ministry of Education, Government of India, New Delhi for the funding through Rajasthan State Higher Education and State Project Directorate Jaipur under RUSA scheme to the Department of Geology, Mohanlal Sukhadia University Udaipur.