
Abstract
The Cretaceous-Palaeogene intertrappean deposits of the Deccan Volcanic Province of India comprise vertebrate, invertebrate and plant fossils of the dominantly continental realm. A 3–4 m thick shaly-sandstone lacustrine intertrappean sequence of Early Palaeogene (Palaeocene-Early Eocene) Bamanbor Formation, Chotila Basin (Saurashtra province, Gujarat State, western India) has yielded a collection of molluscans (bivalve and few gastropods) fauna. High-resolution digital photography and radiography of the recovered bivalves allowed us to gain detailed inferences on shell morphology. In a taxonomical context, we herein propose a new unionid molluscan superfamily Deccanoidea nov. represented by a single new family Deccanoidae nov. that includes two new subfamilies, that is, Deccanoinae nov. (represented by two new genera Deccanoida gen. nov. [including four new species: D. conrugis sp. nov., D. aleta sp. nov., and D. costaria sp. nov.] and Bamanboria gen. nov. [including one new species Bamanboria oblongis gen. et sp. nov.] and Chotiloinae nov. [represented by one new genus Chotilia gen. nov. and three new species: Chotilia trappeansis sp. nov., C. tuberculata sp. nov. and C. deccanensis sp. nov.]). Inorganic geochemistry of host (unionid-yielding) lithologies reveals oxic-freshwater conditions, humid-climate and moderate rainfall. The data on historical biogeography reveals that the Indian subcontinent hosted unionids during the late Cretaceous; however, the presence of new (at family/generic level) unionid fauna during the Early Palaeogene (Palaeocene-Early Eocene) in India indicates changes in the geographic position and ecologic condition linked to northward drift and lava outpouring, respectively. A disjunct distribution of unionid fauna in the erstwhile Gondwanan continents and insular India during the Cretaceous-Palaeogene interval can be explained in terms of both vicariance and dispersal.
Introduction
The fossil record of freshwater bivalves is sparse, particularly with limited evidence of their early evolutionary phases due to taphonomic constraints in continental depositional systems. The early record of the nonmarine bivalve of Pangea and subsequent records from the Gondwana exemplify vicariant species. The earliest nonmarine bivalve Archanodon is reported from the Middle Devonian—Late Carboniferous (Chamberlain & Chamberlain, 2007), Bellvale Sandstone (Givetian) of northern New Jersey, North America and Anthracosiidae from the Upper Carboniferous, UK (Eagar, 1953). The widespread Permian nonmarine bivalves Palaeomutela and Palaeanodonta (Silantiev & Carter, 2015) are reported from the Pangea supercontinent and nonmarine Autobranch (Palaeoheterodont) genera Gondwanaiadites, Bakulia, Gangamya, Raniganjelia, Gondwanadontella and Indonellina are reported from the Permian of the Raniganj Formation of the Raniganj, Jharia and North Karanpura basins of the Damodar valley from the Indian Gondwana Coal Measures (Silantiev et al., 2015).
The earliest unionids are reported from the Olenekian Stage of the Early Triassic Karoo Basin of South Africa (Yates et al., 2012) which are now widespread across all continents except Antarctica (Graf & Cummings, 2007, 2009). The freshwater unionids Tihkia karooensis (Cox, 1932) found in the Middle Triassic Manda Beds of Tanzania and Ntawere Formation of Zambia (Cox, 1932); the Late Triassic (Late Carnian or Early Norian) species Tihkia corrugata reported from Maleri Formation of Tihki region of Madhya Pradesh (formerly a part of Hyderabad state), India (Sahni & Tewari, 1958) and Silesunio parvus and T. corrugata from Late Triassic (Carnian-Rhaetian), Silesia, Poland are described under a suborder Silesunionina (Skawina & Dzik, 2011).
The Pangea started breaking into Laurasia and Gondwanaland in the Early Jurassic (Ali & Aitchison, 2008; Rogers & Santosh, 2004) with India lacking evidence of the unionids until their dramatic appearance in the Late Cretaceous.
The Deccan Volcanic Province of the Indian Peninsula is characterised by a majority of freshwater intertrappean sequences which bear different types of unionids, primarily reported from the Nagpur and Gurmatkal, such as Unio malcolmsoni, Unio deccanensis, Unio hunteri, Unio mamillatus, Unio imbricatus and Unio carteri (Hislop, 1860), along with one species of Sphaeriida, Pisidium medlicottianum from Goraha, Narmada intertrappean beds of peninsular India (Hislop, 1860). P. medlicottianum of Hislop (1860) was later redefined as a Unionoid Lamellidens vredenburgi by Prashad (1921). Hislop’s (1860) descriptions were in Latin which hindered progress in molluscan research, they were later translated to English for potential usage by Hartman et al. (2008). The bivalve-bearing intertrappean sections described by Hislop (1860) were redefined and assigned the latest Cretaceous age based on the presence of dinosaurian and floral remains (Hartman et al., 2008; Prasad & Khajuria, 1996; Sahni et al., 1984; Samant et al., 2020). Recently reported intertrappean unionids include Lamellidens from Rajindragram and Deori of Madhya Pradesh (Gangopadhyay et al., 2011), U. deccanensis from Topidhana of Madhya Pradesh (Srivastava & Kandwal, 2013) and Unio from chertified limestone bed of the Maastrichtian (Cretaceous) from Gurmatkal, Karnataka (Jalal et al., 2020).
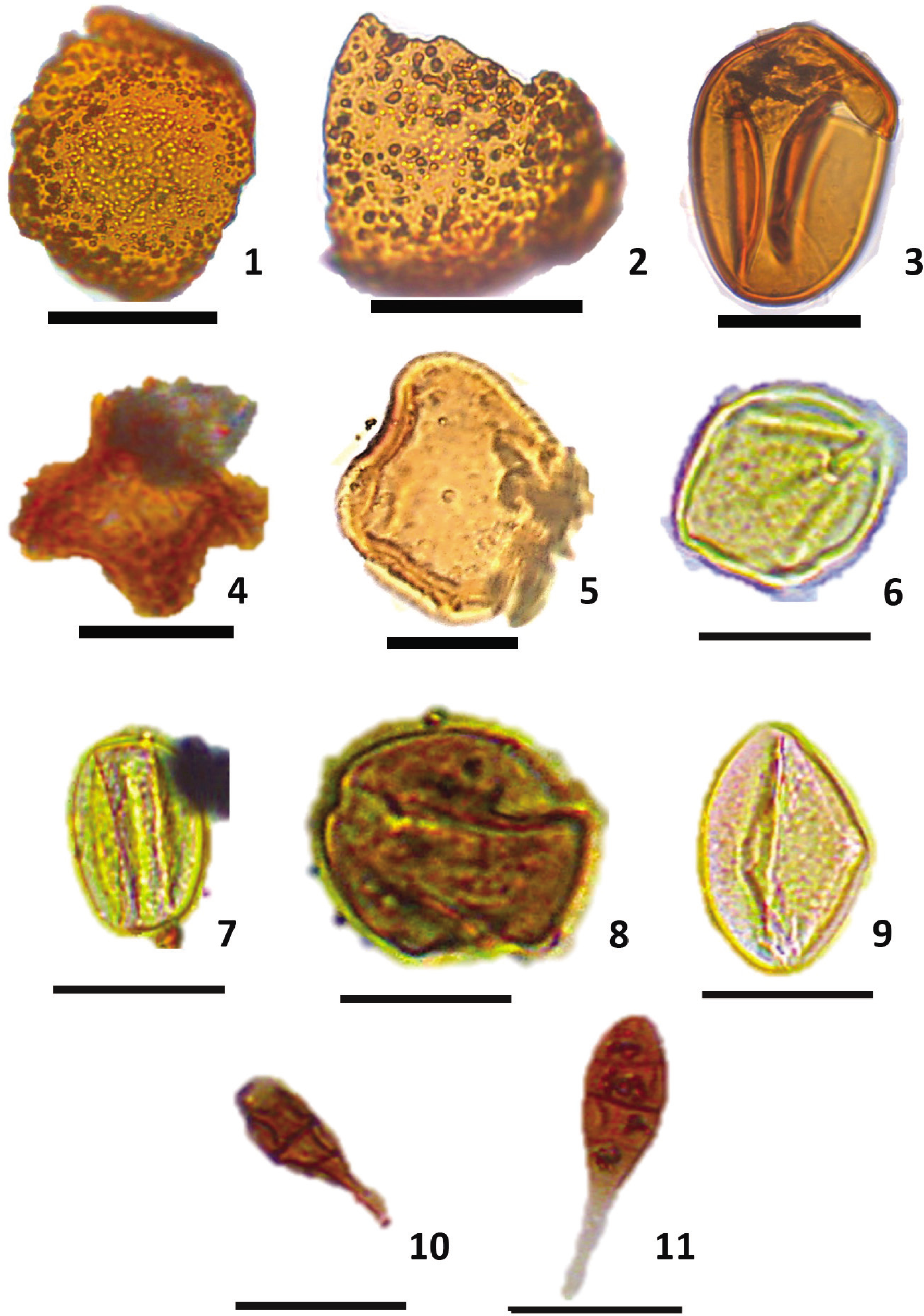
The Deccan volcanic rocks of India range from Late Cretaceous to Early Eocene in age (Agrawal & Rama, 1976; Cripps et al., 2005; Hora, 1938), concurrently the sedimentary deposit associated with lava flows (intertrappeans) are identical in age with similar constraints. However, the Saurashtra intertrappean deposit needs to have an absolute age. In the absence of the diagnostic fauna or flora and absolute geochronological dating, the complexity deepens to resolve the age of these deposits. A few attempts have been made based on paleontological evidence of the Saurashtra intertrappeans. Borkar (1973) reported clupeoid fish such as Horaclupea intertrappean and perciforms Paleopristolepis chiplonkari, and Paleopristolepis feddeni from the Bamanbor area and suggested Paleocene-Early Eocene age. Saxena (1990) and Frederiksen (1994) assigned Palaeocene to Early Eocene for the palynoflora Intrareticulites brevis, Neocouperipollis spp., Striacolporites striatus, Retitricolporites crassimarginatus and Rhombipollis sp. Later, Samant et al. (2014) found a similar palynoflora assemblage from the Ninama intertrappean of the Saurashtra region and suggested the Palaeocene age. Moreover, the presence of palynofloras such as Longapertites retipilatus (Mandal & Kumar, 1997), Matanomadhiasulcites maximus, Rhombipollis geniculatus, Yeguapollis sp and Lakiapollis ovatus from the Ninama intertrappeans suggest an Early Eocene age (Plate 1, Figures 1–5). Bivalve-bearing shaly sandstone of the Bamanbor Formation (Figure 1) (Patel & Shah, 2023) consists of a variety of palynofossils, including pollens like Florschuetzia rajpardiensis, Monocolpopollenites plicatus, Meliapollis ramanujamii and Rhoipites kutchensis, and fungal spores like Polycellaesporonites psilatus and P. saxenae (Plate 1, Figures 6–11). Based on the presence of marker pollen M. ramanujamii (Khanolkar & Sharma, 2019; Saxena & Tripathi, 2012) and F. rajpardiensis (Samant & Phadtare, 1997) an Early Eocene age is assigned.
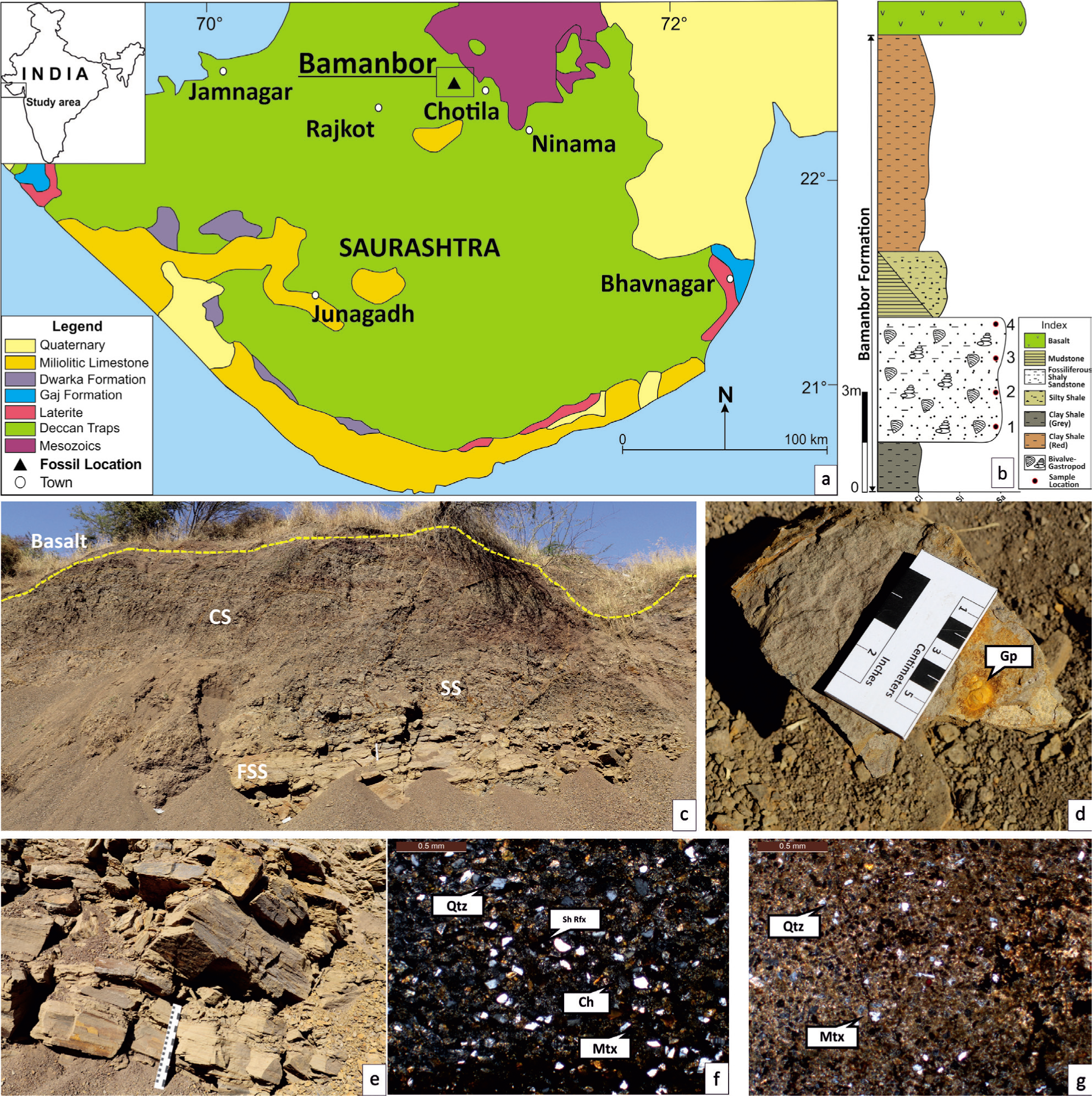
(a) General geological map of the part of the Saurashtra Peninsula showing bivalve bearing location (solid triangle); (b) Litholog of Bamanbor Road Section (BRS) with positions of four geochemically analysed samples; Field photographs of BRS showing; (c) Fossiliferous shaly sandstone, silty shale and clay shale; (d) FSS with embedded gastropod (Gp); (e) Thickly bedded FSS unit; Photomicrographs of cross-polarised view of FSS showing; (f) Coarse silt to fine sand-sized subangular to subrounded quartz (Qtz), shale lithoclasts (Sh Rfx), chert fragments (Ch) with abundant matrix (Mtx), and; (g) Quartz and chert fragments with abundant matrix and opaques (iron oxide).
The present investigation is an attempt to study the taxonomic diversity of unionid bivalves recovered from an early Paleogene (Palaeocene-Early Eocene) intertrappean site, Bamanbor, in Saurashtra province, Gujarat State, western India. Additionally, the geochemical data incorporating the XRF technique together with the recorded molluscan assemblage has been utilised to infer the depositional palaeoenvironment. Further, we herein discuss the palaeobiogeographic significance of the recovered unionid bivalve fauna. Three genera and seven species of the order Unionida are described systematically under the superfamily Deccanoidea nov. and their affinities and palaeobiogeographic implications are discussed.
Geological Settings and Fossiliferous Locality
The intertrappean rocks are exposed on the Chotila–Rajkot Highway (15 km west towards the Rajkot) near Bamanbor village, Gujarat State, India (Figure 1a). The Bamanbor Road section (22° 25’ 38” N, 71° 03’ 23” E) of Chotila Basin has yielded relatively well-preserved fossils of bivalve shells (Figure 1a) (Patel & Shah, 2023). The fossiliferous stratum is dark grey to rusty brown-coloured shaly sandstone and comprises thinly bedded, alternating silty shale and/or clay shale layers, capped by Deccan Trap (Figure 1b). These layers often show thinning, thickening, at times pinching out and reappearing laterally, reflecting the bed geometry of the host sediments. In total, 30 cm thick fine-grained sandy layers (with overall bed thickness of 3–4 m) are common which contain >50% of fine-grained sand-sized particles (Figure 1c–e), dominated by quartz and lithic fragments of chert, approximated based on petrographic analysis (Figure 1f–g). This layer contains moderately abundant bivalve shells along with gastropods (Figure 1c–e) (Patel & Shah, 2023).
Early Eocene palynofossils from the Saurashtra Peninsula. Pollen grains from the Ninama intertrappeans include; Fig. 1, Longapertites retipilatus; Fig. 2, Matanomadhiasulcites maximus; Fig. 3, Lakiapollis ovatus; Fig. 4, Rhombipollis geniculatus; Fig. 5, Yeguapollis sp. Pollen grains include; Fig. 6, Florschuetzia rajpardiensis; Fig. 7, Monocolpopollenites plicatus; Fig. 8, Meliapollis ramanujamii; and Fig. 9, Rhoipites kutchensis. Fungal spores include; Fig. 10, Polycellaesporonites psilatus and Fig. 11, P. saxenae from the Chotila intertrappeans. The scale bar represents 20 µm.
Material and Methods
The specimens were carefully collected, labelled, wrapped in cotton, kept in sample bags and brought to the Palaeontology Laboratory of the Department of Geology, The M. S. University of Baroda, Vadodara district, Gujarat State. The recovered fossils are hard calcareous shells, a few being weathered partially or completely, and some preserved as internal/external moulds and casts. A few complete shells partially replaced and recrystallised are retrieved. The majority are preserved as casts, which are rusty brown to dirty yellow, very fine-grained, soft and crumbly owing to their friable nature, hindering their retrieval from the host rocks. The retrieval of complete shells was also difficult from the host rocks due to softness, minor cracks and the development of fine fissility to a certain extent, resulting in the crumbling of both rocks and shells. It has also further complicated the separation procedures during washing and cleaning of the shells, where fine-grained materials are easily washed away. The specimens were cleaned using a soft nylon brush, needle and penknife, then carefully washed to remove extraneous sediments and subsequently dried; extra precautions were taken owing to their delicacy. The retrieved complete bivalve shells have provided important information on cardinal structures and ornamentation, which are useful for their detailed examination.
For examining the morphology, digital and X-ray photography were employed on the fossils to capture external as well as internal features. X-ray photography is advantageously used to examine the hard parts in sediment-enveloped specimens (Racicot, 2016). The anatomical impressions like retractor and protractor muscle scars, gill structures and pallial lines are obscure due to preservational constraints, but anterior adductor muscle scar and impression is observed in calcitic shells and their internal moulds. To study the shell morphology, a high-power magnifying hand glass (10×), a vernier caliper and a ruler of 10 mm resolution were used. The shell shape, symmetry, dentition, commissure, umbonal position, curvature and sculpture, pointing of beak, ligament position, curvature and arc of the shell, shell sculpture and ornamentation, nature of growth lines were taken into consideration to define and differentiate among specimens (Aldridge, 1999; Anderson, 2014; Bogatov & Prozorova, 2017; Klishko et al., 2018; Prozorova & Bogatov, 2006; Sayenko et al., 2020; Van Bocxlaer, 2011; Zieritz et al., 2015). The complete specimens are also reconstructed as line drawings which provide insights into the internal and external morphological features and also give an understanding of their evolution. XRF analysis of the bivalve-bearing horizons was conducted to interpret the various paleoenvironmental parameters, including salinity, redox conditions, climate, humidity and precipitation. A total of four samples from the bivalve bearing horizon were processed to obtain the concentration of inorganic geochemical elements and major oxide, using the XEPOS HE XRF at the Institute of Seismological Research (ISR), Gandhinagar, India. The samples were dried, homogenised and crushed to <45 μm size; the crushed samples were packed in cups and used for analysis. The analytical precision of major oxide was better than 5% and that of elements was better than 10% (Makwana et al., 2019; Shah & Patel, 2024). All the specimens are housed at the Geology Museum, Department of Geology, The M. S. University of Baroda, Vadodara District, Gujarat State, India under the prefix Geo/Mus/BF/I to Geo/Mus/BF/XXVI.
Field Observations on the Preservation of Unionid Bivalves
Bivalves are ubiquitous in aqueous environments and the majority of the fresh water bivalves secrete aragonitic calcium carbonate shell. The original aragonitic mineral composition is highly unstable and readily recrystallises to calcite (Casella et al., 2017). The preserved materials are affected by taphonomic processes, where the majority of shells are either disarticulated with in situ burial, short distance transport and burial due to necrolysis, or severely affected by the solution, leading to the alteration of the original preserved shell material. Preservation of the shells is controlled by their own fragility, transportation, energy conditions, salinity and the presence of the scavengers. Disarticulated young and adult shells are represented as either right or left valves, lying horizontally, due to post-mortem transportation. Few articulated shells are also observed either closed or open in a butterfly position.
The textural and mineralogical composition of sediments has influenced and substituted the original materials, where original aragonitic shells are leached out and replaced by either clayey sediments or ferruginous materials. It resulted in the preservation of shells in the form of moulds or casts and also influenced the crumbling of the shells during the retrieval. Even though, these altered shells, in some cases, show faithfully defined bivalve outlines and preserve delicate external or internal morphological features like impressions of cardinal characteristics. Occasionally, well-preserved disarticulated, hard shells are also found, with diagenetic alteration of the original aragonitic material into more stable sparry to cleavable calcite (Casella et al., 2017). Percolating rainwater has also caused severe alteration in the original composition, compromising the recovery of the bivalve specimens.
Taxonomic Constraints
The early nonmarine bivalve fossils record is sporadic and poor due to discontinuity in sedimentary record caused by the ephemeral nature of most of the freshwater depositional systems of the continental realm. Hence, there is no systematic record available on nonmarine bivalve fossils that document the lineage since their inception. The freshwater unionid ancestry possessed a crucial weak point due to their allopatric nature, leading to difficulties in recording their evolutionary pathway and subsequently posing challenges in assigning formal taxonomic categories for the older fossils (Chamberlain & Chamberlain, 2007).
The earliest recorded freshwater bivalve is based on a sole species, Archanodon from the Middle Devonian to Carboniferous; its taxonomic status was debatable (Cox et al., 1969), but based on elongate, inequilateral shape and enlarged posterior region, it is considered as an earliest known member of the bivalve order Unionida, even in absence of the soft parts (Chamberlain & Chamberlain, 2007). The status of the Upper Carboniferous nonmarine Anthraconaia and Carbonicola is confined to the family Anthracosiidae of the order Cardiida (Cox et al., 1969), while the Permian widespread bivalves species Palaeomutela and Palaeanodonta are described under the order Actinodontida (Silantiev & Carter, 2015). The first record of the nonmarine bivalve from the Permian Raniganj Formation (Indian Gondwana Coal Measures) records autobranchs, namely Gondwanaiadites, Bakulia, Gangamya, Raniganjelia, Gondwanadontella and Indonellina, which is endemic (Silantiev et al., 2015). However, there is no link between the earliest form of nonmarine bivalves and the freshwater unionid bivalves of 230 Ma., that is, the late Triassic (Skawina & Dzik, 2011), which marks a long time gap in the records. However, the Triassic freshwater bivalves of Poland are to be considered as immediate ancestor of marine extent relative of the Australian Neotrigonia (Skawina & Dzik, 2011).
The earlier unionid bivalves evolved during the Late Triassic in Pangea and subsequently dispersed to all continents except Antarctica (Graf & Cummings, 2007). This group is well-established in Triassic and subdivided into three suborders: Unionidina, Siluesunionidina and Hyriidina (Carter et al., 2011), each bearing distinct morphological features. The Unionidina and Siluesunionidina were separated based on gill structures, the former possess eulamellibranch gills while the latter possess filibranch gills (Skawina & Dzik, 2011). Order Unionida is characterised by both, fossil and living forms and is further classified into several superfamilies, such as Silesunionoidea, Mullerioidea, Trigonioidoidea, Tamesnelloidea (Van Damme et al., 2015), Unionoidea, Etherioidea and Hyrioidea (Carter et al., 2011).
India also evidences unionids from the Late Triassic Pangea (Late Carnian or Early Norian), specifically from the Maleri Formation of the Tihki region, Madhya Pradesh, India. This oldest described freshwater bivalve is poorly documented and represented by only one species belonging to Suborder Siluesunionina, Family Silesunionidae: Tihkia corrugate (Sahni & Tewari, 1958). The incompleteness of the freshwater bivalve record, particularly of Jurassic in the Indian continent poises the difficulty in establishing the relationship with older ancestors. Once again, these freshwater bivalves are recorded and described from the Upper Cretaceous intertrappean of the Nagpur area and are considered to belong to the superfamily Unionidea (Hislop, 1860). However, there is no fossil record of freshwater bivalves from the intertrappean which can be considered as descendants of the Triassic suborder Siluesunionina.
Interpreting the specimen described from the intertrappean of Saurashtra is challenging due to the incomplete fossil record of the Indian continents. Additionally, freshwater bivalves exhibit a high degree of morphological disparities. To evaluate their evolutionary invention, major diagnostic characters are required sequentially across different geological epochs. Skawina and Dzik (2011) proposed filibranch versus eulamellibranch grades to diagnose paraphyletic units at the suborder rank for the Late Triassic freshwater bivalves. The intertrappean specimens are not assigned a suborder category due to a lack of connecting links, poor preservation and unrecognisable gill structures and are kept open to inspect the well-preserved materials yet to be found.
At present five superfamilies such as Silesunionoidea, Tamesnelloidea, Unionoidea, Etherioidea and Hyrioidea are described in the suborder Unionina. The present material collected from the Deccan intertrappean shows characteristically distinct hinge structures, overall shape and ornamentation, which is a crucial character in bivalve taxonomy enabling, the erection of the new category at the Superfamily level, Deccanoidea nov., based on new evidence, although still incomplete and tentative, it should be included in concepts of high-rank unionid taxa. Although the specimens are limited, the analysis of the morphologic features is distinct, that have assisted in discriminating the species. This allopatric population is deemed to fit into separate biological entity until more anatomical evidence on lacustrine unionids is made available.
Systematics
A total of 26 specimens, relatively well preserved in the fine-grained shaly-sandstone of the Bamanbor Formation, Bamanbor Road Section (Figure 1) are studied for their morphological features which allowed differentiation into seven bivalve species. The detailed morphological analysis such as shape, size, ornamentation and dentition enabled to classification of them into a superfamily, Deccanoidea nov., family Deccanoidae nov., divided into the following two subfamilies: Deccanoinae nov. and Chotiloinae nov. The former subfamily consists of two genera, Deccanoida nov., and Bamanboria nov., with four species, Deccanoida conrugis gen. et sp. nov., Deccanoida aleta gen. et sp. nov., Deccanoida costaria gen. et sp. nov. and Bamanboria oblongis gen. et sp. nov.; latter subfamily comprises of one genus Chotilia nov. with three species, Chotilia trappeansis gen. et sp. nov., Chotilia tuberculata gen. et sp. nov. and Chotilia deccanensis gen. et sp. nov.
These freshwater bivalves are described following the systematic classification of Unionida (Carter et al., 2011) and the Treatise on Invertebrate Palaeontology, Part N, Volume 1, Mollusca 6, Bivalvia (Cox et al., 1969).
Order Unionida, Gray (1854)
Superfamily Deccanoidea nov.
Diagnosis: Equivalve or inequivalve, inequilateral, elliptical to trapeziform, extremely prosogyrous umbones, flatly compressed unionids with prominent anterior muscle scar pit, 2–3 diverging cardinal teeth, one lateral tooth below the opisthodetic, parivincular ligament and entirely corrugated commissure. The length of the shells ranges from 15 to 60 mm.
Remark: The defined superfamily from the Saurashtra intertrappean, described under the Order Unionida, is a highly evolved paraphyletic group, owing to its allopatric nature and is assigned a new taxonomic category. The superfamily shows distinctly different shell morphological features as compared to unionid reported from other intertrappean of the Deccan Volcanic Province of India.
Family included: Deccanoidae nov.
Family Deccanoidae nov.
Diagnosis: Equivalve or inequivalve, elongated, prosogyrous, opisthodetic unionid bivalves with anisomyrian muscle scars, one lateral and two to three cardinal teeth. Variable sizes, moderately inflated but with large capacious cavities below umbones. Ornamentation consists of growth lines, costae and prominent corrugations on the posterior-ventral margins. The shell is probably composed of aragonite and calcite with 1.0 to 1.5 mm thickness with the possible presence of the innermost nacreous layer.
Subfamily included: Deccanoinae nov. and Chotiloinae nov.
Subfamily Deccanoinae nov.
Diagnosis: Elongated, equivalve or inequivalve, equilateral to sub-equilateral bivalves, integripalliate, anisomyrian, heterodont with a lateral tooth, prominent posterior wing, opisthodetic ligament and prominently developed ribs and corrugated commissures.
Genera included: Deccanoida gen. nov. and Bamanboria gen. nov.
Genus Deccanoida nov.
Type species: Deccanoida conrugis gen. et sp. nov.
Etymology: The genus name is derived from the Deccan Traps, a large igneous province (LIP) of India where shells are intimately associated with the intertrappean.
Diagnosis: As per the type species.
Genus Deccanoida conrugis gen et sp. nov. (Plate 2, Figures 1–11)
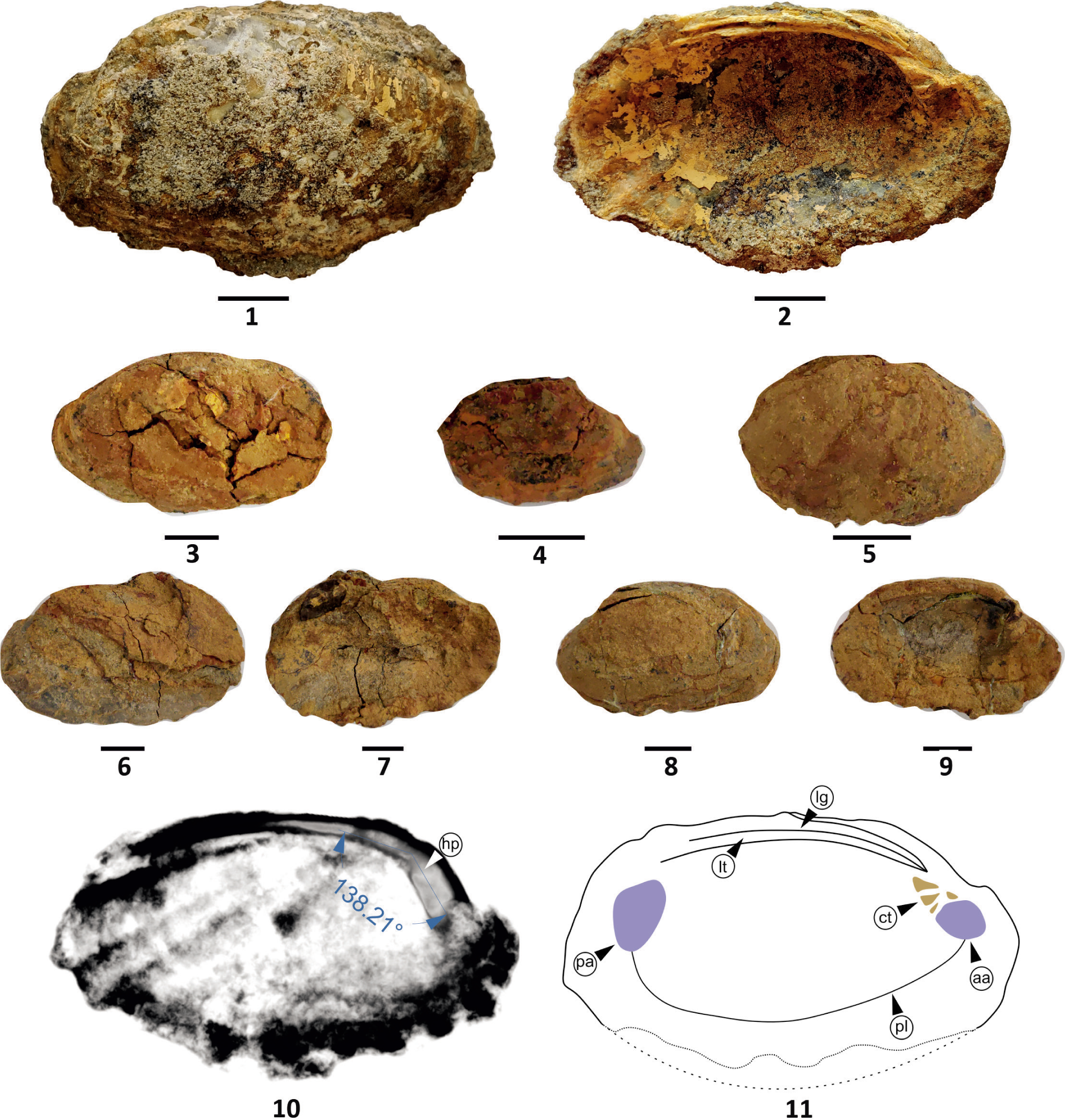
Deccanoida conrugis sp. nov; Fig. 1, External view of holotype; Fig. 2, Internal view of holotype; Figs. 3–9, Paratypes of D. conrugis; Fig. 10, X-ray image of the holotype showing umbonal angle and hinge plate (hp); Fig. 11, Line drawing of the holotype shows internal morphology (ct = cardinal teeth, aa = anterior adductor muscle scar, lt = ligament tooth, lg = ligament, pl = pallial line, pa = posterior adductor muscle scar). All scale bars represent 1 cm.
Material: Four left valves and two right valves with several broken ones are collected.
Etymology: The species name refers to the corrugated nature of the ventral margin (Latin conrugis, meaning corrugated).
Holotype: Geo/Mus/BF/I (Plate 2 Figures 1–2).
Paratype: Geo/Mus/BF/II, Geo/Mus/BF/III, Geo/Mus/BF/IV, Geo/Mus/BF/V, Geo/Mus/BF/VI (Plate 2, Figures 3–9).
Diagnosis: Elliptical, compressed shell, with distinct corrugations towards the posterior margin and a small posterior dorsal wing.
Description: Shell thick, recrystallised and petrified. It is elongated and elliptical (Plate 2, Figures 1–2). The shell is long ranging from 2.3 to 6.9 cm (length approximately 1.5 times its height) and inequilateral. The ornamentation on the umbo is obscure whereas the entire shell shows prominent concentric growth lines (Plate 2, Figure 1). The antero-ventral and posterior-ventral margins of the shell show characteristic corrugation (Plate 2, Figures 1–2). The large and well-spaced radially broad costae in the central and lower part of the shell are typical and they either truncate or fade towards the umbo (Plate 2, Figure 1). The umbo and beak are subcentrally positioned towards the anterior. The beak is prosogyrous and the umbonal angle is 138° (Plate 2, Figure 10). Small anterior and long posterior dorsal wings are noticeable. Parivincular ligament is distinct, low-arched and well-preserved. Three to four hinge teeth, diverging towards the mantel cavity, are discrete. One long lateral posterior tooth parallel to the ligament is prominent. Anterior adductor muscle scar is definitive whereas posterior adductor muscle scar, posterior and anterior retractor muscle scars, and pallial line are obscure. The commissure is corrugated (Plate 2, Figure 11).
Dimensions: see Table 1.
Dimensions of Deccanoida conrugis gen et sp. nov.
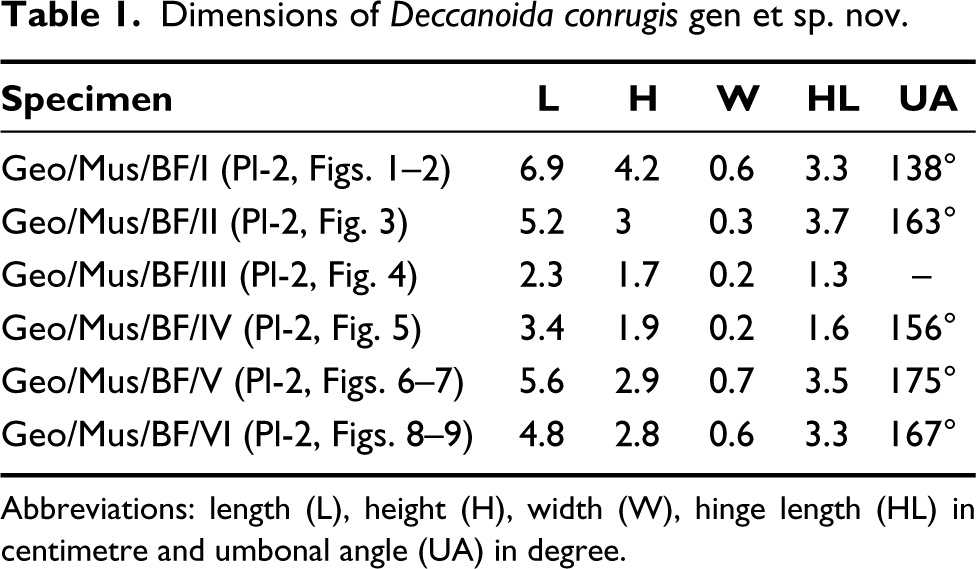
Abbreviations: length (L), height (H), width (W), hinge length (HL) in centimetre and umbonal angle (UA) in degree.
Remarks: D. conrugis gen. et sp. nov., is elliptical and inequilateral with prominent development of corrugations, and consists of 3–4 well-preserved teeth, and prominent arcuate ligament. Comparative shell morphology of the present fossils is difficult due to inadequate records of freshwater bivalves from the Paleogene intertrappean of India. The only resembling available record is of Unio hunter Hislop (1860) described from the Late Cretaceous intertrappean beds of Nagpur, which is a smooth, subquadrangular, inequilateral, posteriorly compressed, unionid bivalve with radiate-rugose apices. The overall morphology of the valves, along with their distinct dentition and shape, distinguishes them from the U. hunteri, and hence, are considered as new unionid species.
Type locality: Bamanbor, Chotila–Rajkot Highway, Gujarat, India.
Type horizon: Bamanbor Formation.
Genus Deccanoida aleta gen. et sp. nov. (Plate 3, Figures 1–12)
Deccanoida aleta sp. nov; Figs. 1–2, Holotype: Fig. 1, External view; Fig. 2, Internal view; Figs. 3–10, Paratypes: Fig. 3, External view and internal shell shows the dentition and ligament (Fig. 4); Fig. 5, Internal view shows the dentition and ligament; Figs. 6–10, External view; Fig. 11, X-ray of holotype with umbonal angle and hinge plate (hp); Fig. 12, Internal shell morphology illustrated of holotype. (ct = cardinal teeth, aa = anterior adductor muscle scar, lt = ligament tooth, lg = ligament, pl = pallial line, pa = posterior adductor muscle scar). All scale bars represent 1 cm.
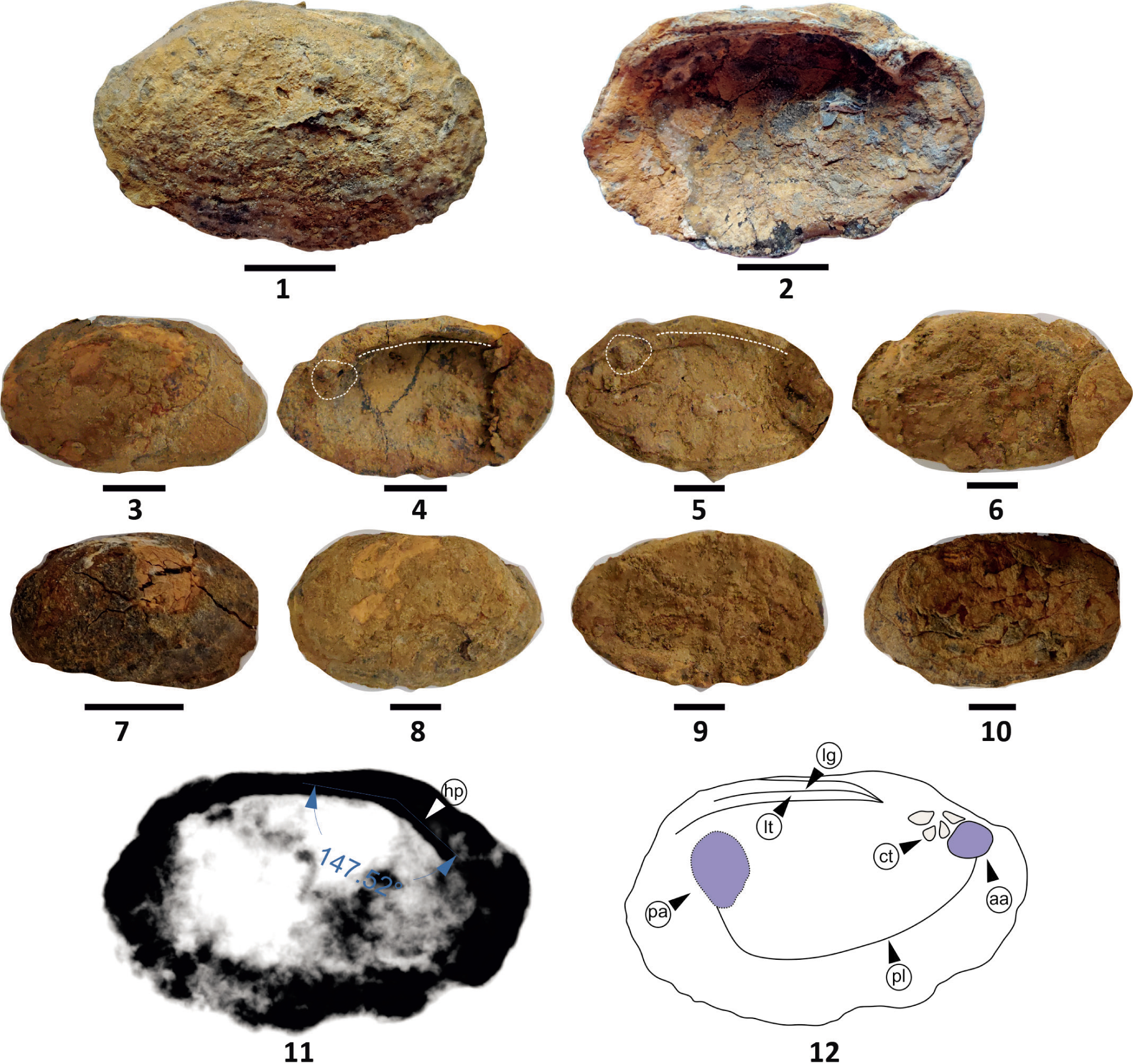
Material: One complete specimen, five left valves and one right valve is preserved.
Etymology: The species name refers to the presence of a prominent posterior wing in the specimen (Latin aleta, meaning wing).
Holotype: Geo/Mus/BF/VII (Plate 3, Figures 1–2).
Paratype: Geo/Mus/BF/VIII, Geo/Mus/BF/IX, Geo/Mus/BF/X, Geo/Mus/BF/XI, Geo/Mus/BF/XII, Geo/Mus/BF/XIII (Plate 3, Figures 3–10).
Diagnosis: Rhomboidal to sub-rhomboidal shape, with a large prominent posterior wing and parivincular ligament.
Description: Elliptical to rhomboidal shape, posteriorly flattened shell, with faint growth lines and length ranging from 2.5 to 5.5 cm (Plate 3, Figure 1–2). Characteristic ribs and folds on the commissure (Plate 3, Figures 1, 2, 5, 6, 9, 10, 12). The umbo is low, subcentral towards anterior with an angle of 147° (Plate 3, Figure 11). Three hinge teeth diverging towards the mantel cavity are discrete. A prominent high posterior dorsal wing, straight ligament and one lateral tooth are distinct. The anterior adductor muscle scar is small but deeply impressed whereas the posterior muscle scar is large but faintly preserved with a small posterior retractor muscle scar; the pallial line is distinct (Plate 3, Figure 1). The anterio-ventral and posterio-ventral margins of the shell also show characteristic corrugations. The posterior part is wider compared to the anterior (Plate 3, Figures 1–2). Ornamentation includes distinct growth lines.
Dimensions: see Table 2.
Dimensions of Deccanoida aleta gen. et sp. nov.
Remarks: The shell morphology helps in designating the D. aleta gen. et sp. nov. a new taxon. It is differentiated from the D. conrugis gen. et sp. nov. based on the presence of a prominent posterior wing. The nearest resembling unionid of the Late Cretaceous is U. carteri (Hislop, 1860) with a smooth sub-elliptical shell, with prominent umbones; whereas D. aleta gen. et sp. nov. is characterised by overall rhomboidal shape; discrete dentition with a prominent posterior wing.
Type locality: Bamanbor, Chotila–Rajkot Highway, Gujarat, India.
Type horizon: Bamanbor Formation.
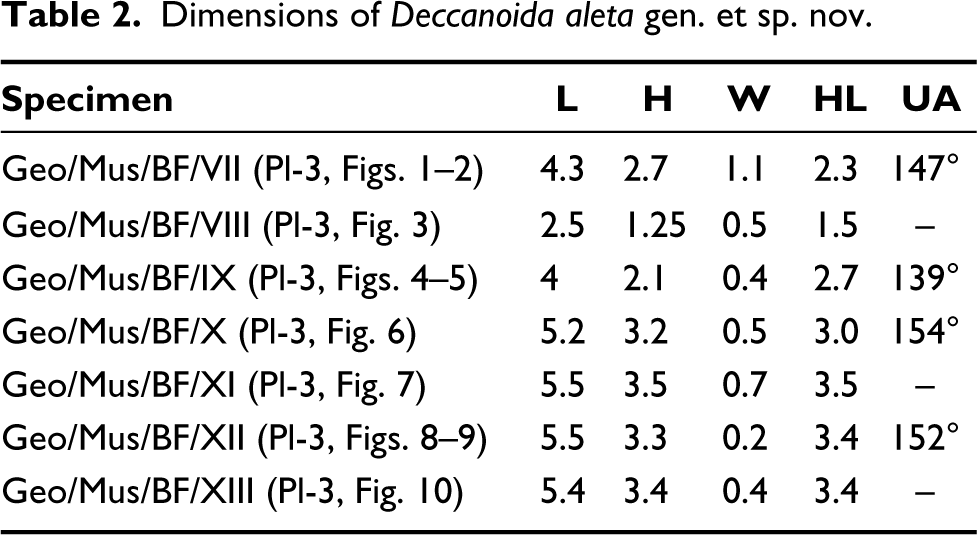
Genus Deccanoida costaria gen. et sp. nov. (Plate 4, Figures 1–6)
Deccanoida costaria sp. nov; Figs. 1–2, Holotype: Fig. 1, Left external view; Fig. 2, Right external view; Figs. 3–4, Paratype: Fig. 3, Left external view; Fig. 4, Right external view; Fig. 5, X-ray of the holotype (left valve) with umbonal angle and hinge plate; Fig. 6, Line drawing of the holotype illustrates the internal shell morphology. (ct = cardinal teeth, aa = anterior adductor muscle scar, lt = ligament tooth, lg = ligament, pl = pallial line, pa = posterior adductor muscle scar). All scale bars represent 1 cm.
Material: Two complete specimens with both the valves closed are preserved.
Etymology: The species name refers to the presence of prominent ribs towards the ventral (Latin costae, meaning ribs).
Syntype: Geo/Mus/BF/XIV, Geo/Mus/BF/XV (Plate 4, Figures 1–4).
Diagnosis: Shell semi-circular, moderate size, low to moderate posterior wing and prominent corrugations towards the posterior margin extending sub-centrally.
Description: Equivalve, inequilateral, posteriorly compressed semi-circular to rhomboidal in shape with 5.7 to 6.4 cm length (Plate 4, Figures 1–4). Anteriorly placed umbo, prosogyre, ligament opisthodetic and parivincular with umbonal angle 154° (Plate 4, Figure 5). The posterior of the left and right valve shows prominent ribs and corrugations which continue till the subcentral anteriorly placed, low umbo (Plate 4, Figures 1–2). It has faint growth lines. The umbo is convex with concentric ornamentation. The growth lines are visible on the anterior dorsal wing of both the valves; a small posterior wing is present.
Dimensions: see Table 3.
Dimensions of Deccanoida costaria gen. et sp. nov.
Remarks: D. costaria gen. et sp. nov. is regarded as a new species due to its sub-rhomboidal to semi-circular shape and prominent costae extending sub-centrally towards the umbo. The external shell morphology is distinct from D. conrugis gen. et sp. nov. and D. aleta gen. et sp. nov. but the articulated nature of the shells prevents further comparison of internal shell morphology. The complete species is redrawn and shows hinge structures, muscle scars and pallial lines (Plate 4, Figure 6).
Type locality: Bamanbor, Chotila–Rajkot Highway, Gujarat, India
Type horizon: Bamanbor Formation
Genus Bamanboria nov.
Type species: Bamanboria oblongis gen. et sp. nov.
Etymology: The genus name refers to the geographic name of the location wherein it was reported (Village—Bamanbor).
Diagnosis: As for the type species.
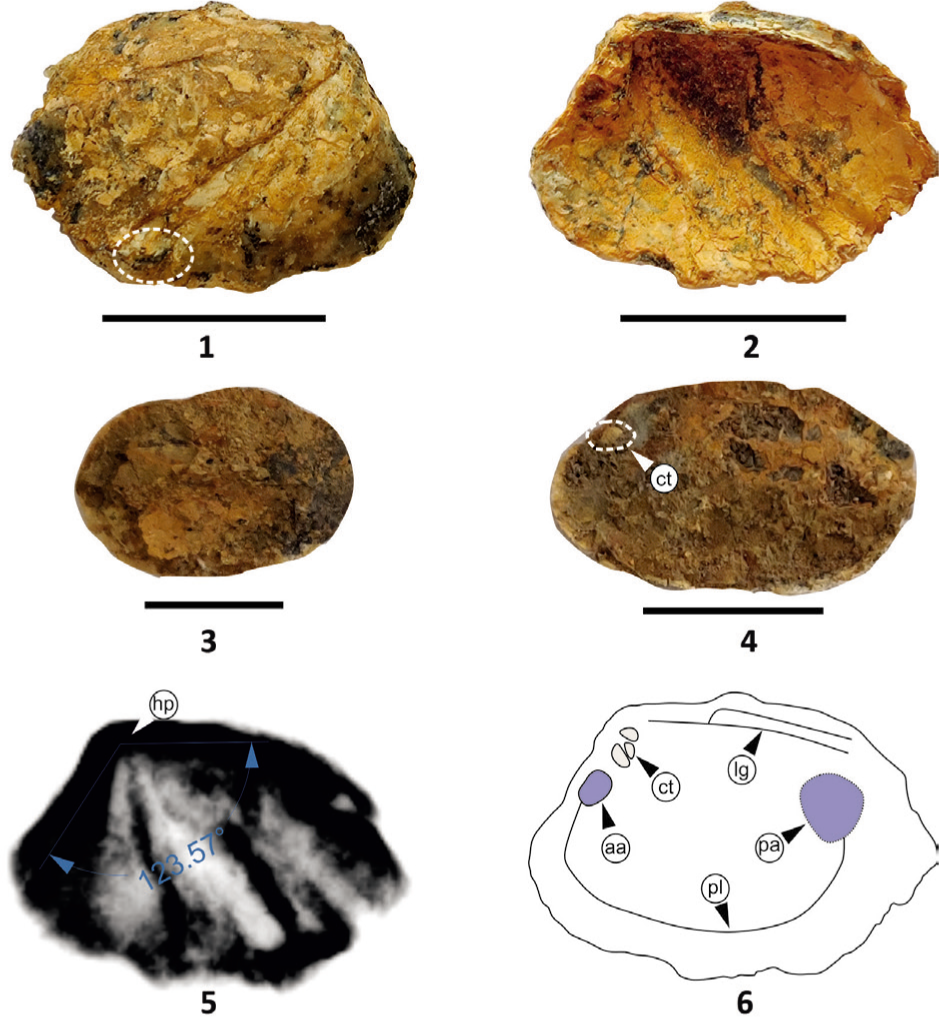
Genus Bamanboria oblongis gen. et sp. nov. (Plate 5, Figures 1–11)
Bamanboria oblongis sp. nov; Figs. 1–3, Holotype: Fig. 1, External view of right valve; Fig. 2, Internal view of right valve with umbonal angle; Fig. 3, Ventral view with highly corrugated commissure and tubercles; Figs. 4–9, Paratype: Figs. 4–6, Internal moulds; Figs. 7–8, External and internal view with cardinal teeth; Fig. 9, External view; Fig. 10, X-ray of holotype; Fig. 11, Internal shell morphology illustrated from the holotype. (ct = cardinal teeth, aa = anterior adductor muscle scar, lt = ligament tooth, lg = ligament, pl = pallial line, pa = posterior adductor muscle scar). All scale bars represent 1 cm.

Material: Two right valves, one complete specimen and three moulds are preserved.
Etymology: The species name refers to its elongated nature (Latin oblongus, meaning elongated, length is considerably more than width and thickness of the shell).
Holotype: Geo/Mus/BF/XVI (Plate 5, Figures 1–3)
Paratype: Geo/Mus/BF/XVII (internal mould), Geo/Mus/BF/XVIII (internal mould), Geo/Mus/BF/XIX (internal mould), Geo/Mus/BF/XX, Geo/Mus/BF/XXI, (Plate 5, Figures 4–9).
Diagnosis: Highly elongated with very wide hinge angle, >160°, one-half height-to-length ratio, small posterior wing and distinct corrugations towards the posterior margin.
Description: Equivalve, inequilateral, elliptical to oblong shell, the length of the valves is nearly twice its height ranging from 1.6 to 5.9 cm (Plate 5, Figures 1, 4–9). The shell is thick but flattened (Plate 5, Figures 1, 4–9). Umbo is prominent, extremely anterior and bears two cardinal teeth, lateral teeth are long and extended quite to the posterior margin (Plate 5, Figures 1, 8). The ligament is long, extending up to 2/3 length of the valve, opisthodetic and parivincular ligament (Plate 5, Figure 1) with a 175° umbonal angle (Plate 5, Figure 10). Anterior and posterior muscle scars are unequal and faintly preserved. The pallial line is straight and prominent. The shell shows allometric growth, on the anterio- and posterior-dorsal margin, young shells are oblong (Plate 5, Figures 1–2) while the adult is elliptical (Plate 5, Figures 7–8) in shape. The ventral outline of the shell has distinct corrugations with tubercles (Plate 5, Figures 1, 3, 7), and size increases towards the posterior side. Small anterior and posterior wings are present in the young shell (Plate 5, Figures 1–2). The valves have distinctly marked growth lines and the ventral margin is symmetrically corrugated (Plate 5, Figures 1–2, 7–9).
Dimensions: see Table 4.
Dimensions of Bamanboria oblongis gen. et sp. nov.
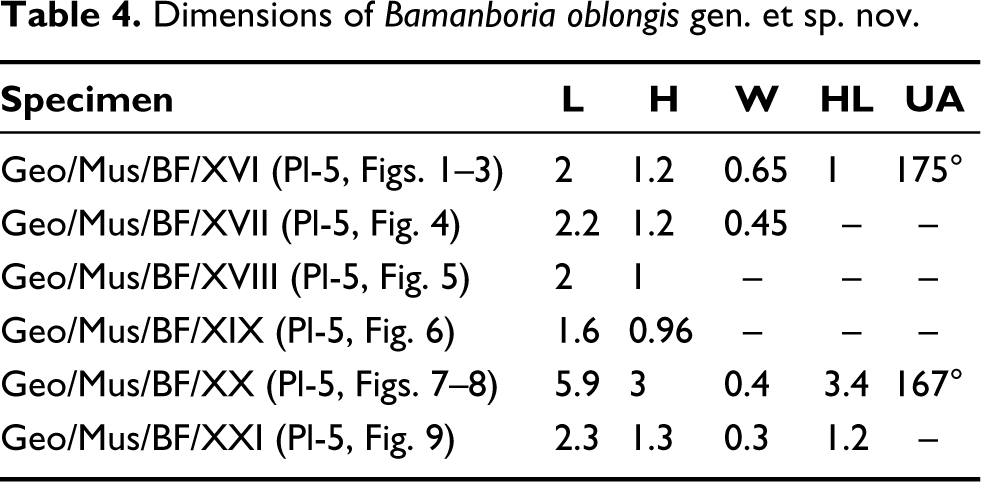
Remarks: B. oblongis gen. et sp. nov. is characterised by its elliptical to oblong shape and a low posterior wing compared to Deccanoida nov. Several internal moulds are observed, showing faithful outlines with identical ventral corrugations, for example, (i) Plate 5, Figure 4—left valve mould/nacreous layer which is elliptical/oblong shows subcentral anteriorly placed umbo. The anterior adductor muscle scar and teeth impressions are distinct (Plate 5, Figure 2, 4, 11). The posterior part shows well-preserved coarse corrugations; (ii) Plate 5, Figure 5—right valve mould/nacreous layer which is elliptical shows an anteriorly placed umbo and the internal mould is smooth; and (iii) Plate 5, Figure 6—the single left valve mould/nacreous layer which is elliptical shows subcentral umbo. The internal mould is smooth.
Type locality: Bamanbor, Chotila–Rajkot Highway, Gujarat, India.
Type horizon: Bamanbor Formation.
Subfamily Chotiloinae nov.
Diagnosis: Trapezoidal to elliptical shape of specimens, equivalve or inequivalve with two diverging cardinal teeth and corrugated commissures.
Genera included: Chotilia gen. nov.
Genus Chotilia nov.
Type species: Chotilia trappeansis gen. et sp. nov.
Etymology: The genus name refers to the name of the geographic location where the type species was reported (Town: Chotila).
Diagnosis: As for the type species.
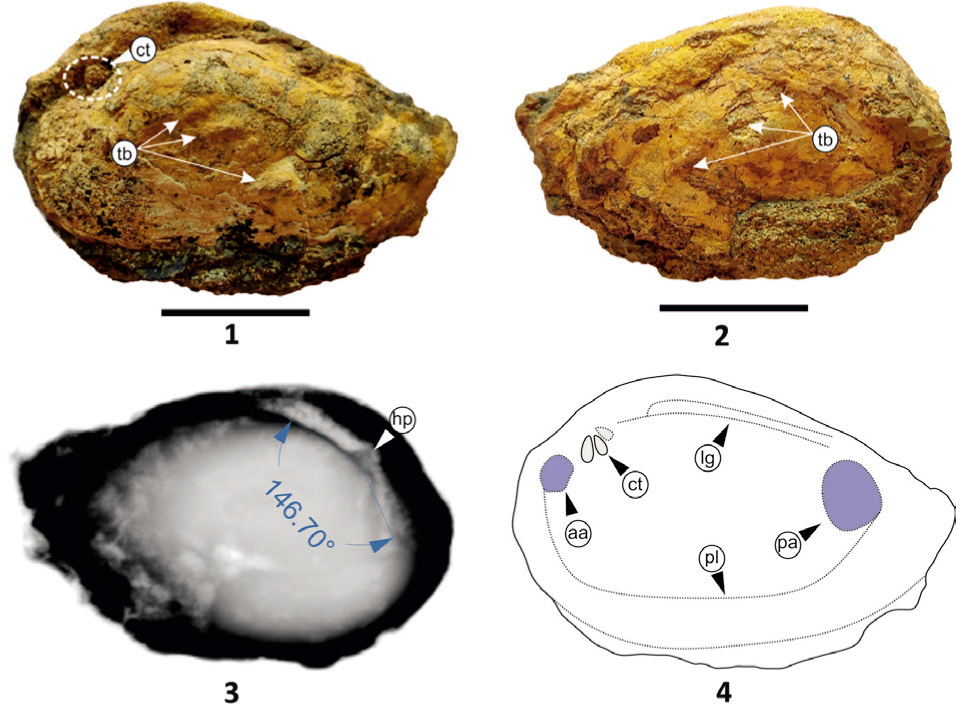
Genus Chotilia trappeansis gen. et sp. nov. (Plate 6, Figures 1–6)
Material: Three specimens, one articulated; and two disarticulated shells, among one right valve and one left valve, are preserved.
Etymology: The species name refers to the name of the sandwiched sedimentary sequences between two volcanic lava flows, deposited during the quiescence period of Deccan Volcanic activity (intertrappeans).
Holotype: Geo/Mus/BF/XXII (Plate 6, Figures 1–2).
Paratype: Geo/Mus/BF/XXIII, Geo/Mus/BF/XXIV (Plate 6, Figures 3–4).
Diagnosis: Trapezoidal shell, with low obtuse hinge angle, two prominent cardinals as well as one lateral tooth. The valves are uniquely characterised by the placement of a single tubercle on the posterior-ventral margin along with distinct corrugations and posterior wings.
Description: Trapezoidal, non-inflated and inequilateral, equivalve (Plate 6, Figures 1–2). The posterior wing is prominent and elevated, with a distinct opisthodetic parivincular ligament (Plate 6, Figure 2). Umbo is prominent, extremely anteriorly placed and has a 123° umbonal angle (Plate 6, Figure 5); and two cardinal and one lateral tooth on the posterior side (Plate 6, Figures 2, 4). Anterior muscle scar leaves a small depression with a faint pallial line. The posterior wing is distinct (Plate 6, Figure 2). One tubercle on the posterior-ventral side with a conspicuous wavy corrugated margin (Plate 6, Figure 1). Growth lines are broad, faint and distinct ventrally.
Chotilia trappeansis sp. nov; Figs. 1–2, Holotype: Fig. 1, External view of right valve; Fig. 2, Internal view of right valve; Figs. 3–4, Paratype: Fig. 3, Left valve view; Fig. 4, Right valve view and also shows prominent cardinal teeth fused; Fig. 5, X-ray of holotype with umbonal angle and hinge plate; Fig. 6, Illustrates the internal shell morphology of the holotype. (ct = cardinal teeth, aa = anterior adductor muscle scar, lt = ligament tooth, lg = ligament, pl = pallial line, pa = posterior adductor muscle scar). All scale bars represent 1 cm.
Dimensions: see Table 5.
Dimensions of Chotilia trappeansis gen. et sp. nov.

Remarks: The trapeziform shape, distinct high perivincular opisthodetic ligament and presence of tubercles in C. trappeansis gen. et sp. nov. differentiate from the other species of the genus Deccanoida and Bamanboria of the Bamanbor Formation. The complete species is reconstructed with the internal features (Plate 6, Figure 6).
Type locality: Bamanbor, Chotila–Rajkot Highway, Gujarat, India.
Type horizon: Bamanbor Formation.
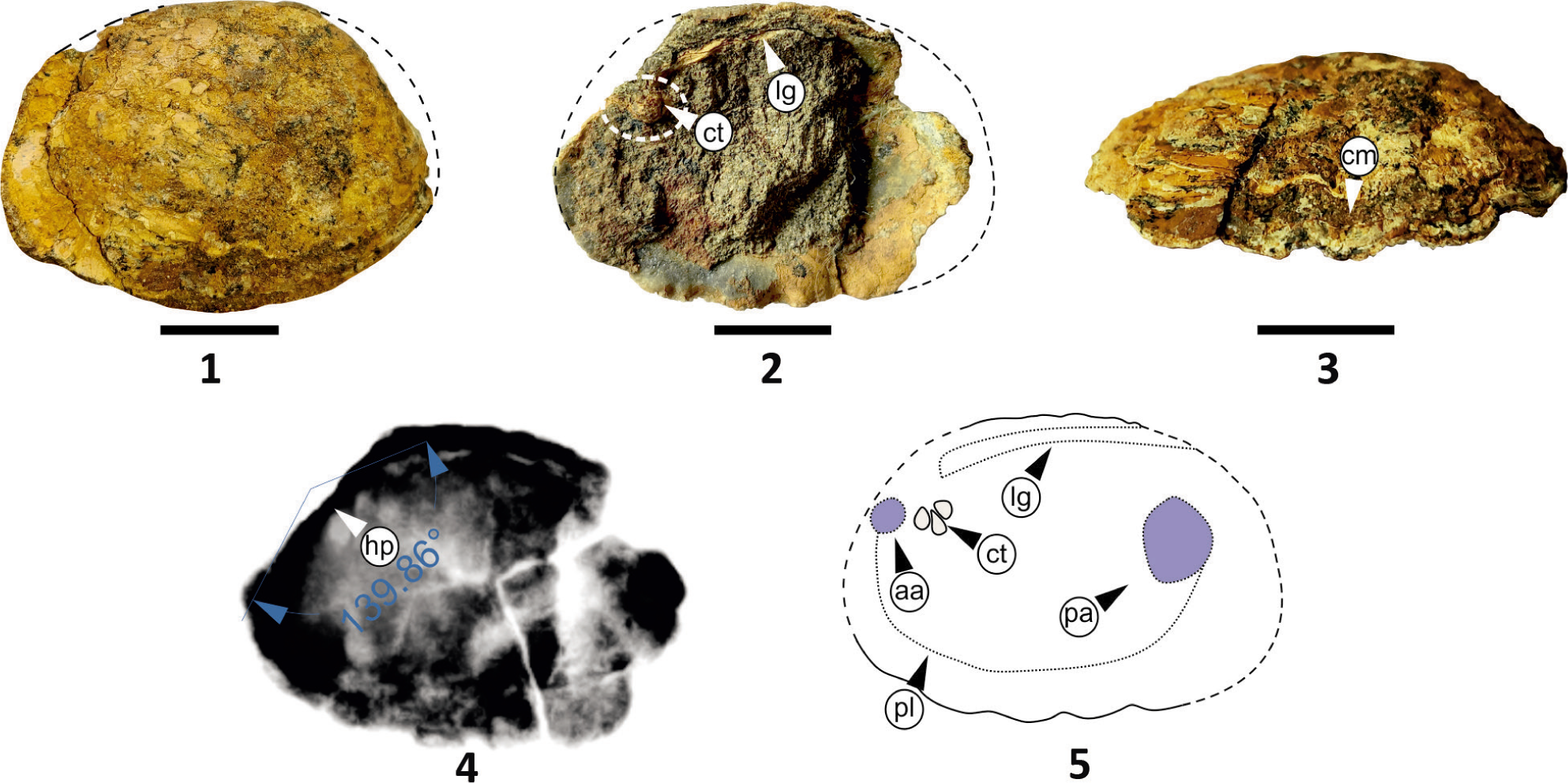
Genus Chotilia tuberculata gen. et sp. nov. (Plate 7, Figures 1–4)
Material: Well-preserved articulated shell.
Etymology: The species name refers to the presence of single-row tubercles (Latin tūberculum meaning lump) on both valves.
Holotype: Geo/Mus/BF/XXV (Plate 7, Figures 1–2).
Chotilia tuberculate sp. nov; Figs. 1–2, Holotype: Fig. 1, Left valve external view showing prominent tubercles and cardinal teeth of right valve; Fig. 2, External view of right valve; Fig. 3, X-ray of holotype with hinge plate and umbonal angle; Fig. 4, Line drawing of the holotype shows the various internal morphological features. (ct = cardinal teeth, aa = anterior adductor muscle scar, lt = ligament tooth, lg = ligament, pl = pallial line, pa = posterior adductor muscle scar, tb = tubercle). All scale bars represent 1 cm.
Diagnosis: Trapeziform to cuneiform bivalve with low obtuse hinge angle, and sub centrally placed umbo. Prominent tubercles at the centre of the valve in a single row from the umbo to the ventral margin. Moderately developed rostrum.
Description: It is a complete specimen, trapeziform to cuneiform in shape with inequilateral and equivalves (Plate 7, Figures 1–2). Valves are preserved in a slightly displaced form. Umbo is high and ventricose, placed sub-centrally on the anterior side, with a 146° umbonal angle (Plate 7, Figure 3); and a slightly curved, opisthodetic ligament is present (Plate 7, Figure 1). Two faint cardinal teeth on the right valve are present (Plate 7, Figure 1). A posterior wing with a moderately developed rostrum (Plate 7, Figures 1–2). The posterior end shows relics of ribs and corrugations. Corrugations are concentric towards the ventral margin. Tubercles are present in a single row on both valves and their size gradually increases from the umbo to the ventral margin (Plate 7, Figure 1). Growth lines are distinct and present on entire valves. The redrawn Figure from the X-ray (Plate 7, Figures 3 and 4) shows the important internal features.
Dimensions: see Table 6.
Dimensions of Chotilia tuberculata gen. et sp. nov.

Remarks: C. tuberculata gen. et sp. nov. is compared with Unio mammilatus, Hislop (1860) which is also a sub-cuneiform, inequilateral unionid with a single row of mamillae and elevated umbones. The presence of concentric growth lines on the entire shell, tubercles on both valves and the highly corrugated margin distinguish C. tuberculata gen. et sp. nov. from U. mammilatus Hislop (1860). Additionally, it features a low posterior wing compared to C. trappeansis gen. et sp. nov.
Type locality: Bamanbor, Chotila–Rajkot Highway, Gujarat, India.
Type horizon: Bamanbor Formation.
Genus Chotilia deccanensis gen. et sp. nov. (Plate 9, Figures 1–5)
Chotilia deccanensis sp. nov; Figs. 1–3, Holotype: Fig. 1, External view of right valve; Fig. 2, Internal view of the right valve with prominent cardinal teeth and opisthodetic parivincular ligament; Fig. 3, Ventral view with distinct coarsely corrugated commissure; Fig. 4, X-ray image with umbonal angle and hinge plate; Fig. 5, Line drawing of the shell shows internal morphological features of the holotype (ct = cardinal teeth, aa = anterior adductor muscle scar, lt = ligament tooth, lg = ligament, pl = pallial line, pa = posterior adductor muscle scar, cm = commissure). All scale bars represent 1 cm.
Material: One right valve is preserved.
Etymology: The species name refers to the name of the LIP wherein it was reported (Deccan Traps).
Holotype: Geo/Mus/BF/XXVI (Plate 9, Figures 1–3).
Diagnosis: Elliptical shell, ventricose umbo placed low on anterodorsally margin with thick hinge plate. Distinct concentric corrugations, towards the posterior margin.
Description: A thick right valve, elliptical in shape, probably equivalved but inequilateral (Plate 9, Figure 1). The umbo and beak are towards the anterior side but placed low (Plate 9, Figure 2) with a 139° umbonal angle (Plate 9, Figure 4). One lateral and two cardinal teeth are prominent with slightly curved parivincular opisthiodetic ligament (Plate 9, Figure 2). Concentric growth lines are present on the entire valve (Plate 9, Figure 1). In the ventral side, it is folded with costae which are reflected as corrugation (Plate 9, Figure 3). The reconstructed species shows unequal adductor muscle scars with prominent pallial lines (Plate 9, Figure 5).
Dimensions: see Table 7.
Dimensions of Chotilia deccanensis gen. et sp. nov.

Remarks: The unique elliptical shape is characteristic of C. deccanensis gen. et sp. nov. which is differentiated from the C. trappeansis and C. tuberculata. It is also different from L. vredenburgi Prashad (1921), redescribed from the Hislop and Hunter (1855) collections of the intertrappeans of Goraha, Narbada (Narmada), which is an elongated sub-rhomboidal, unionid with narrow posterior wing and rounded posterior and somewhat angulate anterior margin of the shell.
Type locality: Bamanbor, Chotila–Rajkot Highway, Gujarat, India.
Type horizon: Bamanbor Formation.
Geochemistry and Palaeoenvironmental Aspects
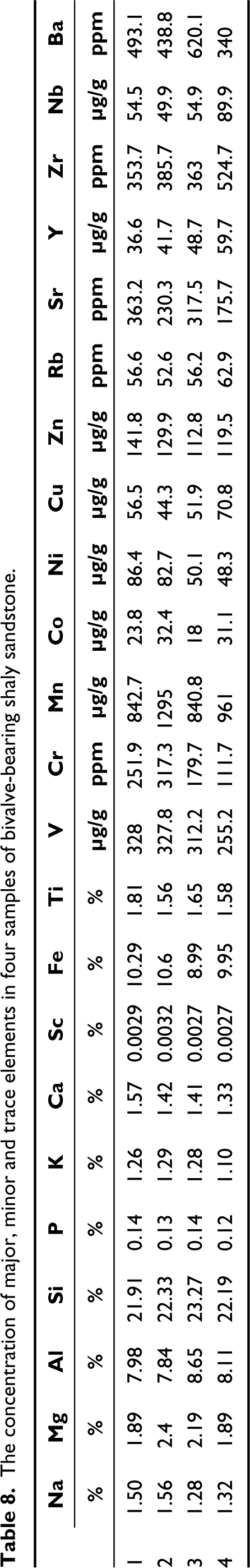
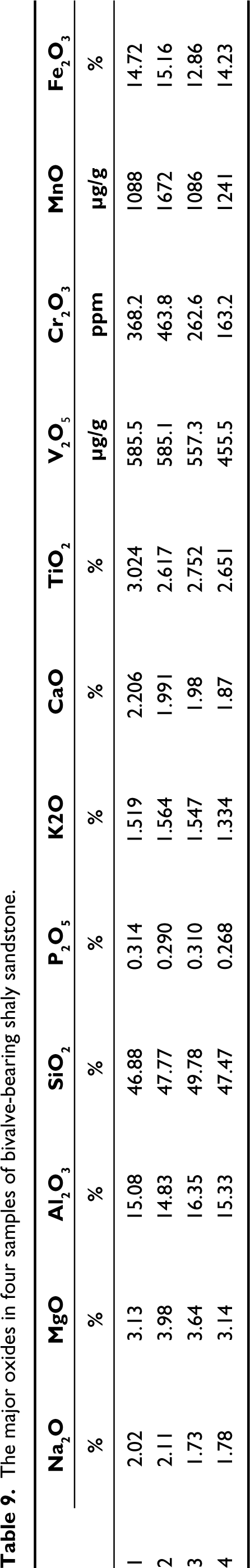
The composition of major oxides and various inorganic geochemical elements (major, minor and trace elements) was determined using an XEPOS HE XRF spectrometer. The concentration of elements like Na, Mg, Al, Si, P, K, Ca, Sc, Fe, Ti, V, Cr, Mn, Co, Ni, Cu, Zn, Rb, Sr, Y, Zr, Nb, Ba and major oxides like Na2O, MgO, Al2O3, SiO2, P2O5, K2O, CaO, TiO2, V2O5, Cr2O3, MnO, Fe2O3 were determined. Four samples of shaly sandstone which bears the assemblage of D. conrugis gen. et sp. nov., D. aleta gen. et sp. nov., D. costaria gen. et sp. nov., B. oblongis gen. et sp. nov., C. trappeansis gen. et sp. nov., C. tuberculata gen. et sp. nov. and C. deccanensis gen. et sp. nov. were analysed, to infer the probable paleoenvironment of deposition. Tables 8 and 9 summarise the average concentration of major, minor and trace elements, and major oxides, respectively. Table 10 shows the standard and observed value of the various paleoenvironmental parameters.
The concentration of major, minor and trace elements in four samples of bivalve-bearing shaly sandstone.
The major oxides in four samples of bivalve-bearing shaly sandstone.
The standard and observed average values of various palaeoenvironmental parameters were calculated using elemental and oxide ratios.
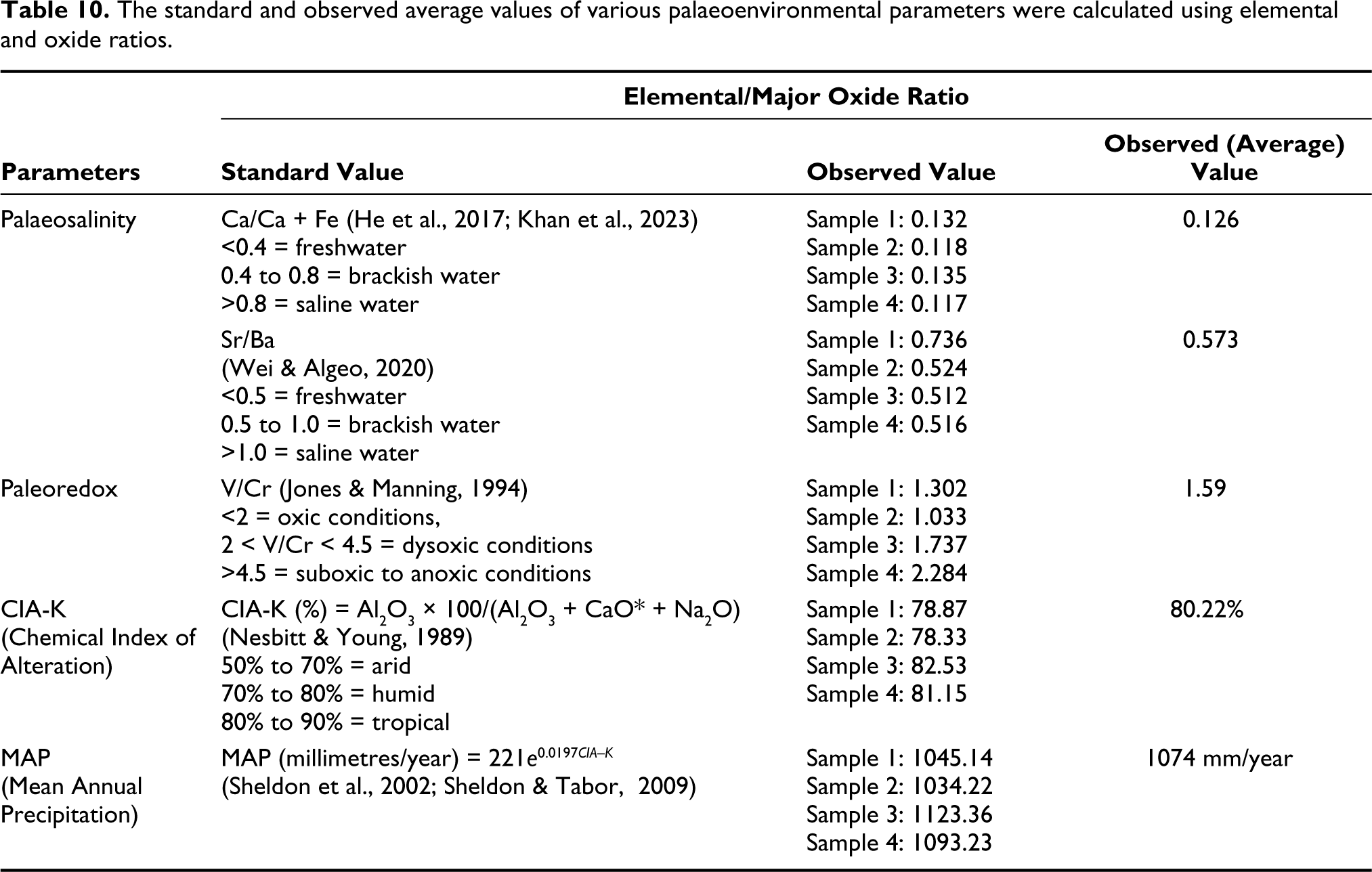
The Ca/Ca + Fe ratio (He et al., 2017; Khan et al., 2023) ratio and Sr/Ba ratio (Wei & Algeo, 2020) can serve as an indicator of salinity levels (Table 10). The Ca/Ca + Fe ratio of 0.8 corresponds to the saline water, ratios ranging from 0.4 to 0.8 indicates brackish water, and <0.4 indicates freshwater (He et al., 2017; Khan et al., 2023). The Sr/Ba ratio of <0.5 suggests freshwater, 0.5–1.0 suggests brackish water, and >1.0 suggests saline water (Wei & Algeo, 2020). The calculated ratios of Ca/Ca + Fe and Sr/Ba are 0.126 and 0.5725, respectively, the former suggests freshwater while the latter values suggest a brackish water environment. The presence of carbonate within the shaly sandstone may affect the Sr/Ba ratio (Eric et al., 2022; Wang et al., 2021), however, the percentage of CaO in the samples ranges from 1.87 to 2.206 (Table 9) which may be insignificant. Thus, based on both the standards, a wider range of salinity, fresh to brackish water conditions is considered for the bivalve bearing shaly sandstone horizon. For paleoredox conditions, the V/Cr ratio (Jones & Manning, 1994) calculated is 1.589, which suggests an oxic environment (<2 indicates oxic conditions, 2 < V/Cr < 4.5 indicates dysoxic conditions and >4.5 indicates suboxic to anoxic conditions [Jones & Manning, 1994]).
The Chemical Index of Alteration (CIA-K) is a measure of chemical weathering caused by climatic changes. It quantifies the degree of weathering by analysing the molar proportions of Al2O3, CaO and Na2O. It excludes potassium (K) due to potential diagenetic processes that can lead to higher K concentrations (Adams et al., 2011; Sheldon et al., 2002). The CIA-K calculation, based on the work of Nesbitt and Young (1989), uses the percentage of aluminium oxide (Al2O3) to the sum of Al2O3, corrected calcium oxide (CaO*) and sodium oxide (Na2O). For accounting for the presence of carbonates, a CaO correction is applied, assuming that CaO* is equivalent to Na2O if the values of CaO exceed Na2O.
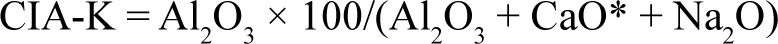
Additionally, mean annual precipitation (MAP) is determined quantitatively using CIA-K values, which provides insights into the relationship between weathering severity and precipitation. The MAP is calculated using a formula derived by Sheldon and Tabor (2009), which incorporates the CIA-K values to estimate annual precipitation in millimetres.

The CIA-K value is 80.203 and MAP is 1074 mm for the bivalve bearing assemblage shaly sandstone. It suggests a humid condition and moderate to high precipitation. The value >80 suggests moderate to intense chemical weathering in the source area which also reflects a humid climate with moderate to high precipitation (Adams et al., 2011; Sheldon et al., 2002; Sheldon & Tabor, 2009).
Palaeobiogeography and Evolution
Early Evolution in Palaeozoic
The earliest known occurrence of the nonmarine bivalve, Archanodon can be traced back to the Bellvale Sandstone in New Jersey, North America, during the Middle Devonian period. This finding, reported by Chamberlain and Chamberlain (2007), provides the first insight into the existence of the oldest known member of freshwater bivalve, Archanodon from the Middle Devonian-Late Carboniferous within Laurussia, a relatively minor supercontinent located in the northern hemisphere (Cocks & Torsvik, 2011). Species A. cataskillensis was reported from channel fill and over-bank deposits of the Catskill Formation (Chamberlain & Chamberlain, 2007). For its widespread presence in Catskill rocks, it was suggested to be classified in the order Unionida while eventual taxonomic revision was also suggested (Chamberlain & Chamberlain, 2007). The Archanodon and Anthracosiidae from the Upper Carboniferous, UK (Eagar, 1953), survived in the assembling early Pangea supercontinent (Cocks & Torsvik, 2011). The nonmarine bivalves Palaeomutela and Palaeoanodonta (Silantiev & Carter, 2015) became widespread during Permian and are reported from Laurentia and Gondwana of the accreting Pangea supercontinent; and Microdonta in the USSR (Cox et al., 1969). Nonmarine Autobranch (Palaeoheterodont) genera Gondwanaiadites, Bakulia, Gangamya, Raniganjelia, Gondwanadontella and Indonellina are reported from the Permian of the Raniganj Formation of the Raniganj, Jharia and North Karanpura basins of the Damodar valley from the Indian Gondwana Coal Measures (Silantiev et al., 2015).
The Late Permian/Early Triassic extinction had severely affected nonmarine bivalves, especially the endemic taxa, suggesting that endemics are more vulnerable during mass extinctions (Yan et al., 2023), the Triassic is represented by new stocks of freshwater bivalves. The harsh environmental conditions followed into the Triassic after the Permian-Triassic Mass Extinction (PTME), which are reflected in the imperfect early Triassic ecosystems and a resultant Lilliput effect in the animals (Chen et al., 2019; Erwin, 2006; Retallack, 1995; Rodland & Bottjer, 2001). As a consequence, there are several gaps in the fossil record of the Early Triassic that attest to the absence of several niches during this inimical stage, a well-known example is the absence of coals from the Early Triassic (Retallack et al., 1996). Bivalve molluscs re-invaded freshwaters soon after the End-Permian extinction event, during the earliest part of the recovery phase during the Olenekian Stage of the Early Triassic. The unionoid-like bivalve reports from the Early Triassic, preserved in vertebrate coprolites reported from the fluvial-lacustrine lower Burgersdorp Formation, Karoo Basin, South Africa are the first reports post PTME (Yates et al., 2012). These unionids were part of the bivalve reinvasion of freshwaters after the End-Permian extinction event had wiped out earlier freshwater bivalve families and its clade remained a major part of freshwater habitats throughout the rest of the Mesozoic and Cenozoic Eras (Yates et al., 2012).
A silesunionid, T. karooensis Cox (1932) (Figure 2) is also reported from the Middle Triassic Manda Beds of Tanzania and Ntawere Formation of Zambia (Cox et al., 1969; Van Damme et al., 2015). With subsequent dispersal in different continents, bivalve shows environmental, physiological and structural modifications resulting in their early diversification (Figure 2). Both supercontinents characteristically observed the freshwater bivalves of order Unionida, and this group became widespread except in Antarctica (Graf & Cummings, 2006, 2007).
Paleogeographic reconstruction of the Indian plate during its northward drift in the Tethys Sea during the Late Cretaceous, Palaeocene and Early Eocene, outlining the vicariance and dispersal events in freshwater unionid stocks of Africa, Madagascar and India (Modified after Smith et al., 1994). (Reconstructed using Corel Draw v. x8 and QGIS v. 3.30.0).
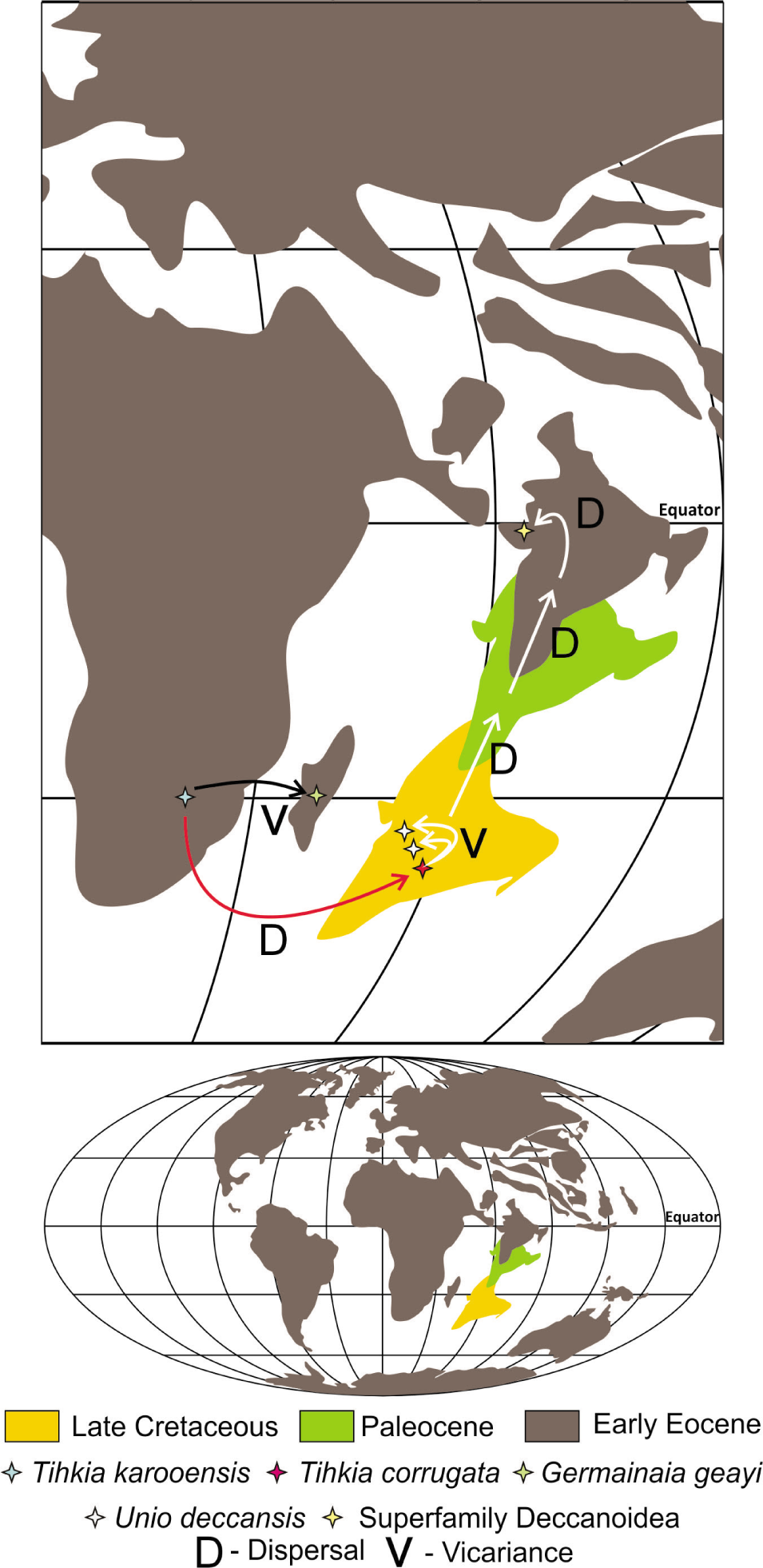
The earliest unionids evolved during the Late Triassic and subsequently dispersed in all the continents of the Pangea. Newell and Boyd (1975) suggested that the probable ancestors of the order—Unionida and Trigonida, eventually evolved in the late Triassic, with its first representatives reported from Arizona, North America. The oldest records at 230 m.y. from Carnian-Rhaetian deposits of Krasiejów clay pit, Silesia, southern Poland are vital. It has yielded numerous fossils from order Unionida, suborder Silesunionina, family Unionellidae and subfamily Silesunionidae, which are filibranch members with free-living larvae, comprising of S. parvus and Tihkia(?) silesiaca (Skawina & Dzik, 2011). The Indian subcontinent was part of the Pangea in the southern hemisphere (Briggs, 2003), and it was also an epicentre of the terrestrial deposits ranging in age from Lower Permian to Triassic. Many different types of terrestrial organisms were residing there, or crossing physical limits, with evolving endemic communities. The Middle Triassic Yerapalli Formation, Late Triassic Tiki Formation and Maleri Formation had yielded well-preserved and highly varied vertebrate fossils such as fishes, amphibians and several reptilian and mammalian amniotes (Mukherjee & Ray, 2012). The fauna of Yerrapalli Formation is known to consist of a Saurichthys fish (Jain, 1984), the dipnoian Ceratodus (Chatterjee, 1967), amphibian Parotosuchus (Roychowdhury, 1970a), two dicynodonts—Wadiasaurus and Rechnisaurus (Roychowdhury, 1970), rhynchosaur Mesodapedon kuttyi (Chatterjee, 1980), cynodont (Chatterjee et al., 1969), two archosaurs and a protosaurian (Jain & Roychowdhury, 1987). The Tiki Formation contains the dipnoian Ceratodus, the metoposaurid Metoposaurus maleriensis, the phytosaur Parasuchus hislopi (Chatterjee, 1978), rauisuchid Tikisuchus romeri, the cynodont Rewaconodon tikiensis along with a morganucodontid mammal Gondwanodon tapani, Tikitherium and Tikiguania estesi. Ray et al. (2016) reported rhynchosaur, Hyperodapedon tikiensis and large traversodontid cynodont Ruberodon roychowdhurii. The vertebrate fossils reported from the Maleri Formation include Ceratodus, a dipnoi fish, Metoposaurid amphibian M. maleriensisi, a rhynchosaur Paradapedon, a phytosaur Parasuclzus, an eosuchian Malerisaurus and a cynodont Exaeretodon are other important members of reptiles (Jain, 1996). Most noteworthy is the discovery of the saurischian dinosaur (Walkeria maleriensis) from Adilabad district in red mudstones facies (Jain, 1996). The Middle and Late Triassic vertebrate record marks distinction in certain basins suggesting endemic nature of the certain group of the animals.
Among the freshwater invertebrates, the Maleri Formation also yields ostracods (Sohn & Chatterjee, 1979) and bivalves (Sahni & Tewari, 1958). Ostracods belonging to families like Darwinulacea and Cytheracea are reported. The record of a freshwater bivalve Tihkia corrugate (Figure 2) of the Maleri Formation of India (Sahni & Tewari, 1958) is parallel with S. parvus and Tihkia(?) silesiaca from Krasiejów clay pit, Silesia, southern Poland (Skawina & Dzik, 2011), marks a crucial example of dispersal.
Evolution Across Gondwanaland
The breakup of Pangea and subsequent splitting of Gondwanaland in Early and Late Jurassic (Rogers & Santosh, 2004), foresees the allopatric speciation in the ancestral unionid stocks of Pangea. The records of the Unionids from the Middle to Late Jurassic from Africa are ubiquitous. It hosted some important endemic (Unionidae, Margaritiferidae, Tegulaedontidae and Tamesnellidae) and non-endemic (Unionelloides, Margaritifera) unionids (Van Damme et al., 2015). Among the invertebrates non-endemic unionids and vertebrates like freshwater fishes (Arratia et al., 2004b) and theropod dinosaurs (Belvedere et al., 2019 and references therein) are symbolic of the terrestrial (including freshwater, aquatic) connections between Europe and Gondwanaland, which had assisted the faunal exchange (Van Damme et al., 2015).
The Middle-Late Jurassic breakup of Gondwanaland (Rogers & Santosh, 2004), separated the India-Madagascar plate from the Somali coast of Africa, which experienced island continent-like biogeography in the southern Tethyan ocean. The record of Etheria elliptica, Unio geayi (Germainaia geayi), Unio madagascariensis, Unio malgachensis from the Mesozoic (Early Cretaceous) of Madagascar and Mascarene, and the evolutionary implications of G. geayi, as a possible vicariant lineage in the island, highly implies that the possibility of speciation of freshwater unionids by disjunct distribution in the separated Gondwanan continents (Figure 2) (Graf & Cummings, 2009). The freshwater deposits have a continuous record in India, after the breakup of Gondwanaland in the Late Jurassic till the Cretaceous. The Jurassic freshwater deposits recorded from Kota Limestone (Govindan, 1975) and Early Cretaceous deposits of thick fluvio-lacustrine deposit, Songir Formation in the Western Lower Narmada Valley (Shitole et al., 2021) lack the bivalves (body fossils) evidence possibly due to preservational constraints, suggesting a hiatus (Jurassic–Early Cretaceous) in the record of unionids in Indian subcontinent. However, a boring structure, Apectoichnus made by freshwater Teredinidae bivalves found in poorly bioturbated sandstone of the Songir Formation marked their presence (Shitole, 2022).
Evolution of Intertrappean Basins
Maastrichtian and Palaeogene intertrappean sequences of the Indian subcontinent are mainly developed in restricted topographic lows rather than structurally controlled basins. Major structurally controlled basins, particularly of Kachchh, Saurashtra and Narmada, where sedimentation was ceased due to rift failing (Biswas, 1987) and subsequent upliftment. During the Maastrichtian-Palaeogene time, the Insular Indian (now, the Indian subcontinent) detached completely from the Gondwanaland and experienced the outpouring of several lavas flows of reunion hot spot (Mahoney, 1988), which resulted in uneven topography. The topographic low led to the formation of sporadic perennial and ephemeral freshwater pools due to seasonal residual water drawing streams and became a host to various life forms. The pools, usually formed across the Cretaceous-Palaeogene, filled by the sediments were covered by the subsequent Deccan lava flows. As a result, it formed intertrappeans in the Indian subcontinent which is characterised by typical terrestrial sedimentary sequence governed by the local parameters and thence it also appears patchy (Blanford, 1872; Chenet et al., 2008; Khosla & Sahni, 2003), but mainly occurs throughout the periphery of the Deccan Volcanic Province (Kapur & Khosla, 2019). The sediment thickness of these units is also highly variable and comprises a wide range of terrestrial fossils of vertebrates, invertebrates, micro- and plant fossils (Kapur & Khosla, 2019; Khosla, 2015; Khosla & Sahni, 2003; Khosla & Verma, 2015; Prasad et al., 1995; Prasad & Khajuria, 1996; Samant & Mohabey, 2014; Wilson et al., 2007). Preliminary data is available on freshwater bivalves of intertrappeans but it lacks systematic attempt, the majority of the work is related to noting their occurrence in different localities. A systematic attempt by Hislop (1860) for the Late Cretaceous intertrappean freshwater bivalves (translated and revised by Hartman et al., 2008) recorded from the Nagpur and Narmada which includes, six Unionidae species, U. malcolmsoni Hislop, 1860 (new name described for U. tumidus Retzius, 1788 and U. tumida J. Sowerby, 1840), U. deccanensis J. Sowerby, 1840, U. hunteri Hislop, 1860, U. mamillatus Hislop, 1860, U. imbricatus Hislop, 1860, U. carteri Hislop, 1860, and P. medlicottianum Hislop, 1860; Lamellidens is also reported by Gangopadhyay et al. (2011) from Deori, Madhya Pradesh, India. P. medlicottianum Hislop, 1860 is redefined as L. vredenburgi by Prashad, 1921. Srivastava and Kandwal (2013) also reported U. deccanensis from the Maastrichitian intertrappean rocks of the Hiradehi and Topidhana area, Betul District, Madhya Pradesh (Figure 2).
Unfortunately, the freshwater bivalves of India have received limited attention, despite the coexistence of a globally endangered taxonomic group within a region widely acknowledged as a biodiversity hotspot during the Late Cretaceous (Joshi & Karanth, 2013). The sporadic occurrence of ephemeral pools might have restricted the distribution of the bivalve, moreover, their low abundance and diversity, and the delicacy of the shells, further constrained the preservation. The preservation is further compromised by the predominant fine-grained shaly nature of intertrappeans, and palaeoenvironmental conditions, particularly with fluctuating salinity which might have dissolved the shells or obscured the major morphological features. As a result, less attention has been paid to the freshwater bivalves of the intertrappeans of India, which has resulted in an evolutionary gap. Thus, this geographical area exhibits the highest suitability for the long-term viability of freshwater mussels, owing to their requirement for maintaining a calcareous exoskeleton and their reliance on fishes as hosts for their parasitic larvae (Kremen et al., 2008).
The Disjunct Distribution of Bivalve
The Deccan intertrappeans of India comprise unique new stock as compared to Gondwana and Laurasia (Hartman et al., 2008; Hislop, 1860; Prashad, 1921). During the northward drift of the Indian plate, Insular India experienced extensive lava flows of the Deccan Continental Flood Basalts, which covered nearly 500,000 sq. km in western and central India. These lava flows changed the ecosystem, climate and physiography of the terrestrial realms across the K-Pg. It is highly implausible to compare the endemically evolved freshwater bivalves on the Indian continent after its separation, either with Gondwana or with Laurasia. Across the Cretaceous and Palaeocene, Insular India aided in the disjunct distribution of freshwater bivalves (Figure 2). The evidence of Insular India acting as sweepstakes, Noah’s ark as well as stepping stones, for the biotic exchange between Insular India and Asia and the presence of Gondwanan elements suggesting vicariance event as well as exchange through Greater Somalia and Oman-Kohistan-Dras Island arcs are not less (Jacobs et al., 2011; Khosla & Lucas, 2020; Verma et al., 2012, 2016).
Up till now the freshwater bivalves of the K/Pg intertrappean localities of the north and eastern part of the Deccan province are paid attention to (Gangopadhyay et al., 2011; Hartman et al., 2008; Hislop, 1860; Srivastava & Kandwal, 2013). The records of the Upper Cretaceous unionids, from the intertrappeans of Nagpur, Narmada, Amravati, Betul and Deori must also be considered as different stocks, and more materials are to be required to differentiate them from the other established categories. These stocks of genus Unio and Lamellidens are evident of the vicariance in the Late Triassic stocks (genus Tihkia) (Figure 2), which is also documented in various vertebrate remains from Gondwana and Insular India, suggesting the possibility of new endemic stocks (Karanth, 2021; Toussaint et al., 2017; Van Bocxlaer et al., 2006; Verma et al., 2016). There is a long gap in the records of freshwater bivalves after Tihkia. The Late Cretaceous freshwater bivalves show different stocks comparatively, due to their highly endemic nature because of the constantly changing landscape as well as changing palaeoecological parameters due to intense volcanic activity. To cope with the changing conditions organisms may have evolved and modified their shells. More detailed studies will fill the evolutionary gap in the Late Cretaceous freshwater bivalve phylogeny.
In the Saurashtra Peninsula, the lake deposits which were hospitable for the different kinds of organisms in the palaeotopographic low of Deccan traps were covered by an eventual lava flow that occurred after a long cessation period. This intertrappean sequence dated based on palynomorphs suggests the Early Eoceneage. For the first time, seven different species of the order-Unionida, superfamily-Deccanoidea are recovered from the shaly sandstone, which is described and their affinity is discussed in context to their biogeography. Though the taxa recovered are meagre nonetheless their presence is significant as they have never been reported from the study area earlier as well as their counterparts; hence, they throw light on terrestrial ecosystems. There is not a single reference available on freshwater bivalves of the Saurashtra intertrappeans. The superfamily Deccanoidea nov. is the new evidence of dispersal in the Insular Indian unionids, further corroborated by changes in the ecological condition of intertrappeans by the waning phase eruption of Deccan Traps.
During the Middle and Late Eocene, the Indian Plate made a connection with Mainland Asia via Burma terrain which acted as a Gondwanan biotic ferry (Bolotov et al., 2022). Before this connection, it exhibited a unique biogeography, characterised by a vast island continent. This isolation led to the evolution of numerous species that were restricted to the Indian landmass. They evolved independently, and played a major role in shaping the distribution of India’s present-day biota (Rao, 1989), which likely originated from Gondwana through vicariance and dispersal, and spread across the Indo-China via Indo-Malaya, Iran-Afghanistan blocks and Kohistan-Dras Island arc (Bhatia et al., 1996; Verma et al., 2016), supporting the ‘Out of Africa Model’ (Hoeh et al., 2002).
Dispersal Mechanism
The superfamily Deccanoidea nov. provides interesting insights into the vicariance and dispersal mechanisms for the geographic isolation and evolution of unionids. Two different types of ectozoochronous mechanisms are considered for the dispersal of Deccanoidean unionids, the role of fish and the role of birds (Figure 3). The Bamanbor Formation had evidenced the fish fossils, first reported by Fedden (1884) was the fragmentary record of fossil fish skeletons from the Bamanbor–Navagam area. Later, Borkar (1973, 1975, 1984) reported fish remains of P. chiplonkari, H. intertrappean, P. feddeni, Perca cf. angusta from the cherty shales of the Bamanbor area and the latest record of Arratia et al. (2004a), reports evidence of Indiaichthys bamanborensis and Percromorpha indet. These freshwater fishes reside in rivers as well as stagnant water bodies like lakes. CIA-K and MAP suggest moderate rainfall which is capable of flooding the river and lake, the flooded lake water is temporarily connected with the river and provides a route for the migration of fish. In the early stage of the unionid, a glochidium larvae has a phoretic relationship with fishes, where they attach to the fish gills or fins, till they undergo metamorphosis into juveniles, and drop off when suitable conditions for survival are attained (Bauer & Wächtler, 2001). During the Early Paleogene time, the fish played a major role in dispersing these Deccanoidean glochidia larvae to the lake (Bauer & Wächtler, 2001).
Model illustrates the dispersal mechanism of bivalves by freshwater fishes and/or birds.
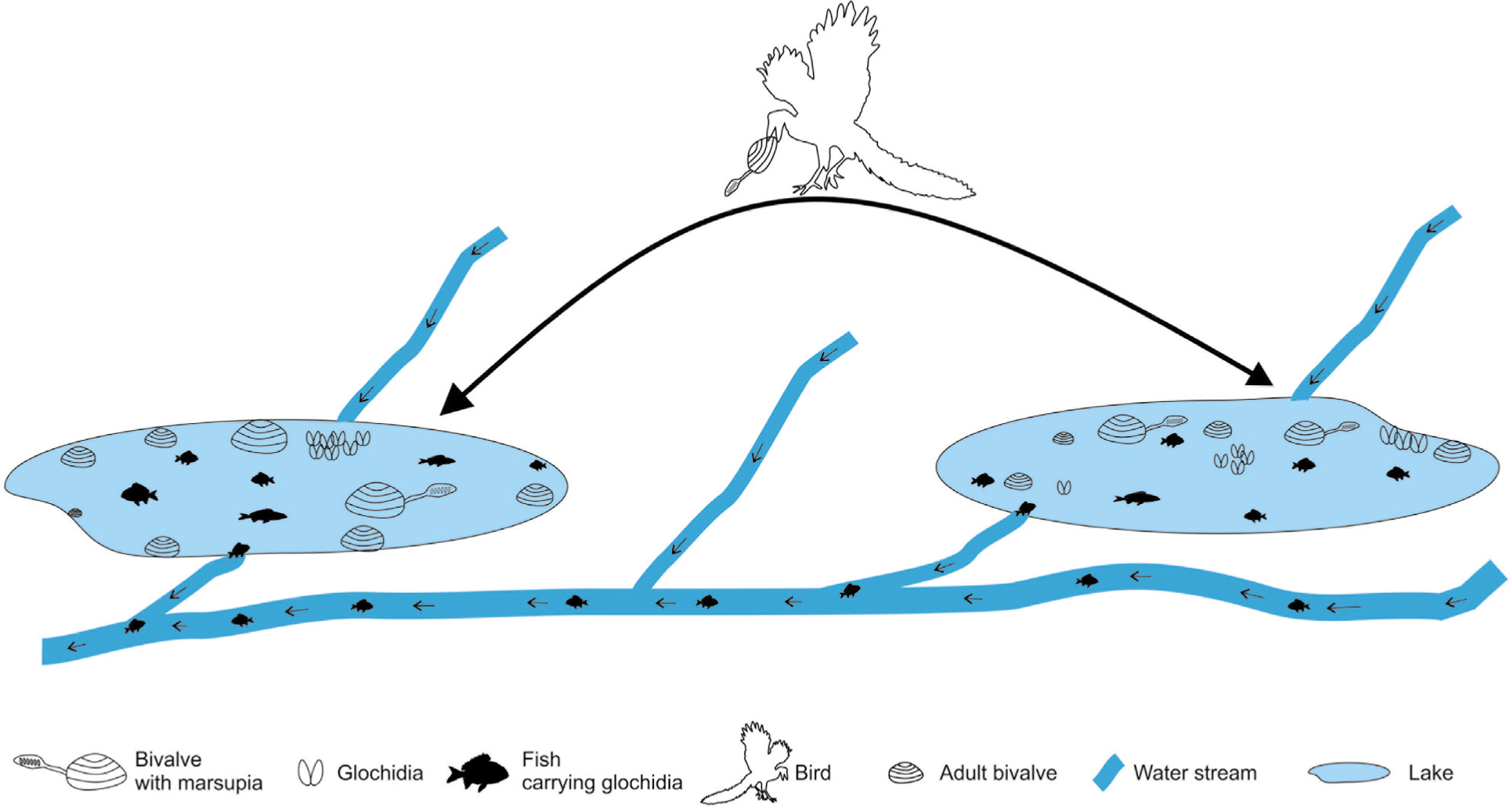
In another mechanism of dispersal, the role of the birds is considered. Many shell-eating birds, pick the shell in their beak and take them farthest on the land area (shell clusters/debris bivalve with gastropod shells observed in the interior of the land) and break the shells with the impact or pick the shells by beak to eat the flesh. While moving the adult mothers with fertilised brood pouches (marsupia), they may slip from their beaks and fall into a water pool where there is the possibility of propagation. The fortuitous event may play a role in the dispersal of bivalve fauna by sweepstakes dispersal.
The elemental and oxide ratios indicate that lake water was fresh and in oxic condition, and it existed in a humid environment where the water was received and renewed regularly. This optimum condition prevailed during the Early Eocene time when bivalves were subjected to phenotypic variation. Later on, they flourished and evolved into a new stock, due to new ecological settings compared to their ancestor in the Deccan Trap country. The bivalve of the intertrappean of Saurashtra indicates a dispersal, where a small geographic barrier of topographic high is crossed by any of the means as discussed.
Conclusions
The waning phase of Deccan volcanics comprises unique bivalve assemblages as compared to surrounding freshwater habitats of the Deccan Volcanic Province of India. During the Early Eocene, the Indian Plate was a huge island continent and its biota experienced allopatrics speciation, which resulted in disjunct distribution and has enabled us to draw the following conclusions.
The Chotila intertrappeans of the Saurashtra Peninsula consist of new stocks of freshwater bivalves comprised of a superfamily, family, 2-subfamily, 3-genus and 7-species of order Unionida. The higher-level category includes the superfamily Deccanoidea nov. consisting of one family, Deccanoidae nov., comprising two sub-families, Deccanoinae nov. and Chotiloinae nov. Three genera Deccanoida nov., Bamanboria nov. and Chotilia nov. include seven species, D. conrugis gen. et sp. nov., D. aleta gen. et sp. nov., D. costaria gen. et sp. nov., B. oblongis gen. et sp. nov., C. trappeansis gen. et sp. nov., C. tuberculata gen. et sp. nov. and C. deccanensis gen. et sp. nov.
Based on palynoflora and fish remains suggest the Early Palaeogene (Palaeocene-Early Eocene) age for the Chotila Basin intertrappean.
The major, minor and trace elements and oxides analysis of the bivalve bearing beds reveals fresh-brackish water conditions, oxic and humid environment and moderate precipitation.
The new unionid species of the Chotila paleolake were a result of the following disjunct mechanism. Stage I: The early dispersal of unionids on Gondwanaland during the Triassic. Stage II: Followed the vicariance in the unionids of Africa, Madagascar and Mascarene in the Early Cretaceous when the Indian Plate experienced island continent biogeography. Stage III: The latest dispersal on Insular India in Late Cretaceous–Early Paleogene (Palaeocene-Early Eocene) intertrappean sequences of Deccan Volcanic Province.
The newly erected freshwater unionid bivalve superfamily and genera therein are examples of disjunct distribution in Insular India during the Early Palaeogene. These stocks bridge the gap in the understanding of their evolutionary history between Gondwana, Insular India and Asia. The present evidence also affirm ‘Out-of-Africa’ or ‘In-to-Insular India’ model. After the Indian–Eurasian plate convergence they may have propagated into Asia during the remaining period of the Cenozoic Era.
Footnotes
Acknowledgements
The authors would like to thank Prof. Mukund Sharma of the Journal of the Palaeontological Society of India and the anonymous reviewers for the fruitful comments that helped improve the quality of this paper. The authors are indebted to the Head of the Department of Geology, for providing the necessary facilities to carry out the field and laboratory work. We extend our thanks to Dr Siddharth Prizomwala, Institute of Seismological Research, Gandhinagar, for granting us access to the XEPOS HE XRF facility. His support and access to this advanced equipment have significantly enhanced our research. We are thankful to Mr Mahendra Sutaria, ONGC Ltd., Vadodara for his help in identifying the palynofossil and to Mr Mohmmad Hussain, for preparing the thin sections.
Declaration of Conflicting Interests
The authors declared no potential conflicts of interest with respect to the research, authorship and/or publication of this article.
Funding
The authors received no financial support for the research, authorship and/or publication of this article.