
Abstract
Due to natural selection, an increase in biotic and abiotic factors develops an adaptive response in bivalves (molluscs) for survival. Morphological variation in a bivalve is strongly correlated with the changes in the structural and compositional properties of a shell. Therefore, the mechanical characteristics of a shell reflect its morpho-functional adaptation against predation and ecological stresses. An initial investigation into the primary mechanical attributes of predominant bivalve shells from the east coast of India, Tegillarca granosa, Meretrix meretrix, Sunetta meroe, Mactra turgida and Donax scortum, has been documented in this study. The quantitative assessment of the centreline thickness, Vickers hardness and density variation of the shells were measured and correlated along with the preliminary investigation of the microstructural patterns. The results from our study are in agreement with the most vulnerable region of a shell, located just below the umbonal zone, which has been verified through predatory boreholes in other studies. Our data shows a high correlation between shell hardness and density, which could also be the potential properties to consider in detail for future studies, apart from the shell thickness, which could provide a broader perspective on the stability of bivalve shells. Different microstructural patterns observed in the species could be phylogenetically driven based on their life modes.
Introduction
The morphological attributes of a bivalve (mollusc) shell play a crucial role in the reinforcement of an organism against predation and environmental challenges (Kocabaş et al., 2023). A molluscan shell consists of calcium carbonate (approximately 95 wt.%) and organic materials (5 wt.%) (Currey, 1977; Okumura & De Gennes, 2001); calcite and aragonite are the two main polymorphs of calcium carbonate (Yang et al., 2011). The process of natural selection optimises the shell structure against several biotic and abiotic factors, providing protection, locomotion, stability and buoyancy, which are appropriate for meeting the challenges of survival (Peter et al., 2023).
The correlation between shell microstructure and functional significance in the survival of a bivalved mollusc dates back to Wainwright (1969). Studies concerning the behaviour of the shell in various bivalve species have been reported in the works of Taylor and Layman (1972), Barthelat et al. (2009), Mondal et al. (2010), Chattopadhyay et al. (2013) and others. The authors (e.g., Currey, 1977; Taylor & Layman 1972) analysed the structure of the shell, thereby providing the first insight into the toughness and strength of mollusc shells. Compression and hardness tests were conducted to understand the mechanical properties of the bivalve shell in Saxidomus purpuratus (Yang et al., 2011). The risk of predation and the effect of hydrodynamic conditions are evident and are represented by the morphological variation of the shell (Bertness & Cunningham, 1981; Denny et al., 1985; Menge, 1978). The current knowledge on the behaviour of bivalve shells in an Indian context is very scarce, so the main objective of this study was to understand the shell properties along the east coast of India, which has a very rich molluscan diversity.
In this study, to validate the implications of the mechanical characteristics of the bivalve shells on their survival strategy, we compared our data with the available information on the prey–predator interaction in the bivalve species, Mactra luzonica (Reeve, 1854) and Donax scortum (Linnaeus, 1758), from the same study area. Carnivore gastropods are the primary predators of bivalves reported from the east coast of India; they tend to make boreholes in the shells of the prey associated with mechanical rasping or chemical secretion (Carriker, 1951, 1959; Fischer, 1922). The east coast of India is dominated by two genera of predatory gastropods, Natica Scopoli (1777) and Polinices Montfort (1810) (Mondal et al., 2010).
The east coast of India is an environmentally dynamic ecosystem with a variation in faunal diversity, substrate type and energy conditions (Chaitanya et al., 2015; Gupta et al., 2020). Locally, abiotic factors such as salinity and temperature in a region could potentially influence the ecological diversity and their interaction (Alexander et al., 2001; Hoffmeister et al., 2001). However, the aforementioned factors do not vary much along the east coast of India, and there is no significant influence on the predation intensity (Mondal et al., 2021). The study on the role of environmental factors influencing the naticid–bivalve interaction is scanty at present (Allmon et al., 1990; Kelley & Hansen, 1993).
The samples collected from Chandipur, which is an intertidal flat with a mesotidal setting, consist of mostly mud-dominated substrate, and Chilika on the outer channel has an intertidal setting with substrates dominated by coarse to fine sand (Mahapatro et al., 2015; Mukherjee et al., 1987). The substrate type, life modes of a bivalve and the predatory preference of gastropods are known to influence the predatory risk in bivalves. Our studied species are based on their life modes: Tegillarca granosa (semi-infaunal), Meretrix meretrix (semi-infaunal), Sunetta meroe (semi-infaunal to infaunal), Mactra turgida (shallow burrower) and D. scortum (infaunal in shallow depth). The foraging preference of a predatory gastropod found along the east coast ranges from shallow burrower to infaunal over epifaunal prey, as documented by the borehole drills on the bivalve shells; small infaunal bivalves that dwell in muddy and sandy substrates face a high risk of predatory drilling (Mondal et al., 2013, 2021).
On a microcosmic level, the arrangement and evolution of microstructures in a shell could also provide another aspect to understanding the strength of the shells in molluscs, pioneered by Wainwright (1969). In this study, we have classified the life mode of five bivalves based on the textural arrangements of microstructures in a shell. The nomenclatures are retained from Boggild (1930).
As shown by our study, the variation in centreline thickness (CT), Vickers hardness (VH), density and the microstructural patterns of the shell in bivalves, T. granosa (Linnaeus, 1758), M. meretrix (Linnaeus, 1758), S. meroe (Linnaeus, 1758), M. turgida (Gmelin, 1791) and D. scortum (Linnaeus, 1758), have been addressed to understand the correlation and its significance in resisting different biotic and abiotic factors. This study provides an initial investigation of the mechanical properties of bivalves from the eastern coast of India, as no such study has been conducted before in India at least, to our current understanding of the subject. A limited number of primary mechanical properties have been chosen for this study using available instruments and laboratory facilities.
Study Area
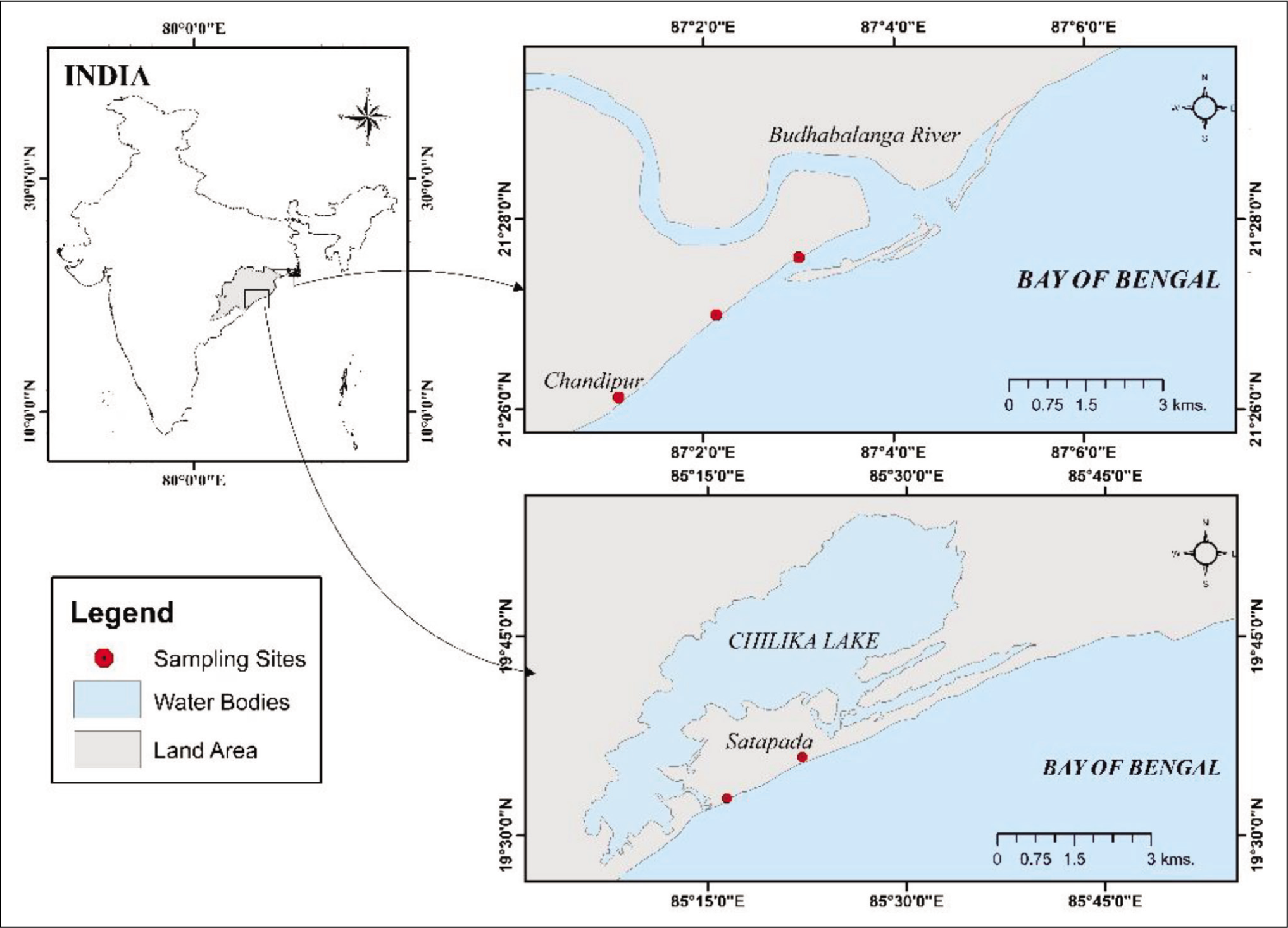
The samples of dead bivalves were collected from Chandipur (21°25′59″N, 87°01′39″E) and Chilika (19°40′46″N, 85°29′60 ″E), which are situated on the east coast of India, adjoining the Bay of Bengal. Chandipur is a coastal region approximately 5 km long with a slender beach of approximately 40 m that features a wide tidal flat of 4 km and comprises a network of bars and inter-bars (Mondal et al., 2010). It hosts a large diversity of molluscs, with bivalves and gastropods comprising 59 and 47 species, respectively (Rao et al., 1991; Sarkar et al., 2016).
Chilika Lake is a 64 km long and 18 km wide water body, covering an area of approximately 900–1,100 sq. km during the summer and monsoon seasons (Mahapatro et al., 2018; Panda et al., 2008). A study on the macrofauna of Chilika Lake by Mahapatro et al. (2018), from 2007 to 2010, reported 61 species of bivalves, 64 species of gastropods and 1 species of Polyplacophora. Chilika Lake is the largest brackish water lagoon in Asia, and since 1981, it has been recognised as a major Ramsar site. The sampling sites are shown in Figure 1.
Map of Sampling Sites on the East Coast of India (Chandipur and Chilika).
Materials and Methods
The five most dominant bivalve species were selected for the present study. T. granosa, M. meretrix, S. meroe, M. turgida and D. scortum are shown in Figure 2. Only the pristine and mature shells of the bivalve specimens were selected for the experimental tests. The estimation of three major mechanical properties—CT, VH and density variations along the centreline for the collected shells—was done, and tests related to the properties were carried out. A preliminary investigation of the microstructural arrangements of the shell textures has also been considered in this study.
Bivalve Species from the East Coast of India; 1. Tegillarca granosa (Linnaeus) (Scale Bar 5 cm); 2. Meretrix meretrix (Linnaeus) (Scale Bar 4.5 cm); 3. Sunetta meroe (Linnaeus) (Scale Bar 4.5 cm); 4. Mactra turgida (Gmelin) (Scale Bar 7 cm); 5. Donax scortum (Linnaeus) (Scale Bar 6.5 cm); 6. A Bivalve Shell Showing Defining Areas (Scale Bar 5 cm).

Centreline Thickness
The thickness variation along the centreline was measured normal to the growth line. Thickness measurement was performed at equidistant points from the margin, the commissure line. The centreline was drawn by joining the midpoint of the commissure line with the hinge point at the umbo, and the specimens were then cut with a low-speed, water-cooled diamond saw.
The locational distance of a point and the measurement were normalised by the actual length of the centreline, l/L, where l is the distance of the measuring point from the commissure line and L is the actual measured length. A minimum of six points were measured to check the variations in the shell. It was carefully measured using a ‘Mitutoyo’ digital Vernier calliper with an accuracy of ±0.0001 mm. The measured values were averaged to obtain the overall estimated mean, used in this study to compare the variation.
Vickers Hardness
At first, the samples were made serially, one by one of 1 cm2, following the centreline of the shell and maintaining an equal distance. The surfaces of all the cut-out samples were diamond-polished to obtain a clear view using a VH metre. Then, micro-indentation testing was done perpendicular to the surface at the centre of 1 cm2 cuts (completely dry) at the same locational points of thickness measurements. It was measured under 100 gram-force (gf) and 50 gram-force (gf) loads.
Density
The same technique of sample preparation, keeping the dimension of 1 cm2, was followed to determine the density variation as it was done for VH. Each sample was weighed first in a weighing machine with a minimum count of 0.1 mg. After obtaining the mass of each sample, the volume was determined. The volume was estimated by dipping it in a 10 ml measuring cylinder and noticing the rise in water level each time the samples were placed inside the cylinder. This procedure was followed for all weighted samples. Then, the density was calculated by simply dividing the mass of each sample by its volume based on density = mass/volume.
Microstructural Textures
The shells of the studied species were cut perpendicular to the growth lines, and the edges were smoothened. The textural arrangements on the shell surface and their microstructures were observed and documented using scanning electron microscopy (SEM).
Results and Discussion
The plot of the CT is represented in Figure 3. For better correlation, the centreline measurement points were made dimensionless by reciprocating the distance of measurement from the commissure line to the entire length of the centreline.
Centreline Thickness (CT) Graph of the Species.
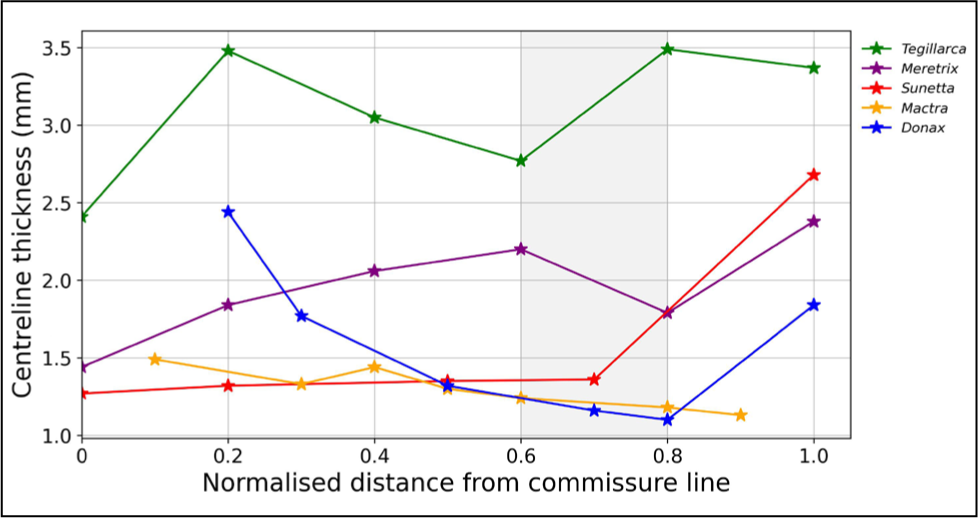
As shown in Figure 3, the overall lowest thickness for all the studied species is located at a normalised distance of 0.6–0.8. The tip of the commissural line shows a low thickness, except for the species D. scortum. The reason could be the surface ornamentation of the D. scortum, consisting of blades and spines on the anterior and posterior regions, respectively. The umbonal region nearest to the beak (oldest) shows the species’ highest CT value.
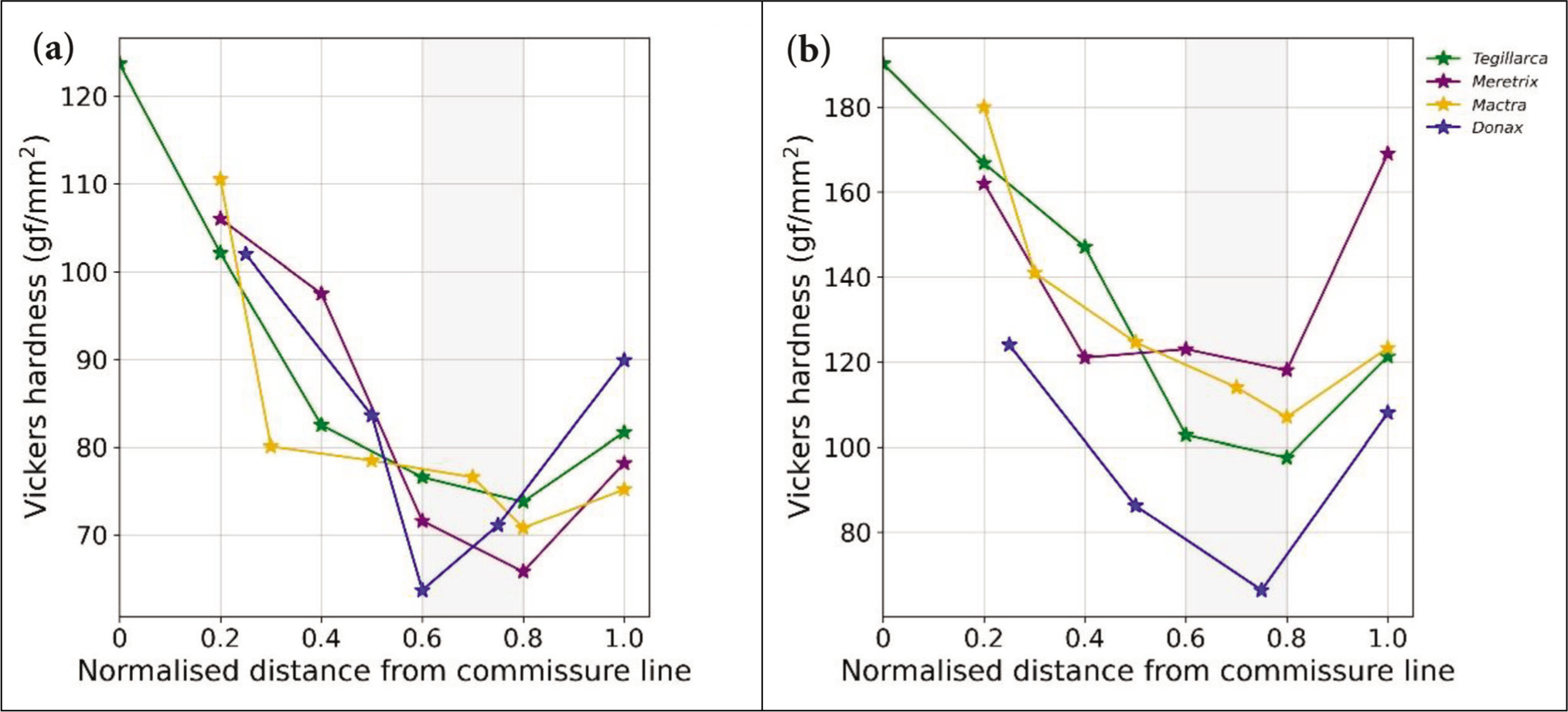
The plot in Figure 4 shows the VH variation from the commissure line towards the umbo. For both 100 and 50 gf, the overall lowest hardness is noticed at a normalised distance between 0.6 and 0.8. It correlates well with the overall lowest CT. The maximum hardness is encountered in the peripheral zone nearest to the commissure line. Both the plots under the applied force of 100 and 50 gf show a very similar trend. An increasing trend is observed towards the umbonal region. The indentation process for the species S. meroe could not be conducted due to its very low shell thickness. The shell could not sustain the test, so no reliable data could be obtained for this particular species.
Vickers Hardness Graph (VH) of the Species Under (a) 100 gf and (b) 50 gf.
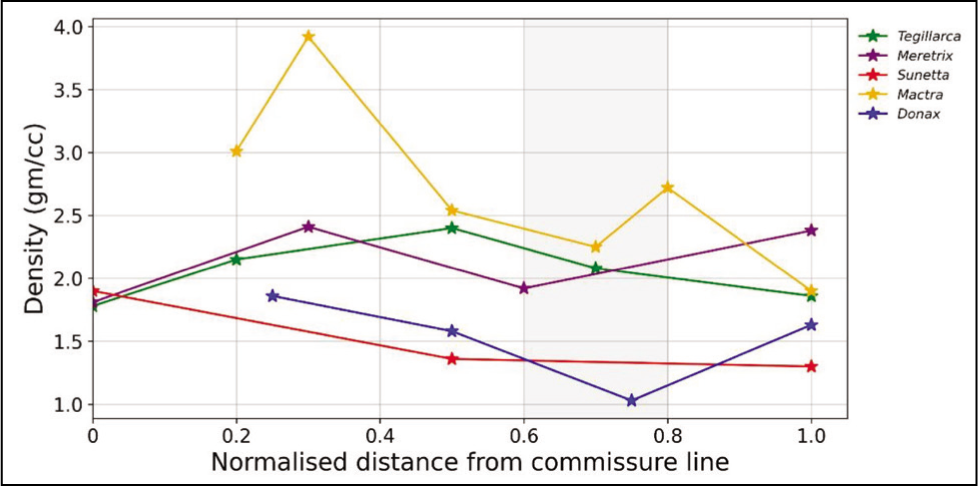
The highest shell density is observed for M. turgida (Figure 5). As illustrated in Figure 5, M. meretrix and D. scortum show an increasing trend in the umbonal region, unlike the other studied species. The comparative study shows that the normalised distance of 0.6–0.8 is the overall lowest density considering all the studied species. It is on par with the CT and VH, with the overall lowest value. The plot in Figure 6 shows the mean values of the mechanical properties of the species considered in this study.
Density Graph of the Species.
Graphs of the Mean Values of the Mechanical Properties.
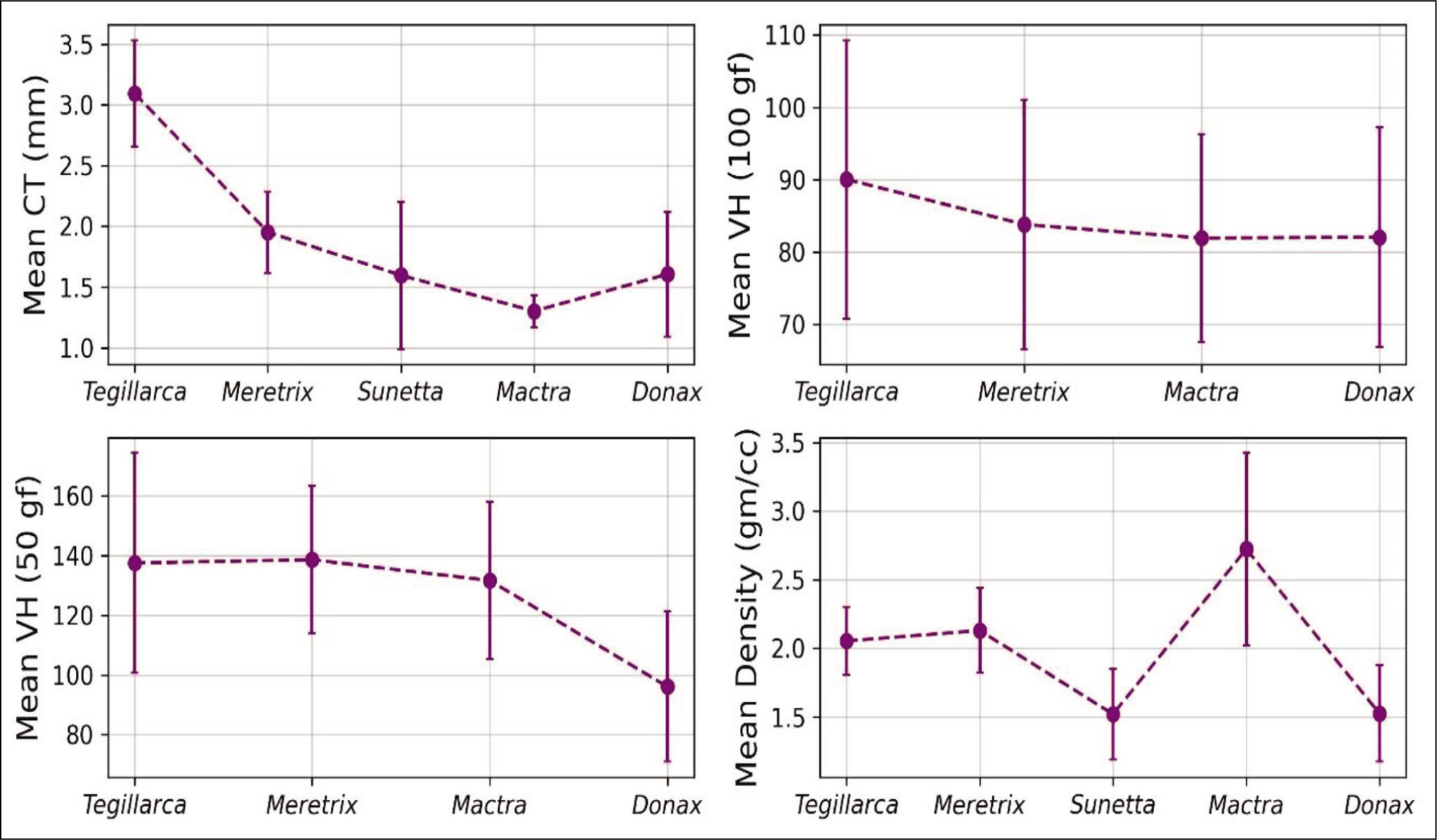
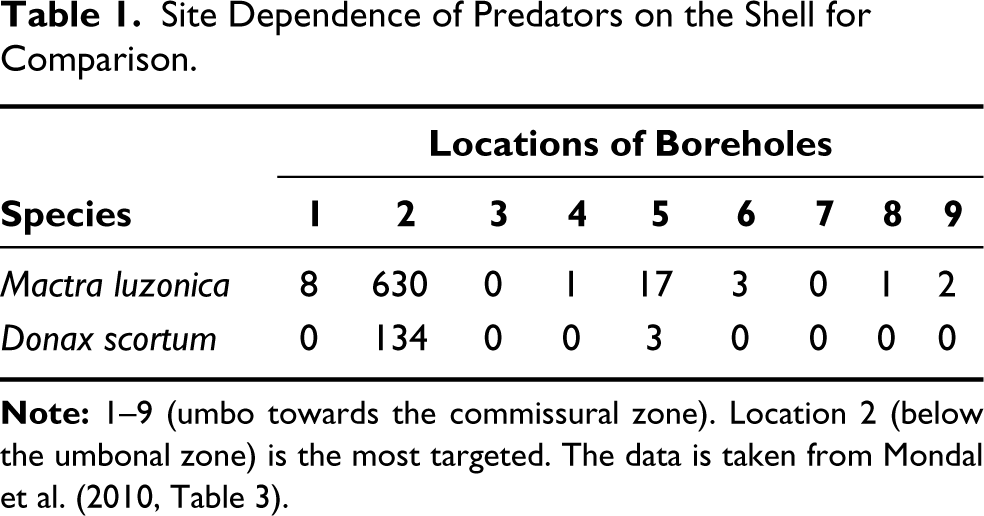
According to site selectivity, predators tend to prefer the thinnest or weakest area in a prey’s shell (Carriker, 1961; Kitchell, 1986; Paine, 1962). The observation of Mondal et al. (2010) states that the most frequent boreholes were noticed below the umbonal region (Table 1), with almost 95% of the cases in M. luzonica and 98% in D. scortum, where the shell area is the thinnest and less ornamented. Our study suggests that the weakest zone in shells for all the five studied species falls at a normalised distance of 0.6–0.8, below the umbonal region. From the correlation between the observation of Mondal et al. (2010) and our data, for the two similar genera (Mactra and Donax), we can confirm that the site below the umbonal zone is the most vulnerable and preferred zone in a shell by the predators. Furthermore, the work of Chattopadhyay et al. (2013) supports this idea, with the observed boreholes in Donax being umbonally located. The mortality rate noticed in M. luzonica was high compared to D. scortum based on drilling frequency, about 24.2% and 8.79%, respectively (Mondal et al., 2010). If the predation preference is primarily based on the shell thickness, the high mortality rate in Mactra compared to Donax stands true based on the mean CT from our data, that is, Mactra (1.301 ± 0.131 mm) and Donax (1.605 ± 0.513 mm); see Figure 6. Based on our data, the same interpretation could be true for the other remaining species, Tegillarca, Meretrix and Sunetta, because the most vulnerable zone in their shell is also found to be below the umbonal region, but unlike Mactra and Donax, it could not be verified due to the lack of availability of data.
Site Dependence of Predators on the Shell for Comparison.
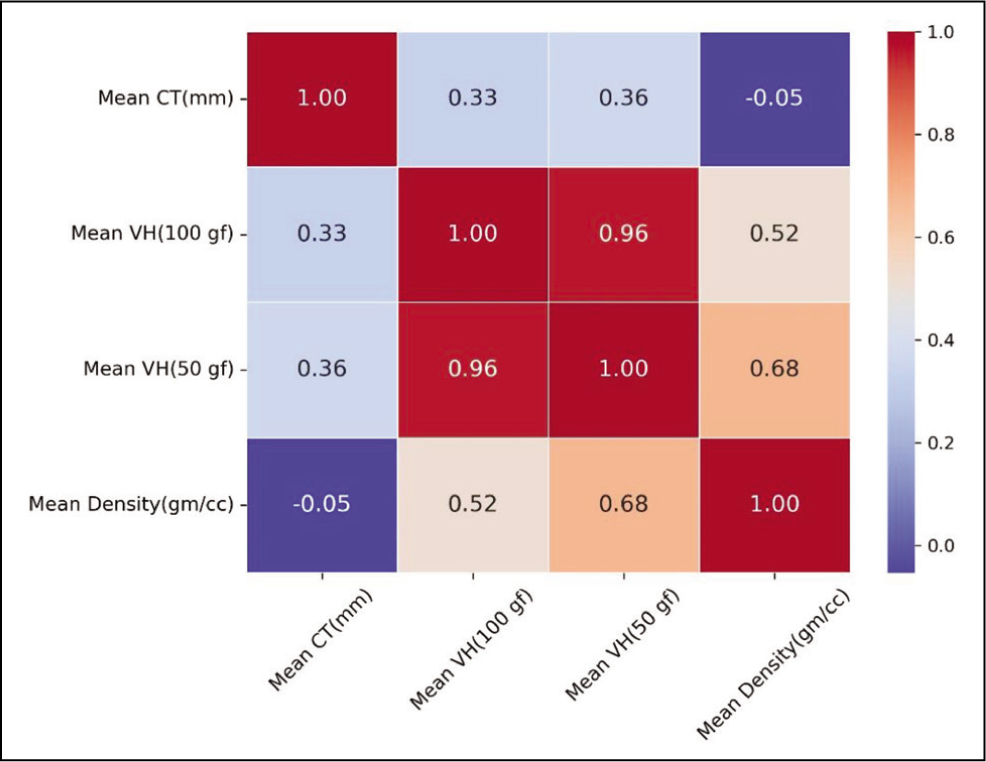
Based on the mean values of the observed mechanical properties, we also calculated the correlation matrix, as shown in Figure 7. There is a high correlation between VH (100 and 50 gf) and density, with a correlation coefficient of 0.52 and 0.68. However, no significant relationship was found between CT and other variables.
Correlation Matrix of the Mechanical Properties.
We also identified five different textures of microstructures from the study, Tegillarca (crossed lamellar), Meretrix (bilamellar), Sunetta (homogeneous), Mactra (complex crossed lamellar) and Donax (foliated), which are illustrated in Figure 8. Based on the mean values of the mechanical properties (Figure 6), there is an inverse relationship between CT and density. We can infer that the species with lower mean shell thickness are compensated by the development of complex microstructural patterns and vice versa. There are no significant trends observed in mean VH for the studied species at both 100 and 50 (gf) loads. The indentation method used in this study is based only on the outer layer of the shell, but a more sophisticated study, such as hardness related to the middle and inner layers of the shell, could potentially help to decipher the functional significance of the different microstructural patterns. The development of microstructural changes in the shell could be phylogenetically related to different species based on the life mode of a bivalve.
Microphotographs (SEM) of the Microstructural Patterns; 1. Tegillarca granosa (Crossed Lamellar); 2. Meretrix meretrix (Bilamellar); 3. Sunetta meroe (Homogeneous); 4. Mactra turgida (Complex Crossed Lamellar); 5. Donax scortum (Foliated); Scale Bar 50 µm.

Other biotic factors not considered in this study that could influence the predation risk of a bivalve are the shell morphology (shape) and shell ornamentation, for example, the development of blades and spines during the late ontogeny in Donax, which acts as a mechanism for protection and makes it a less favourable prey compared to Mactra, which has a smooth shell ornamentation (Mondal et al., 2010, 2021).
The study areas of Chandipur and Chilika host a rich molluscan and other biological communities with a documented list of species, as mentioned earlier. Based on the predominance of the bivalves, the ecological condition of the area and the accessibility of our facility, the chosen sites are most appropriate for this investigation.
During the high tide conditions along the east coast, the risk of predation in bivalves increases due to the presence of secondary predators such as portunid crabs (Mondal et al., 2010). Based on the geographical location, the energy condition is higher in Chandipur compared to Chilika, with a noticeably higher rate of predation, documented in bivalves from Chandipur. In our case, we cannot verify this statement because of the different species, chosen from the two sites, but the dominance of the predatory species could be a major factor. The tendency of predatory gastropods, considered the main predator, with their foraging preference for shallow and semi-infaunal species, makes bivalves such as Mactra more vulnerable compared to infaunal species such as Donax. Based on the life mode, Tegillarca, Meretrix and Sunetta also fall under the range of predatory preference zones, but the statement cannot be justified unless correlated with the predatory evidence. An interplay of biotic and abiotic factors could potentially influence the mechanical properties of a shell in different ways in various bivalve species.
Conclusion
Our observation of the most vulnerable region in a bivalve shell, documented below the umbonal zone, is supported by the evidence of predatory markers (boreholes) from the works of Mondal et al. (2010) and Chattopadhyay et al. (2013). The shell thickness does play a significant role, but the associated contributions of hardness and density of the shell, as shown by our study, are the other essential properties to be considered for understanding the survival strategy of bivalves. There are limitations in understanding the complex control of biotic and abiotic factors on the mechanical properties of the shell, but each effort carries the potential to pave the way for a detailed study in the future. The development and evolution of microstructural patterns in a shell of various life modes in a bivalve can provide further knowledge on the stability of the shell. A detailed study of the spatial arrangement of the crystals in the structure is required. The answer to these observations may lie within the internal structure of the shell and its arrangement (stacking patterns of the crystals) within the molluscs, which is the future scope of this study.
Footnotes
Acknowledgements
We thank Mr Taraknath Pradhan for the sample preparation and Mr Anindya Pal (M. N. Dastur School of Material Sciences and Engineering, IIEST, Shibpur, Howrah) for his help during the testing of the mechanical properties. RS also thanks Dr Abiral Tamang (Department of Physics, Jadavpur University) for the SEM images and Hari Ramudamu for the help related to the study.
Declaration of Conflicting Interests
The authors declared no potential conflicts of interest with respect to the research, authorship and/or publication of this article.
Funding
The authors received no financial support for the research, authorship and/or publication of this article.