
Abstract
Establishing modern pollen analogues under various vegetation types in a location is necessary without which it is difficult to accurately interpret fossil pollen assemblages in any region in terms of the past environment and palaeoclimate. We have analysed the modern pollen dataset of 30 surface soil and sediment samples from the forested, cropland, wetland and river sites of the Karimganj District in south Assam, to comprehend the connection between the recent pollen assemblage and the vegetation patterns in the Barak Valley. The overall pollen data reflect the tropical moist deciduous and semi-evergreen forest comprising Syzygium, Schleichera, Terminalia, Lagerstroemia, Duabanga, Sapotaceae, Ilex, Mesua and Schima under a warm and humid climate in response to rainfall variations in the region. The presence of Euphorbiaceae and Convolvulaceae inferred high monsoonal activity in and around the region. A robust relationship between local herbaceous vegetation and pollen was observed. The steady occurrence of cereal pollen (average 18.67%) along with other cultural pollen taxa like Brassicaceae, Coriandrum and Solanaceae illustrate intense agricultural activity around the valley areas. Multivariate principal component analysis (PCA) and box plot were applied to the quantified data obtained from pollen frequency analyses of the surface samples which clearly revealed a significant variation and similarity in vegetation types. Thus, an attempt has been commenced to precisely observe the behavioural pattern of modern pollen deposition in a varied depositional setting, which could, in turn, help in assessing the expanse of agricultural practices and the depth of deterioration of pristine forests and surrounding areas. This modern training dataset could also help in the precise reconstruction of past climate and vegetation shifts in the Barak Valley region. A correlation of regional palynodata with other regions of the Indian subcontinent and Indo-Burma will also be assessed for better interpretation.
Introduction
The state of Assam situated in northeast India at the foothills of the eastern Himalayas is one of the most climate-sensitive regions owing to its unique topographic features and ecosystem (Dixit & Bera, 2013). The Barak valley in Assam is a part of the Indo–Burma biodiversity hotspot and is mainly characterised by tropical moist deciduous forest along with the scattered growth of semi-evergreen tree taxa under subtropical, warm and humid climate with an average rainfall of greater than 300 cm, most of which is received during the southwest monsoon season (June to September) (Tripathi et al., 2021). Fossil pollen plays a crucial role in deciphering palaeovegetation and past climate since it is a direct proxy record of past vegetation and an indirect proxy for past climate. Pollen, vegetation and climate interactions, however, are not linear, inconsistent and difficult to understand (Dixit & Bera, 2011). The lack of thorough and systematic investigations on contemporary pollen dispersal and deposition processes, particularly on the quantification of these processes, has been one of the major contributors to disparities between pollen data (Bonnefille et al., 1999).
Therefore, a deep knowledge of the linkages between current vegetation and modern pollen rain is required for advancing the interpretation of fossil data and reconstructing past vegetation and climate with certainty. Nevertheless, qualitative studies of the pollen-vegetation relationship in northeast India have been carried out, especially from the lower Brahmaputra plain of Assam and Meghalaya (Basumatary et al., 2013; Dixit & Bera, 2011, 2012; Tripathi & Bera, 2014; Tripathi et al., 2020). Moreover, the Late Holocene vegetation and climatic history of the Barak Valley region were interpreted based on pollen and diatom studies (Tripathi et al., 2021). However, despite the rich biodiversity, the existence of the endangered and endemic plant species and good pollen preservation, no modern pollen reference data have been documented to date from the Barak Valley region of South Assam.
Thus, the objective of the present study is (1) to provide background information in terms of pollen morphological data of important plant taxa in and around the Karimganj District of Barak Valley, Assam and, (2) to analyse the relationships between modern pollen and vegetation from the 30 surface soil and sediment samples procured from the different depositional setting like forest, open land, cropland, wetland and river margin of the Karimganj District. The precise reconstruction of past vegetation and palaeoclimate from the Barak Valley region is important and is regarded as a poorly understood area of the tropics (Dixit & Bera, 2013). Regional pollen data will be correlated/compared with those in the other regions of the Indian sub-continent as well as those in the Indo–Burma region for better interpretation.
The recent creation of a new method for reconstructing global biome distribution utilised for data model comparison in the framework of global change projects has highlighted the need for full global coverage of modern samples (Prentice et al., 1996). It has taken a lot of actual pollen counts from all the habitats observed at all latitudes to establish this methodology. There are already such data sets for Europe (Guiot et al., 1996; Peyron et al., 1998; Tarasov et al., 1998), North America (Williams et al., 1998) and Africa (Julier et al., 2021). For South-east Asia and India, however, where information on pollen rain from contemporary vegetation is still reported incongruously, they have not yet been compiled. This study aims to provide the whole data set for reconstructing realistic biomes in the Barak Valley using actual pollen data from South Assam, India.
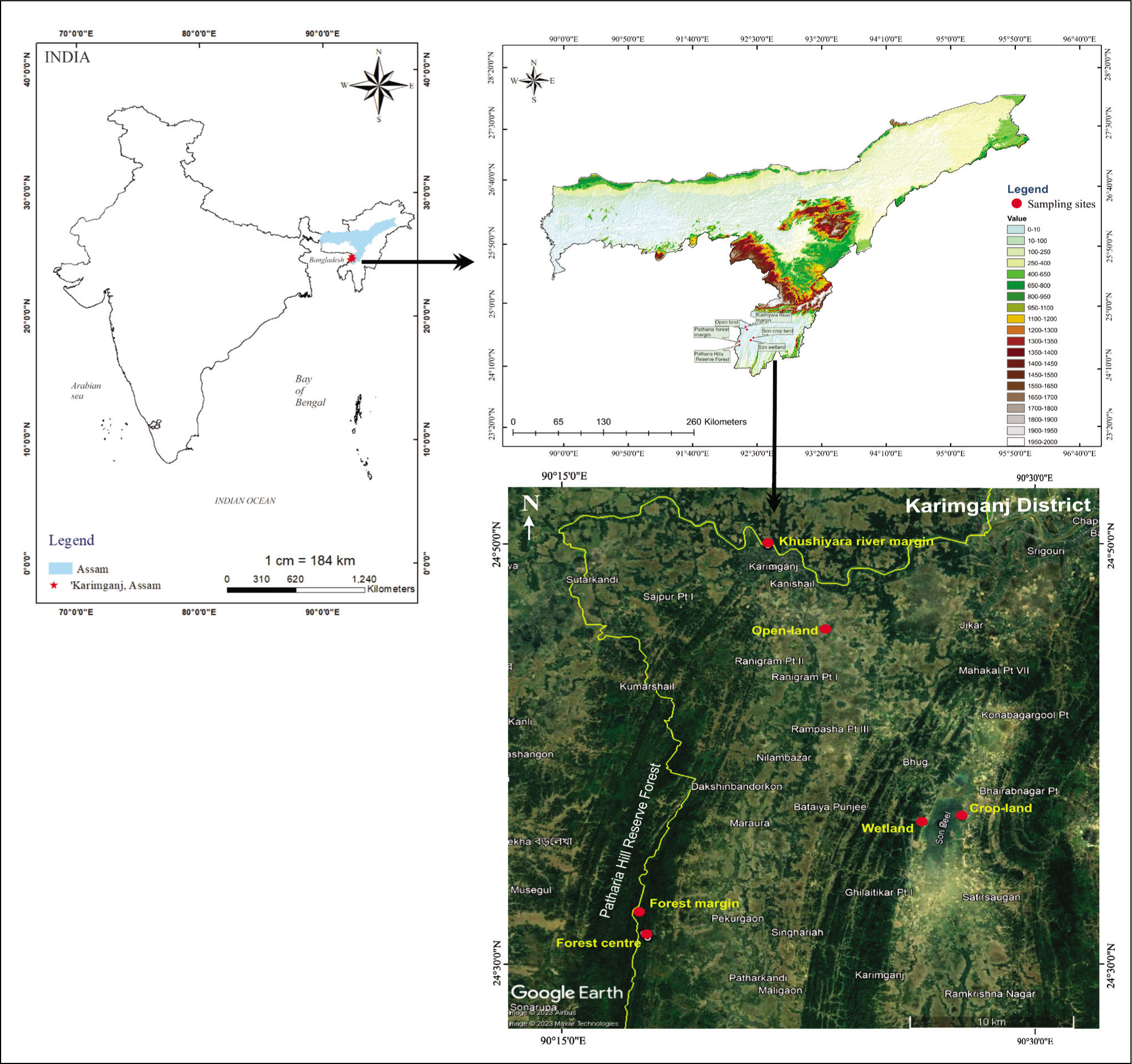
Study Area
The Barak Valley covers an area of about 39,390 sq. km. and encloses the three districts of south Assam (Cachar, Hailakandi and Karimganj) in India (30,770 sq. km), extends into Bangladesh (7,780 sq. km.) and a small part of Myanmar (840 sq. km) (Figure 1). The Barak Valley’s tectonic history and structural configuration has had a significant impact on the evolution of its terrain during the Pleistocene epoch (Tripathi et al., 2021). It encompasses a wide range of alluvial landforms, such as flowing river channels, dried-up river channels, meander cut-offs, bhur ridges and water bodies (lakes and ponds). The Karimganj district is roughly parallel to Alaska’s Afognak Island, covering 1,809 square kilometres (698 square miles). On the northeast, it shares a boundary with Cachar District, on the east and south, with Hailakandi District, on the south, with Mizoram, on the southwest, with Tripura State, and on the west, with Bangladesh (Figure 1). The study sites are located along the Kushiyara River on the northern edge of the district bordering Bangladesh. The forests of Karimganj once boasted an abundance of vegetation and fauna, but they are currently being deteriorated due to human onslaught.
Map showing the sampling sites in the Karimganj District of Barak Valley, Assam (using Arc GIS 10.5 and Google Earth).
Soil and Climate
In the Barak Valley of Assam, the soil is mostly distinguished by younger and older alluvium depositions with colour variations. Often, clay, silt and sand make up this material. It has a very slightly acidic nature. It is significantly less acidic and occasionally marginally alkaline along the riverbanks (Karunakaran, 1974). Typically, the monsoonal breeze starts in June, and severe rains last until October. The primary cause of rain in this area is the South-West monsoon. In Karimganj and the surrounding areas, the average maximum temperature in August is 32°C, and the average minimum temperature in January is 10°C (Mani, 1974;
Vegetation
Assam has a rich vegetation cover due to its varied topography, climate and soil. The vegetation of the state can broadly be classified into (i) tropical evergreen and semi-evergreen forest, (ii) tropical moist and dry deciduous forest, (iii) tropical grassland, (iv) Swamp forest, (v) subtropical mixed forest and (vi) mixed bamboo forest (Tripathi et al., 2021). The Barak Valley is mainly characterised by tropical moist deciduous forests along with intermittent semi-evergreen tree taxa (Figure 2(a) and (b)).
Field photographs in Karimganj District, Assam; a. Tropical moist deciduous forest inside Patharia Hill Reserve forest; b. Tree fern, Cyathea giantea growing inside Patharia Hill Reserve forest; c. Son wetland (dried area); d. A Shrub, Melastoma malabathricum growing luxuriantly at the forest margin; e. Collection of surface soil from the Kushiara River margin, Karimganj District, Indo-Bangladesh border; f. Kushiara River flowing through Indo-Bangladesh border.

Material and Methods
The sampling locations were specifically chosen to encompass various land use zones in order to identify the association between the recent (modern) pollen assemblage and contemporary vegetation patterns in the Barak Valley region of Assam (Figure 2). In the Karimganj District of South Assam, sampling stations were separated into six zones: the forest centre, forest edge, open land, cropland, wetland and river margin (Figure 1). At each sampling site, a topsoil sample was procured at 0–2 cm using a stainless steel trowel, packed tightly in a ziplock plastic bag and subsequently labelled with all the details of the field site including the type of soil, climate, vegetation and date of collection. Below is a description of the procedure used to chemically prepare the living and surface samples for palynological observation using Erdtman’s method (Erdtman, 1943).
Extraction of the Pollen/Spores from the Living Polliniferous Samples for the Pollen Morphological Studies
Collection of flowering twigs for polliniferous materials was made from various localities in and around the Patharia Hill reserve forest of Karimganj District, South Assam. Phenological data like the flowering period has been recorded by serial field observations. The identification of plant specimens was done using relevant floras and standard literature (Kanjilal et al., 1982). The acetolysis process for the extraction of pollen/spores from the flowers was carried out using Erdtman (1943) and provided below:
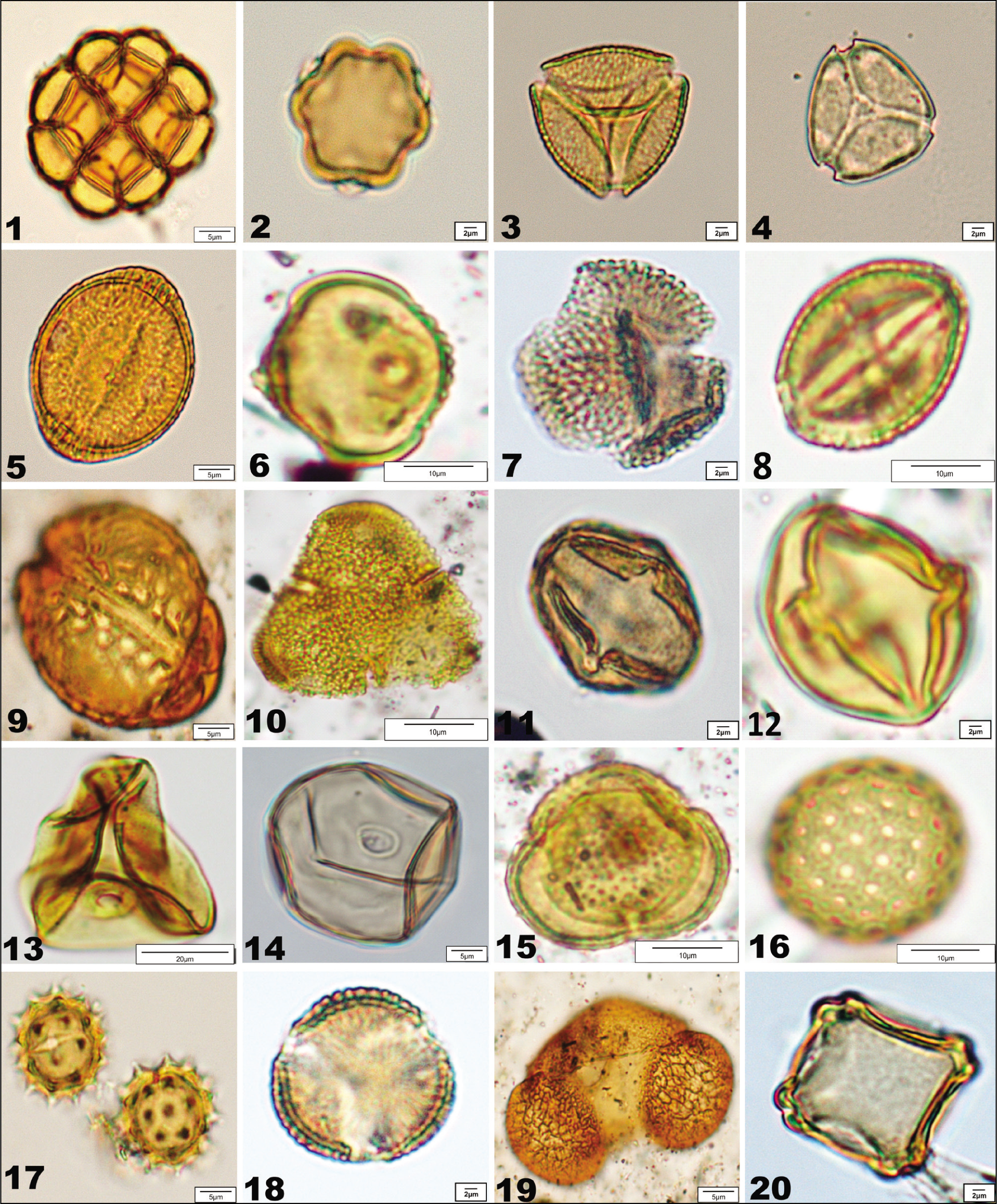
Remove all the petals and sepals from the flower. After that, keep the anthers in a fresh vial tube and gently tease the samples with the help of a glass rod by adding some amount of water. Consequently, the material was then sieved through a 150 µm-mesh sieve to remove the residue materials. After the sieving process, the left filtrate was transferred to the clean 15 ml centrifuge tube. Thereafter, the sample was treated with glacial acetic acid (GAA) and centrifuged in a dehydration process, followed by the boiling in a water bath for 5 minutes after treatment with an acetolysing mixture (9:1 ratio of acetic anhydride and concentrated sulphuric acid). This method is performed for the removal of cellulose and the clear visibility of pollen micro-morphology. The sample was then centrifuged and decanted off to remove the acid. Again, the material was treated with GAA and washed 2 to 3 times with distilled water. Thenceforth, 50% glycerine was added to the residue and a few drops of phenol were also added to protect the samples from microbial decomposition. Permanent pollen slides were made with the help of DPX and polyvinyl for the preservation, identification, photography and studies of pollen morphology under a light microscope (LM) (Plate 1).
Pollen recovered from the flowers and surface soil samples of Karimganj District, Barak Valley of Assam; 1. Acacia; 2. Terminalia bellirica; 3. Schleichera; 4. Syzygium; 5. Lagerstroemia parviflora; 6. Duabanga sonnertioides; 7. Ilex excelsa; 8. Rutaceae; 9. Barringtonia acutangula; 10. Bombax ceiba; 11. Euphorbiaceae; 12. Solanaceae; 13. Cereal; 14. Non-cereal; 15. Xanthium; 16. Amaranthaceae; 17. Asteraceae, 18. Brassicaceae; 19. Pinus; 20. Alnus.
Extraction of the Pollen/Spores from the Surface Soil/Sediment Samples for the Modern Pollen-Vegetation Relationship
Before the examination of the sediment cores from the investigation areas for the palaeoecological assessment, the study of the modern pollen/vegetation relationship is indispensable. The status of pollen preservation in the soil could be well portrayed through the modern palynological analysis. To achieve the target a series of surface soil has to be analysed from the various forest stands, adjoining open-land, cropland and wetland areas, which incorporate swamps and river margins also, to understand the respective representation of the forest constituents in the modern pollen rain as well as their dispersal efficiency and preservation in the sediment concerning their factual composition in the regional forest types. This comparative database generated from pollen rain and the extant vegetation through the analysis of surface soil could be a modern analogue for the appropriate evaluation of the fossil pollen spectra in terms of preservation and reconstruction of past vegetation and climate in and around Barak Valley of Assam.
The materials for the present study include thirty surface samples including 18 surface soil (procured in a transect from the forest to open land and cropland) and 12 sediment samples procured from wetland and river margin areas of the Karimganj District of South Assam and the chemical procedure adopted was stepwise provided below:
In order to dissolve the humic acid, about 20 g of surface soil and sediment sample were boiled in a 10% potassium hydroxide (KOH) solution for 5 minutes to deflocculate pollen from the soil. The samples were then subjected to three washes with distilled water in order to remove the alkalis by the decantation procedure. The materials were sieved using a 150 m-mesh sieve to remove the coarse debris following the KOH treatment. If carbonates were present, they were then removed from the samples using 10% hydrochloric acid (HCL), and the samples were then rinsed twice with distilled water to remove the acid using decantation. To eliminate the silicates, the material was then treated with 40% hydrofluoric acid (HF) and stored in plastic jars for 2 to 5 days. To rid the samples from silica and excess HF, they were again rinsed twice with distilled water. The left filtrate was treated with GAA, dehydrated and centrifuged twice. Then, it was treated in a water bath for five minutes, or until the water began to boil, with an acetolysing solution (9:1 ratio of acetic anhydride and concentrated sulphuric acid). To remove the acid out, the samples were centrifuged and decanted. The remaining material was treated with GAA once more and cleaned with distilled water three or four times. In order to protect the samples from microbial decomposition, 50% glycerine and a few drops of phenol were afterwards added to the residue. The grass pollen has been separated into the Poaceae <45m (non-cereal) and Poaceae >45m (cereal) size categories (Joly et al., 2007; Tripathi et al., 2019, 2020).
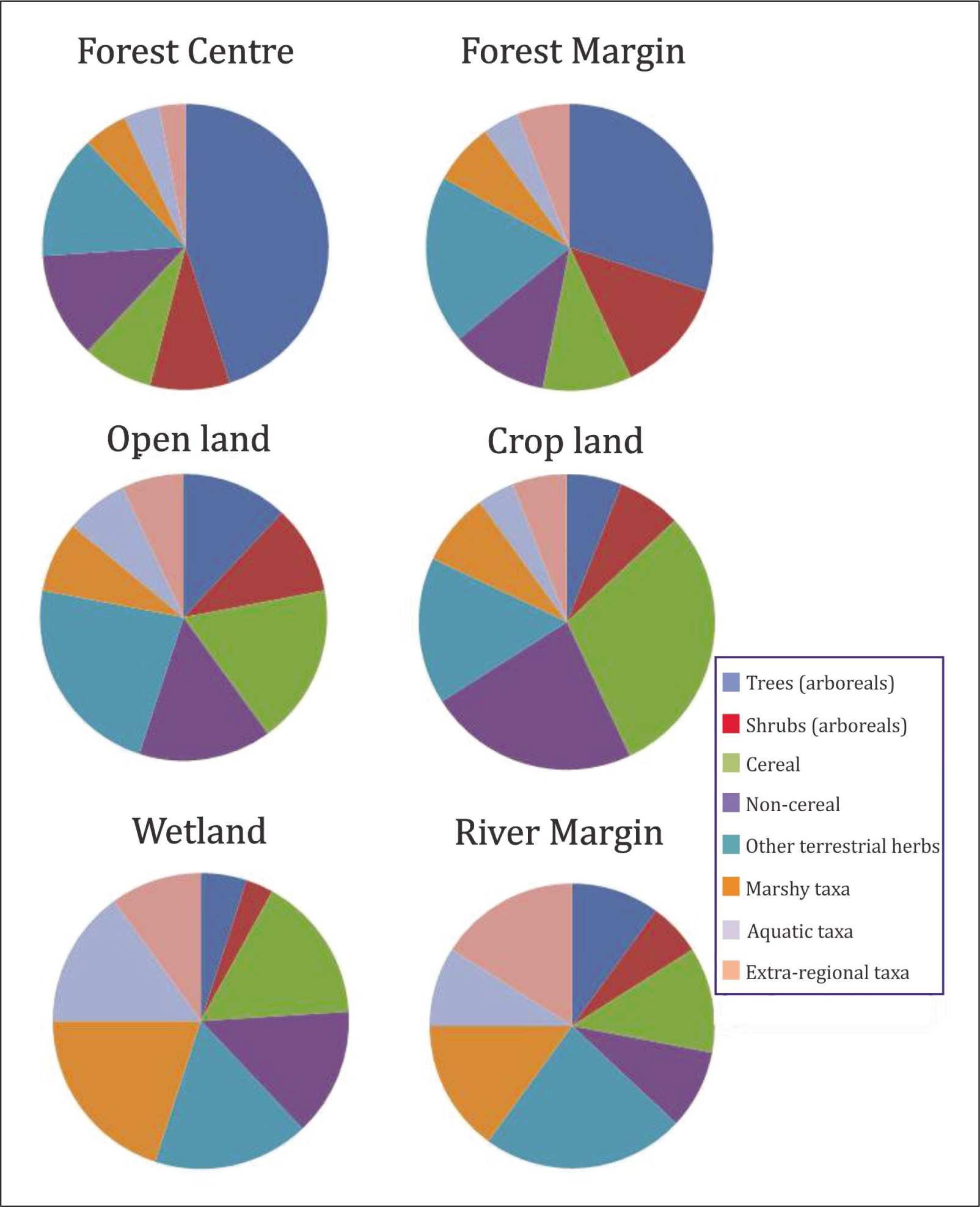
The overall pollen count for each sample ranges from 250 to 450. In the Micro Excel program, a pie chart was used to display the composite pollen spectrum (Figure 3). Arboreals (trees and shrubs), non-arboreals (terrestrial herbs and marshy/aquatics) and extra-regional taxa are the three categories into which the recovered pollen taxa from the surface samples have been divided (Figure 3). Pinus, Betula, Alnus and Quercus, are extra-regional taxa coming from the Himalayan regions. They attain improved value in open-land samples including wetlands and rivers, which may be attributed due to the changing face of the forest (outskirt open land) where strong wind and water found carrying Himalayan pollen taxa from nearby or distant higher reaches which finally incorporated in the soil.
Composite Pollen spectra from the surface soil of forest centre, margin, open-land, crop-land, wetland and river margin areas of Karimganj District, Assam to understand modern pollen frequency and vegetation association in different depositional sites.
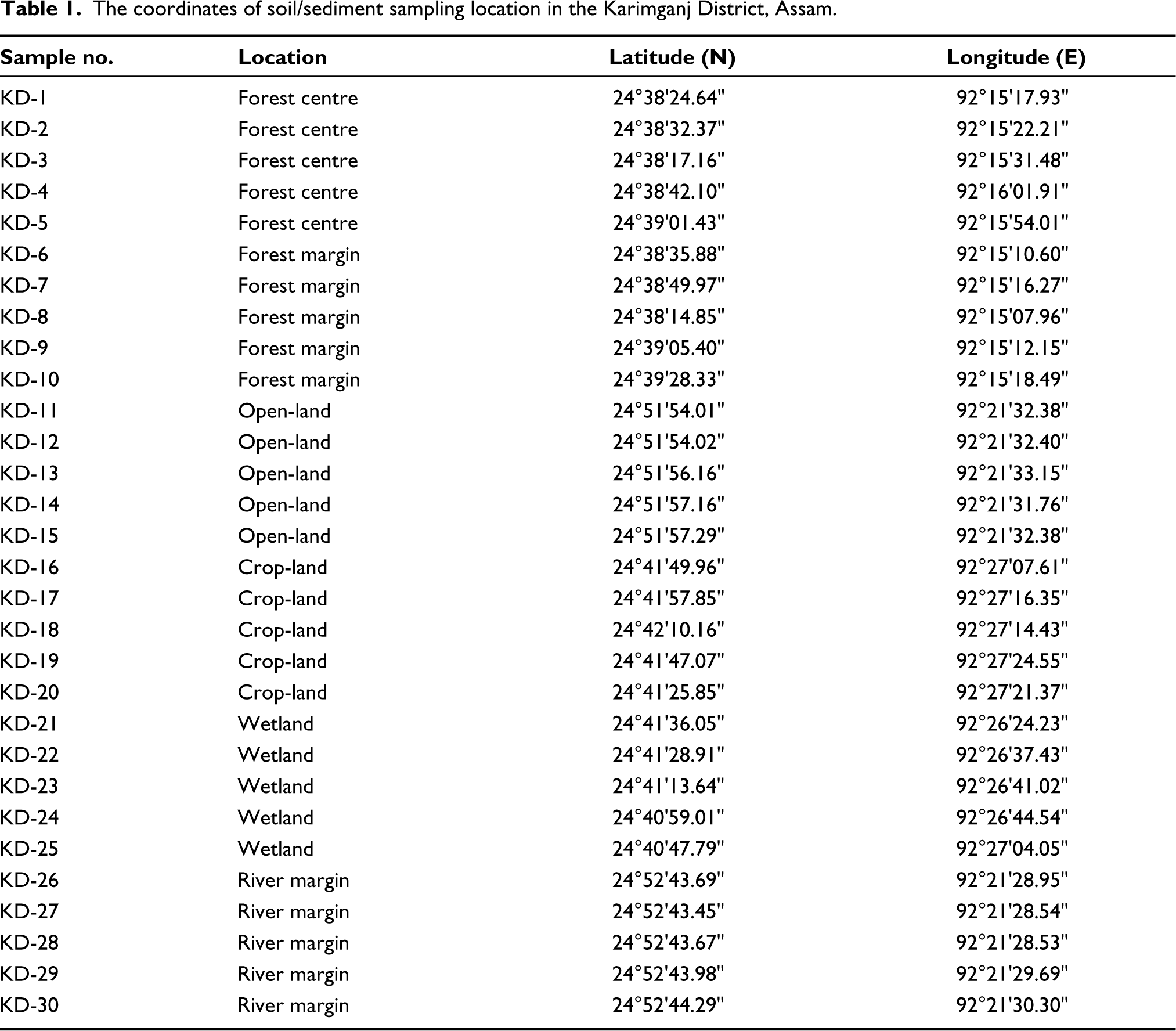
Due to the overgrowth of ferns in the study region, fern spores (both monolete and trilete) are not included in the total pollen count to avoid the bias representation in the palynoassemblage. For the precise identification of pollen and spores, reference pollen slides from the Birbal Sahni Institute of Palaeosciences’ herbarium and published pollen literature (Bera et al., 2009; Chauhan & Bera, 1990) were used. Using an Olympus BX-50 microscope, palynomorphs were photographed and counted (Plate 1). A table has been prepared portraying the coordinates of soil/sediment sampling location (Table 1).
The coordinates of soil/sediment sampling location in the Karimganj District, Assam.
Principal Component Analysis
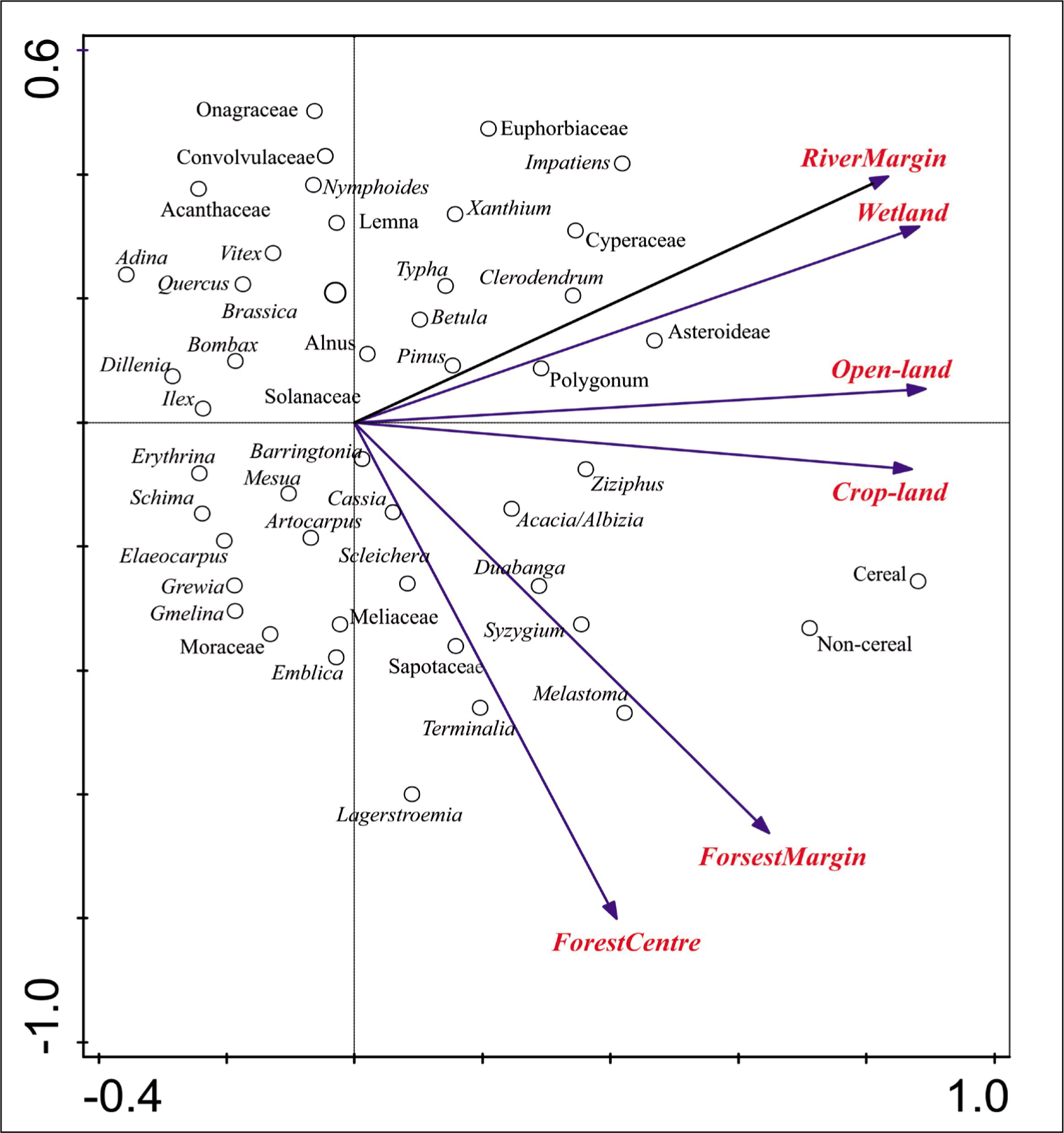
Principal component analysis (PCA) was used with the Canoco version 5 software to analyse the statistical significance of quantified data of 49 pollen taxa and their frequencies obtained from surface soil samples, procured from various depositional sites (forest centre, forest margin, open land, cropland, wetland and river margin) of the Karimganj District of South Assam (Smilauer & Leps, 2014). Moreover, the statistical significance of quantified data of six major vegetation groups like trees, shrubs, terrestrial herbs, aquatic taxa, marshy taxa and extra-regional taxa formed by categorising different pollen taxa was also analysed against the aforesaid surface samples. A p-value of ≤.05 was used to determine the significance of the data. The value was imported into Corel Draw-12 software for minor editing of data labels (Figure 4).
PCA ordination plot showing interrelationships of recovered pollen taxa and surface soil samples collected from different depositional settings of the Karimganj District, Barak Valley of Assam.
Box-Plot Analyses
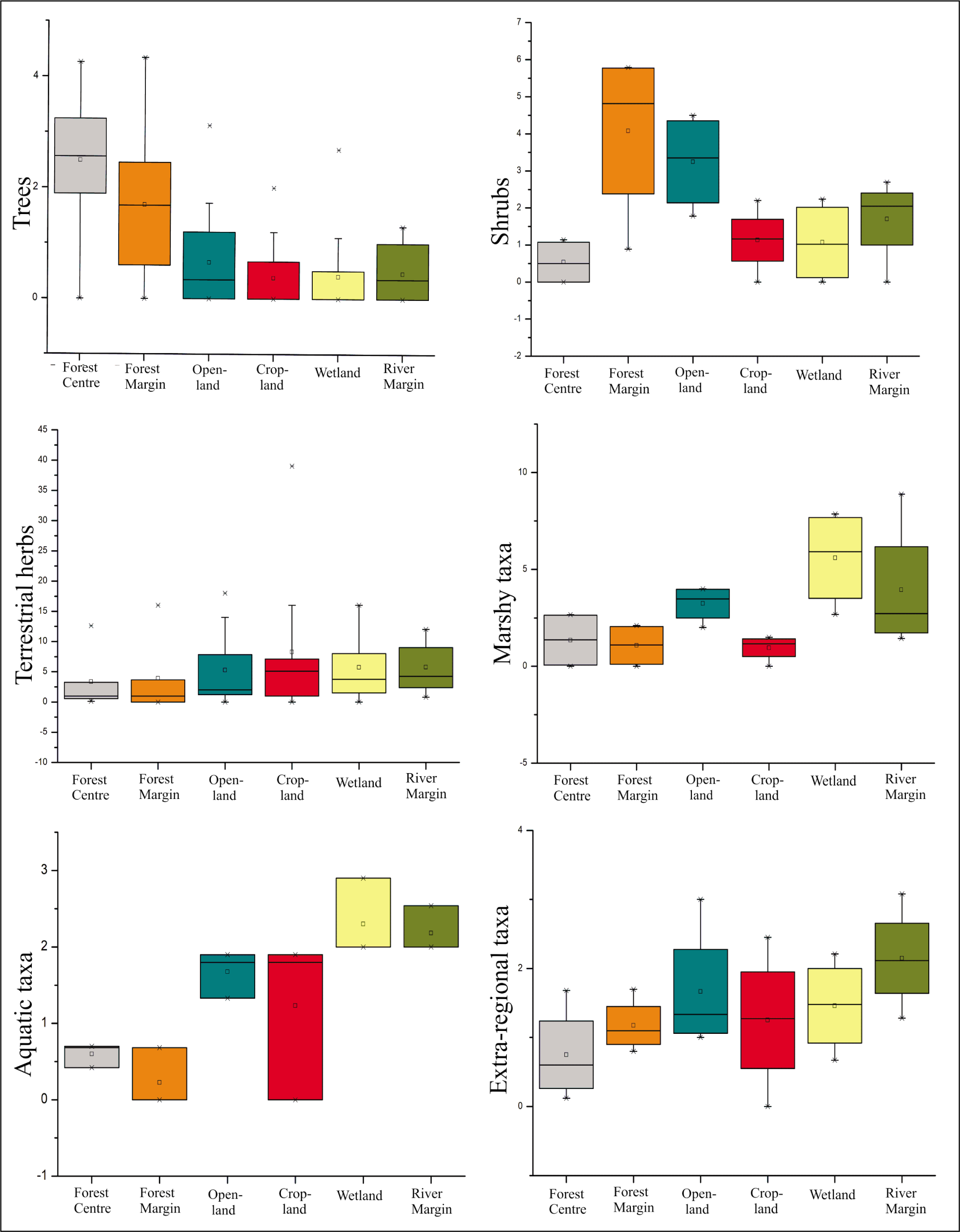
To compare and visualise the main vegetation groups among the various surface pollen substrates of the different depositional sites in the Barak region, box-plot graphs were created using the Origin Pro 8.5 software (Figure 5). The pollen dataset’s presentation was standardised using this statistical technique, which was based on a five-number summary (Tripathi et al., 2023). To determine the minimum to maximum values for six different pollen substrates (forest centre, forest margin, open land, cropland, wetland and river margin), the collected pollen data were divided into the main vegetation groups (arboreals, terrestrial herbs, aquatic taxa, marshy taxa and extra-regional taxa). The box-plot graphs show the variation in abundance and dearth of variable pollen types at respective sites. Some suggestive signs have been mentioned in the graph to show the minimum-maximum tendencies that aid in deciphering the accurate fluctuations among the pollen-vegetation relation. In the box plot, the hollow square (β) represent the ‘mean’, the cross (*) represents the ‘maximum values’ and the ‘minimum values’ and the black line inside the box is the ‘median’ (Figure 5).
Box plots showing distribution of arboreal, non-arboreal and extra-regional taxa in Karimganj District, Barak Valley based on the modern pollen precipitation.
Pollen Morphological Investigations
The pollen morphometrical observation presented here is based upon detailed pollen morphological analysis of 21 plant taxa belonging to 16 families distributed in the Barak Valley of South Assam. The chief objective of preparing this pollen morphological data is to identify modern and fossil pollen grains preserved in the surface soil and sediments, respectively. To avoid repetition, pollen diagnosis of individual taxa is not given here. This section includes the major pollen morpho-characters which have been made sufficiently exhaustive to meet all requirements in identification (Plate 1). In preparing this pollen morphological data, primary importance is given to the number, position and character of apertures and sexine ornamentation. The terminology adopted is after Erdtman (1952); Nair (1966); Faegri et al. (1989) and Punt et al. (2007). This is a regional key applicable to the Barak Valley region under investigation.
Results of Pollen Morphometry of Selected Plant Taxa
Areca catechu Burm. f. Vern. Tamul (Arecaceae) Pollen 1-sulcate, 37 × 34 µm, longest axis 25 µm and 8–10 µm broad. Exine comparatively thin, sexine with rounded warts.
Artocarpus chaplasha Roxb. Vern. Sam (Moraceae) Pollen 3-4-porate, subprolate, 21 × 17 µm. Pores circular, 4 µm, tenuimarginate. Exine 1.5 µm, sexine slightly thicker than nexine, granulate-foveolate, tegillate.
Barringtonia acutangula Gaertn. Vern. Hinsol (Lecythidaceae) (Plate 1, figure 9) Pollen 3-syncolpate, prolate, 42 × 32 µm. Colpi crassimarginate. Exine 3µm thicker at aperture, sexine thicker than nexine, granulate, tegillate.
Bauhinia purpurea L. Vern. Kanchan (Fabaceae) Pollen large 3-colporate, prolate, 70.3 × 50.2 µm. Colpi 60 µm long, broad, crassimarginate. Os distinct, lalongate (12.5 × 6 µm). Exine 3–4 µm thick, sexine thicker than nexine, striatoreticulate sexine pattern.
Cassia fistula L. Vern. Sonaru (Fabaceae) Pollen 3-colporate, prolate-spheroidal, 25.9 × 22.9 µm. Colpi long, broad, tenuimarginate. Os lolongate. Exine 1 µm thick, sexine as thick as nexine, punctate sexine pattern.
Cinnamomum tamala T.Nees & Eberm. Vern. Tejpat (Lauraceae) Pollen non-aperturate, spheroidal, 34 µm. Tenuiexinous, provided with scattered, irregular, short projections or warts up to 1.5 µm in height.
Clerodendrum infortunatum L. Vern. Vatetita (Lamiaceae) Pollen spheroidal, 70 µm. Exine 2.9 µm, scattered spinules, coarsely granulate.
Dillenia indica L. Vern. Owtenga (Dilleniaceae) Pollen 3-colporate, brevicolpate, subspheroidal, 28.5 × 29.5 µm. Colpi broad. Os lalongate. Exine 3 µm thick, sexine slightly thicker than nexine, retipilate, integillate.
Dipterocarpus macrocarpus Vesque. Vern. Hollong (Dipterocarpaceae) Pollen 3-colpate, oblate, 38 × 55 µm. Colpi long, straight. Exines 3 µm, sexine punctitegillate, granulate.
Duabanga sonnertioides Buch.-Ham. Vern. Bandar-kanda (Lythraceae) (Plate 1, figure 6) Pollen 3-colporate, prolate-spheroidal, 42.5 × 37.2 µm. Colpi long, margin rough and thicker. Os slightly lalongate. Exine 2.6 µm, sexine as thick as nexine, finely reticulate, tegillate.
Elaeocarpus rugosus Roxb. Vern. Ruddraksh (Elaeocarpaceae) Pollen 3-colporate, prolate-spheroidal, 12 × 8 µm. Colpi distinct, tenuimarginate. Exine 0.5 µm thick, sexine thicker than nexine, finely granulate.
Emblica officinalis Gaertn. Vern. Golian (Phyllanthaceae) Pollen 4-colporate, oblate-spheroidal, 21 × 21 µm. Colpi tenuimarginate. Os circular. Exine 2 µm thick, sexine thicker than nexine, distinctly reticulate, tegillate.
Erythrina stricta Roxb. Vern. Modar (Fabaceae) Pollen 3-porate, suboblate, 25 × 38 µm. Amb subtriangular. Pores circular. Exine 2.5 µm, sexine as thick as nexine, finely reticulate, tegillate.
Gmelina arborea Roxb. ex Sm. Vern. Gomari (Lamiaceae) Pollen 3-colpate, perprolate, 52 × 21 µm. Colpi long, broad, apices obtuse, margin thick. Exine 2.5 µm, crassisexinous, ±obscure, tegillate.
Ilex excelsa Wall. Voigt. Vern. Kotoki (Aquifoliaceae) (Plate 1, figure 7) Pollen 3-colporoidate, prolate, 33 × 20 µm. Colpi medium. Exine 4 µm, sexine slightly thicker than nexine, retipilate, heterobrochate, pila flattened at top.
Lagerstroemia parviflora Roxb. Vern. Dhauli (Lythraceae) (Plate 1, figure 5) Pollen 3-colporate, prolate spheroidal, 42 × 38 µm. Colpi long, margin thick and slightly undulated. Os lalongate. Exine 3–4 µm thick, sexine thicker than nexine and much thicker at the poles, reticulate, heterobrochate, tegillate.
Lannea coromandelica (Houtt.) Merr. Vern. Rohimola (Anacardiaceae) Pollen 3-colporoidate, subporolate, 37.5 × 26.9 µm. Colpi 32.4 µm long, crassimarginate, constricted at the middle. Exine 3.1 µm thick, sexine thicker than nexine, striate, tegillate.
Mesua ferrea L. Vern. Nahor (Calophyllaceae) Pollen 3-colporate, peroblate, 43.3 × 36.1 µm. Colpi long, broad. Os lalongate, 11.9 µm. Exine 3.6 µm, sexine much thicker than nexine, reticulate, tegillate.
Morus alba L. Vern. Nuni (Moraceae) Pollen 2-3-porate, spheroidal, 18 µm. Pore small, circular, 2.1 µm. Exine slightly thicker at pores, sexine as thick as nexine, granulate to obscure, tegillate.
Schima wallichii (D.C.) Korth. Vern. Noga-bhe (Theaceae) Pollen 3-colporoidate, subprolate, 24 × 19 µm. Colpi long, thick, 17 µm. Exine 3.2 µm, sexine thicker than nexine, pseudoreticulate-canaliculate.
Terminalia bellirica (Gaert.) Roxb. Vern. Bhauri (Combretaceae) (Plate 1, figure 2) Pollen, 3-colporate, heterocolpate, prolate, 19.8 × 16.7 µm. Colpi 10.9 µm long, tenuimarginate. Os circular – slightly lalongate, 2.6 × 1.9 µm, margin indistinct. Exine 1.3 µm thick, sexine thinner than nexine, ±obscure.
Results
Modern Pollen/Vegetation Relationship
The modern pollen dispersal is a physical process of both vertical and horizontal transport of tiny pollen released from anther. Huge quantities of pollen grains are transported throughout the atmosphere when they get released, with some remaining airborne for extended periods of time under favourable conditions while the remaining settles in different locations by various medium. In order to understand and develop a pollen/vegetation relationship model, which in turn can assist in interpreting pollen diagrams and determining the preservation status of pollen from respective areas, the study should be conducted strategically, focusing on (a) pollen dispersal within the forested region, (b) pollen dispersal across open land including wetland and river margin and (c) pollen dispersal across cropland.
The sample collection follows a survey of sampling sites and procurement of samples in a transect from the forest centre (KD: 1 to 5) and margin (KD: 6 to 10) across the open- land (KD: 11 to 15), cropland (KD: 16 to 20), wetland (KD: 21 to 25) and river margin (KD: 26 to 30) (Table 1). The study areas selected for the purpose were in and around the Karimganj District of South Assam, Barak Valley (Figure 1).
Pollen Spectra (Forest Centre and Margin)
The palynological study of surface soil from the forest centre shows that the average value of arboreal taxa (59.37%) is higher than non-arboreal taxa (37.63%) followed by extra-regional taxa (3%). The major non-arboreal taxa are represented by grasses viz., non-cereal (12.60%) and cereal (11%) with a combined value of 23.60% (Fig. 3), followed by other terrestrial herbs like Asteroideae (3.26%) in moderately high values. The other herbaceous taxa like Xanthium (1.12%), Convolvulaceae, Solanaceae, Brassicaceae, Coriandrum, Amaranthaceae, Euphorbiaceae and Acanthaceae were encountered in lower to scattered frequencies. Marshy taxa are represented by Polygonum (2.60%) and Cyperaceae (2.67%) with moderate values. Whereas, aquatic taxa are represented by Lemna, Typha and Nymphoides in scanty values. Extra-regional taxa were represented by Pinus followed by Alnus, Betula and Quercus in scanty amount. Some degraded fern spores were noticed in the forest soil. The fungal spores like Diporotheca, Diploidia and Nigrospora were well observed. Among arboreals, pollen taxa like Terminalia (4.09%), Duabanga (4.03%), Lagerstroemia (3.12%), Syzygium (3.10%), Scleichera (3%), Sapotaceae (4.26%), Meliaceae (4.20%) and Emblica (3.38%) are predominant, followed by other tree taxa like Erythrina, Elaeocarpus and Moraceae. The shrubby taxa were represented by Ziziphus and Melastoma in slightly lower values.
However, the samples collected from the forest margin show that the average value of non-arboreal taxa (55.17%) is slightly higher than arboreal taxa (40.13%) followed by extra-regional taxa (4.70%). The major non-arboreal taxa are represented by grasses viz., non-cereal (12.89%) and cereal (16%) with a combined value of 28.89%, followed by other terrestrial herbs like Asteroideae (3.67%). Other herbaceous taxa like Xanthium and Solanaceae were encountered with a value of 1%. The herbaceous taxa are more or less similar to the forest centre samples but are recorded in relatively higher frequencies (Figure 3).
Pollen Spectra (Open Land and Cropland)
The palynological study of surface soil from the open land shows that an average value of non-arboreal taxa (65.34%) is dominant over arboreal taxa (27.99%) followed by extra-regional taxa (6.67%) (Figure 3). The major non-arboreal taxa are represented as usual by grasses comprising cereal (18%) and non-cereal (14%) with a combined value of 32% followed by other terrestrial herbs like Asteroideae (7.83%) in moderately high values followed by Acanthaceae, Euphorbiaceae, Solanaceae, Coriandrum, Amaranthaceae and Xanthium within the values of 1–3%. Strikingly, Brassicaceae was recorded in a scanty amount. Marshy taxa were represented rank wise by Polygonum (4%), Cyperaceae (3.96%), Impatiens (3%) and Onagraceae (2%). Whereas, aquatic taxa were represented by Lemna (1.90%), Typha (1.80%) and Nymphoides (1.33%) in lower values. Extra-regional taxa like Pinus, Betula, Alnus and Quercus were also encountered in the palynoassemblage with relatively high values as compared to the forest soil (Figure 3). The ferns including monolete and trilete spores were found in relatively higher frequencies as compared to the preceding palynozone. The fungal remains like Helminthosporium, Alternaria, Glomus, Diporotheca and Diploidia were frequently observed in this palynoassemblage.
Among arboreals, a gradual declining pattern of tree taxa like Terminalia, Duabanga, Lagerstroemia, Scleichera, Emblica, Sapotaceae, Moraceae, Sapotaceae and Meliaceae was noticed. The tropical semi-evergreen tree taxa like Elaeocarpus, Schima, Ilex and Mesua were completely absent in the palynoassemblage. The other deciduous arboreal taxa like, Bombax, Artocarpus, Acacia, Gmelina and Cassia were encountered in more or less similar frequencies in the open-land samples. Shrubby taxa like Ziziphus (4.50%) and Clerodendrum (4.20%) were found in moderately high values, followed by other shrubby taxa like Melastoma and Vitex within the values of 1–3%. The fungal spores like Alternaria, Helminthosporium, Diplodia, Ascospores of Cookeina, etc., were encountered in moderately high frequencies. The degraded pollen spores of fern spores and some pollen taxa were also noticed in the surface soil of the open land.
Interestingly the surface samples collected from the cropland shows an average value of non-arboreal taxa (81.91%) which are over-dominant over arboreal taxa (13.09%) followed by extra-regional taxa (5%) (Figure 3). The major non-arboreal taxa are represented by grasses comprising cereal (39%) and non-cereal (16%) with a combined value of 54%, followed by other cultural herbaceous taxa like Brassicaceae, Coriandrum and Solanaceae (more than 5% each) in relatively higher frequencies than the preceding palynozones.
Pollen Spectra (Wetland and River Margin)
The palynological study of surface soil samples collected from the wetland shows that an average value of non-arboreal taxa (80.66%) is over-dominant over arboreal taxa (13.50%) followed by extra-regional taxa (5.84%) (Figure 3). The major non-arboreal taxa are represented as usual by grasses comprising cereal (16%) and non-cereal (14%) with a combined value of 30% followed by other terrestrial herbs like Asteroideae, Xanthium, Amaranthaceae and Convolvulaceae in moderately higher values of 3–8%, followed by moderate to lower frequencies of Euphorbiaceae, Solanaceae, Coriandrum and Brassicaceae pollen taxa. Marshy taxa were found in relatively higher frequencies (22.38%) (Figure 3) and were represented by Polygonum, Cyperaceae, Impatiens and Onagraceae within the values of 2–8%. The aquatics were also found in moderately higher values (6.90%) with taxa like Lemna, Typha and Nymphoides. Extra-regional taxa were also encountered in slightly higher frequencies than the preceding palynozones of forest centre, margin and cropland sites and were represented by Pinus Alnus, Betula and Quercus pollen (Figure 3). The fungal remains like Helminthosporium, Alternaria, Glomus, Diporotheca and Diploidia were frequently observed in this wetland palynoassemblage.
Among arboreal taxa, a gradual declining pattern was continued and the tree taxa like Duabanga, Lagerstoemia, Syzygium, Scleichera and Sapotaceae were observed in lower values. The rest of the tree taxa like Meliaceae, Bombax and Cassia are found in scanty amount. The surface samples collected from the river margin again show the relatively increased frequencies of arboreals than the preceding pollen zones of cropland and wetland. The dominance of non-arboreal taxa (74.12%) over arboreal taxa (17.29%) was observed followed by the extra-regional taxa (16%). The non-arboreal taxa are represented by grasses comprising cereal (12%) and non-cereal (9%) with a combined value of 21%. The extra-regional/highland taxa were observed in abundance in the river samples with pollen taxa like Pinus, Betula, Alnus and Quercus. The local tree taxa like Duabanga, Lagerstoemia, Syzygium, Emblica, Cassia and Acacia were represented in lower values along with the occurrence of Barringtonia, Sapotaceae and Meliaceae in scattered values. The shrubby taxa are mostly represented by Vitex, Clerodendrum and Ziziphus. However, Melastoma pollen is completely absent in the river margin samples.
PCA Results of the Recovered Pollen Taxa from the Surface Soil and Sediment Samples
The PCA score plot illustrated the eigenvalues of 0.6336 (Axis 1) and 0.1934 (Axis 2). Simultaneously the explained cumulative variation for the PCA score plot is 63% for Axis 1 and 82% for Axis 2. The total variance of 74% could be explained by the pollen taxa between the forest centre, forest margin, open-land, cropland, wetland and river margin surface samples in and around Karimganj District. By examining the PCA score plot (Figure 4), we observed that the pollen taxa responsible for the similarity between forest centre and forest margin are mainly comprised of the tree and shrubby pollen taxa like Terminalia, Sapotaceae, Meliaceae, Syzygium, Emblica, Melastoma, Moraceae, Duabanga and Lagerstroemia. The cereal and non-cereal pollen taxa fall near the cropland. Some of the terrestrial herbaceous pollen taxa like Asteroideae, Xanthium, Brassica, Coriandrum, Solanaceae Convolvulaceae, Euphorbiaceae and Acanthaceae lie near the open-land, wetland and river margin sites. The aquatic and marshy taxa like Lemna, Nymphoides, Typha, Cyperaceae, Polygonum, Onagraceae and Impatiens are situated near the wetland and river margin areas. The extra-regional taxa like Pinus, Betula, Alnus and Quercus are located near the wetland and river margin samples.
The overall pollen data show three major clusterings based on differential depositional settings, (1) forest centre and forest margin which are mainly occupied by the tree taxa like the moist deciduous and semi-evergreen patch and Melastoma; (ii) cropland and open land, which mainly constitutes cereal, non-cereal, Ziziphus, Acacia/Albizia, Asteroideae and other terrestrial herbs; and (iii) river margin and wetland which are mainly dominated by mixed herbaceous, marshy and aquatics like Xanthium, Euphorbiaceae, Impatiens, Cyperaceae, Lemna, Typha with an exceptional presence of shrubby pollen, Clerodendrum. Extra-regional pollen taxa also fall near the wetland and river margin areas.
The PCA score plot for the pollen taxa composed of major vegetation groups like trees (arboreals), shrubs (arboreals), cereal, non-cereal, other terrestrial herbs, marshy taxa, aquatic taxa and extra-regional taxa illustrated the eigenvalues of 0.6298 (Axis 1) and 0.2067 (Axis 2). Simultaneously the explained cumulative variation for the PCA score plot is 62.98% for Axis 1 and 83.65% for Axis 2. The overall PCA score plot shows the occurrence of trees and shrubs near the forest centre and forest margin. The cereal and non-cereal pollen taxa lie near the open-land and cropland surface soil, whereas other terrestrial herbs, marshy and aquatic taxa are found to be closer in wetland and river margin samples. The extra-regional taxa are found to be relatively more in open-land, cropland and wetland surface samples (Figure 4).
Box-Plot Results of the Recovered Pollen Taxa from the Surface Soil and Sediment Samples
The differences in the frequencies of the major groups of the pollen taxa in the surface soil and sediment samples collected from different depositional sites (forest centre and margin; open-land and cropland, and wetland and river margin) were observed through comparative box plots for different vegetation groups like trees, shrubs, terrestrial herbs, aquatic taxa, marshy taxa and extra-regional taxa (Figure 5). Both frequency and diversity are greater for the tree taxa (arboreals) in forest centre and margin surface samples, whereas, shrubby taxa are prominently represented in forest margin and open-land samples; terrestrial herbs are relatively more in open-land, cropland, wetland and river margin samples. Marshy and aquatic taxa are dominant in wetland and river sediment samples, while aquatic taxa are more frequently found in wetland samples. Extra-regional taxa like Pinus, Betula and Alnus are found in river margin, open-land, cropland and wetland samples in relatively improved values as compared to forest samples.
Discussion
A major obstacle to understanding the vegetation and climate history of this crucial, northeastern region of India throughout the Late Quaternary is the absence of any modern pollen rain data from the Barak Valley of Assam. As support for palaeoenvironmental reconstructions, the composition of current vegetation, the available environmental data and contemporary pollen deposition are examined in the Karimganj district of Barak Valley for both naturally occurring and human-induced plant types. The interpretation of fossil pollen data is perplexing and ambiguous due to differences in pollen productivity, dispersal and preservation, as well as the physical conditions of the sampling site (Boutahar et al., 2023; Broothaerts et al., 2018; de Nascimento et al., 2015; Hicks, 2001). Hence, studies of modern pollen-rain composition under different vegetation types in a study location are crucial for the appropriate interpretation of fossil pollen data (Bajpai & Kar, 2018; Chen et al., 2017; Giesecke et al., 2010; Quamar et al., 2018; Schuler et al., 2014; Wilmshurst & McGlone, 2005). To correlate the occurrence and quantity of pollen types or assemblages with current climate and vegetation data, the present study primarily focuses on the collection of flowers (polliniferous material) and modern surface soil and sediment samples. The changing frequencies of various arboreal taxa in the pollen assemblage reflect the region’s current vegetation structure. Whereas the herbaceous assemblage offers details on the local flora. Especially throughout the Holocene period, which is influenced by the rainfall regime, the shifting assemblage of marshy and aquatic plants at various stages of pollen sequence largely reflects the lake-level fluctuations/hydrological status of lakes/swamps at the temporal scale. Frequencies of Cereal and other cultural pollen taxa show the extent of agricultural activity, and this modern analogue data may offer a way to assess the relative pace of past agricultural expansion.
In addition to being relevant to the interpretation of past vegetation from fossil sequences (Flenley, 1973) and to palaeoecological reconstruction (Huntley, 1990), the observed variation in the composition of the ‘modern pollen rain’ within existing stands of vegetation, both natural formations like forests, grasslands and thickets as well as human-impacted formations, is also significant in biodiversity, pollen productivity and preservation studies (Calcote, 1995; Xu et al., 2007; Zhang et al., 2017) and also landscape reconstruction. Pollen analysis has been an important proxy tool in revealing climate change for a long (Bradley, 1985). Understanding the interaction between pollen rain and a region’s existing vegetation is crucial to the effectiveness of this strategy. Because pollen deposits in every area represent that location’s vegetation structure and preservation condition, the study of the modern pollen rain can be helpful in reliably establishing the pollen/vegetation relationship (Tripathi et al., 2016). This will help in better understanding the same relationship in fossil samples because it is impracticable to directly reconstruct past vegetation/climate correlations. Moreover, the pollen morphological data of 21 arboreal and non-arboreal plant taxa from the tropical forests of Barak Valley are very significant, which could be utilised for accurate identification of pollen grains procured from surface and sedimentary core samples of this region.
The present study mainly concentrates on the collection of a series of modern surface and sediment samples and correlating the occurrence/abundance of pollen/spore types or assemblages with present climate and vegetation data from tropical forests and surrounding vast open land and cropland with coveted wetland/swamp and river margin areas distributed in the Karimganj District of Barak Valley, Assam.
The database information in the first phase permeated from the palynological studies on surface soil and sediment substrates has envisaged the existence of admixture of tropical moist and semi-evergreen plant constituents which has been acknowledged as long as the study is confined within the woods but these arboreal representatives proportionately decreases in the samples away from the forest stands, that is, open-land, cropland, wetland and river margin areas. Although some important plant taxa including Convolvulaceae, Asteroideae, Chenopodiaceae, Melastoma, Clerodendrum and Ziziphus show no drastic change in most of the cases either in or out of the forest which generally flourishes immediately after forest clearance or other biotic influences need more investigation.
Various issues relating appearance and disappearance of palynomorphs and other microbiota in relation to biodegradation in swampy sediment have also been illustrated precisely. Thus, a complexity in interpreting pollen diagrams constructed from these unique forest stands having scarce tree pollen taxa should not be treated as a forest-less zone. Instead in the absence or scarcity of main forest ingredients, such as a Duabanga, Emblica, Lagerstroemia and Syzygium for the tropical moist deciduous forest, some closely associated arboreal as well as non-arboreal taxa could be taken into consideration for studying the preservation of pollen in ground soil and also defining various developmental phases of a forest at spatial and temporal scale. Similarly, Bauhinia, Acacia, Cassia and Barringtonia are the representative taxa for a transitional open-land complex, and cereal, Coriandrum, Brassicaceae and Solanaceae for the cropland. Keeping all this background information in mind, the following interpretations have been made separately to execute palynodata to resolve many critical problems related to the pollen productivity and preservation in the ground soil and palaeoecological studies from tropical forests distributed in Karimganj District of South Assam.
Interpretation of Pollen Rain in Forest Land
The Palynological investigation has been conducted on five surface soil samples procured from within the forested area of Patharia Hill reserve forest. The study of pollen rain on surface soil has predicted a slightly higher proportion of arboreals (59.37%) and non-arboreals (37.63%). The total pollen assemblage indicates the existence of factual forest composition of tree elements like Terminalia, Lagerstroemia, Duabanga, Syzygium, Scleichera, Emblica, Erythrina, Sapotaceae, Meliaceae, Moraceae, Elaeocarpus, Mesua, Ilex and Schima as major moist deciduous and semi-evergreen forest taxa along with Melastoma and Ziziphus which are observed in lower values (1–2%) as major prominent shrubby taxa in the palynoassemblage. However, the values of shrubby taxa proliferate significantly in the forest margin samples (3–6%) attributed to the increased shrubby population in the marginal areas of the forest. Among non-arboreals, except Asteroideae (3.26%), all other terrestrial herbs such as Chenopodiaceae, Convolvulaceae, Coriandrum, Solanaceae, Brassica and Euphorbiaceae are represented at moderately lower values. Whereas, in the forest margin samples these terrestrial herbs were registered in relatively higher values owing to the gradual opening of the forest.
Grasses played the dominant role representing fair frequencies in almost all the surface samples as a major ingredient of the ground vegetation (23%). Poaceae (grass) pollen have been categorised based on their pollen sizes, that is, <45 µm (mostly wild type; non-cereal) and >45 µm (cereal type) for convenience in pollen interpretation, for instance, the occurrence of more cereal represents the acceleration of pastoral activity and wild types in general as vast open grassland though the same may occur at patches within the forest may be attributable to other anthropogenic factors. The majority of fungal elements are grass pathogens belonging to the Ascomycetes and Deuteromycetes, which are regularly present in the sediment at reasonable levels. The high value of the fungal complex including Helminthosporium, Nigrospora, Diporotheca and Diploidia was observed in forest soil which indicates a humid depositional environment. Besides, degraded pollen spores are noticed in the forest palynoassemblage. Judging from the pollen assemblage it is evident that the setup of the pollen distribution pattern has enabled us to deduce the pollen/vegetation relationship within the forest.
Interpretation of Pollen Rain Along Open Land and Cropland
The study was conducted on 30 samples of which five surface soils were procured from the outskirts of forest land, that is, open land areas of Karimganj District. In this case, the study reflects a relative reduction in arboreals (27.99%) as compared to the preceding soil samples within the forest. Shrubs like Melastoma, Ziziphus and Clerodendrum are represented in moderately fair values (2–5%). However, the almost same group of non-arboreals (71%) were recorded in predominant frequencies uniformly. Prominent ground flora such as Asteroideae, Chenopodiaceae, Convolvulaceae, Acanthaceae, Coriandrum, Solanaceae, Brassica and Euphorbiaceae along with extra-regional taxa (6.67%) like Pinus, Betula, Alnus and Quercus, attain improved value may be attributed due to the changing face of the forest (outskirt open land) where strong wind found carrying Himalayan pollen taxa from nearby or distant higher reaches which finally incorporated in the soil. Ferns are consistent in their values and are represented by monolete and trilete spores. The occurrence of moderate frequency of fungal elements in open-land soil samples is due to the location of adjacent crop fields where generally air gets charged with pathogens and finally incorporated in the sediment. The relatively high occurrence of degraded pollen spores was noticed in the open-land soil.
The availability of water hyacinth, Salvinia and Ipomoea near the grassland at the margin of swamps provides a suitable condition for the growth of saprophytic and parasitic fungi such as Alternaria, Nigrospora, Diploidia, Diporotheca, Ascospores of Cookeina and Helminthosporium which supports the biological degradation of the palynomorphs. Thus, the occurrence of degraded pollen spores festooned with abundant fungal remains in the soil samples could be one of the reasons for the degradation of some pollen taxa, which requires detailed investigation (Bera et al., 2008).
The significant feature of the palynoassemblage observed from the cropland surface soil case is the gradual decline of tree elements which is directly proportionate to the distance covered from the forest to the open land across cropland. The second feature is the reasonable enhancement of non-arboreal taxa (82%) especially the crop taxa including cereal, non-cereal pollen and other cultural pollen taxa (Figure 3). In the ultimate case the samples procured further off the forest areas, the tree pollen becomes less or sporadic in frequency (13%). The under-representation of some pollen taxa which were frequently growing along the crop fencing could be attributed to their poor dispersal efficiency, low pollen productivity owing to entomophily as well as biodegradation of pollen in sediments. Sometimes locally abundant non-arboreal taxa like Poaceae (cereal and non-cereal) and Asteraceae create problems and consequently result in distorted palynoassemblage due to over-representation.
Interpretation of Pollen Rain Along Wetland and River Margin
The wetland and river sediment samples show the over dominance of non-arboreal taxa over arboreal taxa. The tree taxa (9–11%) were scatteredly represented and the overall pollen assemblage does not cohere with the extant vegetation in and around the wetland and river, which may be due to recurring floods and intense river flow that washed away the major local arboreals, though pollen production and dispersal also play a strong role for such biased representation in the wetland and river sediment samples. The extra-regional taxa are represented in relatively higher frequencies in the river margin samples owing to the intense fluvial activity from the eastern Himalayan reaches. The degraded pollen/spores have also been detected in the samples. Hence, while interpreting the pollen productivity of an area, the biological degradation factors must be taken into consideration. Also while translating the fossil pollen spectra in terms of past vegetation and climate, all these factors and the database should be considered with immense care.
Interpretation from PCA and Box-Plot Analysis
A variance of 74% could be explained by the first two principal components. The PCA score plot clearly showed three pollen cluster separations between (1) cropland and open land, (2) forest centre and forest margin and (3) wetland and river margin (Figure 4). The pollen taxa like cereal, non-cereal and other terrestrial herbs fall close to cropland and open land which is well coherent to the present-day vegetation set-up in an around the study region. The arboreal tree taxa fall close to the forest centre and margin which again coheres with the present vegetation. However, marshy and aquatic taxa along with extra-regional taxa fall near the wetland and river margin samples. Strikingly, the shrubby taxa especially the Clerodendrum which must fall near the forest margin and open-land samples, find its close position near the wetland and river samples.
The PC score plot reveals the overall three major cluster separations between the (1) tree pollen taxa; (2) terrestrial herbaceous taxa and (3) cereal and non-cereal pollen taxa (Figure 4). The moderately fair amount of tree pollen taxa like Terminalia, Syzygium, Duabanga, Lagerstroemia, Syzygium, Emblica, Sapotaceae and Meliaceae along with the semi-evergreen tree patch (Ilex, Schima, Mesua and Elaeocarpus) could be well observed in forest samples. The terrestrial herbaceous taxa excluding cereal and non-cereal pollen taxa are relatively more dominant in open-land, wetland and river margin samples of the Barak Valley. According to the score plot, the relatively high occurrence of Poaceae (cereal and non-cereal pollen) and other crop pollen taxa like Coriandrum near the cropland quadrant is the direct indicator of agricultural and anthropogenic activity in the region. The presence of non-cereal pollen (Poaceae) could perhaps reflect human activity, which could indicate changes in the mineral content of the sediment, increased fire frequency or a replacement of pollen types of mesic habitats with those of xeric conditions (Tripathi et al., 2018). By the statistical analysis, we conclude that the variation in pollen frequencies of tree and non-tree taxa is responsible for the major segregation between forest land, open land including cropland and wetland as well as river sites.
The frequency and diversity of arboreals (trees), as revealed by box-plot analysis, are comparatively higher in samples taken from the forest floor, illustrating the luxuriant expansion of the core forest species. Due to the significant fluvial inflow from the nearby high-land Himalayan region, terrestrial plants, marshy taxa and extra-regional taxa including Pinus, Betula and Alnus are substantially more prominent in the wetland and river samples. These pollen taxa may be transported into this depositional environment more easily due to the wide area of the basin and the active river flow in the neighbouring regions. Nonetheless, wetland samples more typically contain aquatic pollen taxa, which is consistent with the current flora observed near lakes (Figure 5).
Quamar et al. (2021) postulated that the pollen of the terrestrial herbaceous taxa like Poaceae, Cereal, Asteroideae/Tubuliflorae and Malvaceae, though showing high values, represented only part of the extant ground vegetation in Jammu region of India which is concurrent to the pollen results from the Barak region, thus, approving the uniform over-representation of ground herbaceous taxa in different vegetation types. The anthropogenic marker pollen taxa have been established which could provide an insight on the commencement and subsequent pace of anthropic practices in and around the Barak Valley of northeast India. Moreover, for surface lake sediments, efforts should be made to sample from the central part of lakes, so that the regional pollen rain is not distorted by the local pollen (Quamar & Kar, 2020). In order to develop techniques for reconstructing the global biome distribution used for data model comparison in the framework of global change projects (Prentice et al., 1996), it is necessary to have a lot of actual pollen counts from all ecosystems, covering all latitudes. The data on pollen rain and modern vegetation in Barak Valley may be useful in this regard.
Conclusion
The pollen data gathered from 30 surface pollen samples show a mixed deciduous forest made up of Terminalia, Lagerstroemia, Emblica, Syzygium and Duabanga in a warm, humid climate in response to local rainfall variation. The pollen spectra have shown the dominance of non-arboreals (especially grass pollen and terrestrial herbs) and relatively lower frequencies of arboreals (APs; trees and shrubs) in open-land, cropland, wetland and river margin areas. The representation of core tree taxa in the palynoassemblage procured from the woods was appreciable when compared to the modern pollen spectra in the Brahmaputra Valley of Assam, attributable to the gentle slope, confined river flow that aids in the fair preservation of the pollen in the soil of Barak Valley. Due to the coherence between contemporary vegetation and surface pollen data, this modern pollen analogue serves as a potential baseline data for the reconstruction of past vegetation and climate change in the Barak Valley of northeast India.
Footnotes
Acknowledgements
The authors would like to thank the Director, BSIP, for providing laboratory facilities and permission to publish the manuscript. The present manuscript is the outcome of an in-house project of BSIP (Project 6). We are also thankful to the forest officials of Assam for their permission and help during the sample collection. We thank Dr. Achuta Nand Shukla and two other anonymous reviewers for their reviews which helped elevate the manuscript manifold.
Declaration of Conflicting Interests
The authors declared no potential conflicts of interest with respect to the research, authorship and/or publication of this article.
Funding
The authors received no financial support for the research, authorship and/or publication of this article.