
Abstract
Background
Migraine is a brain disorder with a multifaceted and unexplained association to sleep. Brain excitability likely changes periodically throughout the migraine cycle. In this study we examine the effect of insufficient sleep on neuronal excitability during the course of the migraine cycle.
Methods
We examined 54 migraine patients after two nights of eight-hour habitual sleep and two nights of four-hour restricted sleep in a randomised, blinded crossover study. We performed transcranial magnetic stimulation and measured cortical silent period, short- and long-interval intracortical inhibition, intracortical facilitation and short-latency afferent inhibition. We analysed how responses changed before and after attacks with linear mixed models.
Results
Short- interval intracortical inhibition was more reduced after sleep restriction compared to habitual sleep the shorter the time that had elapsed since the attack (p = 0.041), and specifically in the postictal phase (p = 0.013). Long-interval intracortical inhibition was more increased after sleep restriction with time closer before the attack (p = 0.006), and specifically in the preictal phase (p = 0.034). Short-latency afferent inhibition was more decreased after sleep restriction with time closer to the start of the attack (p = 0.026).
Conclusion
Insufficient sleep in the period leading up to a migraine attack may cause dysfunction in cortical GABAergic inhibition. The results also suggest that migraine patients may have increased need for sufficient sleep during a migraine attack to maintain normal neurological function after the attack.
Introduction
The connection between migraine and sleep has been well known for 150 years (1) but is still largely unexplained. However, brain regions such as the hypothalamus appear to be involved in both sleep physiology and migraine pathophysiology, possibly producing premonitory symptoms such as yawning and sleep disturbances (2,3). Sleep is also commonly used to end the migraine attack (4), but a large portion of migraine patients still experience postdromal symptoms such as fatigue after waking up (5). Furthermore, sleep disturbances are commonly known to trigger migraine attacks (6). These observations imply a close relationship between sleep physiology and migraine pathophysiology, signifying a potential to discover migraine specific mechanisms by manipulating sleep.
Migraine is suggested to be a cycling disease, possibly driven by periodic changes in the neuronal excitability of the hypothalamo-thalamo-brainstem network. This central nervous dysfunction may be represented by gradual decrease of sensory threshold towards an attack before gradual recovery after attacks (7). The preictal phase just prior to the onset of migraine headache encompass connectivity changes involving dopaminergic centres (8) and the hypothalamus (9). The preictal phase may also be characterised by increased somatosensory excitability in the brainstem and cortex, possibly caused by decreased intracortical inhibition (10). Preictal patterns in electroencephalography (EEG) may be indicative of underlying thalamocortical dysrhythmia (11), able to cause increased cortical synchronicity (12,13), which has been interpreted as reduced cortical preactivation (14). In the ictal phase, the increased somatosensory excitability (10,14), connectivity changes (9,15), and the reduced cortical preactivation before the attack (11,14) may flip or normalise towards interictal levels. Less is known about the postictal phase (16), but one group have found slight alterations of postictal brainstem functionality (17). Another group found an inverse relation between lateral inhibition and days elapsed since the last migraine attack (18). More detailed central nervous system (CNS)-excitability mapping with other methods is needed to understand the neurological changes leading up to the attack and accompanying the gradual symptomatic normalisation after the attack.
Sleep restriction as an experimental model of insufficient sleep has previously been shown to affect cortical responses in healthy volunteers (19) and we have recently shown that sleep restriction alters GABAergic inhibition in interictal migraine patients compared to controls (20). Thus, sleep restriction may amplify differences in neurological functioning which are usually subtle or compensated, allowing us to measure dynamics of migraine mechanisms which are specific to parts of the migraine cycle and reactive to sleep changes.
Several aspects of cortical excitability can be studied with transcranial magnetic stimulation (TMS). TMS-measurements of interest include the cortical silent period (CSP), paired pulse TMS (ppTMS) techniques as short- and long-interval intracortical inhibition (SICI and LICI) and intracortical facilitation (ICF), and short-latency afferent inhibition (SAI). These responses represent different facilitatory and inhibitory mechanisms (21). Thus, combining these responses allows for detailed investigation of cortical excitability and the actual mechanisms behind it.
We hypothesised that changes in sleep duration affect cortical functioning differently throughout the time leading up to an attack and during the time following an attack, and that the effects will be most pronounced in the preictal and postictal phases of the migraine cycle. We measured effects of excitatory and inhibitory systems after both habitual sleep and sleep restriction by recording responses to different TMS protocols. The primary objective was to evaluate how the effect of restricted sleep on TMS-measurements of cortical excitability vary with time from the end of the previous and time to the start of the next migraine headache attack. A secondary objective was to estimate how the effects of restricted sleep on TMS-measures varied with preictal, ictal and postictal states, as compared to the interictal state. We also performed exploratory evaluations of how sleep related effects of TMS varied with clinical migraine characteristics.
Methods
Subjects
Migraine subjects were recruited through media and social media and screened by headache nurses. Migraine subjects considered eligible were further evaluated by a neurologist for inclusion, requiring a diagnosis of migraine with or without aura according to The International Classification of Headache Disorders, 3rd edition (22) and meeting predefined criteria. We included migraine subjects between 18 and 65 years of age having between one and six attacks per month during the last six months. Prophylactic treatment for migraine was not permitted from four weeks before the first examination and throughout the whole examination period. Migraine patients were excluded if they had tension type headache for seven days or more per month. Exclusion criteria also included known sleep disorders, metabolic, endocrine or neuromuscular disorders, connective tissue disorder, neoplastic disease, cerebrovascular disease, symptomatic heart disease, lung disease or neurological/psychiatric disease with reduced function, treated hypertension or blood pressure >160/110 mmHg, acute infectious or painful disease, acute injury, previous craniotomy, pregnancy, alcohol or narcotics abuse, regular treatment with neuroleptics, antiepileptic drugs, antidepressants or other medications affecting neural, vascular or muscular function and contraindications for TMS. Subjects were asked not to exercise or to consume caffeinated beverages or tobacco on the examination days. All subjects answered a questionnaire and a semi-structured interview for collection of clinical data and headache characteristics.
A total of 161 migraine subjects were screened for inclusion. Further exclusion and drop out is described in Figure 1. One subject was excluded for not being adherent to sleep instructions. Two migraine subjects had no ppTMS recordings due to a technical malfunction of the coil thermostat. Thus, a total of 54 migraine patients (Table 1) were examined. We have previously reported data from the interictal phase compared to controls from the same study (20).

Inclusion and exclusion flow chart. Flow chart describing initial number of eligible subjects and further drop out and exclusion throughout the study period.
Demographic and clinical data of 54 migraine subjects with two examinations divided into partially overlapping phase groups.
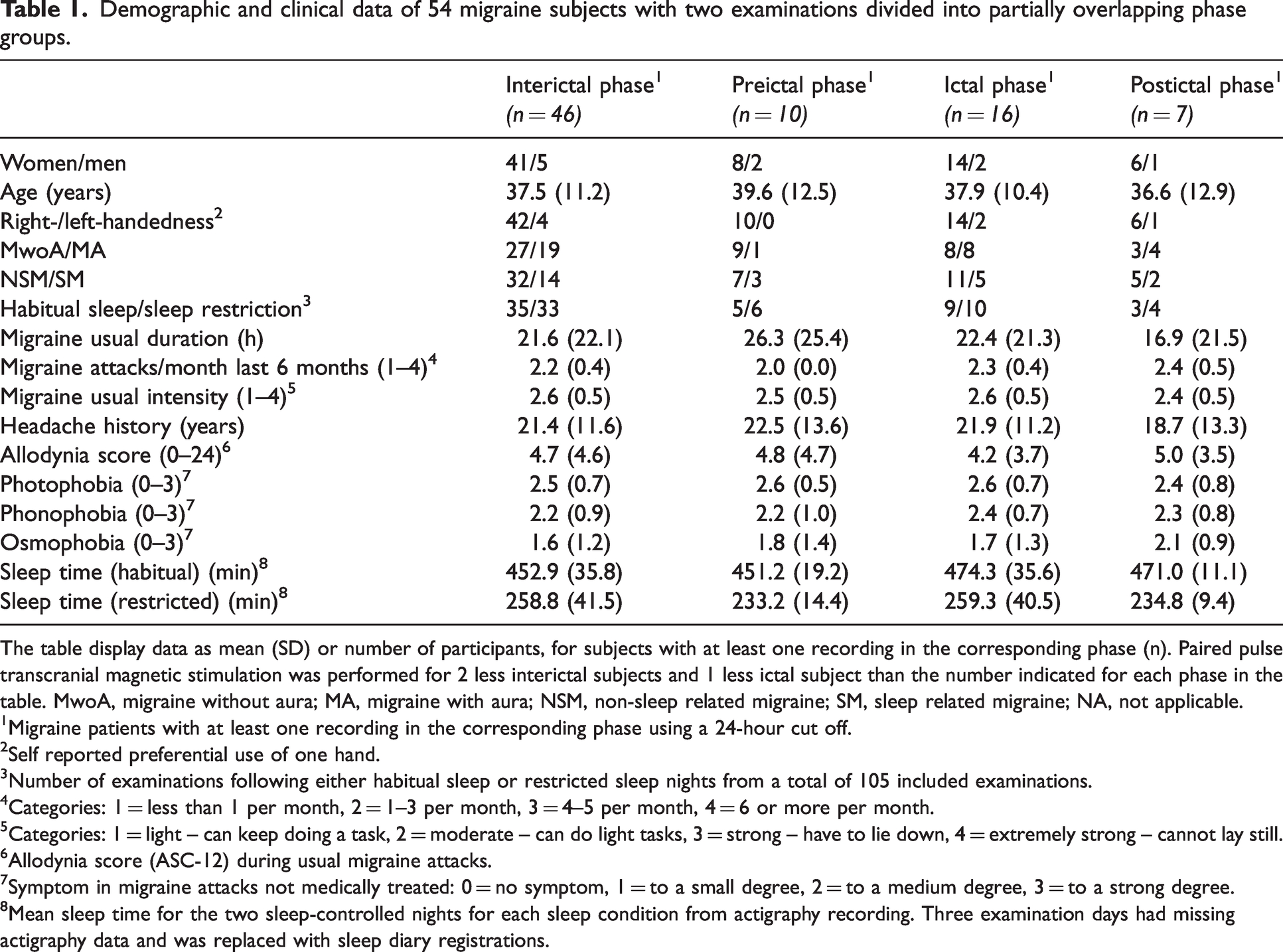
The table display data as mean (SD) or number of participants, for subjects with at least one recording in the corresponding phase (n). Paired pulse transcranial magnetic stimulation was performed for 2 less interictal subjects and 1 less ictal subject than the number indicated for each phase in the table. MwoA, migraine without aura; MA, migraine with aura; NSM, non-sleep related migraine; SM, sleep related migraine; NA, not applicable.
1Migraine patients with at least one recording in the corresponding phase using a 24-hour cut off.
2Self reported preferential use of one hand.
3Number of examinations following either habitual sleep or restricted sleep nights from a total of 105 included examinations.
4Categories: 1 = less than 1 per month, 2 = 1–3 per month, 3 = 4–5 per month, 4 = 6 or more per month.
5Categories: 1 = light – can keep doing a task, 2 = moderate – can do light tasks, 3 = strong – have to lie down, 4 = extremely strong – cannot lay still.
6Allodynia score (ASC-12) during usual migraine attacks.
7Symptom in migraine attacks not medically treated: 0 = no symptom, 1 = to a small degree, 2 = to a medium degree, 3 = to a strong degree.
8Mean sleep time for the two sleep-controlled nights for each sleep condition from actigraphy recording. Three examination days had missing actigraphy data and was replaced with sleep diary registrations.
Study design
The study had a crossover design (Figure 2a) with one training day and two examination days. The training day was utilised for the subjects to become familiar with the laboratory and TMS examination and preceded the first examination by at least three days. The subjects were randomised to have the first examination day preceded by either two nights of eight-hour habitual sleep or two nights of four-hour restricted sleep. The randomisation was done in blocks within the migraine group by a research nurse (GBG, MSS) not involved in the examinations or analyses. The second examination day was preceded by the opposite sleep condition and scheduled at least seven days after the first examination. The investigator (MSM) was blinded for sleep condition and migraine diagnosis both during examinations and during data analysis. Examinations were performed by an investigator who did not know whether the subjects had a migraine diagnosis. Participants were also repeatedly told by a research nurse to not reveal their sleep condition and reminded in SMS notifications. Both examinations started at the same time of day for the same subject, either at 08:00 am or 10:30 am. Participants were instructed to plan their sleep time to wake up as close to 07:00 am as practically possible. Participants kept a headache diary from one week before, until one week after examinations. They marked the time of start and end of migraine headache and were allowed to use acute medication during the attack. They also filled in a sleep diary and used actigraphy throughout the examination period. The diaries used are previously described by our group (23).

Study design. (a) Schematic overview of the crossover study design. Each subject participated in a training day preceding the first examination by at least three days. Subjects were block-randomised to have the first examination preceded by either two nights of habitual eight-hour sleep or restricted four-hour sleep. The second examination followed at least seven days after the first examination and was preceded by the remaining sleep condition. All subjects kept a headache diary from one week before until one week after examinations. Every subject registered sleep by both a sleep diary and a wrist-worn actigraph from the training day until the second examination and (b) Examples of single recordings of short-interval intracortical inhibition (SICI) with 2 milliseconds (ms) interstimulus interval (ISI), intracortical facilitation (ICF) using 10 ms ISI and long-interval intracortical inhibition (LICI) using 100 ms ISI. Conditioning stimuli (CS) was elicited at time 0 ms.
Transcranial Magnetic Stimulation
Subjects sat comfortably during examinations with their arms resting on a pillow. We used a figure-of-eight coil type MCF-B65 with a MagPro X100 stimulator with MagOption (MagVenture A/S, Farum, Denmark). Pulses were biphasic and 280 µs, initially inducing anteroposterior current in the tissue. We recorded electromyography (EMG) from the right abductor pollicis brevis muscle (APB) with Ag/AgCl electrodes through a Dual Bio Amp and a PowerLab 8/35 (ADInstruments, Dunedin, New Zealand). The left hemisphere was stimulated in all participants. Data were recorded in LabChart software version 8 (ADInstruments, Dunedin, New Zealand) with sampling rate 10 kHz, high pass 1 Hz and low pass 2 kHz. Navigation was used to adhere to a determined location using Localite TMS Navigator (Localite GmbH, Bonn, Germany). We stimulated the median nerve for SAI using a Digitimer Constant Current Stimulator model DS7A (Digitimer, Welwyn Garden City, United Kingdom) with 200 µs square wave-pulses.
The TMS protocol started with mapping of the location over contralateral motor cortex eliciting the largest and most consistent peak-to-peak motor evoked potential (MEP) amplitude in the APB. This location was marked in the Localite software and used for the rest of the TMS session. On the training day 70% of maximal stimulator output was used for mapping, while 120% of resting motor threshold (RMT) from the training day was used for mapping during the following examination days.
To determine RMT, we started using 35% of maximal stimulator output and increased intensity by 5% until MEPs were consistently measured above 50 µV. Then, we gradually reduced the stimulation intensity by 1% and used the last intensity eliciting MEP above 50 µV in five or more out of ten stimulations as RMT.
CSP was measured from the APB during isometric thumb abduction of about 50% maximum voluntary contraction against a Velcro band. Six measurements were collected using 120% RMT and 12 second intervals. Contraction level was assessed using visual assessment of EMG activity.
Paired pulse TMS was recorded in one continuous session. We recorded single test stimuli (TS) of 120% RMT, SICI and ICF with 80% RMT as conditioning stimulus (CS) and 120% RMT as TS, and LICI with 109% RMT as CS and 120% RMT as TS (Figure 2b). Interstimulus intervals (ISI) of 2 and 4 ms were used for SICI, 8 and 10 ms for ICF and 100 ms for LICI. The order of these stimulation types was randomised in blocks containing one of each type with randomised four, five or six seconds between each paired pulse train. Twenty blocks were recorded continuously, giving 20 recordings of single test stimuli and each paired stimulation type, and adding up to 120 recorded responses in total.
Alternating 20 single 120% RMT TMS stimulations and 20 SAI were measured using 21 ms ISI and TS of 120% RMT TMS intensity. The lowest median nerve stimulation intensity inducing a visible muscle twitch in APB was used as CS.
Data analysis
EMG recordings were filtered (1 Hz–2.5 kHz) in LabChart and motor evoked potential (MEP) peak-to-peak amplitudes were manually marked by an investigator (MSM) blinded for sleep condition and migraine phase (Figure 2b). LabChart was also used to rectify EMG for CSP analyses.
We used MATLAB R2019b (MathWorks, Natick, MA, USA) with custom scripts to average the rectified EMG for CSP. CSP onset was determined using the onset of MEP, defined as the first data point of the MEP which exceeded the upper limit of the mean consecutive difference threshold method (24). CSP offset was defined as the first data point above the lower limit with 50% or more of the following 5 ms above the same limit. Breakthrough EMG was manually defined by a blinded investigator (MSM) and included in the CSP.
The two examinations of each subject were evaluated for inclusion into the primary analyses of time to the next attack and time from the last attack. Time to the next and time from the last attack for each examination were determined using headache diaries. Time to the next attack was defined as time until reported start of migraine headache. Time from the last attack was defined as starting from the reported end of migraine headache. Examinations with more than seven days without attacks were set to 168 hours. Five examinations (1 time from attack; 4 time to attack) could not be included because of insufficient headache diaries. Ictal examinations were not included in either of the analyses. Preictal (24 hours before attack) and postictal (24 hours after attack) examinations were not included in the time from attack analysis and to attack analysis, respectively. After exclusions, 48 migraine patients for CSP (74 examinations; 38 habitual sleep and 36 sleep restriction), 48 migraine patients for SAI (73 examinations; 38 habitual sleep and 35 sleep restriction) and 46 migraine patients for ppTMS (69 examinations; 34 habitual sleep and 35 sleep restriction) were available for time from attack analyses. For time to attack analyses, 46 migraine patients for CSP (75 examinations; 37 habitual sleep and 38 sleep restriction), 46 migraine patients for SAI (74 examinations; 37 habitual sleep and 37 sleep restriction) and 45 migraine patients for ppTMS (72 examinations; 35 habitual sleep and 37 sleep restriction) were available.
For secondary analyses, examinations were classified as interictal, preictal, ictal or postictal using a 24-hour cut off from the beginning or end of migraine headache according to the headache diary. We applied this 24-hour limit as in previous studies (23,25), because characteristics of the preictal phase are rather specific to this period preceding the ictal phase (26). The preictal phase started 24-hours prior to the reported start of migraine headache. The ictal phase lasted from the start of migraine headache until the end of migraine headache. The postictal phase lasted until 24-hours after the end of migraine headache. Examinations were allocated to the phase the subjects were in when the examination was performed. Examinations performed during overlapping preictal and postictal phases were excluded. Each phase was compared to the interictal phase, and all four phases were included in the same model.
Statistical analysis
We performed statistical analyses in STATA version 17.0 (StataCorp LP). We included CSP duration and every ppTMS and SAI peak-to-peak amplitude in separate linear mixed models for each of the different TMS-responses. Linear mixed models are well suited to handle missing data for when subjects had recordings of different sleep conditions in different phases and is commonly applied for repeated measurements (20,27). Every analysis was performed with an intention to treat basis with intended sleep as sleep condition. Actual sleep duration was recorded with actigraphy (except for three examination days where sleep diary replaced missing data) and is described in Table 1. To better meet assumptions of normality in residuals, the peak-to-peak amplitudes were transformed to the power of 0.2. Box cox transformation was applied to find the most appropriate transformation for normality. We used the model design with the lowest Akaike and Bayesian information criterion (AIC/BIC) values between different theoretically appropriate random intercept and random slope models with the same variables.
The model chosen for primary analysis of CSP was a 2- level random intercept model with time to/from attack, sleep condition and their interaction as fixed effects and subject as level 2. The interaction effect represents difference between sleep conditions in the slope of CSP duration over time. The models chosen for the analyses of ppTMS and SAI were 3-level models with stimulation type, time to/from attack, sleep condition and their interactions as fixed effects, and subject and stimulation type as random intercepts at level 3 and 2, respectively, with time to/from attack as random slope at level 3. Test stimuli, time = 0 and habitual sleep were used as bases in the model. Stimulation type represents the difference between peak-to-peak amplitudes of test stimuli and conditioned stimuli. We conducted separate models for each of the ppTMS stimulation types (SICI 2 ms, SICI 4 ms, ICF 8 ms, ICF 10 ms and LICI 100 ms). The interaction effect in ppTMS and SAI models represent the difference between sleep conditions in the slope of stimulation type differences over time.
We performed secondary analyses replacing time in the model with migraine phase with a 24-hour cut off from start and end of migraine headache. The interictal phase was the base in the model. Post hoc contrasts are reported for habitual sleep separately between phases. Due to the restricted number of individuals in the different phases of the secondary analyses, we applied restricted maximum likelihood estimation with the Kenward-Roger Degrees of Freedom approximation.
We investigated significant findings in primary analyses for correlation with clinical parameters in exploratory analyses. Clinical variables from a questionnaire were included in the model as a fixed effect interacting with the significant interaction. We assessed attacks per month, usual attack duration and usual migraine headache intensity. We report two-sided p-values <0.05 as significant.
The study size was originally planned for a larger migraine group to account for spread of examinations throughout the migraine cycle. By assessing 75 includable subjects we hoped to have 60 subjects completing the study and about 45–50 interictal subjects. Two-tailed Student’s t-test comparing e.g., 30 controls and 45 patients with alpha 0.05 would then have 80% power to detect a medium to large Cohen’s d effect size of approximately 0.67. We did not perform a priori power calculations for analyses of temporal relations to attack or phase comparisons within the migraine group.
Results
Demographic and clinical data are presented in Table 1. Recorded sleep duration is distributed between sleep conditions as expected from the study design.
The average time from examination to the next attack was numerically shorter after sleep restriction (87.0 hours; SD 61.6) compared to habitual sleep (105.6 hours; SD 64.0). The average time from the last attack to the next examination was more similar between sleep restriction (121.6 hours; SD 55.7) and habitual sleep (110.1 hours; SD 60.0).
After a migraine attack
After sleep restriction, SICI 2 ms was reduced with shorter time elapsed since the previous migraine attack (Figure 3). This was seen as a significant interaction effect of time × sleep × SICI 2 ms (p = 0.041; Table 2). In secondary analyses, SICI 2 ms was also reduced after sleep restriction in the postictal compared to the interictal phase (p = 0.013; Table 3; Figure 4).

Estimated marginal mean difference of the significant effect after attacks. Line plots represent estimated marginal mean difference between peak-to-peak MEP amplitude millivolts to the power of 0.2 (mV0.2) after test stimuli and conditioned stimuli for short-interval intracortical inhibition (SICI) at 2 ms. The marginal mean difference is displayed separately for habitual sleep and sleep restriction and over time from the end of a migraine attack until 168 hours after the attack. Marginal mean differences are calculated from linear mixed models. The shaded area indicates estimated marginal standard error. The interaction effect time × sleep × SICI 2 ms was significant (p = 0.041).
Interaction effects of sleep and temporal relation to migraine attacks on TMS-measurements.

Results from the interaction time to/from examination from/to attack × habitual sleep/restricted sleep (× stimulation type (ppTMS and SAI)) in linear mixed models, including number of subjects (n), z-test statistic, p-value, the coefficient and 95% confidence intervals (CI) for difference in stimulation types between sleep conditions per time point (h). CSP, cortical silent period; ICF, intracortical facilitation; LICI, long-interval intracortical inhibition; ppTMS, paired pulse transcranial magnetic stimulation; SAI, short-latency afferent inhibition; SICI, short-interval intracortical inhibition; TMS, transcranial magnetic stimulation. The asterisk (*) and bold text indicate p < 0.05.
Interaction effects on outcome variables for sleep condition and each migraine phase compared to the interictal phase.
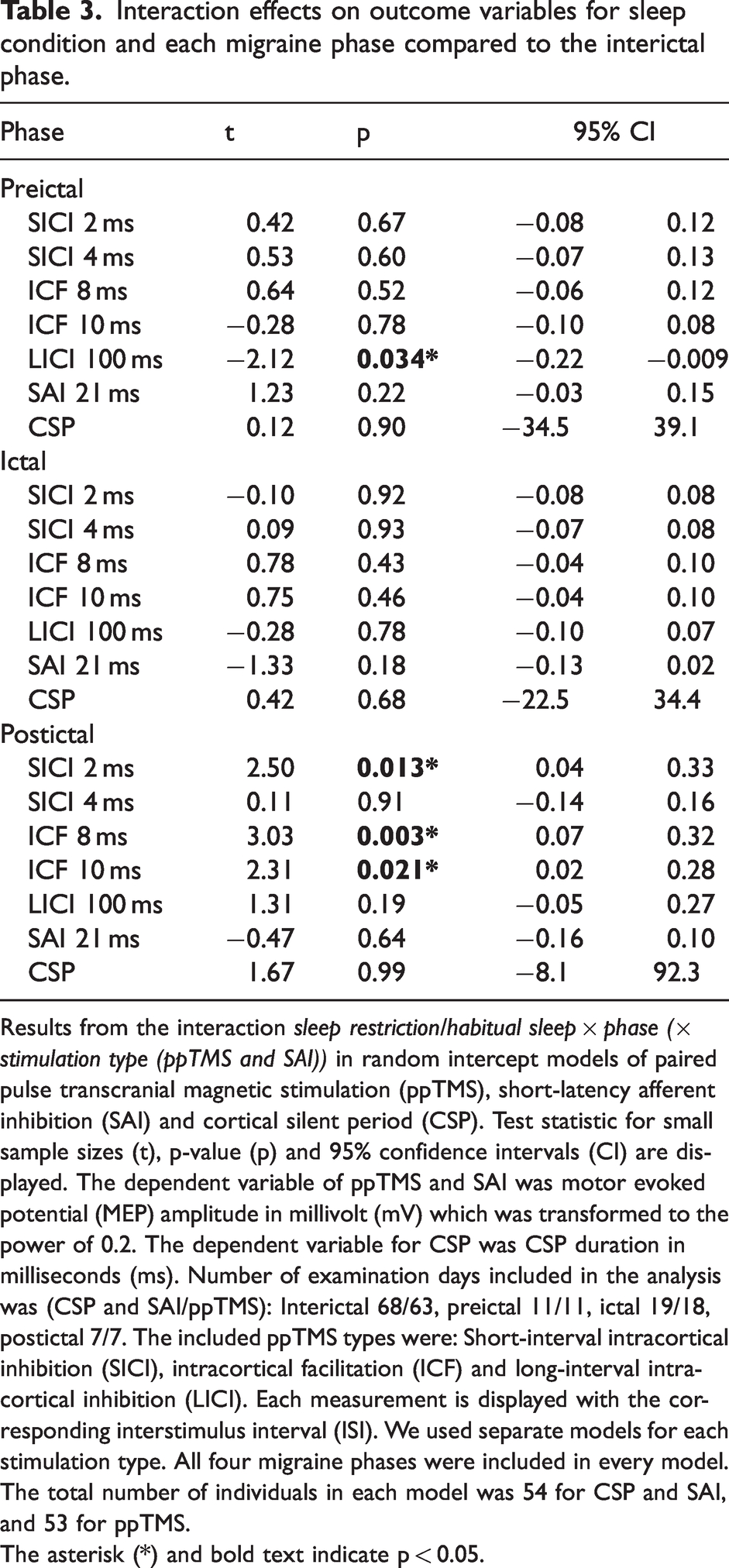
Results from the interaction sleep restriction/habitual sleep × phase (× stimulation type (ppTMS and SAI)) in random intercept models of paired pulse transcranial magnetic stimulation (ppTMS), short-latency afferent inhibition (SAI) and cortical silent period (CSP). Test statistic for small sample sizes (t), p-value (p) and 95% confidence intervals (CI) are displayed. The dependent variable of ppTMS and SAI was motor evoked potential (MEP) amplitude in millivolt (mV) which was transformed to the power of 0.2. The dependent variable for CSP was CSP duration in milliseconds (ms). Number of examination days included in the analysis was (CSP and SAI/ppTMS): Interictal 68/63, preictal 11/11, ictal 19/18, postictal 7/7. The included ppTMS types were: Short-interval intracortical inhibition (SICI), intracortical facilitation (ICF) and long-interval intracortical inhibition (LICI). Each measurement is displayed with the corresponding interstimulus interval (ISI). We used separate models for each stimulation type. All four migraine phases were included in every model. The total number of individuals in each model was 54 for CSP and SAI, and 53 for ppTMS.
The asterisk (*) and bold text indicate p < 0.05.

Paired pulse transcranial magnetic stimulation. Estimated marginal mean differences between test stimuli and conditioned stimuli from linear mixed models of paired pulse transcranial magnetic stimulation (ppTMS) in millivolts to the power of 0.2 (mV0.2). The x-axis displays paired pulse stimulation type with corresponding interstimulus intervals (ISI) in ms. Separate statistical models were applied for each stimulation type and each model included all four migraine phases. Stimulation types were short-interval intracortical inhibition (SICI), intracortical facilitation (ICF) and long-interval intracortical inhibition (LICI). Positive values represent facilitation and negative values represent inhibition. Error bars indicate estimated marginal standard error. Statistical significance bars represent interaction effects of sleep restriction/habitual sleep × phase × stimulation type. Asterisks mark significance level: *p < 0.05, **p < 0.01.
There were no significant effects of time after the previous attack and sleep restriction on the other TMS-measures (p > 0.15; Table 2). However, comparing the postictal and interictal phase in secondary analyses we also found an increase of both ICF 8 ms (p = 0.003; Table 3; Figure 4) and ICF 10 ms (p = 0.021; Table 3; Figure 4) after sleep restriction postictally.
Before a migraine attack
After sleep restriction, LICI 100 ms increased with time closer to the next migraine attack (Figure 5). We saw this as a significant interaction effect of time × sleep ×LICI (p = 0.006; Table 2). This interaction was significantly affected by usual migraine headache intensity (p = 0.002; 95% CI 0.0008 to 0.003), where subjects with higher usual intensity had a larger LICI increase after sleep restriction closer to the attack start. In secondary analyses, we also observed that LICI increased after sleep restriction in the preictal phase compared to the interictal phase (p = 0.034; Table 3; Figure 4). The post hoc contrast effect of preictal/interictal phase × LICI was significant after habitual sleep (p = 0.007; 95% CI 0.03 to 0.19), indicating that it was the LICI effect after habitual sleep that was reduced preictally.

Estimated marginal mean differences of the significant effects before attacks. Line plots represent estimated marginal mean difference between peak-to-peak MEP amplitude millivolts to the power of 0.2 (mV0.2) after test stimuli and conditioned stimuli for long-interval intracortical inhibition (LICI) and short-latency afferent inhibition (SAI). The marginal mean difference is displayed separately for habitual sleep and sleep restriction and over time from 168 hours before a migraine attack and up to the attack onset at time 0. Marginal mean differences are calculated from linear mixed models. The shaded area indicates estimated marginal standard error. The interaction effects time × sleep × LICI (p = 0.006) and time × sleep × SAI (p = 0.026) were significant.
A reduction of the SAI-effect with time closer to the next migraine attack was seen after sleep restriction (Figure 5). This was found to be a significant interaction effect of time × sleep × SAI (p = 0.026; Table 2).
We did not observe any significant interaction effects on SICI 2 and 4 ms, ICF 8 and 10 ms or CSP before migraine attacks (p > 0.44; Table 2).
No other significant interaction effects were found between attacks per month, usual attack duration, usual migraine headache intensity and the TMS-measurements with significant primary interaction effects (p > 0.10).
Discussion
This study shows that the effect of sleep restriction on TMS-measures of cortical excitability may vary within the migraine cycle and specifically with time to the next attack or from the previous migraine attack. After an attack, the SICI 2 ms inhibitory effect was more reduced by sleep restriction the shorter the time that had elapsed from the attack. SICI 2 ms was also reduced after sleep restriction in the postictal compared to the interictal phase in secondary analyses. We found no effects on ICF in the primary analyses, but ICF was increased after sleep restriction in the postictal phase. In the time leading up to an attack we found an increasing LICI inhibitory effect and a decreasing SAI inhibitory effect after sleep restriction the shorter the time that remained until the commencement of the attack. Subjects with higher than usual attack intensity had a greater LICI increase. In secondary analyses, LICI was also more increased after sleep restriction in the preictal compared to the interictal phase, and LICI after habitual sleep was lower preictally compared to the interictal phase.
GABA-A and glutamatergic alterations following migraine attacks
Our findings indicate that sleep restriction induced a larger decrease in GABA-A mediated inhibition in the postictal phase and the shorter the time that had elapsed from the previous migraine attack. We found this effect as changes in SICI which is understood to represent short lasting inhibitory post synaptic potentials from α2 or α3 subtype GABA-A receptor activation, possibly representing a low-threshold cortical inhibitory circuit. This inhibitory effect is thought to be modulated by presynaptic GABA-B mediated autoinhibition of inhibitory interneurons, to be increased by dopamine and nicotine, and to be decreased by noradrenaline (21,28). We also found indications of postictally increased ICF after sleep restriction. ICF represents excitability of excitatory circuits in the cortex; probably as a net facilitation from glutamatergic NMDA-receptor activation and GABA-A receptor mediated inhibition as in SICI (21,29), with contribution from subcortical mechanisms (30).
Previous studies of SICI and ICF after sleep deprivation in healthy subjects have found a reduced SICI inhibitory effect and increased ICF (31,32). This response is thought to accompany a lack of sufficient desaturation of synaptic potentiation by insufficient sleep, which leads to alterations in neuroplasticity and can also compromise learning, memory and attention (32). Thus, migraine patients may display an exaggerated effect of reduced sleep on neurological functioning or a susceptibility to these effects at milder levels of insufficient sleep shortly after the attack. It is well known that most migraine patients are forced to sleep during attacks at least occasionally (6), and that sleep is an effective treatment for migraine headache (4). Sleep restriction before the recordings soon after the attack in this study probably occurred during the ictal phase. Consequently, we propose that sufficient sleep during migraine attacks is of direct importance for maintaining neurological functioning in migraine patients.
GABAergic and cholinergic changes in the period leading up to migraine attacks
We have previously reported decreased CSP duration after sleep restriction in interictal migraine patients compared to controls (20). This finding may be related to the gradually increasing inhibitory LICI effect after sleep restriction leading up to a migraine attack that we observed in this study. LICI is thought to represent slow inhibitory post synaptic potentials acting through GABA-B receptors, which is understood to be dependent on the availability of GABA in the synaptic cleft. GABA-B receptor mediated inhibition is also a mediator of the CSP effect and presynaptic GABA-B receptors may be relevant for these inhibitory effects by regulating the amount of GABA available in the synapse (21,33,34). CSP requires higher levels of GABA availability than LICI before the inhibitory effect is saturated. Thus, presynaptic GABA-B activity may reduce CSP duration without affecting LICI (33). CSP and LICI also differ in the way that CSP possibly represents duration of inhibition, where LICI represents the magnitude of inhibition (34). Shortened CSP has previously been observed as the LICI effect increases (35) and increased CSP has been observed with a concurrent decline in LICI during exercise induced fatigue (33). Thus, our results imply that while the temporal aspect of GABA-B mediated inhibition is decreased by sleep restriction in the interictal phase, the magnitude of inhibition may be elevated by sleep restriction closer to the attack or in the preictal phase specifically. This preictal increase in GABA-B magnitude may occur concurrently with a stable deficit in cortical inhibitory duration as we did not discover changes in CSP towards the start of an attack. While this underlying dysfunction represented by altered CSP was associated with both premonitory and ictal hypersensitivity (20), the LICI change was associated with increased usual attack intensity. Thus, suggesting a closer link between headache and this preictal GABA-B inhibitory affection.
The preictally decreased LICI after habitual sleep (Figure 4) may be in coherence with the theory that migraine patients have increased thresholds for inhibitory homeostatic responses in the preictal phase (36), which may be related to hypoexcitability measured at rest (12,14). This model may also be in line with observations of preictal hyperresponsiveness in previous studies (10,14). indicating an involvement of slow GABA-B receptor mediated inhibition. Inhibitory thresholds may be comparable to interictal levels in examinations after sleep restriction due to sleep deficit induced increase of cortical activity (32,36).
Circadian rhythm related alterations of GABA-B receptor mediated inhibition of motor cortex have been linked to corticotropin-releasing hormone (CRH) from the paraventricular nucleus (PVN) of hypothalamus. GABA-B mediated inhibition decreases during the day, and previous studies suggest that CRH modulates GABAergic neurons postsynaptically on different levels of the central nervous system (37). CRH also contributes to regulation of spontaneous waking (38) and reduced CRH after sleep deprivation has been suggested to mediate temporary relief of depression observed after sleep deprivation (39). Our group has previously described how migraine subjects in the preictal phase display increased sleepiness compared to the interictal phase (40). A dysfunction of PVN and CRH related mechanisms preictally in migraine patients, where CRH release may be more increased in the morning despite foregoing sleep restriction, may be one explanation for increased LICI after sleep restriction closer to the start of an attack as seen in this study.
Cell bodies of PVN also control yawning, which is a common premonitory symptom of migraine patients (41–43), indicating that this nucleus may be relevant for the period prior to attacks. Yawning can also be induced in rats by activating oxytocinergic neurons from PVN to ventral tegmental area, hippocampus and amygdala, parallel to an observed extracellular dopamine increase in the shell of nucleus accumbens (43). Dopaminergic input to the accumbens shell may arise from the dorsal raphe nucleus (44). Schulte et al. reported enhanced functional connectivity between nucleus accumbens and dorsal pons including the dorsal raphe nucleus, as well as left amygdala and hippocampus in the preictal phase (8). Accordingly, central nervous networks with altered preictal function (8) may be related to hypothalamic systems that produce both premonitory symptoms and the sleep related alterations of cortical excitability seen in the present study.
Finally, the effect of SAI which was more reduced by sleep restriction closer to the start of attacks, is likely both mediated by cholinergic input from thalamus to the cortex and by α1 subunits of GABA-A receptors (21,45). Previous findings of preictally reduced rapid eye movement sleep (REM) density in migraine may indicate impaired cerebral cholinergic input during sleep (46). Sleep deprivation may also induce a reduction in cortical levels of acetylcholine in humans (47). Consequently, migraine patients may have a subtle cortical cholinergic deficit which is exaggerated by insufficient sleep demonstrated by a reduction of the SAI inhibitory effect closer to the migraine attack commencement.
Strengths and limitations
We utilised a study design which was randomised and blinded. The investigator was blinded to whether the participants were migraine patients or controls (not included in these analyses), where in the migraine cycle the subjects where and how much they had slept before examinations. This blinding was kept throughout data analyses.
We chose 24 hours as a cut off limit for excluding the preictal and postictal phase from each of the two separate analyses in this study. However, the exact duration of these phases is currently unknown and may vary between subjects. Some previous findings implicate a possible duration of 48 hours for the preictal phase (16). Thus, by this extended definition, some preictal properties may have been included in the analyses of effects after attacks.
Caffeine consumption was not allowed prior to the examinations in this study. This leaves a possibility for withdrawal effects as a confounding factor with unknown effects on cortical excitability.
Most participants reported in a questionnaire to have between one to three attacks per month and a large portion to have between four to five attacks per month for the past six months. Thus, the study population represents episodic migraine patients with an active disease. However, some subjects had a rather low attack frequency.
The phase-grouped secondary analyses only have a posteriori sample sizes sufficient to evaluate large effect sizes, particularly for the postictal recordings. Consequently, smaller effects of these variables may have remained undetected. The chance of detecting effects occurring at random is also greater with small sample sizes. We applied the Kenward-Roger approximation designed to improve the performance of complicated tests of fixed effects in small samples. However, the results of the secondary analyses should only be considered as preliminary, especially when not in accordance with the findings in the primary analyses. The significant findings in the primary analyses for SICI and LICI, but not SAI, had supporting findings in the secondary analyses. Because there were no significant findings for ICF in the primary analyses, the findings for ICF in the postictal phase in the secondary analyses should be considered preliminary.
We observed some numerical differences between the time to attacks and from attacks for the different sleep conditions. Whether these differences occur because sleep changes trigger attacks in some subjects is unknown. It is also uncertain whether subsequent sleep changes or sleep recovery following sleep restriction could affect attack initiation. However, there was no difference in number of preictal or ictal recordings following the two sleep conditions. Thus, we suggest that effects of the different sleep conditions on attack onset were absent or only applicable to a small subset of participants in this study.
Conclusion
We have revealed different effects of sleep restriction on cortical excitatory/inhibitory balance during the course of the migraine cycle. Our findings imply sleep-wake related mechanisms as part of the migraine cycle pathophysiology. Before and close to the start of an attack, we found increased GABA-B receptor mediated inhibition after sleep restriction. This effect may be related to alterations in mechanisms of circadian rhythm in the hypothalamus and may connect previous findings of altered cortical inhibitory capability and hypothalamic dysfunction. We also found reduced cholinergic mediated GABA-A inhibition after sleep restriction before and close to the start of an attack. This finding may indicate a cholinergic dysfunction prior to attacks which may be related to sleep architecture. Shortly after an attack, we observed a decrease of GABA-A mediated inhibition after sleep restriction. This effect may indicate that sufficient sleep during the migraine attack is of increased importance for maintaining normal neurological functioning in migraine patients.
Article highlights
Insufficient sleep gradually increases GABA-B receptor mediated inhibition and reduces cholinergic modulated GABA-A inhibition towards the start of a migraine attack. Insufficient sleep reduces GABA-A inhibition soon after a migraine attack, indicating that sufficient sleep may be of increased importance during migraine attacks to maintain normal neurological function.
Footnotes
Acknowledgements
We are grateful for the contributions from all subjects who participated in the study, and from our collaborators Lise Rystad Øie, Tore Wergeland Meisingset, Erling Tronvik and Marit Stjern. The examinations were performed at the core facility NeXt Move, Norwegian University of Science and Technology (NTNU).
Authors' contributions
All authors have provided substantial contributions to the final version of the manuscript. MSM, MU, TS and PMO planned the study. MSM conducted the examinations, performed the data analysis, interpreted the data and wrote the first draft. MU, TS and PMO contributed to the data analysis and writing of the manuscript. LJS, EB, MSS and GBG evaluated subjects for inclusion and provided feedback on the manuscript.
Declaration of conflicting interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Ethics approval and consent to participate
The work has been carried out in accordance with The Code of Ethics of the World Medical Association (Declaration of Helsinki). The study was approved by the Regional Committee for Medical Research Ethics Central Norway. Written, informed consent was obtained from all participants.
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was funded by The Liaison Committee for education, research and innovation in Central Norway. The project also received a grant from Norwegian Competence Center for Sleep Disorders, Haukeland University Hospital, Bergen, Norway. The funding sources had no involvement in the research.