
Abstract
Background
Migraine is two to three times more prevalent in women than in men, but the mechanisms involved in this gender disparity are still poorly understood. In this respect, calcitonin gene-related peptide (CGRP) plays a key role in migraine pathophysiology and, more recently, the functional interactions between ovarian steroid hormones, CGRP and the trigeminovascular system have been recognized and studied in more detail.
Aims
To provide an overview of CGRP studies that have addressed gender differences utilizing animal and human migraine preclinical research models to highlight how the female trigeminovascular system responds differently in the presence of varying ovarian steroid hormones.
Conclusions
Gender differences are evident in migraine. Several studies indicate that fluctuations of ovarian steroid hormone (mainly estrogen) levels modulate CGRP in the trigeminovascular system during different reproductive milestones. Such interactions need to be considered when conducting future animal and human experiments, since these differences may contribute to the development of gender-specific therapies.
Introduction
Migraine is a complex neurovascular disorder associated with dysfunction of the nociceptive trigeminovascular system (1). Migraine is two to three times more prevalent in women than in men (2). Additionally, women have more frequent and intense headaches, a higher risk of chronification, greater disability and an increased risk of cardiovascular events, including cardiovascular mortality (2,3). Although the precise mechanisms underlying this unique gender-dependent prevalence are not fully understood, they seem to be related to the hormonal fluctuations of ovarian steroid hormones, namely estrogens and progesterone.
Migraine prevalence before puberty is similar between sexes. A notably increased incidence arises in women after menarche, where hormonal fluctuations influence migraine occurrence during different reproductive milestones, such as menstruation, pregnancy, the postpartum state and breastfeeding, perimenopause and menopause (4–6). Furthermore, migraine frequency is affected by the use of oral contraceptive pills, hormone replacement therapy, selective estrogen receptor modulators (i.e. tamoxifen) and medical oophorectomy (7–9). Interestingly, rates of migraine similar to those in females were reported in male-to-female transsexuals under estrogen therapy to induce female sex characteristics (10). This relationship between fluctuations of ovarian steroid hormones and migraine incidence was suggested almost half a century ago by Somerville (11). He stated that in biologically-predisposed women, migraine attacks are triggered by a decline in plasma estrogen concentrations in the late luteal (premenstrual) phase of the menstrual cycle (4,11). On the contrary, phases of rising estrogen levels appear to protect against migraine (12), whereas progesterone withdrawal does not seem to precipitate migraine attacks (13). Additionally, migraine prevalence has been associated with menstrual disorders (e.g. menorrhagia, dysmenorrhea), as well as endometriosis and polycystic ovary syndrome (14–16). Thus, migraine incidence is influenced by the (dys)function of the hypothalamo-pituitary-ovarian system.
In the last years, the calcitonin gene-related peptide (CGRP) pathway has been discovered to play a key role in migraine pathophysiology. Revolutionary experiments revealed that during an acute migraine attack CGRP is released in significant amounts in the extracerebral circulation (17), that these elevated levels of CGRP are normalized after triptan treatment (18), and that infusions of CGRP in migraine patients provoke migraine attacks (19). In addition, all clinical trials with anti-CGRP drugs have been positive (20), consolidating the evidence for the role of CGRP in migraine.
Therefore, it is important to consider that fluctuations in ovarian steroid hormones levels may influence CGRP homeostasis. Indeed, the presence of sex hormone receptors in the trigeminovascular system suggests that trigeminal neurons are sensitive to variations in the levels of these hormones (21). However, the exact functional interactions between ovarian steroid hormones and CGRP, which could increase migraine prevalence in women, are not fully understood.
On this basis, the present review will consider the current knowledge on the effects of a changing hormonal environment on the modulation of the CGRP signaling pathway in the trigeminovascular system, and will address some sex-dependent considerations for future CGRP studies.
CGRP and migraine
CGRP, a neuropeptide with pleiotropic effects, is involved in different biological processes such as neuromodulation, cardiovascular regulation, inflammation, metabolic function, and aging (16). Two homolog isoforms have been identified, namely, α-CGRP and β-CGRP. α-CGRP is a 37-amino acid neuropeptide widely distributed throughout the nervous system and predominantly expressed in sensory Aδ- and C-fibers. It is produced by transcription and alternative RNA splicing of the primary transcript of the calcitonin gene (23–25). β-CGRP is encoded by a second calcitonin gene, differing by only a single amino acid in rats and by three residues in humans (25,26), and predominantly expressed in the enteric nervous system. Considering that the majority of studies focus on α-CGRP, this review will be limited to this isoform, with a focus on the trigeminovascular system because of its relevance in migraine.
Migraine pathophysiology has long been related with activation and sensitization of the trigeminovascular system, a functional pathway consisting of pseudounipolar neurons peripherally innervating the cranial vasculature and the dura mater, whose cell bodies are located in the ipsilateral trigeminal ganglion (27,28), and centrally projecting axons to the trigeminal nucleus caudalis and the upper two cervical divisions, known as the trigeminocervical complex (29). CGRP is highly expressed throughout the trigeminovascular system (30), it is synthetized in neuronal cell bodies and co-stored with tachykinins (especially substance P) in large granular secretory vesicles (31). During the headache phase of a migraine attack, CGRP is released in the perivascular space, producing neurogenic vasodilatation and nociceptive transmission (17,32).
Different experimental migraine models have focused on the various components of the trigeminovascular system. These models indirectly assess peripheral nociceptors in the dura mater and cranial arteries (i.e. neurogenic dilation of pain-sensitive dural blood vessels) or directly study nociceptive trigeminal fibers. The next sections will review animal and human experimental studies focusing on CGRP in the trigeminovascular system, where sex-related differences have been addressed.
Sex hormone effects in animal CGRP studies
Migraine is a female-predominant disorder. Surprisingly, the majority of preclinical studies have used male animals to limit experimental variations resulting from natural fluctuations or effects per se of ovarian steroid hormones (33). A recent study revealed that in naïve rat medulla and trigeminal ganglion, there were sex differences in the mRNA levels of CGRP and the three components of its receptor, namely, calcitonin-like receptor (CLR), receptor activity-modifying protein 1 (RAMP1), and receptor component protein (RCP, 34), with higher CGRP levels in the medulla and lower expression of RAMP1, CLR and RCP-encoding mRNA in female tissues compared to that of males. Even considering that no estrous cycle staging or hormone measurements were conducted, this suggests that fluctuating gonadal hormones may regulate CGRP receptor synthesis, expression, or release in the trigeminovascular system.
Expression of sex hormone receptors in the different components of the trigeminovascular system. Receptor expression for animals is only listed when available in the literature.

GPER: G-protein-coupled estrogen receptor 1; ND: not determined in humans.
Furthermore, androgen and progesterone receptors are expressed in the different components of the trigeminovascular system (see Table 1). Notwithstanding, until now, only one study has revealed that the circadian rhythm of testosterone did not affect CGRP accumulation in the trigeminal nucleus (38), while in the lumbar dorsal root ganglion, CGRP was downregulated by long-lasting testosterone treatment (39). As with the androgen receptors, few studies have focused on the modulation of CGRP by progesterone. In this context, the secretion of progesterone seems to decrease CGRP content in the trigeminal nucleus (38), while it increased CGRP in the dorsal root ganglion and in plasma (40). Since studies with these hormones are very limited, the role of androgen and progesterone receptors in modulating CGRP in the trigeminovascular system remains to be determined.
The effects of estrogen have been studied more widely. There are two classes of estrogen nuclear receptors, alpha (ERα) and beta (ERβ), and a G protein-coupled estrogen receptor (GPER). All estrogen receptors are expressed in different parts of the trigeminovascular system (see Table 1), such as the rodent trigeminal ganglion (41,42) and dura mater (43). Although it is believed that estrogens regulate excitability and sensitization of the trigeminal CGRP pathway, experimental data are scarce and inconsistent. In the female mouse trigeminal ganglion (41) and rat trigeminal nucleus (38), CGRP mRNA levels and CGRP content were constant throughout the phases of the natural estrous cycle, respectively. In contrast, in the rat trigeminal ganglion, ovariectomy significantly increased the gene expression of CGRP compared to cycling (control) rats, and these increased mRNA levels were lowered following estrogen replacement (42). Similarly, in the central nervous system, a deficiency of estrogens increases CGRP levels within the midbrain periaqueductal gray (44), medial preoptic nucleus of the hypothalamus (45) and lumbar dorsal root ganglion (46), while CGRP expression was downregulated after estrogen treatment in the dorsal root ganglion (46) and trigeminal nucleus caudalis (47). However, it has also been described that estrogen positively enhances the expression of CGRP within the dorsal root ganglion (48–50), rat anterior pituitary (51) and medial preoptic nucleus (52). Thus, experimental data suggest that estrogen modulates differently, and in a complex manner, the density of CGRP receptors in the peripheral and central nervous systems. Further studies are clearly required to determine the exact role of estrogens, and which estrogen receptors are involved, in the modulation of CGRP expression in the different components of the trigeminovascular system (Figure 1). The effects of estradiol on the CGRP-ergic system have also been studied using an epigenetic approach, but the variation in the methylation of the CGRP-ergic genes in that study was too large to allow definite conclusions on the role of estradiol (53).
Trigeminovascular system. Sites where estrogens, mainly estradiol (E2), modify CGRP receptor expression/function in rodents (purple) and humans (blue).
Therefore, an alternative approach to assess the functional estrogen-CGRP relationship in the whole trigeminovascular system is with specific in vivo animal migraine models. In this context, Gupta et al. (54) have investigated the modulation exerted by 17β estradiol and progesterone (separately and in combination) on dural vasodilation in ovariectomized rats. Periarterial electrical stimulation of the dural vasculature (i.e. the middle meningeal artery) was applied to release CGRP from the activated trigeminal nerve. The subsequent changes in vessel caliber were measured with intravital microscopy on a closed cranial window. Dural vasodilation was induced by endogenous (released by periarterial electrical stimulation or intravenous capsaicin) or exogenous CGRP. The vasodilatory responses to exogenous CGRP or capsaicin were not affected by ovariectomy or by hormonal replacement. However, estradiol pretreatment significantly enhanced neurogenic sensory vasodilation, suggesting that estrogen modulates prejunctionally the sensory release of CGRP in trigeminal perivascular nerve endings in vivo (54).
Another study performed by Martin et al. (55) explored with an in vivo experimental migraine headache model whether sensitization of the trigeminal sensory system changes during different stages of the rat estrous cycle. In anesthetized rats, an electrode was used to record trigeminal nucleus caudalis neurons activity after cutaneous stimuli and chemical stimulation (with topical capsaicin) of the dura mater. There was an enhanced sensitization of the trigeminal system during the later halves of proestrus and estrus, coinciding with an abrupt decline in ovarian steroid hormones (Figure 2). This finding suggests that sensitization of the trigeminal system is dependent on the varying hormonal milieus throughout the estrous cycle (55).
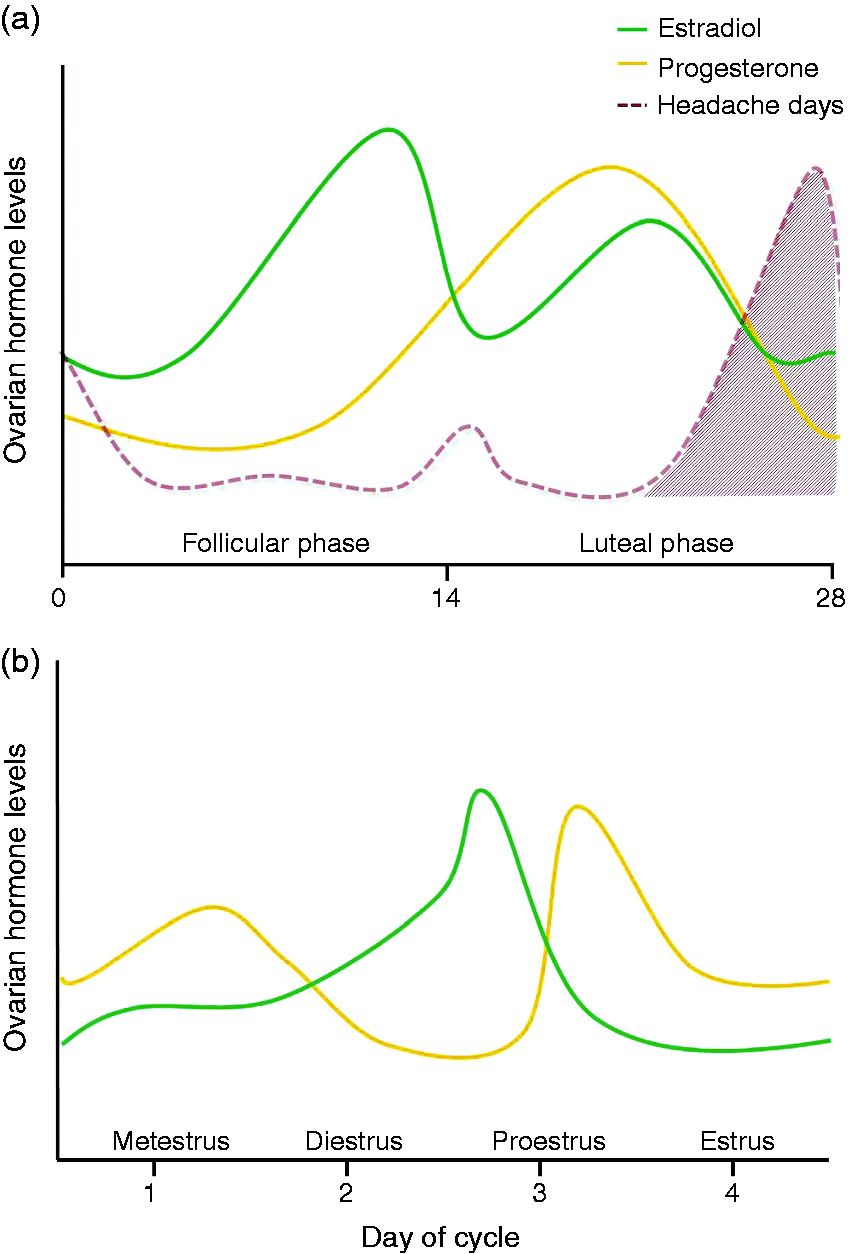
In summary, animal studies have provided evidence that activation of the CGRP system is dependent upon the different stages of the rat estrous cycle, with specific roles for the different sex hormone receptors. However, there are hormonal differences between the rodent estrous cycle and the human menstrual cycle and, therefore, a direct extrapolation to humans should be done cautiously (55, Figure 2). Evidently, further studies will be required to delineate the exact mechanism(s) involved in this (patho)physiological estrogen-CGRP relationship.
Gender effects in human CGRP studies
One of the first clinical studies that discovered a relationship between female sex hormones and CGRP was published in 1986 by Stevenson et al. (56). In this study, concentrations of immunoreactive plasma CGRP (i-CGRP) were measured in healthy controls, throughout normal pregnancy and in the subacute postpartum period. i-CGRP concentrations were significantly increased throughout pregnancy, with the highest concentrations being found towards term. These increased in CGRP levels decreased during the first days after delivery, when they were similar to the controls (56), interestingly, while migraine in many cases seems to be related to increased CGRP levels, during pregnancy and around the delivery, an inverse relationship between CGRP levels and migraine attack incidence seems to be the case. Most likely, this may be attributed to differences between local cranial CGRP levels that are relevant in migraine, and systemic CGRP levels that are increased as part of the cardiovascular adaptation process during pregnancy. Later on, a key study from Valdemarsson et al. revealed that plasma i-CGRP levels are different between both genders. It is remarkable that, in healthy subjects, i-CGRP levels were significantly higher in females than in males, and that the use of combined contraceptive pills was associated with even higher levels of i-CGRP in plasma (57). Accordingly, in postmenopausal women, decreased estradiol serum levels were positively correlated with decreased plasma i-CGRP concentrations (58), ascribing that the CGRP system could be influenced directly by endogenous or exogenous ovarian steroid hormones.
Is there a gender difference in the functionality of CGRP and the trigeminovascular system in humans?
In the last years, some non-invasive human experimental migraine models have relied on laser Doppler perfusion imaging to study cutaneous antidromic vasodilation induced by chemical stimulation of primary sensory neurons. In this way, topical application of capsaicin on the forearm depolarizes nociceptive neurons via activation of prejunctional transient receptor potential ion channels of the vanilloid type 1 (TRPV1), leading to release of vasoactive peptides, neurogenic inflammation, and subsequent increase of dermal blood flow (59). In this pharmacodynamic model, the capsaicin-induced vasodilation is mediated by release of CGRP, as this response was inhibited by CGRP receptor antagonists (i.e. CGRP8–37 and telcagepant) and by a humanized monoclonal antibody directed against CGRP (60–62).
Using this experimental model, a recent study explored gender differences on sensory CGRP release in healthy subjects and migraine patients, as well as the influence of varying levels of estrogen and progesterone during the menstrual cycle (63). In healthy males, dermal blood flow responses did not vary over time and were comparable with the responses of male migraineurs. However, in healthy women, fluctuations of ovarian steroid hormones influenced CGRP-dependent dermal blood flow, with the highest reactivity observed around the menstruation period, when estrogen levels are low. This is in accordance with the estrogen withdrawal hypothesis, where perimenstrual low levels of estrogen are associated with a higher prevalence of migraine (4,11, Figure 2), assuming that an increased reactivity relates to an increased vulnerability to a migraine attack. Interestingly, in female migraine patients, dermal blood flow responses were elevated compared to healthy subjects, but these responses were independent of the menstrual cycle. In this study, serum estradiol and progesterone between healthy women and migraine patients did not differ across the hormonal cycle. Therefore, the mechanisms involved in the increased dermal blood flow in female migraine patients after capsaicin could be the result of an increased TRPV1 expression or an increased release of CGRP (63).
An optimization of the previous model was done by studying a dermatome in the human forehead innervated by the frontal branch of the ophthalmic (V1) nerve, where the increase in dermal blood flow is produced by trigeminal nerve activation via topical capsaicin application or electrical stimulation of the forehead skin (64).
Using this new specific model, a recent study explored the effects of the menstrual cycle on trigeminal nerve-induced vasodilatation in healthy reproductive women with a regular menstrual cycle, postmenopausal women, and patients with menstrually related migraine (MRM). Cycle-dependent changes in trigeminovascular reactivity were observed in healthy women with enhanced responses around their menstruation (65). Notably, these findings are similar to those found with pain perception, sensory and vasomotor responses, where trigeminal sensitization was produced after an intradermal injection of capsaicin in the forehead (66). In contrast, the physiological absence of ovarian steroid hormones in postmenopausal women did not induce any trigeminal variation during all measurements. Surprisingly, in MRM patients, low estradiol levels were detected during the luteal phase (days 19–21 of the cycle), while cyclic dermal blood flow differences during the menstrual cycle were also absent. This could be attributed to a differential activity of the TRPV1 channel produced by estrogens (65), as they alter the expression of the TRPV1 receptor in sensory neurons (67); however, the precise mechanisms involved were not explored.
Another non-invasive technique commonly used to study the trigeminal nociceptive system is functional neuroimaging. Within this context, Maleki et al. (68) used high-field magnetic resonance imaging (MRI) in male and female migraineurs to determine interictal alterations in brain structures using a thermal pain stressor. Female migraineurs had stronger activation of the spinal trigeminal nucleus compared with male migraineurs and healthy controls (68). These findings were attributed to differences in sensitivity of the trigeminal system due to hormonal processes, but female participants were studied without assessing the stage of the menstrual cycle (68).
A further remarkable study recently reported a female migraine patient who had a high-field MRI every day for 30 days, while standardized trigeminal nociceptive stimulation was done during the experiment. It was found that the hypothalamus, depending on the state of the migraine cycle, exhibited an altered functional coupling with the spinal trigeminal nucleus, with the greatest functional coupling in the last 24 h preceding the onset of the migraine attack (69). Regrettably, the relationship between the stage of the menstrual cycle and the activation of the trigeminal nucleus was not reported in this patient. Using MRI seems a promising tool for detecting cyclic activation of the trigeminovascular system during the different stages of the menstrual cycle, and to the best of our knowledge, these studies have not yet been performed.
Overall, sex-related differences in the trigeminal nociceptive system are evident. Notwithstanding, further experimental research will be required to demonstrate whether the fluctuations of ovarian hormones levels produce: (i) a direct effect on trigeminally innervated blood vessels; (ii) a modulation of prejunctional channels that release CGRP in the dura mater; or (iii) a modulation of trigeminal central pain pathways (Figure 1).
CGRP receptor antagonism in females
Because of the unquestionable role of CGRP in migraine pathophysiology, small molecule CGRP receptor antagonists such as olcegepant and telcagepant (gepants) were developed and proved to be effective in the acute treatment of migraine headaches (70,71). The safety and efficacy of telcagepant was evaluated for short-term prevention of perimenstrual headaches. When taken perimenstrually for seven days, telcagepant reduced perimenstrual headaches, but it was associated with transaminase elevations in a small number of patients (72). Unfortunately, clinical trials for chronic migraine treatment were terminated because of pharmacokinetic limitations (73) and safety concerns related to hepatotoxicity (74). Notwithstanding the fact that these drugs have been discontinued due to side-effects, CGRP receptor antagonism has been proven effective in migraine treatment and, indeed, other small molecule CGRP receptor antagonists are currently under development. Moreover, at present, there are four monoclonal antibodies (mAb) in clinical development for migraine prophylaxis (75–78), namely, three humanized mAb targeting CGRP (galcanezumab, eptinezumab and fremanezumab) and one fully human mAb targeting the CGRP receptor (erenumab). In migraine patients of both genders, these drugs have shown efficacy, tolerability, and few adverse effects (compared to the gepants) in phase 2 randomized control trials (75–78). However, the exact site and mechanism of action of these novel drugs or the long-term side effects have not been completely elucidated. Until now, there has only been one randomized double-blind, placebo-controlled study, where prolonged CGRP inhibition on cardiovascular parameters was studied in 31 healthy perimenopausal women. Fortunately, no associated hemodynamic changes were found after chronically inhibiting the CGRP pathway with fremanezumab (79), despite the fact that CGRP (receptor) blockade might theoretically have a larger impact on cardiovascular safety in females than in males (80). Thus, additional randomized control trials with the other mAb should confirm this in women in the reproductive age range.
In summary, CGRP (receptor) blockade is an effective therapeutic option for treating migraine. Clearly, further clinical trials will be required to elucidate whether these novel drugs are safe in individuals with cardiovascular risk factors, if there are any consequences of chronic CGRP inhibition in young reproductive women with a normal menstrual cycle, and whether efficacy depends on the phase of the menstrual cycle.
Future considerations and conclusions
Gender differences are evident in migraine. Animal research is starting to shift from including only male animals to also considering female animals in studies, but there is still much to be done. Consequently, gender-related differences in the CGRP system are emerging. Numerous animal and human studies have shown that cyclic fluctuations of ovarian hormones (mainly estrogen) modulate CGRP in the peripheral and central trigeminovascular system; this is especially relevant now that novel antibodies directed against CGRP or its receptor are currently in clinical trials. Future studies should focus on how fluctuations of gonadal hormones influence migraine pathophysiology in both genders, especially the estrogen-CGRP relationship that seems a key factor in the higher prevalence of migraine in women. Hopefully, these sex-related differences may contribute to the development of gender-specific therapies.
Article highlights
Sex hormone receptors are expressed in the different components of the trigeminovascular system. Fluctuations of estrogen levels modulate CGRP receptor signaling in the trigeminovascular system. The estrogen-CGRP relationship seems to be a key factor involved in the higher prevalence of migraine in women.
Footnotes
Acknowledgement
Figure 2 was modified from Servier Medical Art, licensed under a Creative Common Attribution 3.0 Generic License. ![]() .
.
Declaration of conflicting interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: Antoinette MaassenVanDenBrink was supported by the Netherlands Organization for Scientific Research (Vidi grant 917.113.349), whereas Carlos M. Villalón, Eloísa Rubio-Beltrán and Alejandro Labastida-Ramírez were supported by Consejo Nacional de Ciencia y Tecnología (CONACyT; Grant No. 219707 to CMV and fellowships No. 409865 to ERB and 410778 to ALR; Mexico City).